
Method Article
レジオネラ・ニューモ外膜小胞:マクロファージ上における炎症誘発ポテンシャルの単離と解析
要約
ここでは、 レジオネラ・ニューモフィラ (L.ニューモ )液体培養物から外膜小胞(OMVを)の精製を記載しています。これらの精製された小胞は、次いで、それらのプロ炎症性の可能性を分析するために、マクロファージの治療のために使用されます。
要約
Bacteria are able to secrete a variety of molecules via various secretory systems. Besides the secretion of molecules into the extracellular space or directly into another cell, Gram-negative bacteria can also form outer membrane vesicles (OMVs). These membrane vesicles can deliver their cargo over long distances, and the cargo is protected from degradation by proteases and nucleases.
Legionella pneumophila (L. pneumophila) is an intracellular, Gram-negative pathogen that causes a severe form of pneumonia. In humans, it infects alveolar macrophages, where it blocks lysosomal degradation and forms a specialized replication vacuole. Moreover, L. pneumophila produces OMVs under various growth conditions. To understand the role of OMVs in the infection process of human macrophages, we set up a protocol to purify bacterial membrane vesicles from liquid culture. The method is based on differential ultracentrifugation. The enriched OMVs were subsequently analyzed with regard to their protein and lipopolysaccharide (LPS) amount and were then used for the treatment of a human monocytic cell line or murine bone marrow-derived macrophages. The pro-inflammatory responses of those cells were analyzed by enzyme-linked immunosorbent assay. Furthermore, alterations in a subsequent infection were analyzed. To this end, the bacterial replication of L. pneumophila in macrophages was studied by colony-forming unit assays.
Here, we describe a detailed protocol for the purification of L. pneumophila OMVs from liquid culture by ultracentrifugation and for the downstream analysis of their pro-inflammatory potential on macrophages.
概要
細菌は異なるメカニズム1を介して病原性因子を分泌することができます。よく知られている分泌システムのほかに、グラム陰性細菌は、情報を交換することができ、直径及び二層状膜構造を持つ小さな、回転楕円体小胞10〜300nmである外膜小胞(OMVを)を介して病原性因子を提供します。彼らは、成長環境(液体培養、固体培養、およびバイオフィルム)の様々な、すべての成長段階2、3に分泌されます。 OMVを輸送する重要な手段である( 例えば、OMVの表面上に見出されるタンパク質、アドヘシン、毒素、酵素、ならびにためのLPSのための)4。管腔内の貨物は、タンパク質分解から保護されたので、長い距離にわたって作用することができる、と小胞は、体液や遠隔臓器5、6で見つけることができますされています「外部参照」> 7、8。彼らは認識し、真核細胞9,10によって取り込まが、さらに、それらは、宿主細胞4に細菌およびそれらの浸潤の結合を促進することが可能であることはできません。 レジオネラ・ニューモフィラ (L.ニューモは )のOMVを解放することができ、グラム陰性細菌です。ヒトの肺では、それは主に、その天然の宿主は淡水アメーバ11であっても、肺胞マクロファージに感染します。 L.・ニューモフィラ感染はレジオネラ症、肺炎12の重症型を引き起こす可能性があります。また、宿主細胞内のブロックファゴソーム - リソソーム融合を。複製ニッチ、 レジオネラ -含有小胞(LCV)は、13、14を形成することも、ホスト細胞内小器官を動員します。リソソーム分解だけでなく、エフェクタータンパク質TRAによって阻害されますIV型分泌システムを介して、だけでなく、OMVを15のリリースによりnslocation。
細菌培養物からのOMVの精製は、レシピエント細胞に対する効果を分析するために必要とされます。 L.ニューモフィラのOMVのタンパク質含有量に及び肺胞上皮細胞16が、ヒト肺組織移植とその後の研究に小胞の影響に焦点を当てた初期の研究はL.ニューモフィラのOMVは肺胞マクロファージ17によって取り込まれることを実証しました。
OMVを、本病原体関連分子パターン(PAMP)および他の細菌抗原として、それらは、真核細胞の感染に影響を与えると宿主免疫応答18を調節するかもしれません。 L.ニューモフィラのOMVが急速さらに、それらは、膜TLR2 19を活性化する 、宿主細胞膜と融合し、。それはL. pneumophことが知られているようILAのOMVは、炎症誘発性の様式16、17にマクロファージおよび上皮細胞を刺激し、我々は、ヒトおよびマウスのマクロファージにおける感染プロセスに対するのOMVの影響を分析しました。
ここでは、直接、または感染後、差動超遠心分離により分泌のOMVを分離し、真核生物の宿主細胞に小胞の影響を評価するために、液体培養中でL.・ニューモフィラの栽培のためのプロトコルを説明します。
プロトコル
1.培地および寒天プレートを準備します
- ブロス培地(YEB)の1 Lを用意。 ACES 10gと蒸留水900mL中で酵母エキス10gを溶解させます。 KOH(5 N)で6.9にpHを調整します。 L-システイン(蒸留水10mLの0.4 g)を10 mLを加え、Feの10ミリリットル(NO 3)3 x9H 2 O(蒸留水10mLの0.25 g)を得ました。蒸留水で1Lに埋めるとソリューション(孔径:0.22μm)を滅菌ろ過します。 4℃で保存。
- 寒天プレートの緩衝木炭酵母エキス(BCYE)を準備します。 ACES 10gと蒸留水900mL中で酵母エキス10gを溶解させます。 KOH(5 N)で6.9にpHを調整します。寒天の15グラム、活性炭の2.5グラムを追加します。蒸留水、オートクレーブで1Lに埋めます。
- L-システイン(10mLの蒸留水で0.4グラム)の10 mLを加え、Feの10ミリリットル(NO 3)3 x9H 2 O(10mLの蒸留水で0.25グラム、0.22μmの濾過滅菌の両方BCYE(約50℃)を冷却するための孔)。 4℃でプレートや店舗を注ぎます。
2. L.ニューモフィラの育成
- BCYE寒天プレート上L.ニューモ株コービー(野生型、WT)を広げ、3日間37℃でそれらをインキュベートします。前培養プレートからL.ニューモでOD 0.3の600でYEB 10mLのを接種します。 6時間、回転シェーカー(150rpmで)上で37℃で細菌を培養します。
- 血液寒天プレート上に懸濁液100μLを拡散することにより液体培養物の純度を確認してください。 37℃で一晩インキュベートします。
- 新鮮なYEB培地の90 mLに残りの液体培養を追加し、約16〜20時間を要しOD 3.0から3.5の600に到達するために、回転シェーカー(37℃、回転数150rpm)上でインキュベートします。
3.準備とL.ニューモのOMVを定量化
注:以下の遠心分離工程のすべてを行って無菌条件下で4℃で秒。
- 細菌をペレット化し、20分間4000×gで液体培養を遠心。 、新たな遠心チューブに上清を移し細菌ペレットを廃棄し、遠心分離(20分間4,000×gで)を繰り返します。一度、この手順を繰り返します。
- 滅菌フィルタ残りの上清は二回(孔径:0.22μmで)。 3時間、100,000×gで超遠心管と超遠心分離機に細菌を含まない上清を移します。
- 上清を除去し、それを捨てます。汚染タンパク質およびLPSを除去するために、滅菌リン酸緩衝生理食塩水(PBS)および超遠心機(3時間100,000×gで)にOMVペレットを再懸濁します。
- 上清を捨て、滅菌PBSの500μLにOMVペレットを再懸濁します。準備された小胞の細菌汚染を排除するストリーク血液寒天プレート上とBCYE寒天プレート上で20μL。両方とも37℃(3日間終夜血液寒天プレートおよびBCYE寒天プレートをインキュベート6; C)。
- 製造業者の指示に従ってビシンコニン酸アッセイを用いOMV調製物から得られたタンパク質の量を定量します。
注:100mLのL.のニューモ文化の濃度は、通常を1μg/μLです。 -20℃で調製し、定量化のOMVを保管してください。
4.前処理マクロファージ
- THP-1細胞を準備します。
注:THP-1白血病患者由来の単球細胞株です。- 24ウェルあたり2×10 5 THP-1細胞を添加し、マクロファージ様細胞に20 nmのホルボール12-ミリステート13-アセテート(PMA)を加えることにより、それらを区別します。 37℃で24時間インキュベートします。
- 新鮮な培地の500μLで培地を交換し、さらに24時間インキュベート。 THP-1細胞のための最適な培地を、10%ウシ胎児血清を補充したRPMI1640高グルコースで構成されています。
- 参照ので説明したように、マウス骨髄由来マクロファージ(mBMDM)を単離しますレンス20。
- OMVを用いたTHP-1由来マクロファージやmBMDMを扱います。
- ステップ3で調製したOMVを解凍し、ヒトまたはマウスのマクロファージへのタンパク量(0.1、1、および10μg/ mL)をに従ってそれらを追加します。少なくとも20時間、37℃でのOMVとマクロファージをインキュベートします。 ELISAのために上清を使用するか、手順5に進みます。
5.マクロファージに感染し、コロニー形成単位(CFU)アッセイで細菌の複製を評価します
- ステップ2.1からL.ニューモを使用して 、ステップ4.3からTHP-1細胞またはmBMDMを前処理された、およびコントロール(2×10 5月24日のウェル)としてマクロファージを事前に処理されていません。培地を交換しないでください。
- L.ニューモコービー(0.5の感染多重度(MOI)との両方; 1×10 5 L.ニューモ / 24ウエル)のフラジェリン欠いた変異体とL.ニューモコービーWTとmBMDMとTHP-1細胞に感染し、24インキュベートそれぞれ48時間、。 BOを準備第L.ニューモコービー(WTまたはフラジェリン欠いている変異体)ステップ2.1で説明したように。
- 溶解サポニンを添加することにより、それらの培地中の細胞(最終濃度:0.1%)、5分間37℃でインキュベートします。
- ピペッティングにより細菌を再懸濁し、反応容器に懸濁液を移します。滅菌PBSでメディアを含有L.ニューモの連続希釈液を調製します。
- ストリークBCYE寒天プレート上で必要な希釈液50μL、37℃で3日間インキュベートします。
- 視覚的に形成されたコロニーを数えます。 CFUを計算
 前処理が、100%に設定されたマクロファージを、感染していないためCFUカウント結果を正規化します。
前処理が、100%に設定されたマクロファージを、感染していないためCFUカウント結果を正規化します。
結果
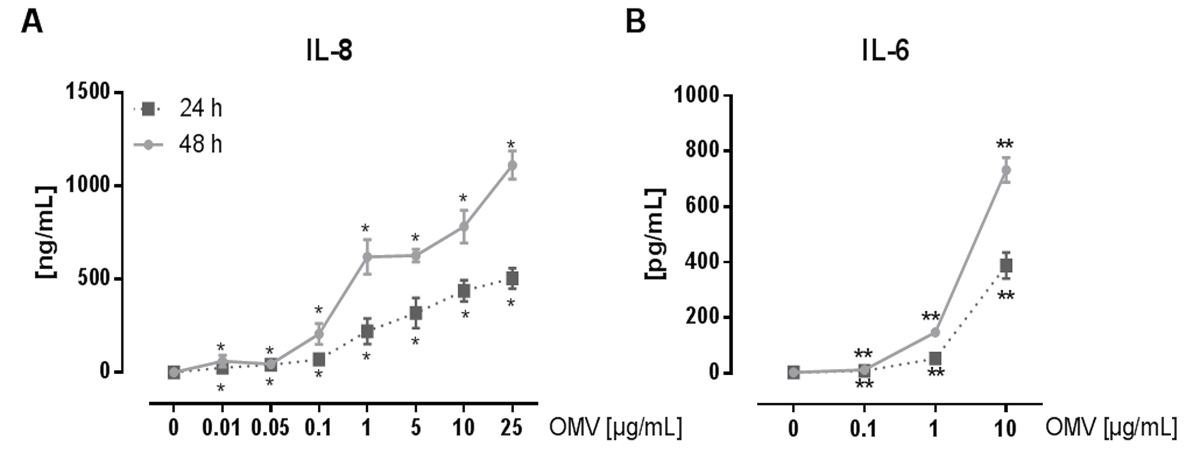
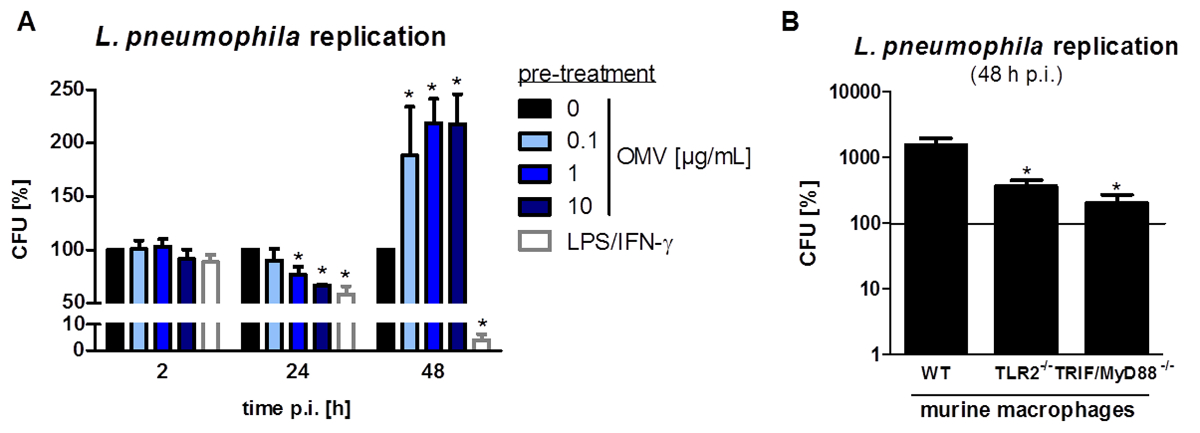
L.ニューモのOMVを調製するための、および感染後のマクロファージの炎症誘発性応答に及ぼす影響を分析する実験は、図1に示されています。調製したOMVの炎症誘発電位は、 図2に示されているPMA分化THP-1細胞上で分析することができます。 THP-1細胞を、IL-8およびIL-6分泌の時間および用量依存的増加に応答します。 図3にCXCL1 ELISAによって提示されるように加え、L。ニューモ OMV認識の異なるTLRの影響は、異なる遺伝的背景からmBMDMを用いて分析することができます。 OMV刺激後CXCL1分泌WTマウスからmBMDM、しばらくmBMDM TLR2 / 4 - / -大幅に少ない分泌。 THP-1マクロファージにおける細菌の複製にL.ニューモのOMVの影響を研究するために、細胞は、OMVを用いてプレインキュベートし、次いで、さらに感染させましたL.ニューモフィラ ( 図4 A)で。 THP-1由来マクロファージの刺激前には、最初の感染の24時間後に細菌の複製を減少させるが、それは後の時点(48時間π)でCFU数の倍増につながります。 図4 Bに提示されるようなマクロファージの感染後のOMV認識にシグナリングToll様受容体(TLR)の影響は、mBMDMによって評価することができます。 OMVのプレインキュベーション後のWT動物からmBMDMで十倍による細菌の複製が増加し、TLR2ながら- / -およびTRIF / MyD88に- / -細胞はL.・ニューモフィラ複製におけるこの増加を示しません。

図1:実験手順。 (A)L. ニューモコービーWT 10センチBCYE寒天プレートは、新鮮なYEBの90ミリリットルに移し、少量の液体培地(10ミリリットル)を、接種するために使用されているから6時間後の培地。少量はまた、純度を確認するために、血液寒天プレート上にプレーティングされます。細菌は、初期定常期(OD 600 = 3.0〜3.5)になるまで37℃でインキュベートします。細菌を除去するために、(B)液体培養物を遠心分離し、滅菌濾過しました。 レジオネラフリー次いで、上清をPBSに再懸濁し、再び超遠心分離されたOMVペレットを得るために超遠心分離されます。単離された小胞は、純度をチェックし、再懸濁し、およびタンパク質の量を定量します。スケールバーは2.5 cmで表します。 (C)ヒトまたはマウスのマクロファージを定量したOMVで刺激します。細胞培養上清をELISAに使用することができ、またはマクロファージは、10センチBCYE寒天プレート上のCFUアッセイによって細菌複製を決定するために、L・ニューモで感染させることができます。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

図2:L.ニューモフィラのOMVによるTHP-1細胞のプロ炎症活性化。 (A)ここで、単球THP-1細胞株は、肺胞マクロファージのモデルとして使用されます。 PMA分化THP-1細胞を、それぞれ、24および48時間L.ニューモのOMV(0.01〜25 / mlの)の漸増用量で処理しました。無細胞上清を、IL-8 ELISAのために使用しました。 SEM±3つの独立した実験の平均値が示されています。 THP-1細胞は、時間および用量依存的であった重要なIL-8分泌、とわずか0.01 / mlのL.ニューモのOMVに反応しました。 (B)L。ニューモフィラのOMV(0.1μgの/ mL)をPMA分化THP-1細胞を刺激するために使用しました。上清をインキュベーションの24および48時間後に収集し、そして放出されたIL-6はELISAにより上清中で測定しました。。 SEM±3つの独立した実験の平均値が示されています。偶数のOMVの最低用量(0.1μgの/ mL)で、IL-6の有意な量を分泌したTHP-1細胞。 IL-6の分泌は、OMV用量の増加に伴って延長し、インキュベーション時間と共に増加しました。統計:マンホイットニー検定。 * P <対応0 / mlのOMVに比べ0.05および**はp <0.01。リファレンス20からの許可を得て転載。この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

図3:マクロファージの炎症誘発性活性化はTLR2 / 4に依存します。 WTからmBMDM及びTLR2 / 4 - / -マウスはL.ニューモのOMV(0.1を1μg/ mL)でインキュベートしました。 CXCL1分泌は、それぞれ、24および48時間後にELISAによって分析しました。ザ3つの独立した実験の平均値±SEMが示されています。 WTマウスからmBMDMはL.ニューモ OMVのインキュベーション後に用量依存CXCL1の分泌と答えました。 TLR2 / 4 - / - mBMDMは、WT mBMDMと比較して有意に少ないCXCL1を分泌し、この分泌は用量依存的に増加しませんでした。統計:マンホイットニー検定。対応する0 / mlのOMVと比較して、*はp <0.05; #P <0.05は等しく扱わWTサンプルと比較します。リファレンス20からの許可を得て転載。この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

図4:マクロファージにおけるL.ニューモ OMVプレインキュベーション増加細菌複製。 (A)分化したTHP-1細胞のOMV(0.1、1、または10μと共にプレインキュベートしました。;グラム/ ml)またはLPS / IFN-γ(200 ngの/ mLの各)またはコントロールの未処理のままとしました。プレインキュベーション(20時間)後、THP-1細胞を、それぞれ、2、24、及び48時間L.ニューモコービーWT(MOI 0.5)で感染させました。 THP-1細胞は、サポニンを添加することによって溶解し、そして細菌をBCYE寒天プレート上にプレーティングしました。 CFUは、すべての時間点の後0 / mlのOMVと比較して計算しました。バーは、3つの独立した実験、技術的な二重に実行される各の平均値±SEMを表します。細菌の取り込みの差異は、前処理されていない細胞と比較して(2時間の感染後(PI))が存在しませんでした。細菌の複製の変化は、それぞれ、24および48時間後に測定しました。 LPS / IFN-γ前処理されたTHP-1細胞は、これはまた、L。ニューモ OMVプレ処理した細胞の用量依存的に観察された細菌負荷の24時間PIの減少を示しました。後の時点(48時間PI)では、OMVの前処理されたTHP-1細胞は、L。ニューモフィラ再で倍増を示しましたLPS / IFN-γ前処理されたマクロファージ一方ひだは、細菌負荷のさらなる低減を示しました。統計:マンホイットニー検定。 * P <対応0 / mlのOMVに比べ0.05および**はp <0.01。 20時間は0.1μg/ mLのL。ニューモフィラのOMVでプレインキュベートし、次いで、フラジェリンを感染させた(B)mBMDMは異なる遺伝的背景を有するマウス(WT、TLR2 - / - 、およびTRIF / MyD88の- / - )から48時間L.ニューモコービー(MOI 0.5)の欠損変異体。 mBMDMは、サポニンを添加することによって溶解し、およびレジオネラを BCYE寒天プレート上にプレーティングしました。 CFUは、実線で示され、0 / mlのOMVに対して計算しました。バーは平均値±±3つの独立した実験のSEM、二重に行っそれぞれを表します。 WTマウスからmBMDMはOMV前処理後のL.・ニューモフィラの複製の増加を示しました。 TLR2 - / -マクロファージは、有意に減少を示しましたレジオネラ複製、 - / - mBMDM。統計:マンホイットニー検定。 WTサンプルと比較してp <0.05。リファレンス20からの許可を得て転載。この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
ディスカッション
細菌性病原体およびそれらの標的細胞上のこれらの膜小胞の影響のOMVは、現在集中的に研究されています。例えば、 クロストリジウム・パーフリンジェンスののOMVが、マクロファージにおけるサイトカインの分泌を誘導由来、Bリンパ球は、 ボレリアブルグドルフェリからのOMVによって活性化され、およびヘリコバクター・ピロリは、膜小胞は、胃上皮細胞21、22、23に作用することができる-releasedできます。 L.ニューモ 、非定型肺炎の重症型を誘導することができる細胞内病原体は、また、肺上皮細胞とマクロファージ16、19を活性化することができるのOMVを解放します。ここでは、肺炎でのOMVの潜在的な役割を研究するための培養液からL.ニューモのOMVの小規模単離するための詳細なプロトコルを提示します。無菌コンディット下で作業することが重要ですイオンおよび純粋L.・ニューモフィラを得るために、他の細菌からの汚染を排除するためには、OMV調製物が由来しました。 OMVを単離は、最大のOMVは、この濾過工程で失われるので、これは、OMVの収率を低下させるにもかかわらず、L。ニューモで得られるOMVペレットの汚染を防止するために、0.22ミクロンの細孔を介して濾過するステップを含みます。
さらに、我々はより密接のOMVが細胞外細菌15でLCVの内側にリリースされたレジオネラ肺炎、の状況に近づけるために、これらの単離された小胞およびL.ニューモで感染した細胞にヒトおよびマウスマクロファージの応答をテストしました。採用OMVの用量は、インキュベーションの24時間後のヒトマクロファージのインビトロ感染における自由OMV量に応じて推定されている(参考文献20に記載)。他のレシピエント細胞の刺激のために、またはin vivo実験において 、他のOMV投与量が必要になるかもしれませんし、確立されなければなりません。 L.ニューモフィラのOMVの効果の分析はイェーガーとシュタイナー24によって記述されたプロトコルに進歩を表しています。
ここでは、PMA分化THP-1細胞は、初代ヒト材料の限られた在庫状況により、肺胞マクロファージのモデルとして役立ちます。 PMAの添加は、マクロファージ様細胞25への単球THP-1細胞を区別する。さらに、それらは、L。ニューモ研究 26のための周知のモデル細胞株です。このヒト単球細胞株のほか、mBMDM細胞が使用されます。 mBMDMは広くL.ニューモ 27、28、29の効果の研究のために受理されています。異なるのTLRまたは他のタンパク質のための遺伝子ノックアウトを使用する可能性は、それらのOMV効果を研究するための貴重なツールとなっています。 ORDで実験あたりのマウスの量を低下させるために、ERは、mBMDMではなく、マクロファージの制限による肺胞マクロファージから使用されています。主な実験は、検証のために肺胞マクロファージを必要とするかもしれません。
OMVを精製するための超遠心分離の本明細書に記載のプロトコルに加えて、Chutkan らのプロトコルに含まれている密度勾配遠心分離を行うことができます。 30。これは、得られたOMV調製物の純度を向上させると共精製されたタンパク質の凝集、フラジェリン、およびLPSの量を減らすことができます。得られたOMV調製物の純度は、品質管理における補助ステップとして、透過型電子顕微鏡法によって、またはナノ粒子追跡分析によって分析することができます。これは、ここで提示タンパク質の測定手順を超えて、定量化の追加の手段を提供することができます。任意に、LPS濃度はリムルス変形細胞溶解物の試験によって分析することができます。 OMV収率が低い場合、遠心フィルターを経由して、追加の濃縮工程は、ここで行われなかった、実行することができます。収量が予想より低かった場合は、OMVを廃棄しました。
OMVを背後にある生物学的メカニズムと機能を解明するための継続的な取り組みの一環として、OMVの生産に異なるストレス条件の影響を試験することができます。栄養欠乏、インキュベーション温度の変化、または有害な薬剤への曝露は、OMV分泌31に影響を与える可能性があります。可能性のあるストレス条件はKlimentovaとStulik 32によってプロトコルで説明されています。また、ハイパーまたはhypovesiculating L.・ニューモ変異体が生成することができます。異なるOMV調製物は、その後、(文献17に記載)、ヒト肺組織外植片、あるいはin vivoモデルにおけるマクロファージの感染実験で分析することができました。細菌の通信における自然免疫シグナル伝達におけるのOMVの役割、彼らの影響力のほかに実験的に対処することができます。さらに、様々な自然免疫シグナル伝達カスケードの影響は、マウスノックアウト細胞の使用またはヒト細胞株におけるCRISPR / Cas9ノックアウトの生成によって分析されることがあります。 OMVのこの基礎研究はすでに髄膜炎菌 33によって送信された髄膜炎Bのために存在する新たなワクチン戦略の開発を支援します。
OMVの単離および特徴上のプロトコルから始めて、1は、他のグラム陰性菌および他の宿主細胞にこれを適用することができます。それは、液体培養における細菌の増殖を調整する必要があります。 Chutkan らによって発表されたプロトコル。 大腸菌や緑膿菌 30からのOMVの生成に関する詳細な情報を提供しています。文化を溶解し、細菌の増加と汚染タンパク質とMEMを避けるために、後期定常期に達するべきではありませんブレーン。さらに、宿主細胞の刺激のために使用されるOMV用量は依然として細胞毒性の低い速度を確保しながら、 インビボでの感染の間のOMVの存在量に応じて決定する必要があります。このように、OMVをの病理学的役割は、種間の通信、およびホスト病原体相互作用に及ぼす影響を調べることができました。
さらに肺炎でL.ニューモのOMVの役割を研究するために、十分な収量と同等の感染実験で標準化されたOMV調製物が必要とされています。このプロトコルは、分離手順を標準化し、他のグラム陰性菌および他の宿主細胞へのOMVの研究を拡張するのに役立ちます。さらに、研究は、 インビボでの設定に実験を拡張するために使用することができる、 インビトロ知識に詳細恩恵を受ける。今後は、このプロトコルは、血清又は気管支LAVとして、主要な生物学的材料からのOMVの単離に拡張することができ年齢流体は、生理学的条件の下でリリースするOMVの組成への洞察を得るために。これは、OMV組成物の重要なパラメータを決定し、in vitroで -generatedのOMVの性質を理解するのに役立ちます。
開示事項
The authors have nothing to disclose.
謝辞
私たちは、TLR2をご提供するための教授マルクスSchnareに感謝- / -およびTLR2 / 4 - / -マウスおよびTRIF / MyD88のための教授カルステンKirschning - / -マウス。 、ドイツ学術協会(SFB / TR-84; BundesministeriumがビルドゥングウントForschung(:: - - http://www.bmbf.de/メッドCAPSYS FKZ 01X1304E FKZ 0316175B、電子バイオmiRSys e)をFURことで、この作業の一部は資金を供給されましたhttp://www.sfb-tr84.de/)、およびHessisches MinisteriumのfürWissenschaftウントクン(LOEWE医療RNomics - FKZ 519/03 / 00.001-(0003); http://www.proloewe.de/medicalrnomics)、すべてのBSへ。
資料
| Name | Company | Catalog Number | Comments |
| 10 cm Petri dish | Sarstedt AG & Co KG (Nuembrecht, Germany) | 82.1473 | |
| 70 Ti rotor | Beckman Coulter Incorporation (California, USA) | 337922 | |
| ACES | Carl Roth GmbH & Co KG (Karlsruhe, Germany) | 9138.2 | |
| activated charcoal | Carl Roth GmbH & Co KG (Karlsruhe, Germany) | X865.2 | |
| agar-agar, Kobe I | Carl Roth GmbH & Co KG (Karlsruhe, Germany) | 5210.2 | |
| Columbia agar with 5% sheep blood | Becton Dickinson GmbH (Heidelberg, Germany) | 254005 | |
| cuvettes | Sarstedt AG & Co KG (Nuembrecht, Germany) | 67.742 | |
| ELISA (human) | BD OptEIA™; Becton Dickinson GmbH (Heidelberg, Germany) | IL-8: 555244 IL-6: 550799 | |
| ELISA (murine) | DuoSet, R&D (Minneapolis, USA) | CXCL1: DY453-05 | |
| Fe(NO3)3x9H2O | Carl Roth GmbH & Co KG (Karlsruhe, Germany) | 5632.1 | |
| Fetal calf serum (FCS) | Life Technologies GmbH (Darmstadt, Germany) | 10270-106 | |
| Heracell 240i CO2 incubator | Thermo Fisher Scientific Germany BV & Co KG (Braunschweig, Germany) | 40830469 | |
| Heraeus Multifuge X3R | Thermo Fisher Scientific Germany BV & Co KG (Braunschweig, Germany) | 75004515 | |
| Inoculation loop | Sarstedt AG & Co KG (Nuembrecht, Germany) | 86.1567.010 | |
| KOH | Carl Roth GmbH & Co KG (Karlsruhe, Germany) | 6751.1 | |
| L. pneumophila Corby | --- | --- | kindly provided by Prof Dr Antje Flieger (RKI, Berlin, Germany) |
| L. pneumophila Corby ΔflaA | --- | --- | kindly provided by Prof Dr Klaus Heuner (RKI, Berlin, Germany) |
| L-cystein | Carl Roth GmbH & Co KG (Karlsruhe, Germany) | ||
| mBMDM | --- | --- | kindly provided by Prof Dr Markus Schnare (Philipps Univeristy Marburg, Marburg, Germany) and Prof Dr Carsten Kirschning (University Duisburg Essen, Essen, Germany) |
| PBS | Biochrom GmbH (Berlin, Germany) | L 1825 | |
| phorbol 12-myristate 13-acetate | Sigma-Aldrich Chemie GmbH (Taufkirchen, Germany) | P8139-1MG | |
| rotating shaker (MaxQ 6000) | Thermo Fisher Scientific Germany BV & Co KG (Braunschweig, Germany) | SHKE6000 | |
| RPMI 1640 high glucose | Life Technologies GmbH (Darmstadt, Germany) | 11875-093 | |
| saponin | Carl Roth GmbH & Co KG (Karlsruhe, Germany) | 9622.1 | |
| Ultrospec 10 | Biochrom Ltd (Cambridge, England) | 80-2116-30 | |
| sterile filter (pore size: 0.22 µm) | Corning Incorporated (new York, USA) | 431096 | |
| THP-1 | Sigma-Aldrich Chemie GmbH (Taufkirchen, Germany) | 88081201-1VL | |
| Sorvall Discovery 100 SE | Thermo Fisher Scientific Germany BV & Co KG (Braunschweig, Germany) | ||
| yeast extract | Carl Roth GmbH & Co KG (Karlsruhe, Germany) | 2363.2 | |
| Pierce BCA protein assay kit | Thermo Fisher Scientific Germany BV & Co KG (Braunschweig, Germany) | 23225 |
参考文献
- Cambronne, E. D., Roy, C. R. Recognition and delivery of effector proteins into eukaryotic cells by bacterial secretion systems. Traffic. 7 (8), 929-939 (2006).
- Ellis, T. N., Kuehn, M. J. Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiol Mol Biol Rev. 74 (1), 81-94 (2010).
- Beveridge, T. J. Structures of gram-negative cell walls and their derived membrane vesicles. J Bacteriol. 181 (16), 4725-4733 (1999).
- Kuehn, M. J., Kesty, N. C. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes Dev. 19 (22), 2645-2655 (2005).
- Chi, B., Qi, M., Kuramitsu, H. K. Role of dentilisin in Treponema denticola epithelial cell layer penetration. Res Microbiol. 154 (9), 637-643 (2003).
- Kolling, G. L., Matthews, K. R. Export of virulence genes and Shiga toxin by membrane vesicles of Escherichia coli O157:H7. Appl Environ Microbiol. 65 (5), 1843-1848 (1999).
- Horstman, A. L., Kuehn, M. J. Enterotoxigenic Escherichia coli secretes active heat-labile enterotoxin via outer membrane vesicles. J Biol Chem. 275 (17), 12489-12496 (2000).
- Dorward, D. W., Schwan, T. G., Garon, C. F. Immune capture and detection of Borrelia burgdorferi antigens in urine, blood, or tissues from infected ticks, mice, dogs, and humans. J Clin Microbiol. 29 (6), 1162-1170 (1991).
- Mashburn, L. M., Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature. 437 (7057), 422-425 (2005).
- Kesty, N. C., Mason, K. M., Reedy, M., Miller, S. E., Kuehn, M. J. Enterotoxigenic Escherichia coli vesicles target toxin delivery into mammalian cells. EMBO J. 23 (23), 4538-4549 (2004).
- Abu Kwaik, Y., Gao, L. Y., Stone, B. J., Venkataraman, C., Harb, O. S. Invasion of protozoa by Legionella pneumophila and its role in bacterial ecology and pathogenesis. Appl Environ Microbiol. 64, 3127-3133 (1998).
- Winn, W. C., Myerowitz, R. L. The pathology of the Legionella pneumonias. A review of 74 cases and the literature. Hum Pathol. 12 (5), 401-422 (1981).
- Ge, J., Shao, F. Manipulation of host vesicular trafficking and innate immune defence by Legionella Dot/Icm effectors. Cell Microbiol. 13 (12), 1870-1880 (2011).
- Hubber, A., Roy, C. R. Modulation of host cell function by Legionella pneumophila type IV effectors. Annu Rev Cell Dev Biol. 26, 261-283 (2010).
- Fernandez-Moreira, E., Helbig, J. H., Swanson, M. S. Membrane vesicles shed by Legionella pneumophila inhibit fusion of phagosomes with lysosomes. Infect Immun. 74 (6), 3285-3295 (2006).
- Galka, F., et al. Proteomic characterization of the whole secretome of Legionella pneumophila and functional analysis of outer membrane vesicles. Infect Immun. 76 (5), 1825-1836 (2008).
- Jager, J., et al. Human lung tissue explants reveal novel interactions during Legionella pneumophila infections. Infect Immun. 82 (1), 275-285 (2014).
- Ellis, T. N., Leiman, S. A., Kuehn, M. J. Naturally produced outer membrane vesicles from Pseudomonas aeruginosa elicit a potent innate immune response via combined sensing of both lipopolysaccharide and protein components. Infect Immun. 78 (9), 3822-3831 (2010).
- Jager, J., Keese, S., Roessle, M., Steinert, M., Schromm, A. B. Fusion of Legionella pneumophila outer membrane vesicles with eukaryotic membrane systems is a mechanism to deliver pathogen factors to host cell membranes. Cell Microbiol. , (2014).
- Jung, A. L., et al. Legionella pneumophila-Derived Outer Membrane Vesicles Promote Bacterial Replication in Macrophages. PLoS Pathog. 12 (4), (2016).
- Jiang, Y., Kong, Q., Roland, K. L., Curtiss, R. Membrane vesicles of Clostridium perfringens type A strains induce innate and adaptive immunity. Int J Med Microbiol. 304 (3-4), 431-443 (2014).
- Whitmire, W. M., Garon, C. F. Specific and nonspecific responses of murine B cells to membrane blebs of Borrelia burgdorferi. Infect Immun. 61 (4), 1460-1467 (1993).
- Ismail, S., Hampton, M. B., Keenan, J. I. Helicobacter pylori outer membrane vesicles modulate proliferation and interleukin-8 production by gastric epithelial cells. Infect Immun. 71 (10), 5670-5675 (2003).
- Jager, J., Steinert, M. Enrichment of outer membrane vesicles shed by Legionella pneumophila. Methods Mol Biol. 954, 225-230 (2013).
- Park, E. K., et al. Optimized THP-1 differentiation is required for the detection of responses to weak stimuli. Inflamm Res. 56 (1), 45-50 (2007).
- Casson, C. N., et al. Human caspase-4 mediates noncanonical inflammasome activation against gram-negative bacterial pathogens. Proc Natl Acad Sci U S A. 112 (21), 6688-6693 (2015).
- Molofsky, A. B., Shetron-Rama, L. M., Swanson, M. S. Components of the Legionella pneumophila flagellar regulon contribute to multiple virulence traits, including lysosome avoidance and macrophage death. Infect Immun. 73 (9), 5720-5734 (2005).
- Isaac, D. T., Laguna, R. K., Valtz, N., Isberg, R. R. MavN is a Legionella pneumophila vacuole-associated protein required for efficient iron acquisition during intracellular growth. Proc Natl Acad Sci U S A. 112 (37), 5208-5217 (2015).
- Zhu, W., et al. Sensing cytosolic RpsL by macrophages induces lysosomal cell death and termination of bacterial infection. PLoS Pathog. 11 (3), 1004704 (2015).
- Chutkan, H., Macdonald, I., Manning, A., Kuehn, M. J. Quantitative and qualitative preparations of bacterial outer membrane vesicles. Methods Mol Biol. 966, 259-272 (2013).
- Macdonald, I. A., Kuehn, M. J. Stress-induced outer membrane vesicle production by Pseudomonas aeruginosa. J Bacteriol. 195 (13), 2971-2981 (2013).
- Klimentova, J., Stulik, J. Methods of isolation and purification of outer membrane vesicles from gram-negative bacteria. Microbiol Res. 170, 1-9 (2015).
- . Novartis.com Available from: https://www.novartis.com/news/media-releases/novartis-bexsero%C2%AE-vaccine-approved-fda-prevention-meningitis-b-leading-cause (2016)
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved