
Method Article
Время дифференциальной окраски Техника сочетании с полноценного двустороннего Gill денервации для изучения Ionocytes в Рыбок
В этой статье
Резюме
This manuscript describes a protocol to track the re-distribution of branchial ionocytes and their innervation using a time differential staining technique coupled with full bilateral gill denervation.
Аннотация
Жаберные ionocytes (ИС) являются функциональными единицами для ионной регуляции у рыб. У взрослых, они находятся на filamental и пластинчатой эпителия жаберных где они транспорт ионов, таких как Na +, Cl - и Са 2+ с помощью различных ионных каналов, насосов и теплообменников. Костистых Гилл внешне иннервируются лица (VI), языкоглоточного (IX) и блуждающего (X) нервы. В IX и X нервов также внешний источник жаберного IC иннервации. Здесь два методы, используемые для изучения иннервации, пролиферацию и распределение микросхем описаны: время методика дифференциальной окраски и полноценного двустороннего техника жаберных денервация. Короче говоря, золотые рыбки подвергаются жизненно митохондрий конкретных красителя (например, Mitotracker красный), которые этикетки (красной флуоресценции) уже существующих ИС. Рыбы были либо оставляли для восстановления в течение 3 - 5 дней или сразу прошел полную двустороннюю жаберные денервации. После 3 - 5 дней восстановления, гбеды собирают и фиксируют для иммуногистохимии. Затем ткань окрашивали с α-5 первичным антителом (цели Na + / K + АТФ-азы, содержащая элементы) в сочетании с вторичным антителом, что этикетки всех (новые и уже существующие) СКВ зеленым. Использование конфокальной микроскопии, было показано, что уже существующие ИС появляются желтые (маркированы как жизнеспособный митохондрий-специфической краски и α-5) и новых микросхем появляется зеленый (помечен α-5 только). Оба метода, используемые в тандеме могут быть применены для изучения иннервации, пролиферацию и распределение ИС на жаберной нити, когда рыба подвергаются экологических проблем.
Введение
ИС являются функциональный блок для ионной регуляции у рыб и находятся на эпителиальные поверхности жаберных лепестках и ламели 4,6-8,10. Хотя различные подтипы были описаны, что обладают уникальными особенности, многие из микросхем характеризуются высокой плотностью митохондрий (следовательно, они также известны как митохондрий богатых клетками) и / или избытка фермента Na + / K + АТФазы (НКА). Как правило, это ИС дом множество других насосов, ионных каналов и теплообменников, участвующих в ионной регуляции (например, Na + / H + обменника, Na + / Cl - совместного переносчика, H + насоса) 2,10,11. Перераспределение и распространение ИС в качестве компенсаторного механизма является центральным для поддержания ионного гомеостаза особенно во время ионной стресса (например, воздействие ионов бедных водой) 4,8,9.
Это исследование описывает разницу во времени окрашивания техниQue 1, чтобы определить недавно распространились ionocytes (ICS) в жабры рыб. Эта методика в сочетании с полной двусторонней денервации жаберных дуг. Золотая рыбка (карась рыбки), виды, используемые в этих исследованиях, хорошо подходят для изучения распространения жаберных эпителиальных клеток, потому что они имеют замечательную способность конструктивно переделывать свои жабры 2. Гилл ремоделирование относится к росту или втягивания в межпластинчатом клеточной массы (ILCM), когда рыба (обычно поддерживают на уровне 15 - 30 ° С) приспособиться к холодной воде (<15 ° С) или гипоксии, соответственно 3. Предыдущие исследования с использованием временной технику дифференциального окрашивания на золотых рыбок были сосредоточены на перераспределении иннервации и распространения ИС на жаберных в контексте жаберных ремоделирования 4,5. Като и Канеко 1 разработали эту новую технику для изучения трансформации и замены жаберных ИС в Killifish (Fundulus heteroclitus) transferred из морской воды (SW) в пресной воде (FW). В этом исследовании, акцент делается на распространение и иннервации ИС в золотой рыбки приспособиться к 25 ° C.
Использование времени технику дифференциального окрашивания было показано, что, в контексте жаберной ремоделирования, золотых рыбок поддерживать постоянное количество микросхем при гипоксии и последующей нормоксических восстановления, однако, процент клеток, иннервируемых уменьшилось по всей нормоксических восстановительного периода 5. Было предложено более 70 лет назад, что ионные механизмы вещества в организм рыб находятся под нейронной контроля 12. Костистых жаберных иннервируется лицевым (VII), языкоглоточного (IX) и блуждающего (X) нервы также именуемые "жаберных нервов" 13,14. Исследования Jonz и медсестра (2003) рыбок данио (Danio rerio) жаберных иннервации показал, что происхождение иннервации внешняя (тело клетки нервного волокна внешними по отношению к жаберных), а также внутренние (корпус ячейкинервных волокон является неотъемлемым жаберных). Те же авторы показали, что жаберные микросхемы внешне иннервируются 7.
В этом исследовании, время методика дифференциальной окраски в сочетании с полной двусторонней жаберных денервации был использован для исследования распространения ИС, не имеющих внешнюю иннервации в золотых рыбок. Полноценного двустороннего жаберных денервация относится к разрывая черепных нервов IX и X. Эти два подхода могут быть реализованы в золотую рыбку, потому что их относительно большой размер (от 30 - 200 г) упрощает тонких хирургических процедур и ionocytes легко идентифицировать с помощью стандартных методов иммуно-гистохимические. В настоящем исследовании, ИС визуализировали с использованием жизненно митохондрии-специфической краски (например, красный) Mitotracker или первичное антитело против α-субъединицы Na + / K + -АТФазы (α-5; с развитием исследований гибридом банка, Университет Айова, Айова-Сити IA). Этот протокол обеспечивает простой метоd визуализации и анализа перераспределения и распространения ИС на рыбу жаберных.
протокол
Оба протокола соответствовала руководящим принципам Канадского совета Уходу за животными (ССАС) и были проведены с одобрения университета Оттавы комитета по уходу за животными (Протокол BL-226).
1. Время дифференциальной окраски Техника: Митохондрия богатых Краска Ванна
- Подготовка 1 мМ Mitotracker Red маточного раствора путем растворения 50 мкг в 94,0 мкл диметилсульфоксида (100% ДМСО). Держите маточного раствора в темноте при -20 ° С, когда он не используется. Избегайте циклов замораживания / оттаивания.
- Подготовка темные коробки (3 - 6 ящиков) с максимальным объемом 600 мл. Заполните ящики с 400 мл воды в системе (вода рыб обычно проводятся в) и поместите воздушный камень в каждой коробке, чтобы обеспечить источник O 2. Получить золотую рыбку (30 - 40 г) и поместите их в коробки с 400 мл воды и воздуха камня. Через 30 мин добавить жизнеспособной митохондрии богатые краситель с получением конечных концентраций 0,1 мкМ и 0,01% ДМСО. Купайтесь рыбу в течение 4 чр.
- Если это контроль рыба (т.е., не денервация), включите расхода воды в боксы, позволяют краситель, чтобы избавиться от и восстановить рыбу на период времени, отведенный в протоколе. Рыба, как правило, восстанавливается в течение 3 - 5 дней.
- После периода восстановления перейдите к разделу 3: Время дифференциальной окраски Техника: иммуногистохимии. Если эти рыбы должны быть денервированные перейдите к разделу 2: Полноценного двустороннего Порядок Денервационная.
2. полноценного двустороннего Порядок Денервационная
- Получить 1 пару студенческих Vannas весенних ножницами (изогнутые), 1 пара стандартных шаблонов щипцов и гетеросексуалов, 1 пара стандартных шаблонов щипцов с загнутыми, 2 пары № 5 щипцы, 1 пара тканей преднатяжителями, небольшой ватные шарики (1 - 2 мм в диаметре), и ватные тампоны (Q Советы или эквивалент).
- Подготовка анестезии водяную баню. Во-первых, растворяют 10 г бензокаин до конечного объема 100 млиз 99% этанола с получением исходного раствора. Для приготовления анестезирующего ванну воды, растворяют 15 мл на складе бензокаин раствора в 30 л газированной воды системы при требуемой температуре. Аэрации воды, поместив воздушный камень, соединенный с воздушным насосом или центральной воздушной линии в резервуар с водой.
- Поместите рыбу в анестезиологической водяной бане (рис 1А).
- После того, как дыхание прекратилось, положите рыбу на хирургический стол и интубация его, как показано на рисунке 1В. Сделайте это, вставив трубку в его ротовой полости для орошения жабры с газированной анестезией. Орошение жабры гарантирует, что рыба подается с достаточным количеством О 2 и анестетика во время процедуры денервации. Расположите рыбу так, чтобы голова слегка наклонена вниз. Это позволяет для лучшего доступа к области за четвертый арки жаберных.
- Аккуратно поднимите крышечку с прямой стандартная схема щипцови поместите тканей преднатяжителями между крышечкой и внутренней части головы. Осторожно откройте тканей преднатяжителями, чтобы сохранить крышечку от головы и держать жабры открытые.
- Убедитесь, что анестезирующий раствор в полива жабры во время этой процедуры. Отдых втягивающее ручки рядом с головой, которая обеспечивает доступ ко всем четырем жаберных дуг.
- Поместите изогнутые стандартная схема щипцы между четвертой арки жаберных и затылок и осторожно открыть их, чтобы создать напряжение в связки, крепящие четвертый жаберные дуги в голову. С парой № 5 щипцы создается небольшое отверстие (2 - 3 мм) путем прокалывания эпителий, соединяющий спинной части жаберных дуг в оперкулярной полости. Будьте осторожны, чтобы не идти слишком глубоко, потому что есть риск повреждения основной кровеносный сосуд.
- С небольшим ватным тампоном, состоявшейся с № 5 щипцы медленно и осторожно расширить разрез, чтобы выставить на IX (языкоглоточного) и Х (блуждающих) нервов. Бесплатныйнервы от любой соединительной ткани с помощью № 5 щипцы, опять стараясь не повредить кровеносные сосуды.
ПРИМЕЧАНИЕ: жаберная IX и X нервов золотая рыбка отдохнуть в глубоком тылу четвертого арки жабры и в непосредственной близости от крупных кровеносных сосудов, питающих в жаберных дуг. - После того, как нервы были определены Используйте изогнутые пружинные ножницы, чтобы тщательно вырезать нервы, удерживая разрез открытый с изогнутыми стандартных щипцов формы. После разрыва нервы, аккуратно сворачивая изогнутые щипцы. Там нет необходимости, чтобы закрыть разрез со швами, поскольку эпителий очень тонок и разрез обычно закрывает по себе в течение 24 - 48 ч. Удалить тканей втягивающим.
- Повторите ту же процедуру с другой стороны головы.
- Включите орошения жабр от анестезии на свежий газированной воды, чтобы восстановить рыбу из анестезии. После того, как Крышечные движения возобновили перемещения рыбы в экспериментальных танков, чтобы восстановиться, по крайней мере, 24 ч. Выполнить процедуру "липовый" на отдельном наборе рыбы. Процедура "обман" включает в себя пирсинг эпителий за четвертый арки жаберных без отсечения нервы.
3. Время дифференциальной окраски Техника: Иммуногистохимия
- Во-первых, получение 4% параформальдегида (PFA) в 1X фосфатным буферным раствором (PBS, 4 г PFA в 96 мл PBS). PFA не легко растворяются в PBS при комнатной температуре. Тепло раствор на водяной бане до растворения PFA. Выполните это в вытяжном шкафу. После того, как PFA в растворе дайте остыть перед использованием. Хранить при 4 ° С в течение 2 недель.
- Перед эвтаназии рыбу и извлечения жаберные ткани, 3 место - 4 мл 4% PFA в сцинтилляционный флакон на общую сумму 8 сцинтилляционные флаконы (1 флакон на жаберной дуги). Кроме того, принять небольшой вес лодки и заполнить его с 1x PBS. Эта информация будет использоваться для мытья ткани после того, как вырезали. Держите все решения на льду.
- После выдержки красителя жизнеспособного митохондрий богатыхЭксперимент закончен, усыпить рыбку, поместив его на водяной бане при передозировке бензокаин.
- Используйте тупые щипцы, чтобы поднять крышечку на одной стороне головы и изогнутые ножницы, чтобы отрезать каждый конец корзине жаберных жаберных. Тщательно подобрать жабры от тычинки с использованием тупых щипцы и поднимите их из оперкулярной полости. Немедленно промыть жабры в ледяной 1x PBS, чтобы удалить избыток бензокаин и кровь.
- Поместите вырезали жабры в отдельных ампулах (флакон для каждого жаберной дуги), заполненной 4% PFA и исправить O / N при 4 ° С.
- После фиксации стирать избыток PFA в 1X PBS, и поместить ткани в 2 мл пули трубки, заполненной 1,5 мл 1% Triton-X на шейкере в течение 6 ч при комнатной температуре или O / N при 4 ° С. Этот шаг permeabilizes ткани. Если вся жаберных слишком большой, чтобы поместить в 2 мл пробирку затем разрезать ткань на части, достаточно маленькие, чтобы соответствовать трубки, стараясь не повредить нити.
- Подготовка первичных антител разведения Мисин 4 мкл исходного раствора каждого первичного антитела (всего 8 мкл) в 992 мкл 1х PBS. Убедитесь, что НКА (этикетки НКА богатых клетки) и Zn-12 (этикетки нейроны) первичные антитела были подняты в том же хосте. Это важно, если исследователи решили определить конкретные подтипы IC в то же время, для которых они должны будут использовать первичные и вторичные антитела, которые выросли в разных видов хозяев.
- Извлеките Тритон-X решение и без промывки ткани добавляют 1 с: 250 разбавление (разбавленной в 1x PBS) моноклонального антитела NKA (α-5) для обнаружения NKA-богатые клеток и нейронов данио конкретного антитела (Zn-12) для обнаружения нервных волокон (первичные антитела) и инкубируют в шейкере в течение 6 ч при комнатной температуре или O / N при 4 ° С.
- Промыть первичного антитела 3 раза по 3 мин каждый из которых использует 1X PBS. Чтобы сделать это, удалите первичный раствор антител от трубки всасывания его с помощью пипетки.
- Подготовка вторичного антитела в1: 200 разбавление путем смешивания 5 мкл вторичного антитела фондовой в 995 мкл 1х PBS. Применение вторичного антитела (Alexa Fluor 488) и инкубируют в течение 6 ч при комнатной температуре или O / N при 4 ° С на качалке.
ПРИМЕЧАНИЕ: Mitotracker Красный возбуждается на ~ 594 нм длины волны, и высветится красным цветом. Вторичное антитело, которое возбуждается на ~ 488 нм длины волны, и флуоресцирует зеленый должны быть использованы для обозначения NKA и Zn-12 первичных антител. - Удалить избыток вторичного антитела путем промывки ткани 3 раза в течение 5 мин каждый (как описано в стадии 3,7).
4. изображений
- После промывки, установите ткани на вогнутой слайд для всей установки конфокальной микроскопии клеток и нервных волокон. Для установки ткани, в первую поместите его в папку (200 мкл) 1x PBS на плоской предметное стекло. Это гарантирует, что ткань не пересушивает.
- Отдельные жаберные hemibranchs с изогнутыми микро-ножниц. Поместите каплю 1x PBS и снижение монтажных СМИ в вогнутой слайд,
- Поместите разделенных hemibranchs в вогнутую слайд с передней кромки нити вверх и накрыть покровным стеклом. Приложите края покровного стекла с лаком для ногтей в целях предотвращения покровное передвигаться и перемещения ткани. Разрешить ткани оседают на дно вогнутой слайд в течение 10 - 15 мин до визуализации.
- Для каждого жаберной дуги, выбрать один из шести жаберных нитей наугад для работы с изображениями, создавая шесть изображений в жаберной дуги. Используйте обычный конфокальной микроскопии для изображения тканей, взяв 1 - 3 мкм оптических срезов.
Примечание: Любые уже существующие ячейки будут помечены Mitotracker красный и позитивным для НКА появится только желтого цвета. Клетки, которые появляются красные только уже существовали ИС, которые не содержат НКА. Любой вновь распространились клеток будет только положительным для НКА и появится только зеленый. Нервные волокна также появится зеленый.
5. Анализ изображений для Ionocyte количественной
- Для ВАСч жаберных нитей, которые были отображены, количественно ИС и связанных с ними иннервации путем прокрутки разделов Z стека и подсчета количества пластинчатых и filamental ИС настоящее и действительно ли они являются вновь дифференцируются, уже существующие и / или Иннервируются ,
- Количественную оценку ИС на нить или на область (мм 2) нити. Сделайте это с помощью инструментов рисования, связанные с программным обеспечением, используемым на приобретение изображения очертить пластинки нити охватывающей область нити, в которых количественно ИС. Большинство компьютерных программ конфокальной микроскопии есть возможность рассчитать площадь контурным области на изображении. Используйте эту опцию в ПО для обработки изображений, которая позволяет это сделать, чтобы получить этот район.
- Разделить подсчитанные ИС в области, рассчитанной с помощью программного обеспечения, чтобы получить меру ИС на единицу площади (например, на 2 мм).
Результаты
Рисунок 1 иллюстрирует заносим в таблицу хирургии (рис 1а) размещение рыбы во время операции (рис 1б), и три самых важных шагов для временного техники дифференциального окрашивания (рис 1в). На шаге 1, рыба выдерживают в течение 30 мин в хорошо аэрированной водяной бане при 25 ° С в темноте. В течение периода 30 мин, исследователь может подготовить аликвоту краситель митохондрии богатые в ДМСО, который добавляют в воду при шаге 2 (фиг 1С). Инкубационный период в шаге 2 позволяет для поглощения митохондрии, богатые краски из воды в митохондрии богатые клеток (например, ИС). Рыба может либо пройти полную двустороннюю процедуру денервации или процедуры фиктивный, в котором рыба под наркозом и opercula манипулировать, но нервы остаются неповрежденными. Создан в шаге 3 представляет собой камеру восстановления событий, предусмотренной проточной водой для рыбы, которая имеет EITher претерпел полную двустороннюю денервации или процедуры "липовый".
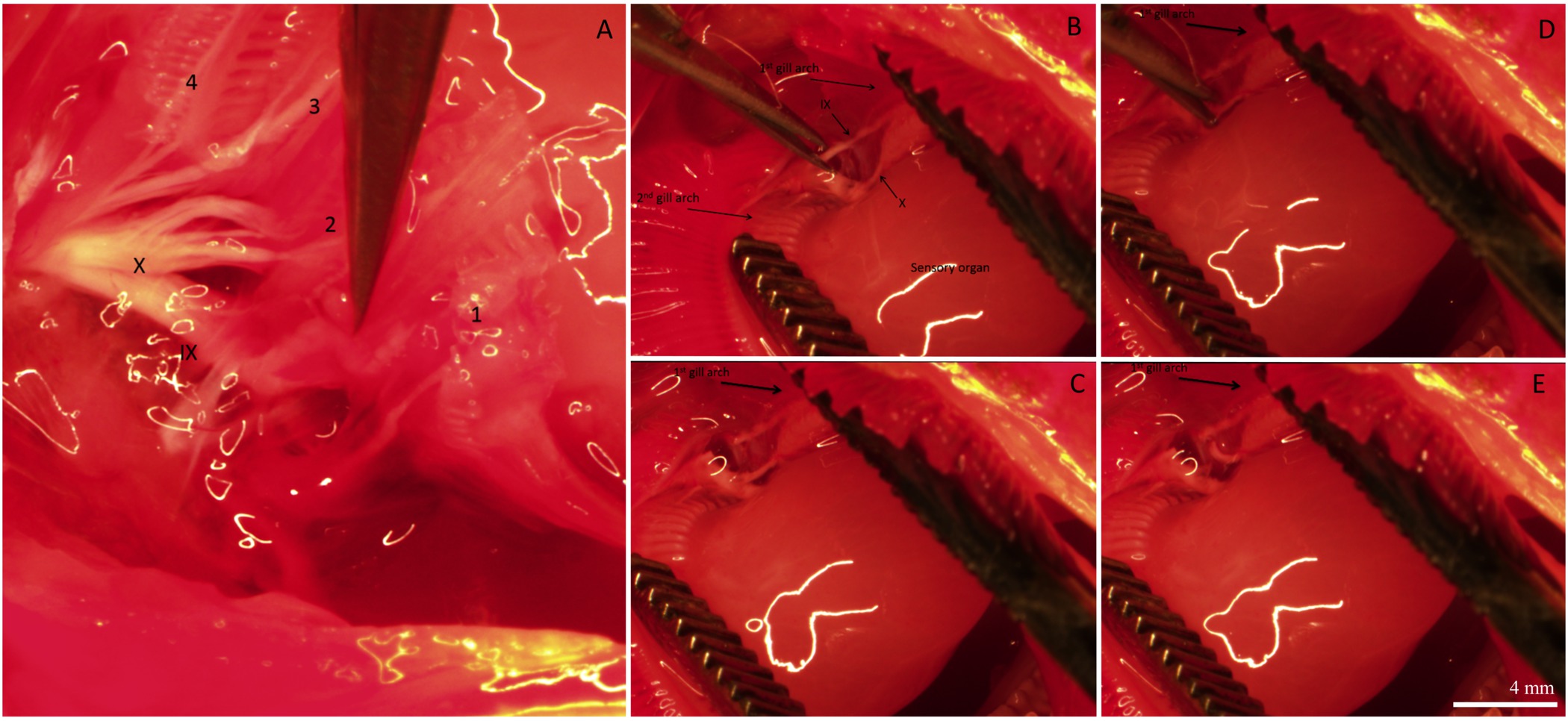
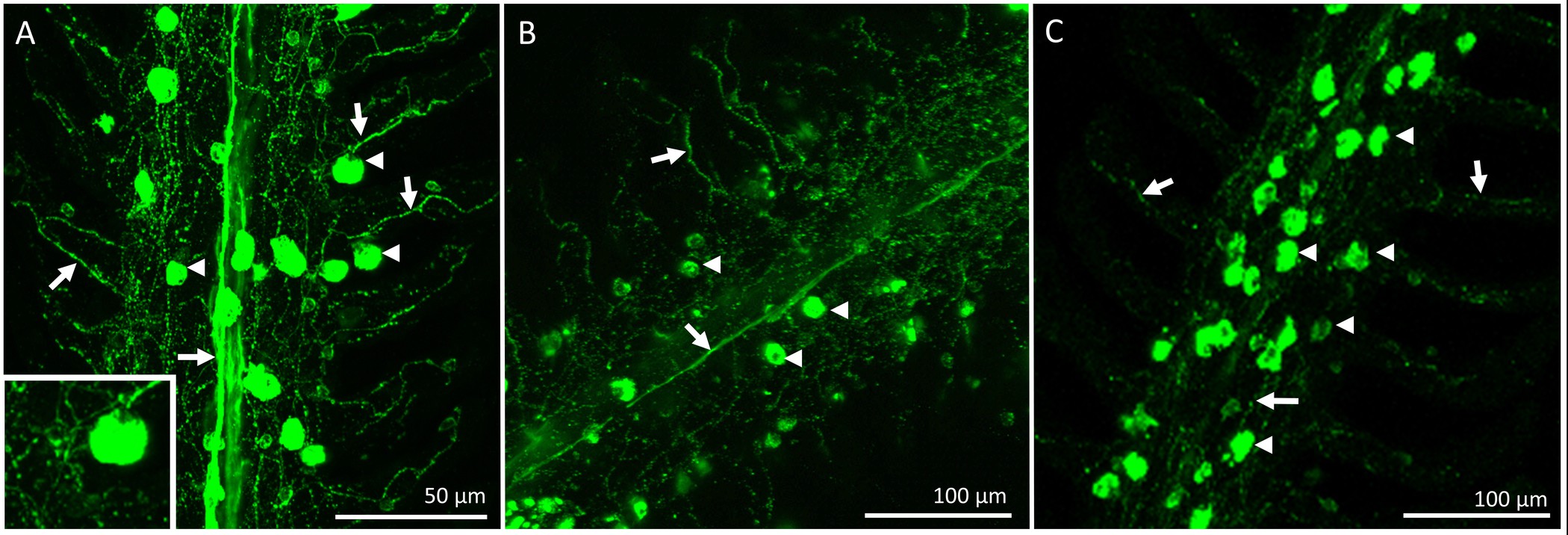
После восстановительного периода, рыба было умерщвлено и жабры вырезали и фиксировали для иммуногистохимии. Общее распределение и иннервации ИС на жаберной нити рыбы, которые подверглись процедуре "фиктивный" изображен на фиг.2. Микросхемы могут присутствовать на filamental эпителия, а также на базе межпластинчатом регионов. На рисунке 2а уже существующие ИС с маркировкой митохондрий богатых красителя (т.е., эти микросхемы существовал до проводились процедуры денервация / бутафорские). Рисунок 2B показывает нервные волокна, иннервирующие ранее существовавшие и вновь образованных ИС (определены НКА иммунореактивности) в filamental и пластинчатые эпителий. Наконец, слияние двух изображений (рис 2С) ясно показывает, ранее существовавшие ICS (отображается желтым цветом) и новые ICS (горит зеленым светом).Рис 2D является представителем график ИК количественного для нити, показанной на фиг.2А-С. На этой конкретной нити, как представляется, быть больше, количество вновь распространились на ИС мм 2, чем уже существующих микросхем (N = 1). Чтобы удалить источник внешней жаберной иннервации, черепные нервы IX и X (внешняя иннервации) были разорваны. 3А показывает спинной области оперкулярной полости после жаберных лепестках и эпителий, охватывающих нервы были удалены, чтобы выставить IX и X черепно-мозговые нервы (указанные с римскими цифрами), которые охватывают с двух основных нервных стволов иннервирующих все четыре арки (рис 3а); жаберные дуги пронумерованы от 1 до 4. Рисунок 3B-E иллюстрируют избирательное денервации первой жаберной дуги. Первый Гилл арки иннервируют обоих IX и X черепных нервов (фиг.3В), которые могут быть удалены без воздействия на иннервацию к остальной уплотнительноеF жаберных дуг (рис 3C-E). Полноценного двустороннего результаты денервации в постепенной потере внешней иннервации на жаберных лепестках (4А-С). Рыба управления обладают очевидной нервного пучка, который охватывает длину нити (фиг.4А). Полноценного двустороннего денервация привело к некоторой потере внешней иннервации после 2 дней восстановления (рис 4б). Кроме того исчезновение внешней иннервации было отмечено после 5 дней восстановления от денервации (фиг.4С). Любой оставшийся иннервации с ИС через 5 дней восстановления предположительно происходит от нервов с клеточных тел в жаберной нити (внутренняя иннервации; фиг.4С).

Рисунок 1. Экспериментальная установка для процедуры денервации и последовательности шагов, используемых вВремя техника дифференциальной окраски. () Хирургическое столовый набор на процедуры денервации, показывая анестезирующие и восстановления танков. (B) Пример размещения в наркоза, интубированного рыбы. (C) Основные этапы используется во время техники дифференциального окрашивания. На шаге 1, рыба находится в контролируемой температурой, статическое аэрированной воды в ванне в течение 30 мин. Митохондрии, богатые краситель добавляют на стадии 2 до конечной концентрации 0,1 мкМ и рыба разрешается ванну в растворе в течение как минимум 4 часов. Расход воды возобновляется в шаге 3, в котором указывают рыба подвергается либо полноценного двустороннего денервации жаберных или фиктивный хирургии. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.
{kind=link}
 />
/>
Рисунок 2. Легкие микрофотографии, представляющая время технику дифференциального окрашивания в золотой рыбки (приспособились к 25 ° C) через 2 дня после полноценного двустороннего денервации. () Распределение уже существующих ionocytes (ИИ; стрелками). На одном жаберных нитей раскрывается с помощью окрашивания красителем митохондрий богатых (B) распределение ИС (новые и уже существующих) и жаберные нервы (пунктирные стрелки) Обнаружено окрашиванием с α-5 и Zn-12 антител, соответственно. Стрелки указывают ИС (C) перекрытие (А) и (В) отличает уже существующих ИС. (Отображается желтым цветом, с указанием стрелки) из вновь распространились ICS (горит зеленым светом, с указанием стрелками). Вставка в (С) представляет собой увеличение в качестве иннервируемой ранее существовавшего ionocyte. (D), характерную диаграмму, СК количественного для нити, показанной на панели (AC). N бар = 1. Шкала в панели (С) 50 мкм и распространяется на всех панелях./ftp_upload/52548/52548fig2highres.jpg "TARGET =" _ пустое "> Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.

Рисунок 3. Типичные изображения, показывающие разные стадии жаберных иннервации. () Спинной зрения ротовой полости, показывая черепные нервы IX и X, иннервирующие все 4 жаберных дуг. Жаберные дуги пронумерованы 1-4. Жаберные нити и ткани, покрывающей нервы были удалены, чтобы лучше визуализировать иннервации. (B) 1-й и 2-й жаберные дуги разделены, чтобы выявить орган чувств и ветви черепных нервов IX и X, иннервирующие 1-й жаберных арх. (CE) Последовательность изображений, показывающих избирательное денервации 1-й жаберной дуги путем разделения ветвей черепных нервов IX и X. Схема устройства Reпрезентация жаберных иннервации костистых рыб, обратитесь к рисунку 1 в Milsom и др 26 Белые линии в изображениях являются отражением воды от микроскопа огней.; они не определяют какие-либо морфологической структуры. Бар Шкала в панели (E) составляет 4 мм и применяется ко всем панелей. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.
{kind=link}

Рисунок 4. Легкие микрофотографии, изображающие распределение и иннервации ionocytes на одной нити 1-го жаберной дуги золотая рыбка приспособиться к 25 ° С Ionocytes (указано стрелкой глав) были окрашены с α-5 антитела и нервов (указывается бу стрелки) были окрашены с Zn-12 антителом. (А) Гилл нить контрольной рыбы, показывающий центральный пучок нервов, предположительно, происходящих из нервов IX и X (внешняя иннервации) с обширным пластинчатые ветвления. Некоторые из указанных ionocytes также иннервируют (вставка). (Б) Gill нити 2 дня после полной двусторонней денервации. Был сокращение центрального пучка нерва в то время как пластинчатые иннервации появился в основном без изменений. (С) жаберные нити 5 дней после полноценного двустороннего денервации, подтверждающие, что внешняя иннервации был в значительной степени отсутствует. Качественный анализ показывает, что полноценного двустороннего денервация вызывает деградацию внешней жаберных иннервации при сохранении нервы с сотового тел в жаберной нити (внутренняя иннервации) создание сети нервов через нить и в ламелями. Пожалуйста,Нажмите здесь, чтобы посмотреть большую версию этой цифры.
{kind=link}
Обсуждение
Время техника дифференциальной окраски может быть полезным инструментом, чтобы понять динамическое регулирование поглощения ионов и изучения временной перераспределение ИС в жаберной эпителия. Хотя простой процедуры, есть ряд ключевых моментов, которые имеют решающее значение для успеха времени техники дифференциального окрашивания. Золотая рыбка должна подвергаться митохондрии богатые краски для времени, отведенного в протоколе. Более короткими выдержками может привести к ухудшению поглощения красителя на митохондрий богатых клетками (например, ionocytes). Во время фиксации, жаберные ткани должны быть изъяты быстро и выдерживают в темноте, чтобы избежать фото отбеливание. Ткани должны быть обработаны для работы с изображениями в течение 2 недель фиксации. Во время двусторонней процедуры нерва секционирования убедитесь, что рыба хорошо анестезированной; нервы и кровеносные сосуды четко определены; и рыба возобновляет полный Крышечные функцию, прежде чем он перемещается в резервуар для извлечения.
"> Окружающая среда FW представляет рыбу с двойной проблемой балансирования пассивные потери ионов и осмотического усиление воды 14. Балансировка пассивных потерь ионов происходит с помощью активного поглощения соли через микросхем, которые локализованы в filamental и пластинчатой эпителия, где они могут прямой контакт с внешней средой 2,4,8,9,16. Тем не менее, расположение микросхем на жаберных не является статичным. За последние три десятилетия число исследований показали, что несколько FW видов рыб, когда они сталкиваются с ионной и / или температуры вызов, перераспределить жаберные ИС от нити или основание пластинки от более дистальных отделах пластинки 1,2,4,5,17-21. Такое перераспределение может увеличивать толщину пластин, которые может поставить под угрозу подачу газа (O 2, CO 2) через жабры эпителий 22. Исследователи использовали технику дифференциального окрашивания раз описанной в этой рукописи, чтобы отслеживать перемещение и emergencе новых микросхем на жаберных эпителия в этих различных экспериментальных условиях (рис 1в) 1,4,5.Жабры, и, предположительно, жаберные ИС, иннервируются IX и X черепных нервов 7,23-25. Эти нервы нести как эфферентные и афферентные входы и из жаберных соответственно. Они расположены на спинной стороне ротовой полости за 4-й жаберной дуги. Доступность нервов и легкость, с которой двусторонних процедура денервация могут совершаться определенные разновидности. В форели, например, острый и плоский анатомия головы позволяет нервы лежат в одной плоскости, за 4-й жаберной дуги под тонким слоем ткани. Это делает нервы видны и легко доступны для исследователя, чтобы выполнить процедуру денервации. В отличие от этого, золотая рыбка имеют более короткий рыло и круглее головой. В IX и X черепных нервов золотой рыбки лежат глубже в спинной сиде полости после 4-го жаберной дуги, занимающих различные самолеты. Эта ориентация ограничивает легкость доступа к нервам и требует более тщательного подхода к определению и разорвать соответствующие нервы. Целью процедуры денервации, чтобы удалить сенсорную афферентные и эфферентные вход и из жабр, соответственно. Денервации жаберных дуг также может быть соединен с потоком ионов экспериментов с использованием радиоизотопов (например, 22 Na). Эти методы могут быть использованы в тандеме для изучения вклада нервной входа на движение ионов через жабры эпителия. Другим ограничением в порядке денервации является неспособность различить сенсорных и моторных нейронов, таким образом, когда отсечения нервного пучка, возможно, что оба типа иннервации удаляются. Разрыв любые двигательные нейроны могут повлиять на жабры и Крышечные движение рыбы. Таким образом, при выполнении экспериментов жаберные денервации важно также контролировать вентиляцию послерыба оправилась от процедуры, чтобы обеспечить, что есть достаточное движение жаберных для газа, так и ионного обмена.
Протоколы, описанные в этой рукописи для использования взрослыми животных, содержащихся на 12:12 свет: темнота цикла и кормили коммерческих пищевых гранул. Эти методы могут быть модифицированы различными способами. Во-первых, период восстановления после воздействия красителя митохондрии, богатые могут быть адаптированы к требованиям протокола исследователя (например, 1, 3, 5 или 14 дней). Длительный период восстановления после воздействия красителя митохондрий богатых в лаборатории был 14 дней 4,5. Там не было значительное снижение интенсивности флуоресценции митохондрий богатых красителя после 14 дней восстановления. Во-вторых, использование первичного антитела α-5 ограничивается только идентификации NKA богатых клетки и не делает различий между различными подтипами бранхиальных ИС. К счастью, в золотых рыбок было установлено, что большинство микросхем являются mitochondrионно-богатых (этикетка с Mitotracker) и НКА богатых (этикетка с α-5) клеток, которые не могут быть дело в всех видов рыб 4,27. Дальнейшие эксперименты могут сосредоточиться на следующих временную перераспределение определенных подтипов IC с помощью антител, направленных именно против различных каналов, насосов и теплообменников (например, NHE, H + насос). Предыдущие исследования показали, что большинство ionocytes окрашенных выставочная краситель митохондрии богатые НКА иммунореактивности 4. Иннервации ИС могут быть обнаружены с помощью первичных антител против данио, полученных нейрона-специфического антигена (Zn-12). В этом исследовании, α-5 и Zn-12 первичные антитела были обнаружены с помощью того же вторичного антитела (Alexa Fluor 488) для обоих. Это ограничение обусловлено как первичные антитела поднимается в хозяине мыши и преодолевается тем, что НКА-богатые клетки и нейроны можно выделить морфологически, даже если они флуоресцируют и тот же цвет. Последовательное окрашиваниес различными вторичными антителами (например, первый α-5 с Alexa Fluor 488, то ZN-12 с Alexa Fluor 594) также может быть использована для устранения проблемы, связанной с обеих маркеров флуоресцирующих тот же цвет. И, наконец, полное двусторонний протокол нерва секционирования может быть изменен целевой нервы на конкретные жаберных дуг. Например, избирательное секционирования нерва может быть выполнена на первом жаберной дуги, осторожно отделяя первый и второй жаберных дуг, чтобы обнажить нервы, ведущие к первому жаберной дуги на спинной части жаберной корзины (фиг.2В-E). Дифференциальный время метод также может быть применен для изучения распределения ionocytes в личиночной данио рыбы, как они развиваются и ионно-транспортные переходы от кожи до жаберных.
Раскрытие информации
The authors declare that they have no competing financial interests.
Благодарности
We would like to thank William Fletcher for animal care at the University of Ottawa. The authors would also like to thank Dr. William Milsom for teaching VT the full bilateral denervation technique at the University of British Columbia. A travel grant for this research was provided by the Faculty of Graduate and Postgraduate Studies at the University of Ottawa. This research is also supported by NSERC PGS-D scholarship to VT and Discovery Grants Program to SFP.
Материалы
| Name | Company | Catalog Number | Comments |
| MitoTracker Red | Life Technologies | M-7512 | |
| Dimethyl sulfoxide | Sigma Alrdich | D2650 | |
| α-5 primary antibody | Develompental Hybridoma Bank | a5 | |
| zn12 primary antibody | Develompental Hybridoma Bank | zn12 | |
| Alexa Fluor 488 (anti mouse) | Life Technologies | A-11001 | |
| Benzocaine | Sigma Alrdich | E1501 | 4-aminobenzoic acid ethyl ester, ethyl 4-aminobenzoate |
| Vannas spring scissors | Fine Science Tools | 15000-10 | |
| Standard pattern forceps | Fine Science Tools | 11000-12 | straight |
| Standard pattern forceps | Fine Science Tools | 11001-12 | curved |
| Dumont No. 5 forceps | Fine Science Tools | 11252-30 | |
| Tissue retractor | Fine Science Tools | 17009-07 | |
| Paraformaldehyde | Sigma Alrdich | P6148 | |
| Triton X | Sigma Alrdich | X100 | |
| Vectashield with DAPI | Vector Laboratories | H-1200 |
Ссылки
- Katoh, F., Kaneko, T. Short-term transformation and long-term replacement of branchial chloride cells in killifish transferred from seawater to freshwater, revealed by morphofunctional observations and a newly established 'time-differential double fluorescent staining' technique. J. Exp. Biol. 206 (22), 4113-4123 (2003).
- Perry, S. F. Relationships between branchial chloride cells and gas transfer in freshwater fish. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 119 (1), 9-16 (1998).
- Sollid, J., Weber, R. E., Nilsson, G. E. Temperature alters the respiratory surface area of crucian carp Carassius carassius and goldfish Carassius auratus. J. Exp. Biol. 208 (6), 1109-1116 (2005).
- Mitrovic, D., Perry, S. F. The effects of thermally induced gill remodeling on ionocyte distribution and branchial chloride fluxes in goldfish (Carassius auratus). J. Exp. Biol. 212 (6), 843-852 (2009).
- Tzaneva, V., Vadeboncoeur, C., Ting, J., Perry, S. F. Effects of hypoxia-induced gill remodelling on the innervation and distribution of ionocytes in the gill of goldfish, Carassius auratus. J. Comp. Neurol. 522 (1), 118-130 (2014).
- Greco, A. M., Fenwick, J. C., Perry, S. F. The effects of soft-water acclimation on gill structure in the rainbow trout Oncorhynchus mykiss. Cell Tissue Res. 285 (1), 75-82 (1996).
- Jonz, M. G., Nurse, C. A. Epithelial mitochondria-rich cells and associated innervation in adult and developing zebrafish. J. Comp. Neurol. 497 (5), 817-832 (2006).
- Laurent, P., Hebibi, N. Gill morphometry and fish osmoregulation. Can. J. Zool. 67 (12), 3055-3063 (1989).
- Perry, S. F., Laurent, P. Adaptational responses of rainbow trout to lowered external NaCL concentration: contribution of the branchial chloride cell. J. Exp. Biol. 147, 147-168 (1989).
- Evans, D. H., Piermarini, P. M., Choe, K. P. The multifunctional fish gill: dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 85 (1), 97-177 (2005).
- Hwang, P. P., Lee, T. H., Lin, L. Y. Ion regulation in fish gills: recent progress in the cellular and molecular mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 301 (1), R28-R47 (2011).
- Krogh, A. The Active Uptake of Ions into Cells and Organisms. Proc. Natl. Acad. Sci. U. S. A. 25 (6), 275-277 (1939).
- Nilsson, G. E., Randall, D. J., Hoar, W. S. Innervation and pharmacology of the gills. Fish Physiology. , 185-272 (1984).
- Sundin, L., Nilsson, S. Branchial innervation. J. Exp. Zool. 293 (3), 232-248 (2002).
- Dymowska, A. K., Hwang, P. P., Goss, G. G. Structure and function of ionocytes in the freshwater fish gill). Respir. Physiol. Neurobiol. 184 (3), 282-292 (2012).
- Witters, H., Berckmans, P., Vangenechten, C. Immunolocalization of Na+/K+-ATPase in the gill epithelium of rainbow trout, Oncorhynchus mykiss. Cell Tissue Res. 283 (3), 461-468 (1996).
- Bindon, S., Fenwick, J. C., Perry, S. F. Branchial chloride cell proliferation in the rainbow trout, Onchorhynchus mykiss: implications for gas transfer. Can. J. Zool. 72 (8), 1395-1402 (1994).
- Bradshaw, J. C., Kumai, Y., Perry, S. F. The effects of gill remodeling on transepithelial sodium fluxes and the distribution of presumptive sodium-transporting ionocytes in goldfish (Carassius auratus). J. Comp. Physiol. B. 182 (3), 351-366 (2012).
- Chou, M. Y., et al. Effects of hypothermia on gene expression in zebrafish gills: upregulation in differentiation and function of ionocytes as compensatory responses. J. Exp. Biol. 211 (19), 3077-3084 (2008).
- Kaneko, T., Katoh, F. Functional morphology of chloride cells in killifish Fundulus heteroclitus, a euryhaline teleost with seawater preference. Fisheries Science. 70 (5), 723-733 (2004).
- Ouattara, N., et al. Changes in gill ionocyte morphology and function following transfer from fresh to hypersaline waters in the tilapia Sarotherodon melanotheron. Aquaculture. 290 (1-2), 155-164 (2009).
- Bindon, S., Gilmour, K., Fenwick, J., Perry, S. The effects of branchial chloride cell proliferation on respiratory function in the rainbow trout, Oncorhynchusmykiss. J. Exp. Biol. 197 (1), 47-63 (1994).
- Dunel-Erb, S., Bailly, Y., Laurent, P. Pattern of gill innervation in two teleosts, the perch and the trout. Can. J. Zool. 71, 18-25 (1993).
- Jonz, M. G., Nurse, C. A. New developments on gill innervation: insights from a model vertebrate. J. Exp. Biol. 211 (15), 2371-2378 (2008).
- Jonz, M. G., Zaccone, G. Nervous control of the gills. Acta Histochem. 111 (3), 207-216 (2009).
- Milsom, W. K., Reid, S. G., Rantin, F. T., Sundin, L. Extrabranchial chemoreceptors involved in respiratory reflexes in the neotropical fish, Colossoma macropomum (the tambaqui). J. Exp. Biol. 205 (12), 1765-1774 (2002).
- Hwang, P., Lee, T. New insights into fish ion regulation and mitochondrion-rich cells. Comp Biochem Physiol A Mol Integr Physiol. 148 (3), 479-497 (2007).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены