Method Article
イメージゼブラフィッシュアイ開発にライトシート蛍光顕微鏡を用いて、
要約
Light sheet fluorescence microscopy is an excellent tool for imaging embryonic development. It allows recording of long time-lapse movies of live embryos in near physiological conditions. We demonstrate its application for imaging zebrafish eye development across wide spatio-temporal scales and present a pipeline for fusion and deconvolution of multiview datasets.
要約
Light sheet fluorescence microscopy (LSFM) is gaining more and more popularity as a method to image embryonic development. The main advantages of LSFM compared to confocal systems are its low phototoxicity, gentle mounting strategies, fast acquisition with high signal to noise ratio and the possibility of imaging samples from various angles (views) for long periods of time. Imaging from multiple views unleashes the full potential of LSFM, but at the same time it can create terabyte-sized datasets. Processing such datasets is the biggest challenge of using LSFM. In this protocol we outline some solutions to this problem. Until recently, LSFM was mostly performed in laboratories that had the expertise to build and operate their own light sheet microscopes. However, in the last three years several commercial implementations of LSFM became available, which are multipurpose and easy to use for any developmental biologist. This article is primarily directed to those researchers, who are not LSFM technology developers, but want to employ LSFM as a tool to answer specific developmental biology questions.
Here, we use imaging of zebrafish eye development as an example to introduce the reader to LSFM technology and we demonstrate applications of LSFM across multiple spatial and temporal scales. This article describes a complete experimental protocol starting with the mounting of zebrafish embryos for LSFM. We then outline the options for imaging using the commercially available light sheet microscope. Importantly, we also explain a pipeline for subsequent registration and fusion of multiview datasets using an open source solution implemented as a Fiji plugin. While this protocol focuses on imaging the developing zebrafish eye and processing data from a particular imaging setup, most of the insights and troubleshooting suggestions presented here are of general use and the protocol can be adapted to a variety of light sheet microscopy experiments.
概要
形態形成は、胚を整形し、一緒に成長および分化と成熟した多細胞生物への受精卵から個体発生を駆動する方法です。動物発生中の形態形成のプロセスは、無傷の生きた検体1-3の撮像によって最良に分析することができます。このような全胚イメージングが駆動し、シグナル伝達分子の傾き、細胞外マトリックス、血管系、神経支配だけでなく、周囲の組織の機械的性質などの開発を規制するすべてのコンポーネントを保存するためです。形態形成が発生するスケールを、埋めるために、高速な細胞内イベントは、時間または数日間にわたる全組織の発達との関連で分の時間スケール上に捕捉されなければなりません。これらのすべての要件を満たすために、直交する平面照明顕微鏡5の現代の実装4を開発しました。もともと、それは選択的平面照明顕微鏡(SPI命名されましたM)4。今包括的な用語ライトシート蛍光顕微鏡(LSFM)は、典型的に使用されます。レーザ走査またはスピニングディスク共焦点顕微鏡6,7未満の光毒性を誘導しながらLSFMは、高時間分解能での撮像を可能にします。今日では、そこに基本的な光シート照明原理の多くの実装は既にあり、それは研究者8月11日に以前にアクセスできない標本やプロセスの多種多様な画像に使用されてきました。
まず、従来の共焦点顕微鏡のアプローチよりLSFMのいくつかの重要な利点を強調したいと思います:
ライブイメージング顕微鏡実験から意味のある結果を得るためには、観察のみ最小限試料に影響を与えることが重要です。しかし、ゼブラフィッシュを含む多くの生物は光毒性エフことなく、高時間分解能を持つ共焦点顕微鏡で画像彼らにそれが困難に、レーザ光照射に対して非常に敏感ですストールや発達の遅れ6,7のようなfects。 LSFMは現在、試料7上の少なくとも破壊的な効果を持つ蛍光イメージング技術です。薄いレーザー光シートは、特定の時点で撮像された試料の一部のみを照射するので、光シート顕微鏡は非常に効率的に光子を使用しています。その結果、低露光量は、健康な標本の長いタイムラプス観察、 例えば 12-17を可能にします。さらに、LSFMの最小侵襲性のおかげで、取得された画像の数はもはや、むしろ処理して保存することができますどのくらいのデータでサンプルが許容できる光の量によって決定されていません。
近くの生理的条件で試料を維持するという点では同じラインに沿って、LSFMは、ライブ胚に非常に適して代替サンプル実装戦略が付属しています。 LSFM技術では、胚は、典型的には、低い割合アガロースの薄いカラム内に埋め込まれています。 mountinアガロースシリンダへGは、回転の完全な自由を可能にするため、試料は完全な角度(LSFMにビューと呼ばれる)のと同時に、複数のビューから観察することができます。マルチビュー撮影とその後の多眼融合が大きな散乱標本のために特に有益であると高い、等方性の解像度でそれらをキャプチャすることができます。他の可能LSFM実装戦略の概要はE.レイノーのラボによって書かれた試料調製の章では、公式の顕微鏡取扱説明書に記載されています。それは、ここで説明するよりも、ゴールはイメージの異なるサンプルにある場合は特に、推奨読み取りです。
LSFMの画像取得は、レーザ走査型共焦点顕微鏡とは対照的に、広視野、カメラベースです。これは、より高い信号対取得された画像のノイズ比(SNR)になると(数十秒あたりのフレーム数百)非常に高速であることができます。 LSFMの高感度をさらに弱い蛍光SAMPのイメージングを可能にします近い将来の内因性レベル18か、で発現する転写因子、CRISPR / Cas9を使用してタグ付けされた内因性タンパク質のようなレ、。高いSNRはまた、成功した下流の画像解析のために重要です。高速は十分に速く、複数のビューから全胚をするだけでなく、迅速な細胞内プロセスをキャプチャするために必要なだけでなく、画像にされています。観察された現象は、別々のビューからこれらの複数のZスタックの取得中に変化しない場合、複数のビューのシームレスな融合は、達成することができます。
LSFMの利点は、典型的には、画像品質を犠牲にして来ることはありません。 LSFMの横方向の解像度は、共焦点顕微鏡の解像度よりも若干悪くなります。 LSFMに使用される検出の目的は、標準的な共焦点セットアップに水またはシリコン液浸対物レンズの1.2〜1.3と比較してより低い開口数(通常1.0以下)を有するためです。さらに、LSFMで広視野検出のために(absenピンホールのCE)は、共焦点顕微鏡に比べ、より焦点の外れた光があります。ピンボケの光の量を光シート厚さによって決定されます。それにもかかわらず、これらの欠点はLSFMより高いSNRによって補償されます。実際には、これは、例えば、スピニングディスク共焦点取得15と比較して同様の品質の画像が得られます。したがって、これは15,19をトレースする細胞系譜用の細胞膜や核、 例えば 、などの機能の信頼性の抽出を可能にします。
LSFMの軸方向の分解能は光シート厚によって、検出目的に加えて、決定されます。 LSFMの軸方向の解像度は、いくつかのケースでは、共焦点顕微鏡の解像度を上回ることができます。光シートは、典型的には、低倍率の対物レンズと撮像された大規模な標本のために発生し、検出目的の軸方向の解像度よりも薄い場合まず、解像度の向上が付属しています。 LSFMがACHできるか第二の方法、より良好な距離分解能をieve、異なるビューからの高XY解像度情報は、1つの画像スタックに結合されたマルチビュー融合です。得られたマージされたスタックは、横方向20,21に解像度の値に近づく等方性解像度を有します。この資料に記載されてお互いの上に複数のビューを登録するための戦略は、試料20,21の周囲のアガロース中に埋め込 まれた受託者のマーカーとして蛍光ポリスチレンビーズを使用することに基づいています。
LSFMの商業化の結果として、この技術は、科学者22の広いコミュニティに利用可能になりました。したがって、このプロトコルを記述するための動機はLSFMで実務経験を欠く発達生物学者にこの技術がアクセスできるようにすることで、これらの科学者たちは、彼らのサンプルを用いて、この技術の使用を開始します。私たちのプロトコルは、概念的には単純顕微鏡トンを構成している商用光シート顕微鏡を使用しています帽子は、操作が簡単です。我々はさらに、特定の質問23-25 に答えることが適当であるかもしれない自作LSFMのセットアップ、とゼブラフィッシュを撮像するための他の最近のプロトコルに言及したいと思います。 LSFMへの他のエントリのオプションは、より広範なコミュニティにシート光顕微鏡をもたらすためにオープンアクセスの原則を使用するオープンプラットフォーム26,27、です。ハードウェアとソフトウェアの両面のドキュメントはhttp://openspim.orgとhttps://sites.google.com/site/openspinmicroscopy/で見つけることができます。
このプロトコルでは、我々はLSFMと発達過程を研究するためのモデル系として硬骨魚類のゼブラフィッシュを使用します。ゼブラフィッシュの眼の形態形成はLSFMの多くの利点を強調した例です。 LSFM既にメダカ28およびゼブラフィッシュ29,30に眼の発達を研究するために過去に使用されてきました。眼の開発の初期段階では、従来の顕微鏡検査のために正しく胚を配向する複雑さ、かさばる卵黄として胚が客観的に面した目で側にうそをつくことはできません。しかし、LSFMアガロースカラムに搭載して、サンプルを再現可能に位置決めすることができます。また、光学カップステージへ眼胞からの移行中に、目は大のzスタックおよびビューの大きなフィールドを取り込む必要とする、成長に伴う主要な形態形成の再編成を受けます。また、これらの課題のためにLSFMは、従来の共焦点イメージングに優れています。眼杯の形成プロセスは、したがって、理解し、一つのビューからの撮像のみによって可視化することが困難であり、三次元です。これは、等方性の解像度を持つマルチビュー撮影が有益になります。眼杯形成後、網膜は、レーザー露光にますます敏感になります。したがって、LSFMに関連する低毒性長期イメージングのための大きな利点です。
ここでは、古いゼブラフィッシュの胚1〜3日のイメージングのために最適化されたプロトコルを提示します眼の開発を中心とした目の幼虫。我々の方法は、高い空間分解能と時間分解能で12-14時間までカバーするタイムラプス動画の撮影が可能になります。この技術は、常に多くの場合、テラバイトの範囲で、大きなデータセットを生成するように重要なのは、我々はまた、LSFMに不可欠なステップであるデータ処理用のパイプラインを示しています。
プロトコル
注:すべての動物の作業は、欧州連合(EU)指令63分の2011 / EUとドイツの動物保護法に従って行いました。プロトコルは、サンプルを撮像するために実装から、中断することなく続くことを意味します。実践的な経験に応じて、タイムラプス実験を開始するために、2〜3時間かかります。データ処理は、この時間の計算には含まれません。実験に必要なすべての材料は、補足文書として提供され、その開始前に必要とされる材料のチェックリストに記載されています。手順1、2、3と4の手順2、3、4の場合、プロトコルはまた、公式顕微鏡取扱説明書を参照してくださいの5中に粉末フリーの手袋を着用してください。
イメージング実験の前に1.準備作業
- 蛍光ビーズ原液
- このプロトコルは、(赤色の発光蛍光色素で標識された)500または1000nmの直径のポリスチレンビーズを使用します。ビーズの希釈率は1:4、000まず、ボルテックスで1分間ビーズ原液。 ddH 2 Oを990μlのビーズを10μlを希釈4℃で暗所でのソリューションを保存し、この1使用:1:40でさらに希釈用原液として100希釈を。
- 試料室のためのソリューションを準備
- 100ミリリットルのビーカーミックスメチレンブルーせずにE3培地の38.2ミリリットル、10 mMのN個の -phenylthioureaの0.8ミリリットルと1ミリリットル0.4%MS-222で。後に試料室内に浮遊粉塵の小粒子または溶解していない結晶を避けるために、濾過E3媒体を使用することが有益です。
- 蛍光魚の胚の選択
- 実験前の日では、蛍光タンパク質を発現する胚を準備します。右実験前、蛍光信号の望ましい強さのための蛍光ステレオスコープの下で胚を並べ替えます。 5-10健全な胚を取り出し、ピンセットを使用してそれらをdechorionate。
注:このプロトコルは、16から72時間の古いために最適化され胚。
- 実験前の日では、蛍光タンパク質を発現する胚を準備します。右実験前、蛍光信号の望ましい強さのための蛍光ステレオスコープの下で胚を並べ替えます。 5-10健全な胚を取り出し、ピンセットを使用してそれらをdechorionate。
2.試料室の設定
- スリー商工会議所のWindowsの組み立て
- (厚さ0.17ミリメートル選択直径18mm)試料室4のウィンドウ、目的のための1つの三カバースリップで密封することがあります。 70%エタノールでこれらのカバースリップを保管してください。エーテルで使用する前に、きれいにそれらをワイプ:エタノール(1:4)。
- 細かい鉗子を使用してウィンドウにカバースリップを挿入し、それが最小の溝に入っていることを確認します。 17ミリメートルの直径のゴム製のOリングでそれをカバーし、チャンバー窓ツールを用いた照明アダプタリングにねじ込みます。他の二つのウィンドウのためのプロセスを繰り返します。
- 残りの商工会議所のパーツの取り付け
- チャンバーの第残りの側に適切な目的のためのアダプターをねじ込みます。アダプターの中央に直径15mmのOリングを挿入します。
- 茶中の右下の開口部に白のルアーロックアダプターにねじ込みmberと左上開口部の灰色のドレインコネクタ。黒ブラインドプラグが付いている3つのすべての残りの開口部をブロックします。
- ペルチェブロックと六角レンチを用いてチャンバの金属ダブテールスライド底部を取り付けます。ルアーロックアダプタに50ミリリットルの注射器でホースを取り付けます。チャンバー内に温度プローブを挿入します。
- 顕微鏡に目標と商工会議所の挿入
- すべての目標はきれいでステレオスコープの下で確認してください。 10X / 0.2照明の目標と計画-アポクロマート20X / 1.0 W検出目的を使用し、その屈折率補正カラーは水が1.33に設定されていることを確認します。覆われた照明の目標を維持しながら、顕微鏡に検出目標をねじ込みます。
- 照明の目的をカバーするプラスチック製の保護キャップを取り外してください。注意深く顕微鏡にチャンバーをスライドさせて固定しているネジでそれを締めます。
- 温度プロを接続します顕微鏡でペルチェブロックであると。
注:ペルチェブロックの冷却液を循環させる2つの管は、両方のコネクタと互換性があります。それらにかかわらず、接続の方向の機能回路を形成します。 - 室の窓の上端までのステップ1.2で調製した溶液にシリンジを介してチャンバを埋めます。チャンバが漏れていないか確認してください。
- 顕微鏡、インキュベーションおよび制御およびストレージコンピュータを起動します。顕微鏡操作ソフトウェアを起動し、28.5℃のインキュベーション温度を設定します。
注:それは完全に平衡に1時間かかります。その間にサンプルを準備します。
3.サンプルの調製
- アガロースミックスの準備
- アガロース混合物の製造前に、15分、70℃に設定した加熱ブロックに(E3培地に溶解)を1%低融点アガロースの1ml量を溶かします。アガロースが完全にモルになると10、E3培地250μlの0.4%MS-222を50μlとボルテックスビーズストック溶液25μlを追加し、新しい1.5mlチューブに600μlのを転送します。
注:追加の75μlのは、胚と後で一緒に添加される液体のために計算されている。これは、ミックスの925μLを作ります。 - 38-40°Cで第2の加熱ブロックにチューブを維持またはアガロースがそれにサンプル胚を入れる前に、そのゲル化点に非常に近いことを確認してください。
- アガロース混合物の製造前に、15分、70℃に設定した加熱ブロックに(E3培地に溶解)を1%低融点アガロースの1ml量を溶かします。アガロースが完全にモルになると10、E3培地250μlの0.4%MS-222を50μlとボルテックスビーズストック溶液25μlを追加し、新しい1.5mlチューブに600μlのを転送します。
- 胚をマウントします
- (〜1ミリメートル、内径、ブラックマーク付き)20μl容量の5ガラスキャピラリーを取り、それらに一致するテフロン先端プランジャを挿入します。テフロン先端がキャピラリーの底にあるように、毛細管を通してプランジャーを押してください。
- ボルテックス37°C暖かいアガロースミックスのチューブにガラスやプラスチックピペットを用いて転送5胚(一度に取り付けることができる番号)。
注:液体一緒にウィットの最小体積をキャリーオーバーしてみてください時間の胚。 - ミックスにキャピラリを挿入し、プランジャーを引き上げによって内部1胚を吸います。胚の頭部が尾前毛細血管に入ることを確認してください。プランジャと試料との間の気泡を避けてください。アガロース胚上記の±2 cmであり、その下の±1センチメートルがあるはずです。残りの胚について繰り返します。
- アガロースが完全に固化するまで数分以内に起こる、待ってから、粘土やテープを入れたビーカーの壁にそれらを貼り付けることで、E3培地でサンプルを格納。キャピラリーの底部開口部は、サンプルへのガス交換を可能にする溶液中で自由にぶら下がってされるべきです。
注:セクション3.1および3.2と同様のプロトコルもOpenSPIM wikiページのhttp://openspim.org/Zebrafish_embryo_sample_preparationで見つけることができます。
4.サンプルポジショニング
- サンプルホルダ・アセンブリ
- 正しいサイズの2プラスチック製スリーブ(黒)を挿入サンプルホルダーステムに互いに反対。そのスリット辺が外側に直面しなければなりません。それに2-3ラウンドを回して緩くクランプネジを取り付けます。クランプネジを介してキャピラリを挿入し、黒色のバンドは他の側に見えるようになるまで、ホルダーを介して、それを押してください。プランジャーを触れないようにしてください。
- クランプネジを締めます。キャピラリーの外胚以下アガロースの過剰1cmにプッシュし、それをカット。試料ホルダーのディスクにステムを挿入します。
- 顕微鏡ステージがロード位置にあることをソフトウェアで確認します。顕微鏡に垂直下方サンプルと全体ホルダーを滑空する案内レールを使用してください。そのため、磁気ホルダーディスクロックの位置に、それを回します。
- キャピラリーの検索
- これからは、ソフトウェアによってサンプルの位置を制御します。 探しタブで見つけ毛細管オプションを選択し、ちょうど検出オブジェクトの上に焦点にxの毛細血管、yおよびzを配置アイブレンズ。指導のための検体ナビゲータでグラフィカルな表現を使用してください。
- それは、検出目的の瞳の前にあるまで、キャピラリーの外に静かに胚を押してください。
注:「見つけキャピラリー」は、顕微鏡の上蓋が開かれるべきであり、試料が押し出される時に、残りのプロトコル、で唯一のステップです。
- サンプルの検索
- 「サンプルを見つけ 'オプションに切り替え、0.5ズームで視野の中心にゼブラフィッシュアイをもたらします。それは目に到達する前にシート光が試料の任意の高屈折率または吸収部を通過しないように、胚を回転させます。同様に、放出される蛍光は、試料のうち明確なパスが必要です。 「設定ホームポジション」をクリックします。
- 顕微鏡のフロントドアを開き、蒸発を避けるために、チャンバーの上部に3mmの開口部を有するプラスチック製のカバーを置きます。
注:液面が下回る場合イメージングレベルは、実験が損なわれることになります。 - 全体的な健康のためのプロキシとして胚の心拍を確認してください。それは遅すぎる場合は、別のサンプルを使用します(非装着され、コントロールと比較し、特定の値は、発達段階に依存)。最終的なズーム設定に切り替えて、胚の位置を再調整します。
5.多次元の取得を設定します
- 取得パラメータ
- 「買収」タブに切り替えます。レーザーライン、検出目的、レーザーブロッキングフィルタ、ビームスプリッタやカメラなどの光パスを定義します。
- ピボットスキャン]チェックボックスを有効にします。ビット深度、画像フォーマット、光シートの厚さのような他の取得設定を定義し、片面照明を選択します。
- プレス '連続'とレーザパワーとカメラの露光時間を変更して得られた画像の輝度に応じました。
注:すべての画像化の設定を調整するためのレを使用試料への不要な光損傷を避けるために、実際の実験よりもレーザー出力(100mWのレーザーの0.5%、30ミリ秒の露光時間)。
- 光シート調整
- 「両面イルミネーション」とに切り替え 「オンラインデュアルサイドFusio'nチェックボックスを有効にしてください。 「Lightsheet自動調整ウィザード」を起動します。ステップの指示のステップに従ってください。
注:このウィザードは、検出目的の焦点面に光シートを移動させ、それが傾いていないことを保証し、その腰には視野の中心です。自動調整を終えた後、左右の光シートの位置が自動的にソフトウェア更新されます。画質の改善が現在明らかです。 「Z-スタック」チェックボックスを有効にします。 - XZとYZオルトビューで蛍光ビーズで与えられる点広がり関数(PSF)の対称性を検査することにより、光シート調整を確認してください。それはノーである場合トンの対称は、手動で上下対称の砂時計形のPSF( 図1A)を達成するまで光シートのパラメータの位置を調整します。
- 「両面イルミネーション」とに切り替え 「オンラインデュアルサイドFusio'nチェックボックスを有効にしてください。 「Lightsheet自動調整ウィザード」を起動します。ステップの指示のステップに従ってください。
- 多次元取得の設定
- 「最初のスライス」と「最後のスライス」オプションを使用して、zスタックを定義し、1ミクロンへのzステップを設定します。
注:この顕微鏡の光シートは、静的であり、zの切片を介して試料を移動させることによって達成されます。常に「連続駆動 'オプションは、zスタックの速い獲得のために使用します。 - 「時系列」チェックボックスを有効にします。時点の数と、それらの間で間隔を定義します。
- 「マルチビュー」チェックボックスを有効にします。 X、Y、Zと角度情報が格納されているマルチビューリスト、に現在のビューを追加します。キャピラリーを回転させて他の所望のビューを定義するためにステージコントローラを使用してください。各ビューでzスタックを設定し、マルチビューリストに追加します。
注記:画像が取得されている間に、毛細管は、一方向になっているように、ソフトウェアは、シリアル方式でビューをソートします。
- 「最初のスライス」と「最後のスライス」オプションを使用して、zスタックを定義し、1ミクロンへのzステップを設定します。
- ドリフト補正と実験を開始
- セットアップ買収が完了すると、実際の実験を開始する前に15〜30分間待ってください。
注:サンプルは、最初のx、y、zのに数μmを漂うが、30分以内に停止する必要があります。サンプルが長いために漂流し続ける場合は、別のサンプルまたは再マウントを使用しています。 - 「メンテナンス」タブに切り替え、「ストリーミング」オプションで各ビューし、チャンネルの時点または個別のファイルごとに1つのファイル、 例えば 、データを保存する方法を定義します。プレス '実験を開始します」とファイル名、それは保存する場所とファイル形式(使用.czi)を定義し、戻って「取得」タブに移動します。
- すべてが完璧に動作していることを確認するために、最初の時点の取得を確認します。すぐに登録を進めますそしてステップ6及び7に記載のように、第1の時点の融合、データセット全体を処理することが可能となることを確認します。
- セットアップ買収が完了すると、実際の実験を開始する前に15〜30分間待ってください。
6.マルチビュー登録
- マルチビュー復興アプリケーション
- イメージングセッションの終了時に、データ処理用コンピュータに顕微鏡でデータストレージコンピュータからデータを転送します。 「マルチビューを使用します ReconstructionApplication「20,21,31は、データ処理のためのフィジー32( 図1B)に実装されています。
- データセットを定義します。
- 更新フィジー:フィジー>ヘルプ>アップデートのImageJとフィジー>ヘルプ>フィジーを更新します。メインのImageJとフィジー更新サイトを使用してください。
- 一つのフォルダにデータセット全体を転送します。処理の結果と中間ファイルがこのフォルダに保存されます。フィジー>プラグイン>マルチビュー復興> Multivi:マルチビュー復興アプリケーションを起動します。EW復興アプリケーション。
- 「新しいデータセットを定義する」を選択します。データセットの種類としてはMEUオプション」ツァイスLightsheet Z.1データセット(LOCI Bioformats)」を選択し、.xmlファイルの名前を作成します。その後、データセット(インデックスなしつまりファイル)の最初の.cziファイルを選択します。なお、画像データだけでなく、記録のメタデータが含まれます。
注:プログラムが最初.cziファイルを開くと、メタデータがプログラムにロードされます。 - アングル、チャンネル、イルミネーションの数ていることを確認し、メタデータからボクセルサイズを観察します。フィジーのエラーメッセージの表示、処理およびその結果、「ViewSetupエクスプローラー'とコンソールの進捗状況を示す、ログウィンドウ:[OK]を押すと、開いている3つの別々のウィンドウ( 図1C)を観察。
注:「ViewSetup Explorerは、「各ビュー、チャンネルや照明を示しているとの選択を可能にするユーザーフレンドリーなインターフェイスであります興味のあるファイル。さらに、「ViewSetup Explorerは、「すべての処理ステップのステアリングを可能にします。 - 処理する必要のあるファイルを選択し、エクスプローラの中にマウスの右ボタンを押してください。異なる処理ステップ( 図1C)で開いているウィンドウを観察します。
- データセットを定義する際に、データのフォルダ内の.xmlファイルが作成されていることを確認します。
注:このファイルは、以前に確認されたメタデータが含まれています。 - 右上の二つのボタンは、 'info'を観察し、「保存」。 「情報」を押すと、.xmlファイルの内容の要約が表示されます。 「保存」を押すと、処理結果を保存します。
注:「マルチビュー復興アプリケーション」で処理している間に別の処理ステップは、フィジーを閉じる前に、.xmlファイルに保存する必要があります。
OAD / 53966 / 53966fig1.jpg "/>
図1:マルチビュー復興ワークフローと関心点検出蛍光ビーズイメージングに基づいて(A)ライトシート整合。ビーズの画像は、XZ及びYZ突起対称砂時計形状を有する場合、システムは、(中心)位置合わせされます。いずれかの方向にずれて光シートの例には、左と右に示されています。 XZで500 nmのビーズの最大強度投影は、YZとXY軸が示されています。 lightsheetが正しく調整不良の状況に比べて整列されたときにサイドローブへ(XYで)エアリーディスクの中心ピークの強度の比が、大きくなることに注意してください。画像は0.7ズームで20X / 1.0 Wの目的で撮影しました。スケールバーは5μmで表します。 (B)のデータセットを定義し、その後HDF5形式に再保存されています。ビーズは、セグメント化され、その後、登録されています。時系列の各時点を基準時点に登録されています。データは、最終的に融合されています単一等方性ボリューム。 (C、トップ)ViewSetup Explorerは 、データセットの異なる時間点、角度、チャネルおよび照明側面を示しています。 (Cは 、下の隅を左)BigDataViewerウィンドウがViewSetupエクスプローラで選択された図を示しています。 ViewSetupエクスプローラに(C、右中) を右クリックして、処理オプションを開きます。 (Cは 、右下隅)進捗状況や処理結果がログファイルに表示されます。 (DおよびE)の検出の目的は、ここでは、対話型ビーズセグメンテーションからのスクリーンショットのように示すことが可能である試料中のわずかの検出とセグメントとして多くの関心点(ビーズ)にあります。 (DとE、左上隅)セグメント化は二つのパラメータ、 シグマ1およびスレッシュホールドのための違い-のガウス値によって定義されています。(D)が正しく検出されたビーズの拡大図で成功検出例。あまりにも多くの偽陽性および単一のビーズの複数の検出と(E)セグメンテーション。 (C)中のスケールバーは50μmで表す。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- HDF5形式でデータセットを再保存
- データセット全体を再保存するには、Ctrl /アップル+ aと、右クリックですべてのファイルを選択します。その後、 再保存データセットを選択し、HDF5など 。
- ウィンドウには、現在のデータセットのすべてのビューが再保存される警告表示が表示されます。 [ はい ]を押します。
注:このプログラムは、すべてのファイルを開くとHDF5形式の異なる解像度レベルを再保存することにより、HDF5する.cziファイルを再保存していきます。これは、完了時に「行って」で確認します。 usuall再保存yは、時点ごとに数分かかります( 表1参照)。
注:HDF5形式のファイルは非常に高速にロードすることができますので、エクスプローラに「右クリック」とトグル」(オン/オフ)BigDataViewerの表示」で登録されていないデータセットを表示することが可能になりました。 BigDataViewerウィンドウが選択されたビュー( 図1C)が表示されます。 BigDataViewer 33のコア機能は、 表2で説明しhttp://fiji.sc/BigDataViewerされます。
- 関心点を検出
- 特徴点を検出し選択するには、Ctrl /アップル+ Aですべての時間ポイントを選択し、右クリックします。
- 差のガウス 34 関心点検出のタイプのを選択します。ビーズベースの登録は、ラベルの関心点のためにフィールドにタイプ「ビーズ」、ここで使用されているので。 「セグメンテーションの前に画像のダウンサンプリング」アクティブにします。
- 次のウィンドウで、DETを観察ection設定。 「サブピクセルローカライズ者用「3次元二次フィット」34とi'nterestポイントの指定」の「インタラクティブ」ビーズセグメンテーション用差のガウス値と半径を決定します。
- 「ダウンサンプルXY「使用」マッチZ解像度(以下ダウンサンプリング)」および「ダウンサンプルZ 'の使用1×ください。 zのステップサイズは、従って、単にダウンZ分解能が十分であると一致するように、XYサンプリング、X-Yピクセルサイズよりも大きいです。 「CPU(JAVA)上の計算」を選択します。押して「OK」。
- ポップアップウィンドウで、ドロップダウンメニューからパラメータをテストするための1つのビューを選択します。ビューがロードされると、フィジー>イメージ>調整>明るさ/コントラストまたはCtrl + Shiftキー+ Cでダニとビーズ検出用のボックス」の最大値のための外観(緑)」ウィンドウの明るさとコントラストを調整します。
- aroun 'viewSetup」として緑色のリングにセグメンテーションを守ってD検出最小値のための最大値と赤を探していたとき。セグメンテーション、2つのパラメータ、 シグマ1用差のガウス値としきい値 ( 図1D)によって定義されています。セグメントのサンプルの周りなど、多くのビーズと( 図1D)できるだけ試料内のわずか誤認検出にそれらを調整します。一度だけではなく、複数回( 図1E)を各ビーズを検出します。最適なパラメータを決定した後、Enterキーを押して[完了します]。
注:検出時点の各個々のビューをロードし、ビーズをセグメント化することによって開始します。ログファイルでは、プログラムは、ビューごとに検出されたビーズの数を出力します。検出は、数秒で行われるべきである( 表1参照)。 - 検出量が適切であるときにEnterキーを押して、(ビュー当たり数千600)を保存します。
注:フォルダには、情報が含まれているであろう、データディレクトリに作成されます検出されたビーズの座標についてのn。
- インタレスト・ポイントを使用して登録
- Ctrlキー/アップル+を持つ全ての時点を選択し、右側の「インタレスト・ポイントを使用して登録する」をクリックして選択します。
- 「登録アルゴリズム」などのビーズ検出のための「高速3D幾何学的ハッシング(回転不変) 'を使用します。
注:このアルゴリズムは、互いに対して異なるビューの向きや位置についての事前の知識を負いません。 - お互いへのビューの登録については、「登録の種類」として「個別時点を登録する」を選択します。選択したチャンネルの「関心点」については、関心点のために以前に指定したラベルは、現在( すなわち 「ビーズ」)を表示する必要があります。
- 次のウィンドウで、登録のための事前設定値を使用します。 「修正タイル:最初のタイルとバックマップしないでください使用を修正」を選択することにより、第1のビューを修正ゴマ場合(これを使用エスは、「タイルをバックマップ」セクションに)固定されていません。
- 「正則」と「アフィン変換モデル」を使用してください。
注:RANSACのための許容される誤差は5pxのであろうと、ディスクリプタ一致の重要性は、 ''正則」変換が10%剛性および90であることを意味し0.10のラムダ、と「剛体モデル」を使用してください10.になります%アフィン35。 OKを押して登録を開始します。
注:ログ・ウィンドウに表示されるように、最初の各ビューは、他のすべてのビューと一致しています。そして、ランダムサンプルコンセンサス(RANSAC)36は、対応をテストし、偽陽性を除外します。堅牢な登録のために、RANSAC値が90%よりも高くする必要があります。二面図の間で対応する真の候補の十分な数が発見された場合、変換モデルは、画素における平均変位に各マッチの間で計算されます。そして、反復グローバル最適化が行われ、すべてのビューされます固定ビューに登録されています。登録が成功すると、変換モデルが計算され、そのスケーリングおよびピクセルの変位で表示します。平均誤差は、最適に1ピクセル未満でなければならないし、1に近い転換登録のスケーリングが( 表1参照)秒単位で実行されます。 - サンプル、 例えば、細胞膜内の微細構造に観察されるように異なるビューの間にずれがないことを確認してください。その後の.xmlファイルにビューごとに変換を保存します。
注:今すぐ登録ビューがBigDataViewer( 図2A)で互いに重複しているとビーズの画像も同様にオーバーレイされるべきである( 図2B)。 - それらの上で右クリックした時点を選択することによって変換を削除してから、変換を削除]を選択>最新/最新の変容。
2再 "SRC =" /ファイル/ ftp_upload / 53966 / 53966fig2.jpg "/>
図2:マルチビュー再構成の結果 (A)は 、それらの間の重複を示すために異なる色でそれぞれ、登録されたビューをオーバーレイさ。 (B)は、異なるビューから撮像ビーズのPSFの重なりを示す図に拡大します。融合後のビーズのアップを閉じる(C)は 、PSFは異なるビューの平均です。 PSFは一点に潰れることを示すマルチビューデコンボリューション後の(D)(C)と同じビーズのPSF。膜は、GFPはより深い組織におけるzにおける信号の劣化を示すで標識された眼胞の単一のビューの(E)のxy断面および(F)YZ断面。 (G)のxyセクションと少しdegraで約20度離れて4ビューの加重平均融 合後の同じビューの(H)yz断面全体のDED XY解像度が、増加のz解像度。 (I)のxyセクションと解像度とXYであり、zの両方の信号のコントラストの有意な増加を示すマルチビューデコンボリューション後の同じデータの(J)YZ断面、。 (EJ)内の画像は、単一の光のスライスです。スケールバーは50μmで(A、B、EJ)および10ミクロン(C、D)を表し、この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- タイムラプス登録
- 全体の時間の経過を選択し、右の時間をかけてタイムラプスを安定化させるために「インタレスト・ポイント使用して登録を」をクリックして選択します。
- では「基本登録パラメータウィンドウ登録のタイプごとに1つの基準時点に対するマッチ(なしグローバル最適化)を選択します」。柯EP tは 、彼の他の設定個々の時点の登録と同じ。
- 次には窓、時刻を選択する基準は、時間経過の途中で、典型的には、時刻として使用されます。各時点内の個々のビューがすでに互いの上に登録されているので、ボックスにチェックを入れ、「剛性単位として各時点を考えます」。
- 「ショーの時系列統計」のボックスにチェックを入れ。その他の登録パラメータが正則含む以前のように残ります。 [OK]を押します。
注:ログウィンドウでは、同じ出力は、個々の時点登録のように表示されます。個々の時点の登録が成功し、堅調に推移しました場合は、RANSACは今99から100までパーセントであり、平均値、最小値と最大誤差は、通常、1ピクセル以下です。先に進む前に、この登録を保存します。
7.マルチビューフュージョン
注:登録手順から得られた変換は、複数のビューのうち融合等方性スタックを計算するために使用されます。このスタックヘクタールZ間隔は現在XYにおける元の画素サイズと同じであるため、元のデータと比較してZスライスの数を増加させました。■。融合は、コンテンツベースのマルチビュー融合21,31または両方マルチビュー再構成アプリケーションに実装されているベイジアンベースのマルチビューデコンボリューション31のいずれかによって行うことができます。
- バウンディングボックス
注:フュージョンは計算コストの高いプロセスである( 表1参照)、このように境界ボックスを定義することによって、データ量を削減することはかなりの処理速度を増大させます。- 全ての時点を選択し、右クリックして、「バウンディングボックス」を定義を選択します。 「BigDataViewerで定義 'を使用し、バウンディングボックスの名前を選択します。
- 関心領域を決定した後、「OK」を押して、すべての軸に「分」と「最大」のためのスライダーを移動します。バウンディングボックスのパラメータが表示されます。
注:バウンディングボックスは内のすべてのものが含まれています透明マゼンタ層に重ねられる緑色のボックス。
- コンテンツベースマルチビューフュージョン
注:コンテンツベースのマルチビューの融合21の代わりに、単純な平均を使用してのより良い画質のために、より高い重みを考慮にスタックの上の画質の差(Zにおける信号のすなわち劣化)を取り、適用されます。- 「ViewSetupエクスプローラー」に融合されるべき時点(複数可)を選択し、右クリックして、「イメージフュージョン/デコンボリューション」を選択します。
- 画像フュージョンウィンドウで、ドロップダウンメニューから「加重平均融合」を選択し、[選択した境界ボックス用に事前定義された境界ボックスを使用してください」。融合された画像の出力のために選択し、すでにHDF5既存のファイルに新しいHDF5ファイルを書き込み、一緒に登録された融合していない景色と融合された画像を使用することを可能にする、「現在のXMLプロジェクトに追加します」。押して「OK」。
- そして、「事前定義バウンディングボックス&#で39;以前に定義されたバウンディングボックスの名前を選択し、ウィンドウをポップアップし、「OK」を押します。
- 次のウィンドウでバウンディングボックスのパラメータを確認します。迅速な融合のために、融合したデータセットにダウンサンプリングを適用します。
注:融合スタックが一定のサイズを超えている場合、プログラムはより多くのメモリ効率的な「ImageLib2のcontaine'rを使用することをお勧めします。より大きなデータの処理を可能にする」PlanarImg(大画像、表示しやすい)またはCellImg(大きな画像)」コンテナーへの「ArrayImg」から切り替えます。それ以外の場合は事前に定義された設定を使用すると「混合およびコンテンツベースの融合」を適用します。 「OK」を押して進んでください。 - 次のウィンドウにHDF5の設定を確認します。事前定義されたパラメータを使用して、融合プロセスを開始します。フィジーに十分なメモリが割り当てられていることを確認します [編集]> [オプション]>メモリ&スレッド。
- マルチビューデコンボリューション
注:マルチビューデコンボリューション31は anotです多眼融合の彼女のタイプ。ここにさらに融合する、画像システムのPSFは、信号の画像および収量高解像度とコントラストをデコンボリューションするために考慮される( 図2C-J、 図5及び映画3に比較して)。- デコンボリューションする時点(複数可)を選択し、右クリックを押して「画像融合/デコンボリューション」。
- 「マルチビューデコンボリューション」と使用」事前定義されたバウンディングボックスを、現在のXMLプロジェクトに追加」を選択します。バウンディングボックスを選択して続行します。
注:デコンボリューションのための事前定義されたパラメータは、良い出発点です。最初の試用'20の反復のために、デバッグモード ''と使用してデコンボリューションの影響を評価します」。最後に、512×512×512ブロック内への計算を設定します。 - 次のウィンドウでは、事前定義された設定を使用します。すべての5 '信奉結果を表示するには、「デバッグモード」を設定します配給」。
注:デコンボリューションの出力しかしBigDataViewerは現在、16ビットのデータをサポートする、32ビットのデータです。既存のHDF5データセットにデコンボリューションの出力を追加するためには、16ビットに変換する必要があります。 - 変換のために、実行 '(時間をかけて強度を飽和かもしれません)最初の画像の最小/最大を使用してください」。
注:デコンボリューションは、その後、画像をロードし、デコンボリューションのためにそれらを準備することによって開始されます。
- BigDataServer
- 非常に大規模なXML / HDF5データセットを共有するために33 BigDataServer httpサーバを使用します。 http://fiji.sc/BigDataServerで発見することができますどのようにセットアップすると、サーバーへの接続をご紹介。
- 既存のBigDataServerオープンフィジー>プラグイン> BigDataViewer> BrowseBigDataServerに接続します。
- ウィンドウにポートを含むURLを入力します。
注:この公報に記載の映画はこの広告を経由してアクセスできますドレス:http://opticcup.mpi-cbg.de:8085 - 利用可能な映画を選択することができますウィンドウを観察します。ダブルBigDataViewerウィンドウを開き、前述のようにデータを表示します。
補足:開始する前に必要とされる材料のチェックリスト
- ゼブラフィッシュの胚/蛍光タンパク質を発現する幼虫(メチレンブルーなしゼブラフィッシュE3培地中で胚を保管してください。ステージのために24時間以上経過は最終濃度0.2mMにPTUを追加することにより、色素沈着を打ち消します。)
- 蛍光ステレオスコープ
- 毛細血管(ブラックマーク付き20μl容量)および適切なプランジャー(毛細血管を再使用しないでください。プランジャは、他の一方で、いくつかの実験のために再利用することができます。)
- 1.5ミリリットルプラスチックチューブ
- 鋭いピンセット
- ガラス(火研磨)やプラスチックピペット(プラスチックは24時間以前の胚のために使用することができます)
- ガラスやプラスチック皿、直径60ミリメートル(プラスチック製とすることができます24時間以前の胚のために使用されます)
- 2 100ミリリットルビーカー
- 輸液150センチ延長ホースで50mlルアーロックシリンジ(ホースとシリンジは、微生物による汚染を避けるために、実験の間に完全に乾燥した状態で保管する必要があります。)
- プラスティシーン
- 低融点(LMP)アガロース
- E3培地(5のNaCl、0.17のKCl、0.33mMのCaCl 2を、0.33 mMのMgSO 4)し
- MS-222(トリカイン)
- フェニルチオ尿素(PTU)
- (ここで、ビーズとも呼ばれる)、蛍光ミクロスフェア
- 二重蒸留H 2 O(のddH 2 O)
結果
LSFMはスケール全体に発達過程を画像化するための理想的な方法です。いくつかのアプリケーションは、細胞内の構造だけでなく、細胞全体の組織の短いと両方の長期的なイメージングを示すここにコンパイルされます。これらの例はまた、LSFMは、網膜の神経発生に眼杯形成から眼の発達の様々な段階で有用なツールであることを示している。 映画の1が最初に無傷の胚のズームアウトビューを示す、一般的なLSFMアプローチの実例として働きます明視野でのアガロースシリンダーを埋め込み、それ以降の蛍光チャネルにおける網膜の詳細図を示します。

ムービー1:ゼブラフィッシュ網膜のLSFM LSFMのアプローチを説明するためには、この映画は、胚がAの内部にそのまま画像化される明視野で最初示しています蛍光に切り替える前に、シリンダをgarose。後でビューの大きなフィールドが全体の網膜の観察を可能にすることが示されています。次に、映画は良い細胞下解像度を強調するために、網膜の拡大小さな領域を示しています。 Ath5:ギャップ-GFP 37トランスジェニックゼブラフィッシュ胚は、イメージングのために使用しました。この導入遺伝子は、網膜(主に神経節細胞と感光体前駆体)内の異なるニューロンを標識します。映画の蛍光部分は、5分間隔で40X / 1.0 Wの対物レンズを用いて、両面照明で記録単一のビューとして捕獲されました。厚さ30μmの体積の最大値投影が示されている。 このファイルをダウンロードするにはこちらをクリックしてください。
ムービー2は、非常に高速な細胞内事象を高解像度で撮像することができるか、を示しています。この場合における微小管の成長そのプラスは、網膜神経前駆細胞で終わります。映画の中で含まれている情報は、微小管プラス端の成長の追跡および定量を可能にします。

ムービー2:単一細胞における微小管ダイナミクスこの映画は、プラスの先端マーカータンパク質EB3-GFP 38によって標識微小管の成長にプラスのヒントをキャプチャします。タンパク質は、単一の網膜前駆細胞で発現されます。微小管は頂端から基底(下へ)の方向に優先的に成長しています。 EB3彗星の平均速度は0.28±0.05ミクロン/秒と測定されました。高微小管の核形成活性を示す細胞の頂端側の明るいスポットは中心体です。 EB3-GFPプラスミドDNA:野生型胚は、HSP70を注射しました。映画は、約28 hrpost fertilizでヒートショック(37℃で15分間)した後、4時間を取得しましたエーション63X / 1.0 W客観的かつ1秒の時間間隔を使用して片面照明で記録単一のビューとして(HPF)。単セルは、網膜の大部分をカバーする視野からトリミングされました。 2つのスライスの最大値投影が示されている。 このファイルをダウンロードするにはこちらをクリックしてください。
図3に示すように、どのように細胞内構造は、多くの時間にわたって追跡することができます。ここでは、網膜神経節細胞(RGC)トランスロケーション内の中心体が捕捉されます。

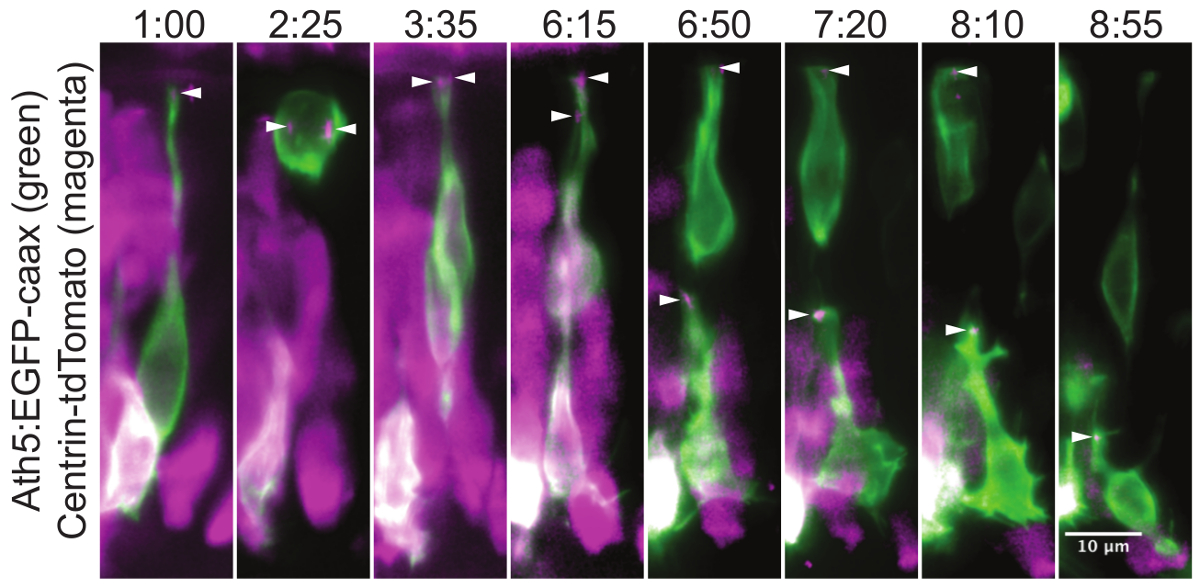
図3:網膜神経節細胞の転座の間に中心体の局在タイムラプス実験のこのモンタージュは、網膜神経節細胞(RGC)の成熟を 通して中心体の位置を示しています。神経前駆細胞では中心体は、頂端処理(1時)の非常に先端にローカライズされています。細胞分裂時には、2の中心体は紡錘体(2時25分)のための極として機能します。この部門は、RGCおよび光受容細胞の前駆体となり、第二の娘細胞に分化1の娘細胞になります。頂端処理が頂端側に取り付けられたまま分裂した後、RGCの細胞体は、網膜の基底側に移動します。 RGCは、基底側に達すると、その頂端処理は切り離し、中心体は(6時15分)で移動します。徐々に頂端処理(午前6時50分、7時20分、8時10分)と一緒に後退しながら、中心体に従うことができます。中心体がまだ頂端局在している間に、最後のフレームでは(8:55)神経節細胞は、その基底側から軸索を成長しています。モザイク発現は、1つの細胞段階での野生型胚へのプラスミドDNAの注射によって達成しました。 GFP-CAAX(緑)のconst:細胞がAth5によって可視化されていますRGCおよび他のニューロンを標識ruct、。中心体(矢頭)はCentrin-tdTomato 29式(マゼンタ)によって標識されています。網膜の頂端側には、下部の画像および基底側の最上部にあります。厚さ30μmの体積の最大値投影を示しています。画像は全体の網膜をカバーする映画のクロップました。映画は約34時間後に受精(HPF)で始まります。 Zスタックは、40X / 1.0 Wの目的で、5分ごとに取得しました。ミリメートル:時間はHHに示されています。スケールバーは10μmで表す。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
図4に、それが表示され、そのような映画1のように全組織を捕捉したデータから抽出することができる方法を、単一細胞の挙動。RGCの移行を容易に追跡し、その頂端および基底処理することができesが続きます。

図4: 図3で説明したように、単一の網膜神経節細胞の転網膜の基底側への頂端からRGC転端末有糸分裂後に発生するRGCがAth5の発現によって標識されています。ギャップ-GFP 37導入遺伝子。厚さ30μmの体積の最大値投影を示しています。画像は全体の網膜をカバーする映画のクロップました。映画は約34 HPFで始まります。 Zスタックは、40X / 1.0 Wの目的で、5分ごとに取得しました。ミリメートル:時間はHHに示されています。スケールバーは5μmで表す。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
図5は、組織の皮下をキャプチャするマルチビューLSFMの能力を実証します光学小胞は、眼杯に変身する時に眼杯の形態形成の例にセルラー解像度、とエール形態形成のプロセス。最終的な画像は、等方性分解能でつのZスタックに(この場合)5つの異なるビューからの情報から合成されたときに画質が大幅に、多視点画像化することによって改善することができます。この図は、マルチビューデコンボリューション後の加重平均融合および画像コントラストのさらなる利得および解像度後の画質の改善を示しています。図は、データセットを介して異なる向きに2つの光のスライスを示しています。また、逆畳み込みデータセットからのモンタージュは、時間をかけて眼杯の形態形成を示しています。全ての時点は、次にムービー3に示されています。

図5:単一のビューとmultivの2つの方法の間の画質の比較IEW融合。(A)側面図及び(B)背側のビューから示された単一のビューデータの1光学スライス。ストライプのアーティファクトと試料内部のより深い信号の劣化は明らかです。また、融合したデータ(CF)で可視画像の一部は、この特定のビューでキャプチャされませんでした。 (C)同じ光学スライス、今多視点融合したデータは、側面図および(D)から示されるように背ビュー。ストライプアーチファクトが抑制され、サンプル中の深部構造をよりよく解決されることに注意してください。 (E)と同じ光学スライス今マルチビューは、側面図及び(F)背ビューから表示されるデータをデコンボリューションとして。個々の細胞膜と核が十分に区別することができるように増加し、コントラストや解像度に注意してください。解像度は、サンプルの内部、特に深い劣化しません。データセットは、約20度離れて5ビューから両面照明で取得されました。 ABのzスタック1.5μmのステップサイズで100μmのは、20X / 1.0 Wの目的と10時間10分間隔で各ビューで取得した外。マルチビュー融合およびデコンボリューションのための入力画像がダウンし、画像処理を高速化するために2回サンプリングしました。マルチビューデコンボリューションの15回の反復が実行されました。 (G)モンタージュは、眼杯のステージへ眼胞からの早期眼の開発中に形態形成のイベントを強調するための逆畳み込みデータから背ビューのトリミングされた領域を示しています。最初に同様の円柱上皮である光学小胞の二層は、異なる細胞集団に分化します。表皮に近い遠位層は、網膜神経上皮(RN)になると、神経管に近い近位層は、網膜色素上皮(RP)となります。 RNの細長いと陥入した中の細胞は眼杯(5時00分に1:40)を形成します。同時にRP細胞は平ら。表面外胚葉はWH、レンズ(1:40)を形成するように誘導されますichの陥入以降(午前5時00分)。映画は17 HPFで始まります。ミリメートル:時間はHHに示されています。すべての細胞膜は、βアクチンにより標識された:RAS-GFP導入遺伝子とすべての核は、HSP70によって標識されている:H2B-RFP導入遺伝子。スケールバーは30μmで表します。 FBの前脳、LEレンズ、OP嗅板、RN網膜神経上皮、RP網膜色素上皮。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

映画3:眼杯の形態形成は、単一のビューとマルチビュー融合の二つの方法で示したタイムラプスムービーは眼杯の舞台に眼胞から眼杯の形態形成の完全なプロセスを示しています。これは、側面図(上)と背側のビューから単一の光のスライス(下)から単一の光のスライスを示しています。開発眼胞の細胞は、最終的に内網膜神経上皮と外側の網膜色素上皮と半球状の光学カップを形成するために複雑な再配列を受けます。レンズは、表面外胚葉から形成されます。これは、神経上皮と一緒に陥入し、眼杯に座っています。すべての細胞膜は、βアクチンの発現により標識された:RAS-GFP(緑色)導入遺伝子および核はHSP70で標識されていますH2B-RFP(マゼンタ)。映画は約17 HPFで始まります。データセットは、約20度離れて5ビューから両面照明で取得し、約100μmのaからzスタックは、20X / 1.0 Wの目標に10分毎に取得しました。ミリメートル:時間はHHに示されています。スケールバーは50μmで表します。 このファイルをダウンロードするにはこちらをクリックしてください。
ディスカッション
データ収集のための1.重要なステップとトラブルシューティング
GFPおよびサンプルを表現するRFPのための典型的な画像設定は、シリンドリカルレンズによって形成された光シートは静的で説明した顕微鏡のセットアップ、 表3に記載されています。 2つの照明目的は、空気レンズであり、検出目的は、水浸漬レンズです。 20X / 1.0または40X / 1.0目標とズーム1.0は、230 nmおよび115 nmのピクセルサイズは、それぞれ441 X 441ミクロンまたは221 X 221ミクロンの視野を提供します。 2:国境比1に中心を持つデフォルトの光の板厚を使用することをお勧めします。 20X / 1.0の場合、この厚さは4.5μmとし、40X / 1.0中心部のμmの3.2にするために相当します。イメージング速度は、一次優先されていない場合、チャンネル間の蛍光発光のクロストークを回避するために、マルチサンプルの場合には別個のトラックを使用します。買収の最高速度は、以下によってZステップごとに50ミリ秒に制限されていますZ-ドライバの移動速度。目標は、例えば 、の場合には、最大画像形成速度を達成することである場合、Zのステップごとに撮影した全ての画像の合計が50ミリ秒未満であるように、両面照明を有する2つのトラックは、露光時間を設定しなければなりません。単一の画像をz工程毎に取得された場合に、50ミリ秒よりも短い露光時間を設定することは有益ではありません。
| 1920 x 1920画像サイズ |
| 16ビットの |
| ピボットは、上のスキャン |
| オンライン融合で両面照明 |
| 10X / 0.2照明目的 |
| 20X / 1.0 Wプランアポクロマート検出目的 |
| トラック1:100mWのレーザーの励起は488nmで、典型的には2%、550 nmのSP発光フィルター |
| トラック2:励起561 nmの75 mWのレーザーの一般的に3%、585nmのLP発光フィルター |
| 100ミリ秒の露光時間まで |
| Zスタックの厚さ50〜100μmで |
| 連続Z駆動モードで1~1.5ミクロンのzのステップサイズ |
| 28.5°Cでのインキュベーション |
表3:イメージングの設定。
実験後の試料を検査します
試料は、実験の終了時に、まだ健康であることを確認することが重要です。第1の読み出しとして、ステレオスコープの下で試料のハートビートを確認してください。鋭いピンセットでサンプルをアガロースから取り出すことができる、それがイメージングによって影響を受けたかどうかを確認するために、更なる開発をインキュベーターに移動しました。あるいは、抗体染色のために固定することができます。
マウントとドリフト
チャンバsの浸透圧を維持することが不可欠です近くアガロース包埋の浸透圧にolution、アガロースおよびサンプルのその後の不安定性のそうでなければ、膨潤/収縮が発生します。従って、チャンバを充填するために、同じ溶液(メチレンブルーなしE3媒体)を使用し、1%低融点アガロースのアリコートを調製します。それは、そのゲル化特性を失うことができるように加えて、以上2時間70℃の加熱ブロック内のアガロースを放置しないでください。
これは胚の熱ショック応答または死亡につながることができますように、あまりにも熱いアガロースに魚を埋め込まないでください。胚上の温かいアガロースの効果がわからない場合は、尾が曲がらないことと心拍数が遅くなることはありませんことを確認してください。この問題が発生した場合、実験のために別の胚を使用しています。
サンプル短い(2周りセンチ)でアガロースカラムの全体の長さを保持し、プランジャの先端に向けられ、その頭とゼブラフィッシュをマウントします。同様に、アガロースシリンダーはcapillから押し出し進はできるだけ短くする必要があります。これらの措置は、映画全体のサンプルの安定性を確保します。同時に、アガロースカラムには、これは、主要な屈折や反射を引き起こすように、ガラスキャピラリー自体は、光路に到達しないように、十分に長くなければなりません。
試料の初期ドリフトをアガロースシリンダー自体の体積変化によって引き起こされます。プランジャーの摺動するとの理由ではありません。したがって、それは粘土やマニキュアでプランジャーを固定するためには役立ちません。胚は、あまりにもその自然な成長のために、ムービー中の位置を変更することがあります。したがって、視野の中央に関心領域を中心とこれらの動きに対応するために、縁でいくつかの部屋を維持することをお勧めします。
光路中の包埋剤の量の減少
サンプルを正しく配向することは可能な限り最高の画質15を達成するのに役立ちます。遺伝子ラリーは、励起および発光光ができるだけ少ない組織と取り付けメディアを通過する必要があります。最適解は、アガロースフリー実装です。これは、メインルートがphytagelに装着され、側根がその後完全にゲルカラムの外に成長してみましょうされたシロイヌナズナ側根イメージング14、のセットアップ、例えば達成されました。アガロースフリー実装はまた、2日間12にわたってモドキTriboliumカブトムシの完全な胚のイメージングのために開発されました。画質改善は、その場合の主な動機はありませんでした。モドキTribolium胚は、単に十分な長アガロース内部に生存しません。絶対に埋め込みメディアフリー実装は、ゼブラフィッシュにおける長期的なイメージングのために達成されていません。それでも、我々はアガロース固化、ほとんどの胚は、アガロースの深い位置する1つ目でキャピラリー内で斜めに配置され、第二の目は埋め込み柱の表面に近いされているという事実を利用することができます。 T表面に近い彼の目は、優れた画像品質を提供し、したがって、優先的に画像化されるべきです。
アガロース濃度と長期的なイメージング
実装用アガロースの濃度は、試料の安定性と、それに酸素の胚の成長および拡散に対応する可能性の間の妥協です。 1%よりも高いアガロース濃度を用いて、サンプルの安定性には追加の利得はありません。実験を最適化するための出発点として、我々はまた、1%アガロースに装着するにはあまりにもデリケートで24 HPFより若い胚に適しており、0.6%アガロースを、お勧めします。古い胚および幼虫anaesthetizeするために、MS-222濃度は、副作用13なしでは200μg/ mlまで上昇させることができます。
それは胚とコーの成長を制限するため、場合に発生中の胚は、取り付けアガロースは推奨されない、より長い±12時間結像され、エステール変形。この問題は、水13,39と同様の屈折率を有するFEPポリマー管に胚を装着することによって、ゼブラフィッシュのために解決しました。マウス胚は、一方で、中空アガロースシリンダー40または注射器41に取り付けられたアクリルロッドの穴に固定化することができます。チューブの壁はアガロースよりわずかに光を屈折するのでFEPチューブの装着は、しかし、デフォルトの方法として推奨されていません。
光シート整合
良好な画質のためには、すべての実験の前に自動調光シートの位置合わせを行うことが重要です。ズーム設定を変更した場合は特に、目的は、取り出し、または別の液体がチャンバ内で使用されました。
イルミネーション
光シートのピボットスキャンは常にアクティブにする必要があります。大きな散乱標本の場合は、オンラインfに両面照明を適用する必要があります視野全体に均一な照明を達成するためにusion。両面照明はまた、胚のレンズにより、入射光シートの屈折であるアイ・イメージング、特定の問題を減少させます。より小さい、より少ない散乱標本を効率半分撮像時間を短縮し、両面照明に比べわずかに良好な画像品質をもたらすことができる片面照明を使用して画像化することができます。 2つの照明アームのための光路が常に異なっていて、より効率的な一つが選択されることができるからです。さらに、それぞれの側から来る二つの光シートは、融合後軽度のぼけが生じる一つの面で完璧になることはありません。左右からの照明で画像がモーションブラーにつながる可能性があり、順次取得されているので、成長している微小管( 動画2)のような非常に高速な細胞内事象については、両面照明は、適切ではありません。
光退色そして、光毒性
少ない蛍光体光退色はしばしばLSFMの主な利点として挙げられています。我々は目的が全く光退色あってはならないことを主張するだろう。ライブイメージング実験に顕著な光退色がある場合、試料は、許容レーザー露光のその生理的範囲外におそらく既にあります。スピニングディスク顕微鏡におけるゼブラフィッシュの胚を撮影するときは、私たちの経験では、高い光毒性が顕著であっても、蛍光シグナルの漂白剤の前に胚発生を停止することができます。ほとんどまたは全く光退色が観察されるように、したがって、LSFMの撮像設定を調整すべきです。 LSFMを試料に穏やかであっても、その後のデータ分析のための十分な信号対雑音比を達成するのに必要なだけのような多くのレーザパワーと露光時間を使用することが賢明です。
Zスタック、時間間隔とデータサイズ
LSFMによって生成されたファイルは、通常は非常に大きいです;時にはテラバイトの範囲です。画質とデータサイズとの間の妥協をすることがしばしば必要です。これは、特にタイムラプス買収におけるスタックと間隔のz間隔の場合です。 zの間隔を定義するには、Z-スタックツール]タブで最適なボタンは、理想的には、データセットが、後にデコンボリューションされる場合は特に、使用されるべきです。これは、隣接する光学スライス間の50%のオーバーラップを達成するために、間隔を算出します。それでも、いくらか大きいzの間隔は、通常は許容されます。それらは、zスタック、ならびに最終的なファイルサイズを取得するために必要な時間を削減します。最適な時間サンプリングは、目的のプロセスに依存します。全体的な眼の開発のために5〜10分間隔は、通常は許容されます。いくつかの構造が自動的に追跡する場合は、その後の時間点は十分に類似していなければなりません。
蛍光ビーズ
蛍光ビーズは、主に異なるビューを登録するための基準マーカーとして役立ちます互いの上にマルチビューデータセットの。必ず使用前に完全にビーズソリューションをボルテックス。これは、蛍光色素の損失につながることができますようにビーズを加熱しないでください。マルチビューの登録のための最適なビーズ濃度は、実験的に決定されなければなりません。説明プラグインは、各ビューを通じて約1,000検出ビーズに最適です。より大きな(500nm以下千nm)のビーズが小さい(500nm未満)のビーズよりもロバストに検出されました。より大きなビーズは明るく、試料中の構造の誤認検出することなく、セグメントに容易であるためです。より大きなビーズの欠点は、それらが最終的な融合と復元画像に非常に顕著であるということです。それぞれの新しい蛍光マーカーについては、適当なビーズのサイズおよび蛍光発光を最適化しなければなりません。 図5 ムービー3からの試料の例を与えるために、100nmの緑色発光ビーズは膜-GFPチャンネルにあまりにも多くの偽陽性検出を与えたが、千のnM赤色発光ビーズを確実に試料内部の非常に少ない正検出とH2B-RFPチャネルで検出されました。ビーズの検出は、蛍光マーカーを有するチャネルで失敗した場合、唯一のビーズを含有する別個のチャネルを取得することができるが、これは非常に実用的ではありません。サブ解像度サイズのビーズは、デコンボリューション( 図2C-D)のために使用することができる顕微鏡の点広がり関数(PSF)の直接読み出しを与えます。登録と融合が大きなビーズ( 例えば 1000nmで)とのより良い動作する場合、PSFの別の画像は、サブ解像度、 例えば 、100nmのビーズで取得することができます。多色のビーズを使用すると、マルチチャンネル取得の登録時に、完全にそのチャンネルのオーバーレイを検証するために有用です。
後続のマルチビューの登録と融合することなく、単一のビューから撮影する場合、蛍光ビーズの添加は必要ありません。しかし、それらの例にはビーズがイニシア時に役立つことがありますリットル光シート調整は、光シートの品質をチェックするため、一般的に光学収差を明らかにします。このような光学収差が損傷や汚れの目的、アガロースでチャンバーまたは不均一性の汚れた窓のような様々なソースから生じ得ます。ビーズはまた、多視点登録フィジープラグイン20によってドリフト補正のために使用することができます。
マルチビュー
マルチビュー再構成目的のためには、互いに対向されていないビュー3、というように5と、奇数番号を取得した方が良いです。 PSFは異なる方向から撮像されているので、これはデコンボリューションを向上させます。ビューの間で十分な重なりがあること時間経過取得の開始時に確認することも重要です。これが最良の直後に最初の時点でのビューが正常に登録することができることを確認し、 すなわち 、経験的に行われています。マルチビュー取得の目的は、分解能を増加させることである場合大きな散乱標本の画像の、各ビューで試料全体の画像にはお勧めできませんが、信号が劣化試料の中心を停止します。試料の後半から低品質の取得は、マルチビューの再構築に有用な情報を追加しないでしょう。
データ処理のための2.重要なステップとトラブルシューティング
現在、十分に文書化および採用することが比較的容易されている光シート顕微鏡から多視点データを処理するためのいくつかの可能性が存在します。私たちは、フィジー32(ステファンPreibisch未発表で実装されたオープンソースソフトウェアであるマルチビュー再生アプリケーション、使用するリンク部1aとLink1b材料の一覧でを)。このプラグインは、以前のSPIM登録プラグイン20の主要な再設計され、レビューBigDataViewerとそのXMLとSPIM登録ワークフローとHDF5フォーマット33( 図1B、統合Schmied ら 42によるリンク2 、 リンク3 )。この出願はまた、かなりの処理43を高速高性能コンピューティングクラスタのために適合させることができます。このマルチビュー登録申請が活発にさらに開発され、改良し続けます。問題や記載されているソフトウェアの要求を特徴とする場合には、(それぞれのGitHubのページに問題を提出してくださいリンク4マルチビュー復興のためのリンク5 BigDataViewer用)。
2番目のオプションは、顕微鏡と一緒に利用できる商用ソフトウェアを使用することです。このソリューションは、うまく機能し、採用します異なるビューを登録するために蛍光ビーズを使用して同じ原理。しかし、それは速いBigDataViewerと同じようにデータセット全体を可視化するためのオプションを欠いています。ソフトウェアのための追加ライセンスを購入しない限り、また、ソフトウェアは、他のユーザーのための顕微鏡をクラスタに適合し、さらに処理ブロックすることはできません。
また、オープンソースソフトウェアである第三の選択肢は、最近ケラーラボ44から発行され、処理および光シートデータの下流の分析のための包括的なフレームワークを提供しました。このソフトウェアは、従って、試料の周りの蛍光ビーズの存在を必要としない、マルチビュー融合を行うために、サンプル内の情報を使用しています。しかし、同時に、撮像ビュー(目標)の直交方向を想定し、それは、任意の角度44から取得したデータのために使用することができません。
ハードウェア要件
ntent ">処理のために使用されるハードウェアは、画像の取得がその後の分析よりも高速である。先に実際の実験の十分なストレージ容量と使用可能なデータ処理のための明確なパイプラインを、が存在しなければならない。 表4に見ることができる、それが簡単です。未処理のデータが殺到取得する。融合されたビュー、最大強度の突起又は球状の突起45のようにすべての生のイメージではなく、トリミングされたバージョンまたは処理された画像を保存することがしばしば非現実的です。| プロセッサ | 二つのインテル®Xeon®プロセッサーE5-2630(6つのコア、2.30 GHzのターボ、15メガバイト、7.2 GT /秒) |
| メモリ | 128ギガバイト(16×8ギガバイト)1600 MHzのDDR3 ECC RDIMM |
| ハードドライブ | 4×4 TB 3.5インチのシリアルATA(7.200 rpm)でハードドライブ |
| HDDコントローラ | PERC H310 SATA / SASコンDell Precision用コントローラ |
| HDDの構成 | C1 SATA 3.5インチ、1-4ハードドライブ |
| グラフィック | デュアル2ギガバイトのNVIDIAのQuadro 4000(2カードワット/ 2 DP&1 DVI-I各)(2 DP-DVI&2 DVI-VGAアダプタ)(MRGA17H) |
| ネットワーク | インテルX520-T2デュアルポート10ギガビットイーサネットネットワークインターフェイスカード |
表4:ハードウェア要件。
データ処理速度
データ処理に必要な時間は、データの寸法にし、使用するハードウェアに依存します。 表1に、我々は4ビューと2チャンネルを1時点から成っ例8.6ギガバイトのマルチビューデータセットを処理する際の重要なステップに必要な時間の概要を説明します。
| 処理■TEP | 時間 | プロトコルステップ |
| HDF5として再度保存 | 6分30秒 | 6.3 |
| 関心点を検出 | 20秒 | 6.4 |
| 興味のポイントを使用して登録します | 3秒 | 6.5 |
| コンテンツベースのマルチビュー融合 | 4時間 | 7.2 |
| マルチビューデコンボリューション(CPU) | 8時間 | 7.3 |
| マルチビューデコンボリューション(GPU) | 2時間 | 7.3 |
表1:データ処理時間。
マルチビュー再構成のための入力データ・フォーマット
フィジープラグインマルチビュー復興は .czi、tifファイルとome.tiffフォーマットをサポートすることができます。 .czi形式のデータ構造に、不連続なデータセットではありません前処理なしでサポート。不連続は、( 例えば 、試料のドリフトによる位置の再調整が)記録を再起動する必要がありましたことを意味します。この場合、.cziファイルは、.tifファイルとして保存する必要があり。 tifファイルの場合は、各ビューをファイルし、照明方向は、別々のファイルとして保存する必要があります。
ピクセルサイズのキャリブレーション
顕微鏡オペレーティング・ソフトウェアは、選択された目的に基づいて、X-Yピクセルサイズのキャリブレーションを算出します。しかし、Zの画素サイズはステップサイズで独立して定義されます。間違った目的は、ソフトウェアで指定されている場合は、z比にxyが間違っていると、登録は失敗します。
新規登録
データセットを定義した後で登録数が1になり、関心点の数がViewSetup Explorerで0になります。初期登録は、データセットのキャリブレーションを表します。数oを両方F登録や関心点は、処理中に増加します。
関心点の検出のためのダウンサンプリング
ファイルの読み込みと分割がはるかに高速になりますので、ダウンサンプリングを使用することは、推奨されます。検出パラメータは、このように異なるダウンサンプル設定間検出の設定を転送し、ダウンサンプリングに応じて変更可能ではないであろうことに注意することが重要です。
関心点の検出
彼らはかなり登録を妨げないので、それは、さらにいくつかの偽陽性検出を取得する価格で、各ビューにできるだけ多くの真のビーズのようにセグメント化することをお勧めします。スプリアス検出は、数字にはいくつかの場合には、(関心点の登録を参照してください)登録時に除外されます。しかし、大規模な偽陽性の検出は、アルゴリズムに問題を提起します。それだけでなく、検出のためのパフォーマンスを低下させ、登録、それはビュー間でこれらのビーズを比較するだけでなく、セグメントにはるかに長いイメージがかかりますが、それはまた、登録の精度を低下させるからです。これは、より厳格な検出パラメータを使用することによって対処することができます。さらに、ビーズ(1ビーズ図1Eにすなわち複数の検出)のセグメンテーションを介して登録に有害であると避けるべきです。
関心点の登録
お互いに意見を登録するには、各ビューの各ビーズの位置は、その3、最も近い隣のビーズに関して、その位置によって記述されています。これらの星座は、地元の幾何学的記述子を形成し、ビュー間、各ビーズを比較できるようにしています。 2ビューの間のマッチング記述子を有するビーズは、その後、候補の対応として考えられています。これが唯一の局所記述は、典型的には、各ビーズのためのユニークであるためにランダムに分布ビーズ、のために働くことに注意してください。一つは、このような登録のために、試料中の核として他の構造を使用することができます。しかし、サンプル中の非ランダムに分布している核を検出するために、他の方法は、20,21が適用されます。
候補対応はその後、偽陽性を排除するためにRANSAC 36に対してテストされています。それぞれの対応がお互いに意見を重ねるための変換モデルを提案しています。外れ値が異なるものに各ポイントを希望のに対し、真の対応は、おそらく、1変換モデルに同意するだろう。真の対応関係は、2つの比較ビュー間のアフィン変換モデルを計算するために使用されます。 反復最適化アルゴリズムと大域的最適化は 、すべてのビューがビュー20,21の間の最小変位を目的とした最初のビューに登録されている間に、行われます。
タイムラプス登録
移動のために顕微鏡ステージのアガロースおよび不正確なモータ運動のメンターは、各スタックの位置は、時間をかけて適度に変化します。個々の時点の登録は、この時点の見解の違いを削除するのに対し、タイムラプスも、全体として登録する必要があります。この目的のために、各個々の時点を基準時点に登録されています。
基準時点
多くの時点で時系列に処理される場合、ビーズ強度が脱色による経時分解することができるので、代表的な時点は、通常、時系列の途中から基準として選択されます。この文献では、関心点検出、登録、バウンディングボックスとの融合のためのパラメータを決定することができます。これらのパラメータは、各個々の時点での特定の変換モデルを計算するために全体の時間経過に適用されます。タイムラプス登録時に、他のすべての時間の見所にはレジストされていますこの基準時点に空間的にered。したがって全体の記録のために境界ボックスパラメータは、この特定の時点に依存しています。
マルチチャンネル登録
複数のチャンネルを画像化すると、理想的には同じ蛍光ビーズは、全ての画像形成されたチャネルで表示されるはずです。検出および登録は、アカウントへの変換に異なる光波長の影響をとる、個別に各チャンネルに対して行うことができます。ビーズはすべてのチャンネルには表示されませんまたはビーズは、1チャンネルでも多くの画像を支配していると、他のチャネル(複数可)で暗すぎているため、多くの場合、これは、不可能です。典型的なソリューションは、検出および登録し、取得した変換モデルのための1つのチャンネルに見えるビーズを使用することである( すなわち 、検出、登録、タイムラプス登録後)その後、 フィジー>プラグイン>マルチビューReconstrにより他のチャネルに適用されますuction>バッチ処理]> [ツール]> [重複変換。 他のチャンネルに選択1チャネル の変換を適用するためのドロップダウンメニューで。次のウィンドウでXMLを選択し、[OK]をクリックします。そして、 ソースチャネルとしてビーズが含まれており、 ターゲットチャネルとしてビーズなしのチャネル(複数可)を選択チャンネルを選択します。 重複のための変換は、 すべての変換 、[OK]を押しますを 交換 して使用します 。変換は、他のすべてのチャンネルにコピーされ、XMLで保存されます。ViewSetup Explorerで新しい変換を確認するには、 マルチビュー復興アプリケーションを再起動します。
バウンディングボックス
複数のビューの融合は非常に計算集約的です。しかし、大きな画像は、典型的には、サンプルだけでなく、その周囲のビーズだけでなく、に対応するために取得されます。登録パラメータいったんsは、ビーズから抽出され、それらはもはや画像の一部として有用ではありません。したがって、融合効率を高めるために、試料を含む画像スタックの一部のみを一緒に融合されるべきです。関心(バウンディングボックス)の領域は、サンプルを含むように定義され、可能な限り周囲のアガロースのわずかする必要があります。境界ボックスとボリュームが1634×1746×1632ピクセルに縮小され、メモリ要件が低減されているのに対し、 図2の例では2229×2136×2106ピクセルとボリューム全体の加重平均融 合は、RAMの38196メガバイトを必要とします17729メガバイトまで。
コンテンツベースのマルチビュー融合
多視点データを融合することで課題は、ビューは通常、突然終了し、同じボクセルに対して同じ画像品質を含んでいないということです。ビューの単純平均は、したがって、ブレンディングアーティファクト、不要な画像の劣化につながります。コンテンツベースのマルチビューのfuシオンは考慮に入れ、これらの問題の両方を取ります。まず、一つの画像が終了し、他の一つは開始し、第2に、局所的な画像品質を評価し、融合21で高画質に高い重みを適用する異なるビューを、ブレンド。単一のビューと比較すると、XYにおける信号のわずかな低下( 図2E-H、図5A-D)でのzの分解能の向上があります。
マルチビューデコンボリューション
マルチビューデコンボリューションは、景色の融合を達成するための別のアプローチです。この方法では、異なるビューのPSFは、顕微鏡の光学系によって畳み込まれた画像を回復するために考慮されます。この方法は、大幅に画像ボケを除去し、信号31( 図2C-D、図2I-J、 図5E-G、 映画3)の解像度とコントラストを増加させることによって、画像品質を向上させることができます。
ve_content ">デコンボリューションは、このように処理のためにGPUを使用すると、このプロセスの速度が向上し、( 表1を参照)非常に計算集約的である。また、ダウンサンプリングされたデータにデコンボリューションを実行する必要があります。ダウンサンプルに、 フィジー>マルチビューの復興を使用>バッチ処理]> [ツール]> [変換を適用します 。これは、.xmlのファイルに保存されますビューに新しい変換モデルを適用します。BigDataViewerとBigDataServerためのHDF5ファイル形式
BigDataViewer 33は、テラバイトサイズのデータを簡単に可視化することができます。どのようにBigDataViewerを制御するために、表2にまとめられている。プログラムの基本的な動作のスクリーンキャストは、元の出版物33におけるサプリメントとしても利用可能です。 BigDataViewerは、メタデータを含む.xmlファイル、および画像データを含むHDF5ファイルを中心とします。画像データがPRESあります3Dブロック内に複数の分解能レベルでHDF5でENT。フル解像度がロードされる前に、複数の分解能レベルは、低解像度で高速にデータを視覚化することを可能にします。必要なときに、個々のブロックは専用メモリにロードされます。このように、HDF5の画像フォーマットはBigDataViewer 33を介してデータを直接かつ高速な可視化を可能にします。ファイルのロードをより効率的に行われるので、それはまた、処理が高速化されます。それは厳密に処理するために必要ではないので、我々は、この形式にデータセットを再保存をお勧めします。データフォーマットのさらなる説明については、を参照してくださいリンク3 。また、データセットは、共同研究者やBigDataServer 33(使用して、パブリックと共有することができますリンク6 )。
| キー | 効果 |
| F1 | BigDataViewerとその基本的な操作の簡単な説明とヘルプを示しています |
| <>またはマウスホイール | Zの動き |
| 上下矢印 | ズームイン、ズームアウト |
| 右クリックしてドラッグ | ビューアでサンプルを動かします |
| 左クリックしてドラッグ | カーソルの周りにサンプルを回転させます |
| ビューアまたはTab、左または右の矢印の下にあるスライダー | 時間軸上の移動 |
| さらに、シフトを押します | 任意の軸に沿ってより速く移動や回転 |
| xと、左と右矢印 | x軸の周りを回転します |
| yおよびその後左と右矢印 | y軸を中心に回転します |
| z及びその後左と右矢印 | z軸の周りを回転します |
| シフトであり、x | x軸に沿ってビューを配向 |
| シフトとy | y軸に沿ってビューを配向 |
| シフトおよびz | z軸に沿ってビューを配向 |
| 私 | 異なる補間モード間の切り替え( すなわち最近傍とトリリニア) |
| Sまたは[設定]> [明るさとコントラスト | チャンネル、明るさとコントラストの色を変更します |
| F6または[設定]> [可視性とグループ化 | 異なるグループをオーバーレイし、数字キーを介してグループを呼び出すにグループ化できます、表示されたグループを変更します |
| F10または[ツール]> [記録映画 | 現在表示されているスライスの時系列を取得します。 |
表2:ビッグデータビューア。
LSFMの説明実装の3制限事項
低いスループット
典型的なLSFM実験では、実験ごとに1つのサンプルが撮像されます。それでも、我々の経験では、多くの有用な情報は、単一のサンプルから抽出することができます。複数の胚のハイスループットイメージングは、最近のサンプルの位置と回転の自由を犠牲にしているが、典型的には、ホーム建てLSFMのセットアップ46-48で達成されています。
深部組織への不十分な浸透
ゼブラフィッシュ胚は、半透明であっても、散乱と吸収による組織内深部を撮像すると、得られる画像の品質がすぐに悪化しています。部分的にこれはサンプルによって放出される蛍光の散乱や吸収の影響で、現在の設定で補正することはできません。不均一な画質のもう1つの原因は、不規則な照明です。 T彼光シートは、左右どちらかの側から入射し、ストライプのアーティファクトとぼかしになり、そのパスそれを屈折、内の任意のオブジェクトです。両面照明や多眼融合は、最終的な画像にアーチファクトを減少させることができます。最後に、画質が原因で、エッジに向かって厚くなってきている光シート、自然の形状に視野のマージンに向かってわずかに悪化する傾向があります。
限られた化学操作
薬剤または阻害剤の使用は、ゼブラフィッシュ研究で広まっています。薬のこの顕微鏡の使用においてによる試料室と同じ室を共有する機器の他のユーザーの考慮事項の大ボリュームに、拘束されています。薬物実験専用の余分室を使用することで、この問題を解決することができます。ガラスビーズで部分的にチャンバーを充填することは必要とされる液体の体積を減らすことができます。
いいえ写真編集ません
カレンTLYこの顕微鏡において光変換、またはレーザアブレーションのような局所的な光学的操作の可能性はありません。それにもかかわらず、ホーム内蔵セットアップは、特定の用途に使用することができます。
4.意義と今後のアプリケーション
LSFMは、ライブ胚の大ボリュームの高速撮像の日付に利用可能な最良の方法です。共焦点顕微鏡上で考えた実験のほとんどは、上述の利点を有する光シート顕微鏡で行うことができます。眼の発生のイメージングの場合には、LSFMの速度が重要なパラメータではありません。その代わりに、サンプルの位置が低い光毒性と柔軟性が決定的な利点です。
LSFMデータは、良好なデコンボリューションの結果を達成するのに役立ち、また、自動画像解析及びオブジェクト追跡のために有益である高いSNRを有します。結論として、LSFMは、胚発生とOveraの上で定量化可能なデータを生成するための素晴らしいツールですLL細胞およびその後のモデリングと、問題のプロセスの物理的な説明については、組織の特徴。
開示事項
このビデオ - 記事の発行はツァイスによってサポートされています。
謝辞
We want to thank Tobias Pietzsch for providing his powerful open source software BigDataViewer. We thank the Light Microscopy Facility of the Max Planck Institute of Molecular Cell Biology and Genetics (MPI-CBG), namely Jan Peychl, Sebastian Bundschuh and Davide Accardi for technical assistance, perfect maintenance of the microscopes used in the study and for comments on the manuscript and H. Moon (MPI-CBG Scientific Computing Facility) for the BigDataServer. We thank Julia Eichhorn for assembling the Movie 1. The Norden lab members and Svea Grieb provided many helpful comments on the manuscript. Jaroslav Icha, Christopher Schmied and Jaydeep Sidhaye are members of the International Max Planck Research School for Cell, Developmental and Systems Biology and doctoral students at TU Dresden. Pavel Tomancak is supported by the ERC Starting Grant: Quantitative Analysis of the Hourglass Model of Evolution of Development and Human Frontier Science Program Young Investigator grant RGY0093/2012. Caren Norden is supported by the Human Frontier Science Program (CDA-00007/2011) and the German Research Foundation (DFG) [SFB 655, A25].
資料
| Name | Company | Catalog Number | Comments |
| Lightsheet Z.1 microscope | Carl Zeiss Microscopy | ||
| Low melting point agarose | Roth | 6351.1 | |
| Low melting point agarose | Sigma | A4018 or A9414 | |
| Ethyl 3-aminobenzoate methanesulfonate (MESAB/MS-222/Tricaine) | Sigma | E10521 | |
| N-Phenylthiourea (PTU) | Sigma | P7629 | |
| 500 nm red fluorescent beads F-Y 050 | Millipore (Estapor) | 80380495 | |
| 20 μl (1 mm inner diameter, marked black) capilllaries | Brand | 701904 | sold as spare part for transferpettor |
| Teflon tip plungers for 20 μl capillaries | Brand | 701932 | sold as spare part for transferpettor |
| Circular glass coverslips diameter 18 mm, selected thickness 0.17 mm | Thermo scientific (Menzel-Glaser) | ||
| O-rings for chamber windows 17×1.5 mm | Carl Zeiss Microscopy | ||
| Tweezers, style 55 | Dumont | 0209-55-PO | |
| 50 ml Luer-Lock syringes | Becton Dickinson | 300865 | |
| 150 cm extension cable for infusion compatible with Luer-Lock syringes | Becton Dickinson | 397400 | |
| Links | |||
| Link 1 Multiview reconstruction application | https://github.com/bigdataviewer/SPIM_Registration http://fiji.sc/Multiview-Reconstruction | ||
| Link 2 BigDataViewer | https://github.com/bigdataviewer | ||
| Link 3 BigDataViewer | http://fiji.sc/BigDataViewer | ||
| Link 4 Multiview reconstruction application-issues | https://github.com/bigdataviewer/SPIM_Registration/issues | ||
| Link 5 BigDataViewer-issues | https://github.com/bigdataviewer/bigdataviewer_fiji/issues | ||
| Link 6 BigDataServer | http://fiji.sc/BigDataServer |
参考文献
- Pantazis, P., Supatto, W. Advances in whole-embryo imaging: a quantitative transition is underway. Nat Rev Mol Cell Biol. 15 (5), 327-339 (2014).
- Truong, T. V., Supatto, W. Toward high-content/high-throughput imaging and analysis of embryonic morphogenesis. Genesis. 49 (7), 555-569 (2011).
- Keller, P. J. Imaging Morphogenesis: Technological Advances and Biological Insights. Science. 340 (6137), 1234168-1234168 (2013).
- Huisken, J., Swoger, J., Del Bene, F., Wittbrodt, J., Stelzer, E. H. K. Optical Sectioning Deep Inside Live Embryos by Selective Plane Illumination Microscopy. Science. 305 (5686), 1007-1009 (2004).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J Microsc. 170 (Pt 3), 229-236 (1993).
- Jemielita, M., Taormina, M. J., DeLaurier, A., Kimmel, C. B., Parthasarathy, R. Comparing phototoxicity during the development of a zebrafish craniofacial bone using confocal and light sheet fluorescence microscopy techniques. J Biophoton. 6 (11-12), 920-928 (2012).
- Stelzer, E. H. K. Light-sheet fluorescence microscopy for quantitative biology. Nat Meth. 12 (1), 23-26 (2015).
- Chen, B. C., Legant, W. R., et al. Lattice light-sheet microscopy: Imaging molecules to embryos at high spatiotemporal resolution. Science. 346 (6208), 1257998-1257998 (2014).
- Keller, P. J., Ahrens, M. B. Visualizing Whole-Brain Activity and Development at the Single-Cell Level Using Light-Sheet Microscopy. Neuron. 85 (3), 462-483 (2015).
- Pampaloni, F., Chang, B. -. J., Stelzer, E. H. K. Light sheet-based fluorescence microscopy (LSFM) for the quantitative imaging of cells and tissues. Cell Tissue Res. 360 (1), 129-141 (2015).
- Weber, M., Mickoleit, M., Huisken, J. Light sheet microscopy. Quantitative Imaging in Cell Biology. , 193-215 (2014).
- Strobl, F., Stelzer, E. H. K. Non-invasive long-term fluorescence live imaging of Tribolium castaneum embryos. Development. 141 (11), 2361-2361 (2014).
- Kaufmann, A., Mickoleit, M., Weber, M., Huisken, J. Multilayer mounting enables long-term imaging of zebrafish development in a light sheet microscope. Development. 139 (17), 3242-3247 (2012).
- Maizel, A., von Wangenheim, D., Federici, F., Haseloff, J., Stelzer, E. H. K. High-resolution live imaging of plant growth in near physiological bright conditions using light sheet fluorescence microscopy. Plant J. 68 (2), 377-385 (2011).
- Swoger, J., Muzzopappa, M., Lòpez-Schier, H., Sharpe, J. 4D retrospective lineage tracing using SPIM for zebrafish organogenesis studies. J Biophoton. 4 (1-2), 122-134 (2010).
- Keller, P. J., Schmidt, A. D., Wittbrodt, J., Stelzer, E. H. K. Reconstruction of zebrafish early embryonic development by scanned light sheet microscopy. Science. 322 (5904), 1065-1069 (2008).
- Wu, Y., Ghitani, A., et al. Inverted selective plane illumination microscopy (iSPIM) enables coupled cell identity lineaging and neurodevelopmental imaging in Caenorhabditis elegans. Proc Natl Acad Sci USA. 108 (43), 17708-17713 (2011).
- Sarov, M., Barz, C., et al. A genome-wide resource for the analysis of protein localisation in Drosophila. bioRxiv. , 028308 (2015).
- Amat, F., Lemon, W., et al. Fast, accurate reconstruction of cell lineages from large-scale fluorescence microscopy data. Nat Meth. 11 (9), 951-958 (2014).
- Preibisch, S., Saalfeld, S., Schindelin, J., Tomancak, P. Software for bead-based registration of selective plane illumination microscopy data. Nat Meth. 7 (6), 418-419 (2010).
- Preibisch, S., Rohlfing, T., Hasak, M. P., Tomancak, P. Mosaicing of single plane illumination microscopy images using groupwise registration and fast content-based image fusion. SPIEMed Imaging. 6914, 69140E (2008).
- Reynaud, E. G., Peychl, J., Huisken, J., Tomancak, P. Guide to light-sheet microscopy for adventurous biologists. Nat Meth. 12 (1), 30-34 (2015).
- Keller, P. J., Schmidt, A. D., Wittbrodt, J., Stelzer, E. H. K. Digital Scanned Laser Light-Sheet Fluorescence Microscopy (DSLM) of Zebrafish and Drosophila Embryonic Development. Cold Spring Harb Protoc. 2011 (10), (2011).
- Pinto-Teixeira, F., Muzzopappa, M., Swoger, J., Mineo, A., Sharpe, J., Lòpez-Schier, H. Intravital imaging of hair-cell development and regeneration in the zebrafish. Front Neuroanat. , (2013).
- Keller, P. J. In vivo imaging of zebrafish embryogenesis. METHODS. , 1-11 (2013).
- Pitrone, P. G., Schindelin, J., et al. OpenSPIM: an open-access light-sheet microscopy platform. Nat Meth. 10 (7), 598-599 (2013).
- Gualda, E. J., Vale, T., Almada, P., Feijò, J. A., Martins, G. G., Moreno, N. OpenSpinMicroscopy: an open-source integrated microscopy platform. Nat Meth. 10 (7), 599-600 (2013).
- Martinez-Morales, J. R., Rembold, M., et al. ojoplano-mediated basal constriction is essential for optic cup morphogenesis. Development. 136 (13), 2165-2175 (2009).
- Strzyz, P. J., Lee, H. O., Sidhaye, J., Weber, I. P., Leung, L. C., Norden, C. Interkinetic Nuclear Migration Is Centrosome Independent and Ensures Apical Cell Division to Maintain Tissue Integrity. Dev Cell. 32 (2), 203-219 (2015).
- Young, L. K., Jarrin, M., Saunter, C. D., Quinlan, R., Girkin, J. M. Using SPIM to track the development of the focal power of the zebrafish lens. SPIE BiOS. 9334, 933408 (2015).
- Preibisch, S., Amat, F., et al. Efficient Bayesian-based multiview deconvolution. Nat Meth. 11 (6), 645-648 (2014).
- Schindelin, J., Arganda-Carreras, I., et al. Fiji: an open-source platform for biological-image analysis. Nat Meth. 9 (7), 676-682 (2012).
- Pietzsch, T., Saalfeld, S., Preibisch, S., Tomancak, P. BigDataViewer: visualization and processing for large image data sets. Nat Meth. 12 (6), 481-483 (2015).
- Brown, M., Lowe, D. G. Invariant Features from Interest Point Groups. Proceedings of the British Machine Vision Conference. , 23.1-23.10 (2002).
- Saalfeld, S., Fetter, R., Cardona, A., Tomancak, P. Elastic volume reconstruction from series of ultra-thin microscopy sections. Nat Meth. 9 (7), 717-720 (2012).
- Fischler, M. A., Bolles, R. C. Random sample consensus: a paradigm for model fitting with applications to image analysis and automated cartography. Commun ACM. 24 (6), 381-395 (1981).
- Zolessi, F. R., Poggi, L., Wilkinson, C. J., Chien, C. -. B., Harris, W. A. Polarization and orientation of retinal ganglion cells in vivo. Neural Dev. 1 (1), 2 (2006).
- Stepanova, T., Slemmer, J., et al. Visualization of microtubule growth in cultured neurons via the use of EB3-GFP (end-binding protein 3-green fluorescent protein). J Neurosci. 23 (7), 2655-2664 (2003).
- Weber, M., Mickoleit, M., Huisken, J. Multilayer Mounting for Long-term Light Sheet Microscopy of Zebrafish. J Vis Exp. (84), e51119 (2014).
- Udan, R. S., Piazza, V. G., Hsu, C. W., Hadjantonakis, A. K., Dickinson, M. E. Quantitative imaging of cell dynamics in mouse embryos using light-sheet microscopy. Development. 141 (22), 4406-4414 (2014).
- Ichikawa, T., Nakazato, K., et al. Live imaging and quantitative analysis of gastrulation in mouse embryos using light-sheet microscopy and 3D tracking tools. Nat Protoc. 9 (3), 575-585 (2014).
- Schmied, C., Stamataki, E., Tomancak, P. Open-source solutions for SPIMage processing. Methods Cell Biol. 123, 505-529 (2014).
- Amat, F., Höckendorf, B., Wan, Y., Lemon, W. C., McDole, K., Keller, P. J. Efficient processing and analysis of large-scale light-sheet microscopy data. Nat Protoc. 10 (11), 1679-1696 (2015).
- Schmid, B., Shah, G., et al. High-speed panoramic light-sheet microscopy reveals global endodermal cell dynamics. Nat Commun. 4, 1-10 (2013).
- Gualda, E. J., Pereira, H., Vale, T., Estrada, M. F., Brito, C., Moreno, N. SPIM-fluid: open source light-sheet based platform for high-throughput imaging. Biomed Opt Express. 6 (11), 4447-4456 (2015).
- McGorty, R., Liu, H., Kamiyama, D., Dong, Z., Guo, S., Huang, B. Open-top selective plane illumination microscope for conventionally mounted specimens. Opt Express. 23 (12), 16142-16153 (2015).
- Jemielita, M., Taormina, M. J., et al. Spatial and Temporal Features of the Growth of a Bacterial Species Colonizing the Zebrafish Gut. mBio. 5 (6), e01751-14-8 (2014).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved