
Method Article
変動アッセイを用いて微生物突然変異率を測定する
要約
ここで、フェノールトマーカーを用いて変動アッセイを行い、微生物変異率を推定するプロトコルが提示される。このプロトコルにより、研究者は多様な微生物や環境における突然変異をアッセイし、遺伝子型や生態学的文脈が自発的な突然変異率に与える影響を判断することができます。
要約
リライクアッセイは、液体環境で増殖する微生物の変異率を推定するために広く使用されています。多くの培養物は、それぞれ数千個の細胞を接種し、それぞれがフェノタイプと言える選択的マーカーに敏感である。これらの並列培養は、フェチピックマーカーがない場合、多くの世代のために成長する。培養物のサブセットは、突然変異の危険性がある細胞の総数(すなわち、成長期間終了時の母集団サイズ、またはNt)を推定するために使用される。残りの培養物は、選択的寒天にめっきされる。次に、並列培養における観察された耐性変異体の分布を使用して、数学モデルを使用して、予想される突然変異事象の数mを推定します。mをNtで除算すると、世代当たりの軌跡あたりの突然変異率の推定値が示される。アッセイには、選択された表現型マーカー、選択された並列培養量、および選択的寒天の表面がインキュベーションの前に完全に乾燥していることを確認するという3つの重要な側面があります。アッセイは比較的安価であり、標準的な実験装置のみを必要とします。また、突然変異の蓄積や単一細胞アッセイなどの代替アプローチよりも手間もかではありません。アッセイは、多くの世代を急速に通過する生物に作用し、マーカーと細胞死のフィットネス効果に関する仮定に依存します。しかし、最近開発されたツールと理論的研究は、これらの問題が分析的に対処できることを意味します。アッセイは、単離またはコミュニティで成長する異なる遺伝子型を有する細胞における異なる表現型マーカーの突然変異速度推定を可能にする。複数のアッセイを並行して行うことで、生物の環境文脈が自発的突然変異率にどのような影響を与えるかを研究するために、アッセイを用いることができ、抗菌性、発がん、老化、進化を理解する上で極めて重要です。
概要
1901年にオランダの植物学者ヒューゴ・ド・フリースは、用語突然変異1を造語しました.26年後、ヘルマン・ジョセフ・ミュラーがX線2の変異原性作用を発見したとき、突然変異はすでに進化の原動力の一つとして認識されていました。しかし、突然変異の性質は明らかではなかった。突然変異が自発的に出現するのか(すなわち、自発的な突然変異)、または選択に応答して(すなわち、誘導突然変異)、突然変異事象を観察する方法が必要であったかという根本的な疑問に答えるためには、このような方法は、細胞分裂当たりの予想数の突然変異、または既に突然変異率3、4として知られていたものを測定するであろう。

図1:96の深井戸板に微生物株を用いて変動アッセイを行う方法の概略図。(A)5 つの異なる環境 (「赤」、「青」、「緑」、「紫」、「オレンジ」アッセイ) を含む 50 mL チューブ内の細胞を接種および順応させます。(B) 96の深い井戸板に少数の感受性細胞を用いた並列培養物を調製する。「赤」アッセイには20の並列文化があり、「青」、「緑」、「紫」、「オレンジ」アッセイはすべて19の並列文化を持っています。96の深い井戸板上の平行培養物の位置はランダムである。ランダム化は、補足レイアウトジェネレータ.Rスクリプトまたは他のツールを使用して行うことができます。右上のレイアウトはランダム化の結果です。(C) 96の深い井戸プレートをインキュベートし、細胞が分裂し、自発的に突然変異することを可能にする。深井戸A1、B1、C1、E1、F1、およびG1からの6つの培養物は、変異体の数が変動する方法を示す:4、0、2、2、1、および4赤い細胞は、それぞれ第3細胞分裂後に。突然変異体の数は、自発的突然変異の数が異なるだけでなく(最初の赤い細胞で示される0、1、または2)、培養サイクル中に抵抗性突然変異が自発的に出現する場合に重要であるため(細胞分裂1、2、または3)。(D)96深井戸板のインキュベーション後、変異体の数は81平行培養物をめっきすることによって決定される。レイアウト上、これらは太字のエッジのない円です。全体の平行培養は、選択的寒天を含む6ウェルプレートの1つのウェルにめっきされる。(E)残りの15種類の培養物を希釈し、非選択的寒天上にめっきして、細胞の平均数(Nt)を決定する。レイアウト上では、これらは Ntwells としてラベル付けされ、太字のエッジを持っています。各アッセイについてNtは、3つの並列培養にわたって平均される。右下には、深い井戸D1(「緑」アッセイの一部)で成長した希釈培養物の25 CfUsを有する非選択的寒天プレートを含むペトリ皿があります。(F)選択的6ウェルプレートのインキュベーション後に観察された変異体の数と予想される突然変異事象数をカウントし、m、最大尤度推定器を用いて推定した。(G)突然変異の数、m、及びアッセイ当たりの細胞数の両方を知って、Nt、変異率はm/Ntと推定した。この図の大きなバージョンを表示するには、ここをクリックしてください。
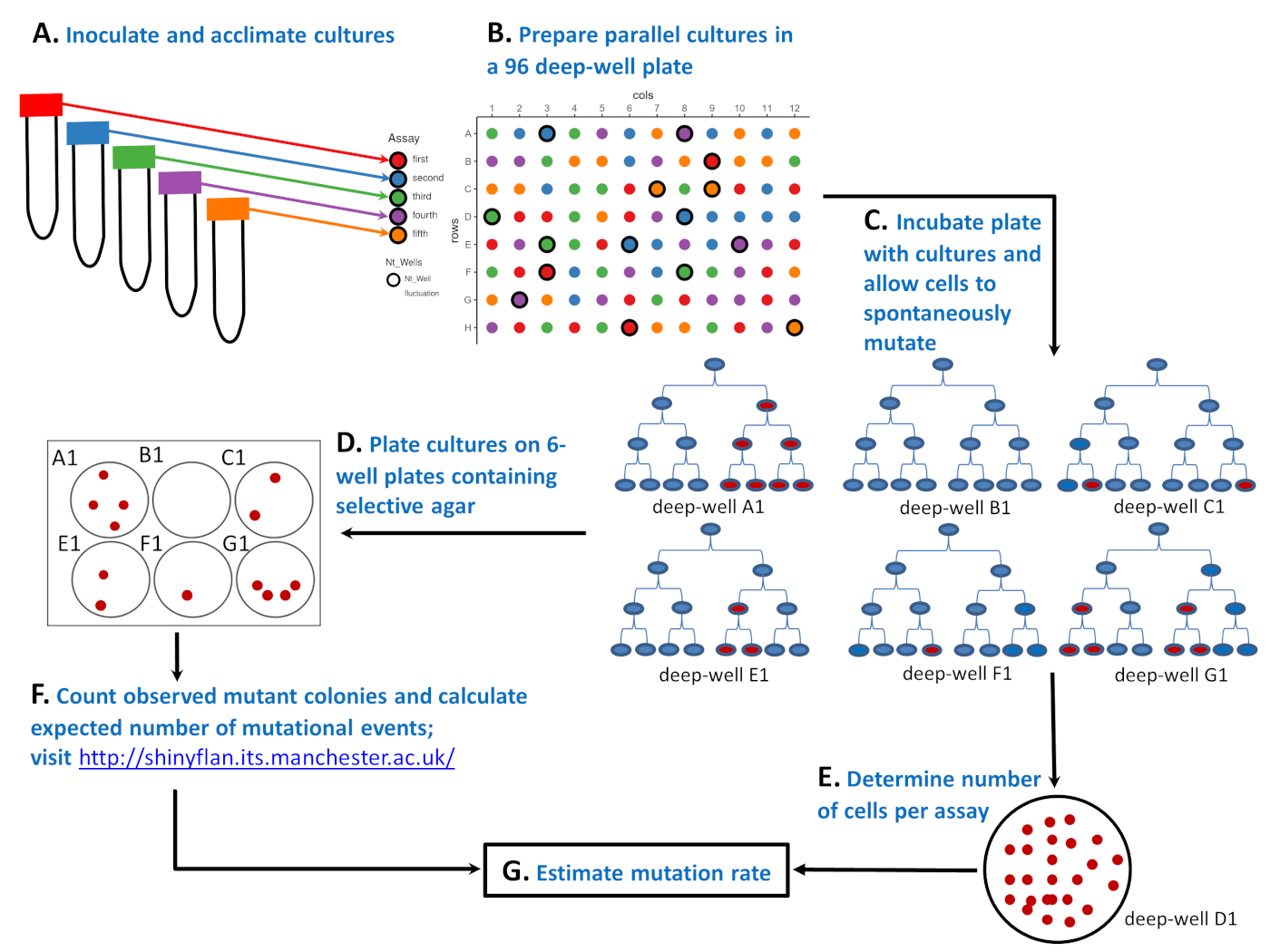{kind=link}
1943年のサルバドール・ルリアとマックス・デルブリュックは、変動アッセイ5でこの問題に対する独創的な解決策を提供しました(図1参照)。アッセイは、少数の微生物細胞で開始される複数の集団(名前付き並列培養)から始まります(図1A,B)。良性の非選択的環境で増殖した後(図1C)、耐性変異を有する細胞のみが生存し、コロニーを産生することができる選択的マーカー(ファージ、抗生物質など)を含むプレート上に並列培養物が転写される(図1D)。主な期待は、抵抗突然変異が誘発される場合、突然変異を運ぶ細胞の数は、平均が分散と等しい異なる集団間で分布する必要があるということです。ルリアとデルブリュックが変動アッセイで見つけたのは、突然変異体の数が急激に変動し、異なる集団間の突然変異体数の分散が平均よりもかなり大きかったことです。ルリアとデルブリュックはそれによって突然変異が自発的であることを実証した。その結果、DNAが複製されるたびに突然変異が自発的に出現し、突然変異の数は集団の増殖中に突然変異がいつ起こるのかによって異なる。図 1Cを参照してください。集団A1、E1、およびF1は1つの単一突然変異(最初の赤い細胞)を経験したが、培養サイクル中に様々な時点で単一の突然変異が自発的に出現するため、集団は非常に異なる数の突然変異(それぞれ4、2、1)で終わった。一方、集団C1とG1は、1つではなく2つの突然変異事象を経験したにもかかわらず、E1およびA1と同じ数の観測変異体で終わった。集団間で観察された突然変異体の変動は、アッセイに名前を与えただけでなく、変異頻度(すなわち、変異細胞の割合)が突然変異率の不十分な指標であることを示した。
変動アッセイの全体的な目標は、特定の液体環境で増殖する細菌または他の単細胞生物の特定の遺伝子型の自発的突然変異率を推定することである。変動アッセイは、微生物突然変異率の環境依存性を研究するための最も適切なツールであり、迅速かつ安価な突然変異率推定を可能にします。最大深度シーケンシング6、母集団シーケンシング7、突然変異蓄積実験8、または子孫のゲノム配列と親9のゲノム配列の比較など、突然変異率推定に対する代替アプローチは、はるかに手間がかかり、したがって環境依存を検出する可能性に適しに欠ける。しかし、突然変異の生成および修復の動的な側面は、変動アッセイまたは上記の突然変異率をアッセイする方法のいずれにもほとんどアクセスできない。突然変異の数が時間、空間、または集団内の個々の細胞間でどのように変化するかを研究するには、単一細胞が11、12に近づく必要があり、これは変動アッセイよりも手間がかけることに加えて、高度に専門的なスキルと機器を必要とします。
実際には、変動アッセイは、そのマーカーの選択を欠く環境で発生する突然変異のために表現型マーカーを得る細胞をカウントしています。数百の公開アッセイ10のメタ分析は、1943年のアッセイ開始以来、少なくとも39種類の表現型マーカーが使用されていることを示している。変動アッセイは、実験室、臨床、非ミューター、および寛容な環境で成長する突然変異株間の突然変異率の平均と環境依存性を比較するために使用することができます。アッセイは、最小限または豊かな環境で成長する異なる遺伝的背景を持つ細胞の突然変異速度推定を可能にします。このアッセイは、単一培養として増殖する集団に適しているだけでなく、細胞間相互作用が突然変異率11に及ぼす影響を研究するためにも使用できる。目的の株が第2の株と共培養され、中性マーカーが株を区別するために使用される場合、突然変異率は、同時に同じチューブ内の2つの株に対してアッセイすることができる。
変動アッセイは、自発的突然変異率が細胞の遺伝子型とその環境12の両方に依存し、それ自体が13を進化させる形質であることを明らかにした。ある特定の遺伝子型の突然変異率が環境とともに変化するたびに、変異率可塑性11と記載される。プラスチック突然変異率は、ストレス誘発性突然変異誘発(SIM)14に対して最も徹底的に対処されている。さらに、変動アッセイを用いて、細胞の集団が増殖する密度(典型的には、運搬能力におけるバッチ培養)は、細菌および単細胞真核生物間の突然変異率と密接に関連していることが最近示されている。世代当たりのゲノム当たりの突然変異率は、密度の高い集団で23倍10、11倍も減少する。この密度関連変異率可塑性(DAMP)は、クォーラムセンシングシステム15に依存し、SIM16とは独立して作用することができる。
ここでは、グルコース最小限の媒体環境における抗生物質リファンピシンに対する耐性を得る大腸菌株K-12の研究に用いられる変動アッセイに関する詳細なプロトコルが提示される。しかし、このプロトコルは、単に突然変異の培養条件および表現型マーカーを変更することによって、多種多様な微生物を研究するために利用できる基本的なテンプレートとして見るべきである。プロトコルは、その開始から進化しました5,17,18,19,20,21,22,23,24,25,26,27,28,29広い範囲の微生物および癌細胞30での使用を通じて変更されました。微生物突然変異率10、11、16の環境依存性を適切にテストするために不可欠であったスループット。ここで説明するプロトコルは、文献で既によく議論されている変動アッセイの方法論的および分析的問題、特に耐性突然変異31のフィットネス効果、表現型遅延32、細胞死33、および突然変異率26、34を推定するために利用可能な種々のアルゴリズムの適合性をカバーするものではない。これは、例えば、フィットネス効果の環境依存性が突然変異率推定値35の誤った変動を引き起こす可能性がある場合に重要である。しかし、ここで使用する分析ツールは、突然変異体フィットネスと細胞死の変動に対処できることに注意してください。ノートと議論で取り上げられているように、同じ環境依存性フィットネス効果を持つ可能性が低い複数の表現型マーカーを考慮することをお勧めします。このプロトコルは、人々が微生物株および環境の多様性における突然変異率の環境依存性を日常的にアッセイすることを可能にする。異なる環境における突然変異のアッセイはまだ十分にテストされておらず、一旦集団密度が考慮されると、変動アッセイは突然変異率10のより正確な推定値を与えることができる。このプロトコルは、進化、発癌、老化、抗菌性を理解するために不可欠な突然変異率を支えるメカニズムを理解するために必要なように、より多くの変動アッセイを行うことを可能にします。
プロトコル
1. 1日目:文化の接種と順応
- 液体リゾゲニースープの3 mLを接種し(LB、補足表1を参照)、大腸菌MG1655グリセロールストック(18%グリセロール、-80°)からの氷の擦り傷を有する。LB培養を120rpmで37°Cで~7時間振る。
注:この実験では、LBで増殖する大腸菌K12 MG1655が用いられるが、このアッセイは、任意の大腸菌株または他の培養可能な微生物種で行うことができる。インキュベーション温度、インキュベーション時間、および成長媒体の栄養レベルはすべて、種または株によって変動する可能性がある。 - 生理線溶液を用いて培養液2,000倍希釈する。希釈溶液の100 μLを3つの50 mLスクリューキャップ円錐底ポリマーチューブ(50 mLチューブ)に10mLの液体デイビス最小培地(DM、補足表1を参照)を加え、それぞれ80mg/L、125 mg/L、または250mg/Lのグルコースを含みます。これは、突然変異率が推定されるのと同じ培地(すなわち、環境)である。培養物を37°Cで一晩120rpmで振る。
注:メディアの選択は、種、ひずみ、または研究の質問によって変動の対象となります。
2. 2日目:並列培養における変異体の生成
- まず、細菌が培養される環境を準備する。常に必要以上に10%多く準備します(すなわち、20ml培養所は22mLを必要とします)。ブドウ糖の80 mg/Lで22 mL、2)ブドウ糖の125 mg/LのDM、3)グルコースの250 mg/LのDM、4)グルコースの80 mg/LのDM、および5つの50 mLチューブで250 mg/Lのグルコースを有するDMを調製します。それぞれ GLC-80A、GLC-125、GLC-250A、GLC-80B、GLC-250Bとしてラベル付けしてください。
- 環境に対するイノクラを準備します。環境の 1 mL の接種に 1,000 -5,000 個のセルが含まれていることを確認します。これを行うには、次の手順に従います。
- 600 nmで一晩培養物の光学密度(OD)を測定する(ステップ1.2から)。
- グルコースを有するDMのmL当たり1,000〜5,000細胞の最終密度に達するために各一晩培養を希釈する。私たちの手の中で、2.2.1で測定されたODが0.3の場合、これは一晩培養物の100倍の希釈(生理食合い溶液中)を作り、22 mL環境にこの溶液の11μlを加えることを意味します。
- 並列カルチャを準備します。
注:このプロトコルは、3つの環境を使用して、5つの変動アッセイ(1つの96ウェルプレート上の最大妥当な数)を実行し、1つの96深いウェルプレートのために書かれています。経験によって、複数の深井戸プレートが並行して実行される場合があります。- 1つの96の深い井戸プレートのための平行培養のランダムなレイアウトを作成します。補足 R スクリプト LayoutGenerator.R (図1Bのレイアウトを参照) を使用できます。レイアウトに従って、96の深い井戸プレートに各平行培養を配置します。
注:LayoutGenerator.R を実行すると、最初のアッセイに 20 個の並列カルチャがあり、2 番目、3 番目、4 番目、および 5 番目のアッセイにはそれぞれ 19 個の並列カルチャが含まれます。 - 無作為化されたレイアウトに従って、接種したメディアを96の深い井戸プレートの各ウェルに転送します。
- テープで深い井戸板のふたを固定します。培養の成長は通気量に敏感であるため、蓋をしっかりと固定しないでください。
- プレート全体を蓋とテープで計量し、プレートを250 rpmで250rpmで37°Cで振ります。2 Lの蒸留水をインキュベーターに入れ、実験セット間の蒸発量を安定させる。
- 1つの96の深い井戸プレートのための平行培養のランダムなレイアウトを作成します。補足 R スクリプト LayoutGenerator.R (図1Bのレイアウトを参照) を使用できます。レイアウトに従って、96の深い井戸プレートに各平行培養を配置します。
- 非選択的テトラゾリウム(TA)寒天板上の各接種媒体の10μLをめっきして接種サイズを決定する(補足表1参照)。寒天表面が乾燥するまで無菌のL字型スプレッダーを使用してください。TA寒天プレートを37°Cで一晩ふくらまします。
注:TA寒天は、糖L-アラビノースと水溶性染料2,3,5-トリフェニルテトラゾリウムクロリドを含む豊富な寒天であり、これは酸化された形態では無色である。細菌が染料を減少させると、フォルマザンの形成により赤色に変わります。L-アラビノースを利用できないTA寒天上のコロニーは濃い赤色です。MG1655などの他の株はピンクがかった。色付きの細菌コロニーを見つけやすくするため、標準的なLB寒天ではなくTA寒天の使用が推奨され、コロニーの数え方がより信頼性が高く、より迅速になります。 - 6ウェルプレートにリファンピシンを含む選択的TA寒天を調製します。選択的TA寒天のピペット5mLを6ウェルプレートの各ウェルに入れます。TA寒天に添加する直前に抗生物質リファンピシンを準備します。
注:ヒロセリンをマーカーとして使用する場合は、寒天とL-アラビノースを添加したグルコース250mg/Lと2,3,5-トリフェニルテトラゾリウムを選択的寒天として使用してください。すなわち、TA寒天は、トリプトンと酵母エキス(TA寒天の必須成分の両方)がアミノ酸D-アラニンを含んでいるため、シアクロクリンに耐性のある細胞のみを選択しない。このアミノ酸は、シアクロクリンの抗菌効果を拮抗し、すべての細胞がコロニーを作ることができます。- 選択的な非選択的プレートと残りの非選択的プレートは、室温で暗い(例えば、箱の中)に残しておきます。
3. 3日目:選択的および非選択的寒天プレート上のめっき培養
- 非選択的寒天プレート上のコロニー形成単位(CFU)をカウントし、CFUに100を掛けてイノクラのサイズを決定します。並列培養の体積(1mL=1,000μL)とめっき体積(10μL)の比率です。
注:非選択的プレートが保存されている場合、冷蔵CFUは後でカウントすることができる。 - 24時間のインキュベーション後、深い井戸板全体を計量して蒸発量を決定する。これは10%前後である可能性が高い。
- 開始体積V[0h]=1,000μlとプレートの重量をマイクロリットルに変換し、成長培地の密度をmg/μlで測定して、マイクロリットルで24時間インキュベーション(V[24h])後の並列培養の平均体積を計算します。この研究では、1 mg/μl の密度を使用します。

- アッセイごとにランダムに選択された3つの培養物(合計15、図1Bのレイアウトで黒い円で強調表示)を標識されたマイクロ遠心管に移します。最終的な母集団サイズ (またはNt)を決定するために、後で使用するためにベンチに残します (手順 3.6 を参照)。
-
残りの81の並列培養物を深いウェルプレートからリファンピシンを含む選択的TA寒天にプレートします。ピペットは、96の深い井戸プレートから6ウェルプレートの1つのウェルに1つの並列培養全体をピペット。任意の6ウェルプレートに複数の変動アッセイからの並列培養が含まれていることを確認します。
- 選択的寒天プレートから蓋を取り出し、滅菌状態で覆い隠します。選択的TA寒天の表面上のすべての液体を乾燥させます。
注:寒天が完全に乾燥していることは非常に重要です。ただし、選択的TA寒天を過乾燥させないでください。
- 選択的寒天プレートから蓋を取り出し、滅菌状態で覆い隠します。選択的TA寒天の表面上のすべての液体を乾燥させます。
-
選択的TA寒天プレートが乾燥している間、数時間かかる場合があり、コロニー形成単位(CFU)を用いてステップ3.4で調製した15の培養物のNtを決定する。
- マイクロ遠心管から培養物を希釈してCFUを決定する。5つの10倍希釈ステップを使用して、各ステップで100°Lの培養物で900 μLの生理油溶液を混合およびボルテックスします。最後の希釈のプレート40μL(希釈係数105)上の非選択的TA寒天およびインキュベートプレートは37°Cで一晩蓋をする。
注:96ウェルプレートの希釈シリーズは、このステップの速度を高めることができるマルチチャンネルピペットで行うことができます。
- マイクロ遠心管から培養物を希釈してCFUを決定する。5つの10倍希釈ステップを使用して、各ステップで100°Lの培養物で900 μLの生理油溶液を混合およびボルテックスします。最後の希釈のプレート40μL(希釈係数105)上の非選択的TA寒天およびインキュベートプレートは37°Cで一晩蓋をする。
- 6ウェルプレート上のすべての井戸に培養液が含まれなくなったら、蓋を元に戻し、6ウェルプレートがすべて乾くまでベンチに蓋をして6ウェルプレートを置きます。すべてが乾いたら、プレートを37°Cで44-48時間インキュベートします。
注:他のマーカー(ナリジキシン酸、シクローセリン、ハイグロマイシンB、または5-FOA)の場合は、プレートを68〜72時間インキュベートする。インキュベーターの湿度が高いことを確認します。選択的寒天プレートが潜伏期間中に乾燥しないことが重要です。
4. 4日目:培養中の細胞数の決定
- 非選択的寒天プレート上のCFUをカウントします。CFUに希釈係数(105)を掛け、マイクロリットル(ステップ3.3を参照)とめっき体積(40°Lを参照)で計算された平均体積V[24時間]の比率を乗じて、培養中の生存細胞の数を推定します。

- 特定の環境で増殖する特定の遺伝子型のNtは、3つの培養物からのこれらの値の平均である。
5. 5日目:突然変異率の推定
-
選択的TA寒天プレート上の抗生物質に耐性のあるコロニーの数を数える(すなわち、2-3日の深井戸プレートの自発的な突然変異を通じて生じた耐性細胞の数)。特定のアッセイに対して観察された変異体数の並列培養物(例えば、16または17の値、ゼロカウントを含む)の分布を記録する。
注:選択的プレートが冷蔵保存されている場合、抗生物質に耐性のあるコロニーは後で数えることができる。- 各分布を使用して、Rパッケージフラン36を使用して、m の突然変異イベントの数を推定します。
注:同様の機能を持つRパッケージrSalvadorもあります29. - 1 つのアッセイの観測された突然変異体の分布を 1 つの列としてテキスト ファイルに保存します。
- 各分布を使用して、Rパッケージフラン36を使用して、m の突然変異イベントの数を推定します。
-
Shinyflanソフトウェア(http://shinyflan.its.manchester.ac.uk/)を使用して、以下の詳細としてmを推定します。
- タブ仮説テストのデフォルト値(すなわち、未知のフィットネスがチェックされ、推定方法=最大尤度(ML)、ミュータント寿命=指数(LDモデル)の分布、Winsorパラメータ= 1,024、突然変異数とフィットネス=1、信頼水準= 0.95、クラス数と最大値= 100)。
- [参照]をクリックし、観測された突然変異体の分布を含むテキスト ファイルを選択します。最初に補足データ ファイルを使用してみます。
- ファイルをアップロードしたら、[テストの実行]をクリックします。[テストの結果] の右側にある[1 サンプル ML-テスト(LD モデル)] で、[変異番号]を見つけます。これはmであり、予期される突然変異イベントの数です。
- 突然変異数の95パーセント信頼区間の下でmの上限と下限を見つける。
- mおよびNt(CFUによって決定される)が利用可能になったら、特定の環境における特定の遺伝子型の突然変異率を.
 として推定する。mの上限と下限をNtで除算して、突然変異率に信頼区間を生成するのと同じ方法で (NB これはNtの不確実性を考慮しません)。
として推定する。mの上限と下限をNtで除算して、突然変異率に信頼区間を生成するのと同じ方法で (NB これはNtの不確実性を考慮しません)。
注:結果は、世代ごとのrpoB軌跡当座あたりの突然変異率として提示される。塩基対当たりの突然変異率は、rpoB遺伝子座当たりの突然変異率を79で割ることによって生成され、これはrpoB遺伝子内の何点突然変異がリファンピシン37に対する耐性を付与するかについての我々の現在の知見である。ヌクレオチド当たりの突然変異率に染色体の大きさ(大腸菌K-12 MG1655=4,639,675 bp)を掛けると、ゲノム当たりの変異率が与える。 - 残りの 4 つの変動アッセイで手順 5.1-5.3 を繰り返します。
結果
結果は、3人の異なる個々の研究者によって4つの異なる週に報告されたプロトコルで収集され、毎週96の深い井戸プレートを使用して5つの突然変異率推定値を生成しました。大腸菌K-12 MG1655(図2A、プロトコルで使用される場合)で15個の変異率推定値(±95%信頼区間、CI)を生成した3つの96個の深井戸プレート(図2B)で、1つの96深いウェルプレートを大腸菌K-12 BW25113ΔmutTの5つの推定値(±95%CI)に使用しました。図 2では、推定mの四分位範囲の中央値精度は 17.5% (1.00%−28.9%,n = 20) です。精度は、σm/m x 100%として計算される予想数の突然変異事象(m)の変動の係数です(σは前に示したように計算されます)。図 2の生データは、補足データ ファイル"Krasovec_etal_JoVE_data.csv" で使用できます。補足データ ファイルには、図 2を生成するための R スクリプトが付属しています。細菌株の詳細については補足表 2と、データ・ファイル内の列について説明する補足表 3も参照してください。リファンピシンとナリジキシン酸で取得したデータポイントは、クラショベックら10で以前に公開され、最初に公開されたときと同じIDを持っています。
図2では、3つの異なるフェノチピックマーカー、シクロセリン、リファンピシン、およびナリジキシン酸のMG1655突然変異率が示されている。ここで、プロトコルで説明したように、グルコースの80、125、および250mg/Lを有するデイビス最小培地で突然変異率を評価した。1つのケースでは、グルコースの1,000 mg/Lを使用しました(図1B)。実際には、任意の初期グルコース濃度を使用することができる。予想通り、シクロセリン耐性の突然変異率は高く、ナリジキシン酸耐性は最も低く、リファンピシン耐性の割合は真ん中であった。これは、これらの3つの抵抗の既知の目標サイズに従い、最大はシクロセリン用で、ナリジキシン酸の最小です。議論と図3は、目標サイズ、並列培養量、環境中の栄養素のレベルを利用して、変動アッセイによる微生物突然変異率の測定を最適化する方法について詳しく説明します。
変動アッセイは、どの株が構成型突然変異器であり、どの株が正常な突然変異率を有するかを明確に示す。ΔmutT株は、MG1655としてナリジキシン酸耐性(図2B)に対して約50倍高い突然変異率を有していた(図2A)。しかし、異なる環境における特定の遺伝子型の突然変異率を決定するには、1つの表現型マーカーのみを使用して、環境当たりの遺伝子型当たり少なくとも5回の反復を行うことをお勧めします。この種の実験の分析に以前使用した統計的手法の詳細については、クラショヴェツェらの補足情報を参照してください。

図2:大腸菌集団における変動アッセイによって推定される突然変異率の代表数値。(A)野生型MG1655(円)で報告されたプロトコルによって決定される突然変異率。図は、リファンピシン(水色の円)、ナリジキシン酸(赤丸)、およびシクロセリン(濃い青色の円)耐性の存在下での突然変異率を示す。ナリジキサン酸の場合は、10 mLの大きな培養量が使用されました(補足データファイルを参照)。リファンピシンおよびナリジキシン酸のデータは、図2aからクラショベックら10で再プロットされる。(B)慶應義塾39Δ変異変異体(赤三角形)におけるナリジキシン酸耐性に対する変異率。突然変異率軸の対数スケールに注意してください。誤差範囲 = プロトコルで説明されているように計算された信頼区間は 95% です。データは図4 aからクラショベックら10で再プロットされる。この図の大きなバージョンを表示するには、ここをクリックしてください。
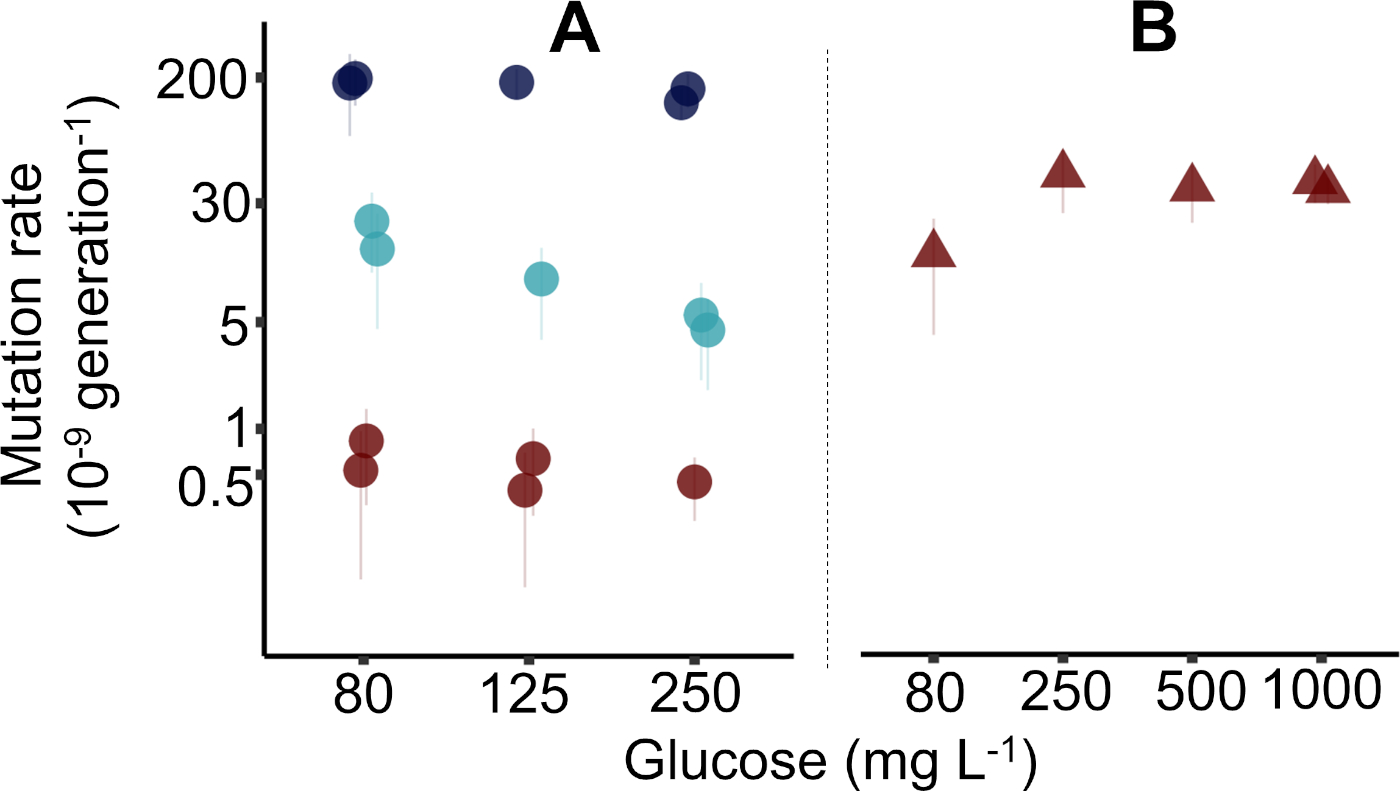{kind=link}

図3:変動アッセイのトラブルシューティング用フローチャートフローチャートは上から続き、黄色のダイヤモンドの3つの質問に順番に対処し、得られた緑色のボックスの内容に従ってプロトコルを調整し、赤い楕円に示すようにプロトコルを実装します。トラブルシューティングの方法の詳細については、説明の最初の 3 つの段落を参照してください。この図の大きなバージョンを表示するには、ここをクリックしてください。
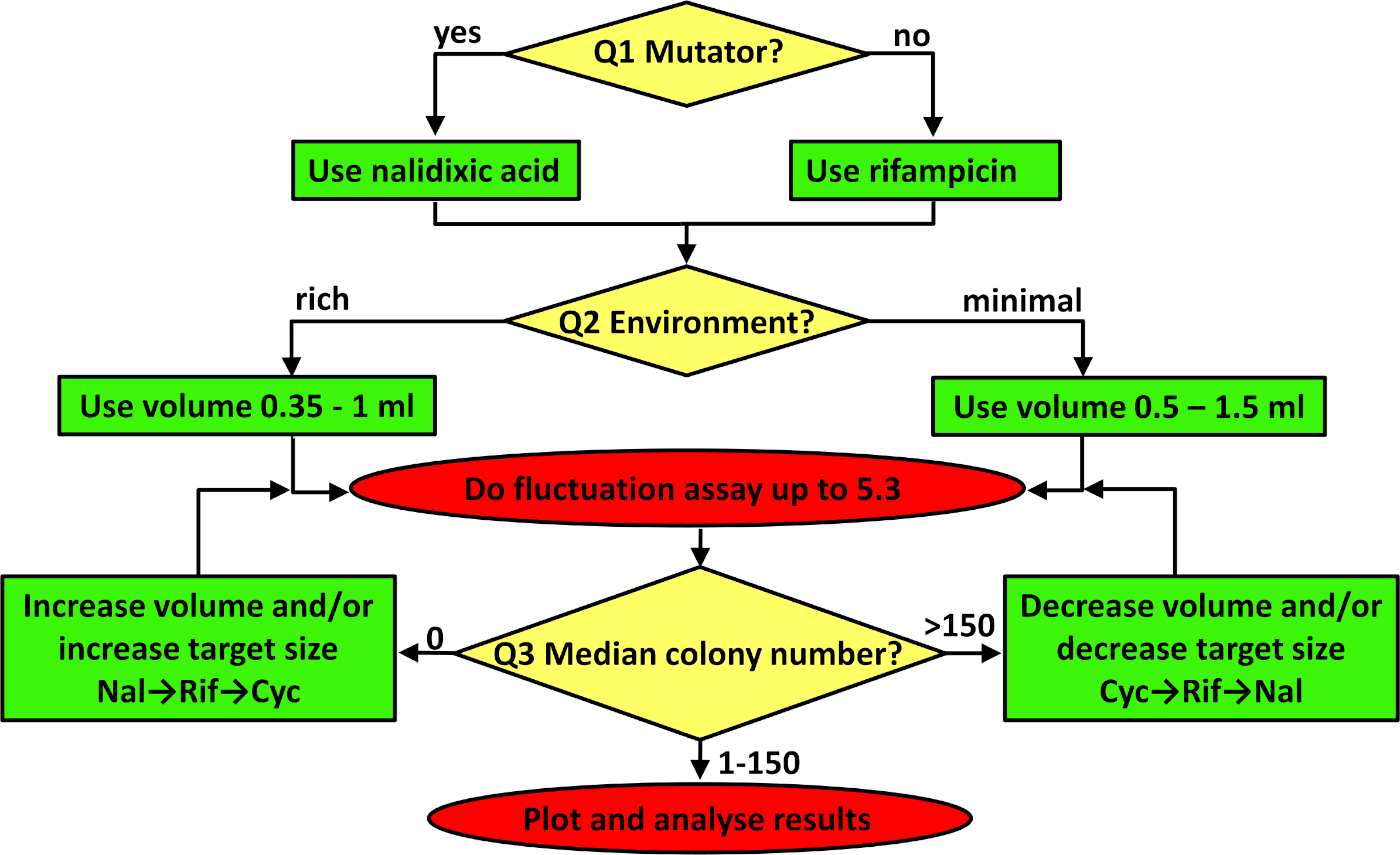{kind=link}
補足表 1: プロトコルで使用されるメディアの数式。ここをクリックしてこのテーブルを表示してください (右クリックしてダウンロードしてください)。
補足表2:細菌株。ここをクリックしてこのテーブルを表示してください (右クリックしてダウンロードしてください)。
補足表 3: 生データ ファイル Krasovec_etal_JoVE_data.csv の列の詳細な説明。ここをクリックしてこのテーブルを表示してください (右クリックしてダウンロードしてください)。
補足データ ファイル。これらのファイルをダウンロードするには、ここをクリックしてください。
ディスカッション
突然変異率の任意の推定は、研究34内および研究間の再現性および再現性を確保するために達成される精度を最大化する必要がある。変動アッセイには、3つの重要な考慮事項があります。最初の 2 つは、指定されたプロトコルで設定されていますが、プロトコルが異なる株または環境で動作するように適合している場合は、トラブルシューティングが必要です (図 3参照)。まず、適切なフェチピックマーカーを選択する。細菌の場合、rpoBまたはgyrAの2つのマーカーロチのいずれかで速度を推定することをお勧めします, 抗生物質リファンピシンとナリジキシン酸に対する耐性をそれぞれ与えます.これら2つの遺伝子座における抗生物質耐性に対する突然変異の標的サイズは異なる。リファンピシン37およびナリジキシン酸40に対する耐性をそれぞれ付与する79および20のユニークな突然変異がある。実際には、これは平均して、リファンピシン耐性変異体がより頻繁に観察されることを意味する。したがって、最初に答える必要がある質問 (図 3の Q1) は、ひずみがミューテーションターであるかどうかです。多くの観察された変異コロニーが予想される構成変異体を研究する場合、より小さい標的サイズ(例えば、ナリジキシング酸)を有するマーカーを使用する方が良い。図2Bを参照し、構成変異体大腸菌K-12 BW25113ΔmutTの変異率を、ナリジキシング酸をマーカーとして用いて推定した。野生型(すなわち、通常)突然変異率を有する非変異体細菌株を用いて作業する場合、リファンピシンはより良い選択である(図2Aを参照)。何らかの理由でより多くの観察された突然変異体が必要な場合、関連するマーカーは、ヒクロセリンに対する耐性である。このマーカーの場合、抵抗突然変異は10個以上の遺伝子41で出現する可能性があり、標的サイズがリファンピシンよりもさらに大きいことを意味する。酵母と古素を研究する場合は、25Sリボソームタンパク質とURA3の突然変異率を推定し、それぞれハイグロマイシンBおよび5-フルオロオロオロ酸(5-FOA)に対する耐性を与えることをお勧めします。
2 番目に重要な考慮事項は、並列カルチャのボリュームです。使用するボリュームは、実際に観測された突然変異体の数によって異なります。観測された突然変異体の予想数は、選択したフェオチピックマーカーの目標サイズ、突然変異を修復して回避する株の能力(平均突然変異率)、および使用される媒体と培養体積の両方によって影響を受ける環境の運搬能力の影響を受ける。並行培養物にリファンピシンに耐性のある変異コロニーが含まれていない場合は、シクロセリンを使用するか、めっき細胞の数を増やす必要があります。これは、最小限の培地にブドウ糖を追加するか、より豊かな(または完全な)環境で細胞を成長させることによって達成することができる。しかしながら、多くの場合、このような集団密度の増加は突然変異率の低下に関連しており、その結果、限られた結果、存在する場合には、10を観察する変異コロニーの数の増加が生じる。栄養素を増やすことが解決策でない場合は、各並行培養の体積を増やすことで細胞数を増やすことが選択肢である。炭素とエネルギーの唯一の供給源として砂糖を含む最小限の塩が含まれている場合(すなわち、図3のQ2に対する答えは「最小媒体」)、0.5~1.5mLの体積を使用する必要があります。環境が豊富な場合、並列カルチャのボリュームは 0.35-1 mL の間である必要があります。最後の質問は、耐性コロニーの中央値に関するものです。あまりにも少ない突然変異コロニーが観察され(すなわち、図3のQ3に対する答えは0である)、環境が改変されない場合は、並列培養物の量を増やすか、より大きな標的サイズ(例えば、cycloserine)を有する抗生物質を使用すべきである。一方、すべての選択プレートで多くの変異コロニーが観察される場合(1プレートあたり〜150以上、図3を参照)、めっき細胞の数を減らすべきである。
ボリュームが選択されると、1つの96の深い井戸板上のすべての平行培養物が同じ体積を持つことをお勧めします。これは、プレートの重量から並列培養の実際の体積のより正確な決定を可能にします。特定の遺伝子型の突然変異率を異なる環境間で比較する場合、すべての環境で同じ量の並列培養を使用するのが最善です。野生型(すなわち、正常)突然変異率または他の表現型マーカーの推定にナリジキシング酸を使用する場合は、ナリジキシング酸よりもさらに小さい目標サイズを有するが、体積をさらに増加させる必要がある。1つのオプションは、最大15 mLの容積を有する50 mLチューブで並列培養を行うことです。例えば、10mLの並列培養物を、大腸菌K-12 MG1655変異率をナリジキシン酸に推定する際に50mLチューブで調製した(図2A参照)。平行な10 mL培養物を選択的TA寒天にめっきし、標準の90mmペトリ皿の代わりに大きな150mmプレートに注いだ。50 mLチューブで並列培養を調製する欠点は、96の深い井戸板のアッシャリ突然変異率に比べてスループットがかなり低いことです。1 つの解決策は、並列カルチャの数を減らすことです。ただし、これは mの推定値の精度に影響を与えます。14-17並列培養(図2で行われたように)で観察された変異体の分布を得ることは、固体スループットと許容可能な精度レベル26の20%との間の良好なバランスである。中央値の精度レベルが 17.5% は、はるかに大きなデータセット10から計算された四分位範囲が 16.4% (5.7%-38.9% n = 580) の中央値精度に似ています。したがって、96の深いウェルプレートまたは50mLチューブで並列培養を調製する場合、観察された変異体の分布は、少なくとも14の並列培養で得られることをお勧めします。異なる環境で突然変異率が推定される場合は、1つのプレート上のすべての96の並列培養が同じ環境で成長するマルチプレート実験を行って、精度レベルをテストすることをお勧めします。また、並列培養物を調製する際には、接種菌に耐性細胞が存在する可能性が低いため、イノクラに含まれる細胞数が少ないほど重要です。既存の耐性変異体は、数が増加し、選択的プレート上に芝生を作成し、突然変異率の推定が不可能であるため、接種体では望まれていません。例えば、ほとんどの非変異体大腸菌集団において、リファンピシン耐性に対する突然変異率は〜10−8の順である。したがって、既存の耐性変異体で培養物を接種しないように、108細胞未満(例えば、103−104細胞)で接種しなければならない。 最後の重要なステップは、選択的寒天プレートがインキュベートされる前に、選択的寒天上の表面が完全に乾燥していることを確認することです。たとえば、6 ウェルプレートを使用し、並列培養の初期体積が 1 mL の場合、スプレッダは使用できません。プレートは、表面の液体を乾燥させるために滅菌条件で覆い隠しておく必要があります。この時間は、周囲の条件とプレートの状態に応じて、非常に可変にすることができます。この時間は最小限に抑える必要がありますが、最大時間を使用できます。
変動アッセイには固有の制約があります。それは、ゲノムの小さなサブセットでのみ突然変異の表現マーカーをアッセイする。したがって、アッセイは、十分な数の世代を経て、速度を全く推定するのに十分な突然変異を観察する大きな集団を必要とする。これは、変動アッセイは、細菌、パン酵母42、または液体培養哺乳動物細胞30のような多数の世代を急速に通過することができる生物に対してのみ使用できることを意味する。また、突然変異は、特定の細胞の特定の生化学的状況で発生する稀な事象である。変動アッセイが時間の経過とむほどに大量の細胞に見えるということは、それらの状況が大きく異なる可能性があることを意味します。このアッセイを用いて、ラグ相から早期および後期指数位相、そして最終的に静止相までの特定の集団の突然変異率の進行を研究することは困難である。集団内の単一細胞間の突然変異率の分化は、変動アッセイから完全に隠されています。単細胞突然変異ダイナミクスは、DNA修復タンパク質MutS43の単一分子追跡または蓄積されたMutLタンパク質44の病巣をカウントすることによって研究することができる。ハイスループットシーケンシングの最近の進歩はまた、親子孫トリオ9、45および多世代血統46からの突然変異率を直接推定することを可能にした。このような方法論的進歩は、単一世代内で起こる突然変異の直接カウントを可能にし始めている。しかし、この直接的なアプローチには、蛍光顕微鏡、マイクロ流体、全ゲノムシーケンシングなどの高価で最先端の技術が必要です。一方、変動アッセイは比較的安価であり、標準的な実験装置のみが必要である。より多くの変動アッセイを行うことは、より直接的な単一細胞アプローチでテストされる可能性のある新しい仮説の生成を容易にします。
突然変異の研究には長年の関心があるので、変動アッセイは広く用いられている方法のままである可能性が高い。過去4年間(2015-2018年)のルリアとデルブリュック5によるセミナルペーパーの引用数は、すべてこの論文の引用のトップ5に入っています。しかし、変動アッセイを適切に行うために必要な精密な手作業が多いため、ほとんどの研究はほんの一握りの変動アッセイしか行っていない。しかしながら、これは突然変異率の環境依存性を明らかにするには不十分である。本論文で説明したように、マルチウェルプレートを用いて変動アッセイを合理化することにより、現在可能な最大スループットは、ここで説明するように11個の深井戸プレート(55変動アッセイ)を並行して行う。1日ずつずらした2組の変動アッセイを並行して実行することで、週に最大110のアッセイを行うことができます。スループットの別のステップの変化は、与えられた純粋に手動プロトコルから変動アッセイの様々なステップを自動化することによってまだ可能であり得る。また、突然変異率の環境依存性を研究するためには、集団密度を考慮する必要がある。前の結果10は、突然変異率に影響を与える既知の因子が考慮されると、母集団密度を制御することで、突然変異率推定値の変動を90%以上減少させることができることを示している。密度を制御するには、集団密度の決定に使用される方法とは別に、Nt (突然変異率の推定に使用) を決定することをお勧めします。細菌において、Ntは、例えば、ATPベースの発光アッセイ10を用いてCFUおよび密度によって決定することができる。
生物の生態学的文脈が自発的突然変異率に及ぼす影響を調べるとき、高いスループットと制御密度は両方とも不可欠です。突然変異率可塑性の存在を知ることは重要であるが、その原因と効果を理解することは、変異率可塑性をより広い生物学的文脈に組み込む場合に満たす必要がある重要な課題である。変動アッセイは、結果が急速に得られ、アッセイは他の方法に比べて安価であるため、多くの仮説をテストするために使用できる素晴らしいツールです。ステージは、例えば、細菌群集および微生物叢における突然変異率の環境依存性を研究するために設定される。変動アッセイを共培養に適応させることで、株が小分子を介して互いの突然変異率に影響を与えるという仮説をテストできる。共培養で何千もの変動アッセイを行うことで、株が互いの突然変異率を変更する能力と、他の人によって突然変異率を改変する感受性の両方で変化するかどうかを判断できます。おそらく、突然変異速度操作に対する感受性における株間の変動は、特定の遺伝的変異に起因する。これは、抗菌性がどのように出現するかなど、広い重要性の例では、複雑なコミュニティにおける進化の仕組みに関する我々の見解を変えるかもしれない。
開示事項
著者たちは何も開示する必要はない。
謝辞
RKはBB/M020975/1とマンチェスター大学生物科学部の支援を受けました。HR は BB/J014478/1 でサポートされています。GGはBBSRCドクタートレーニングパートナーシップBB/M011208/1の支援を受けています。DRGはUKRI賞番号MR/R024936/1によってサポートされました。
資料
| Name | Company | Catalog Number | Comments |
| 1.5 mL Microcentrifuge tubes | Starlab International GmbH | S1615-5550 | |
| 2,3,5-Triphenyltetrazolium chloride | Sigma-Aldrich | T8877-10g | |
| 6-well plates | Greiner Bio-One | REF 657102 | |
| 90 mm Petri Dishes Triple Vented | ThermoFisher Scientific | REF 120189 | |
| 96 deep-well plate (Masterblock 2 mL) | Greiner Bio-One | REF 780270 | |
| Ammonium sulfate | Fisher Chemical | A/6440/53 | |
| Bacto Agar | Becton, Dickinson and Company | REF 214010 | |
| Bacto yeast extract | Becton, Dickinson and Company | REF 212750 | |
| Cycloserine | Sigma-Aldrich | 1158005-250MG | Only for assaying an alternative phenotypic marker |
| D-Glucose anhydrous | Fisher Chemical | G/0500/61 | |
| 50 mL Centrifuge Tube | Corning | REF 430828 | |
| L-(+)-Arabinose | Sigma-Aldrich | A3256-500g | |
| Magnesium sulfate heptahydrate | Fisher Chemical | M/1050/53 | |
| Nalidixic acid | Sigma-Aldrich | N8878-5G | Only for assaying an alternative phenotypic marker |
| Potassium phosphate dibasic trihydrate | Sigma-Aldrich | P5504-500g | |
| Potassium phosphate monobasic | Sigma-Aldrich | P0662-500g | |
| Rifampicin | EMD Millipore Corp, USA | 557303-1GM | |
| Sodium chloride | Fisher Chemical | S/3160/60 | |
| Spectophotometer | Jenway | 6320D | |
| Thiamine hydrochloride | Sigma-Aldrich | T4625-25g | |
| Trisodium citrate dihydrate | Sigma-Aldrich | S1804-500g | |
| Tryptone | Fisher Chemical | 1278-7099 |
参考文献
- de Vries, H. Die mutationstheorie. Versuche und beobachtungen über die entstehung von arten im pflanzenreich. 1, (1901).
- Muller, H. J. Artificial transmutation of the gene. Science. 66 (1699), (1927).
- Muller, H. J. The measurement of gene mutation rate in Drosophila, its high variability, and its dependence upon temperature. Genetics. 13 (4), 279-357 (1928).
- Sturtevant, A. H. Essays on evolution. I. On the effects of selection on mutation rate. The Quarterly Review of Biology. 12 (4), 464-467 (1937).
- Luria, S. E., Delbrück, M. Mutations of bacteria from virus sensitivity to virus resistance. Genetics. 28 (6), 491-511 (1943).
- Jee, J., et al. Rates and mechanisms of bacterial mutagenesis from maximum-depth sequencing. Nature. 534 (7609), 693-696 (2016).
- Fusco, D., Gralka, M., Kayser, J., Anderson, A., Hallatschek, O. Excess of mutational jackpot events in expanding populations revealed by spatial Luria-Delbrück experiments. Nature Communications. 7, (2016).
- Halligan, D. L., Keightley, P. D. Spontaneous mutation accumulation studies in evolutionary genetics. Annual Review of Ecology Evolution and Systematics. 40 (1), 151-172 (2009).
- Jónsson, H., et al. Parental influence on human germline de novo mutations in 1,548 trios from Iceland. Nature. 549, (2017).
- Krašovec, R., et al. Spontaneous mutation rate is a plastic trait associated with population density across domains of life. PLoS Biology. 15 (8), (2017).
- Krašovec, R., et al. Mutation rate plasticity in rifampicin resistance depends on Escherichia coli cell-cell interactions. Nature Communications. 5, 3742 (2014).
- Massey, R. C., Buckling, A. Environmental regulation of mutation rates at specific sites. Trends in Microbiology. 10 (12), 580-584 (2002).
- Lynch, M., et al. Genetic drift, selection and the evolution of the mutation rate. Nature Review Genetics. 17 (11), 704-714 (2016).
- Foster, P. L. Stress-induced mutagenesis in bacteria. Critical Reviews in Biochemistry and Molecular Biology. 42 (5), 373-397 (2007).
- Krašovec, R., et al. Where antibiotic resistance mutations meet quorum-sensing. Microbial Cell. 1 (7), 250-252 (2014).
- Krašovec, R., et al. Opposing effects of final population density and stress on Escherichia coli mutation rate. The ISME Journal-Multidisciplinary Journal of Microbial Ecology. 12, 2981-2987 (2018).
- Lea, D., Coulson, C. The distribution of the numbers of mutants in bacterial populations. Journal of Genetics. 49 (3), 264-285 (1949).
- Armitage, P. The statistical theory of bacterial populations subject to mutation. Journal of the Royal Statistical Society. Series B (Methodological. 14 (1), 1-40 (1952).
- Koch, A. L. Mutation and growth rates from Luria-Delbrück fluctuation tests). Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis. 95 (2-3), 129-143 (1982).
- Cairns, J., Overbaugh, J., Miller, S. The origin of mutants. Nature. 335 (6186), 142-145 (1988).
- Stewart, F. M., Gordon, D. M., Levin, B. R. Fluctuation analysis: the probability distribution of the number of mutants under different conditions. Genetics. 124 (1), 175-185 (1990).
- Drake, J. W. A constant rate of spontaneous mutation in DNA-based microbes. Proceedings of the National Academy of Sciences. 88 (16), 7160-7164 (1991).
- Ma, W. T., Sandri, G. V., Sarkar, S. Analysis of the Luria-Delbrück distribution using discrete convolution powers. Journal of Applied Probability. 29 (2), 255-267 (1992).
- Shapiro, J. A. Adaptive mutation - Who's really in the garden. Science. 268 (5209), 373-374 (1995).
- Matic, I., et al. Highly variable mutation rates in commensal and pathogenic Escherichia coli. Science. 277 (5333), 1833-1834 (1997).
- Rosche, W. A., Foster, P. L. Determining mutation rates in bacterial populations. Methods. 20 (1), 4-17 (2000).
- Rosenberg, S. M. Evolving responsively: Adaptive mutation. Nature Reviews Genetics. 2 (7), 504-515 (2001).
- Lynch, M. Evolution of the mutation rate. Trends in Genetics. 26 (8), 345-352 (2010).
- Zheng, Q. A new practical guide to the Luria-Delbrück protocol. Mutation Research. 781, 7-13 (2015).
- Boesen, J. J. B., Niericker, M. J., Dieteren, N., Simons, J. How variable is a spontaneous mutation-rate in cultured-mammalian-cells. Mutation Research. 307 (1), 121-129 (1994).
- Melnyk, A. H., Wong, A., Kassen, R. The fitness costs of antibiotic resistance mutations. Evolutionary Applications. 8 (3), 273-283 (2015).
- Sun, L., et al. Effective polyploidy causes phenotypic delay and influences bacterial evolvability. PLoS Biology. 16 (2), (2018).
- Frenoy, A., Bonhoeffer, S. Death and population dynamics affect mutation rate estimates and evolvability under stress in bacteria. PLoS Biology. 16 (5), (2018).
- Foster, P. L. Methods for determining spontaneous mutation rates. Methods in Enzymology. , 195-213 (2006).
- Wrande, M., Roth, J. R., Hughes, D. Accumulation of mutants in "aging" bacterial colonies is due to growth under selection, not stress-induced mutagenesis. Proceedings of the National Academy of Sciences of the United States of America. 105 (33), 11863-11868 (2008).
- Adrien, M., Rémy, D., Stéphane, D., Bernard, Y. flan: An R package for inference on mutation models. The R Journal. 9. 334, (2017).
- Garibyan, L., et al. Use of the rpoB gene to determine the specificity of base substitution mutations on the Escherichia coli chromosome. DNA Repair. 2 (5), 593-608 (2003).
- Stewart, F. M. Fluctuation tests: how reliable are the estimates of mutation rates?. Genetics. 137 (4), 1139-1146 (1994).
- Baba, T., et al. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Molecular Systems Biology. 2 (1), (2006).
- Yamagishi, J. -. I., Yoshida, H., Yamayoshi, M., Nakamura, S. Nalidixic acid-resistant mutations of the gyrB gene of Escherichia coli. Molecular and General Genetics MGG. 204 (3), 367-373 (1986).
- Chen, J., et al. Identification of novel mutations associated with cycloserine resistance in Mycobacterium tuberculosis. Journal of Antimicrobial Chemotherapy. 72 (12), 3272-3276 (2017).
- Lang, G. I., Murray, A. W. Estimating the per-base-pair mutation rate in the yeast Saccharomyces cerevisiae. Genetics. 178 (1), 67-82 (2008).
- Uphoff, S., et al. Stochastic activation of a DNA damage response causes cell-to-cell mutation rate variation. Science. 351 (6277), 1094-1097 (2016).
- Robert, L., et al. Mutation dynamics and fitness effects followed in single cells. Science. 359 (6381), 1283-1286 (2018).
- Kong, A., et al. Rate of de novo mutations and the importance of father/'s age to disease risk. Nature. 488 (7412), 471-475 (2012).
- Narasimhan, V. M., et al. Estimating the human mutation rate from autozygous segments reveals population differences in human mutational processes. Nature Communications. 8 (1), 303 (2017).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved