
Method Article
의 효율적인 생산 및 정제 재조합 쥐과 Kindlin-3 생물 물리학 연구 곤충 세포에서
요약
Kindlins는 인테그린을 통해 세포 부착 해 기본적인 그러나 그들의 연구는 세균의 호스트에서 재조합을 표현에서 발생하는 어려움에 의해 방해되었다. 우리는 여기에서 배큘로 바이러스에 감염된 곤충 세포에서의 효율적인 생산 방법을 설명합니다.
초록
Kindlins는 세포 표면 수용체 인테그린 중에서, talin로 필수적 coactivators 있고 외부에서 인테그린 신호 전달에 참여하고, 세포 핵의 유전자 전사의 조절. kindlins은 ~ 75 kDa의 멀티 도메인 단백질과 NPxY 모티브 인테 β-서브 유닛 세포질 꼬리의 상류 T / S 클러스터에 결합한다. hematopoietically 중요한 kindlin 이소, kindlin-3, 혈전 형성시 혈소판 응집을 위해 중요하다, 감염과 염증 및 골 흡수의 파골 세포 족 세포 형성에 대한 응답으로 굴러 백혈구. 이 과정에서 Kindlin-3의 역할은 다양한 세포 및 생리 학적 연구 결과는. 그러나, 추가 연구에 대한 단백질의 고품질 밀리그램 수량을 취득하는 효율적인 방법이 필요하다. 우리는 배큘-D를 사용하여 재조합 뮤린 kindlin-3의 효율적인 발현 및 정제를 위해, 여기에 설명 된 프로토콜을 개발했다그 생물 리 학적 특성화를 허용하도록 고순도 전장 단백질의 충분한 양을 수득 인 Sf9 세포에서 발현 시스템을 떨어져 나간. 같은 접근 방식은 다른 포유 동물 kindlin 이소 형의 연구에 걸릴 수 있었다.
서문
kindlin 가족의 단백질은 중요한 초점 접착 어셈블리의 구성 요소 및 복잡한 생활에 필수적이다. 3 포유류의 아형 (kindlin-1, kindlin-2 및 kindlin-3)가있는 Kindlins은, talin 1과 함께 세포 수용체의 인테그린의 coactivators 간주됩니다. 인테 - 매개 세포 접착 높은 진핵 생물의 세포 외 기질 (ECM)에 세포 표면을 연결합니다. 그것은 조직의 무결성, 배 발생, 뼈 대사, 지혈, 면역력을 포함 생리적 현상의 과다에 중요한 일반적인 과정이다. 인테 - 매개 세포 부착은 자신의 보존 NPxY 모티브에서 인테 β-서브 유닛 세포질 꼬리 (CTS)에 talin과 kindlin의 결합을 통해 내부 아웃 신호 전달을 통해 활성화됩니다. kindlin 단백질의 생물 의학 중요성은 지금까지 kindlin-2는 전사에 관여하는 몇 가지 최근의 보고서에서 밝혀졌다 핵, 그러나 확장알은 2,3를 제어 할 수 있습니다.
Kindlins는 F2 하위 도메인의 중심에 pleckstrin 상동 (PH) 도메인에 의해 중단되는 양자 C-말단 FERM (4.1 밴드, ezrin, radixin, moesin) 도메인의 소유에 의해 hallmarked 약 75 kDa의의 멀티 도메인 단백질이다 4,5. kindlin-2와 kindlin-3 PH 도메인의 연구는 지질 두 번째 메신저 포스파티딜 이노시톨 - (3,4,5) - 트리스 포스페이트 및 포스파티딜 이노시톨 - (4,5) - 비스 포스페이트 6-8에 바인딩 것으로 나타났다. 그러나 kindlin -1 PH 도메인의 연구는 바인딩으로부터 지질을 방지 이소 특정 염다리 의해 kindlin-1에서 설명 될 수 훨씬 낮은 친화력으로 PtdIns (3,4,5) P (3)에 결합한다는 것을 보여 9. 또, 전개 될 것으로 예측되지만 세포막 10,11의 내부 전단에 세린에 결합되어 kindlins의 F1 도메인에 삽입 ~ 100 아미노산 루프가있다. kindlin의 FERMtalin의 FERM 도메인 pleckstrin 상동 도메인을 소유하지 않더라도 도메인, talin의 FERM 도메인에 일치하는 것으로 간주됩니다. 두 kindlins 및 talin는 FERM 도메인의 F3 지역을 통해 인테 β-꼬리에 NPxY 모티프와 상호 작용하지만, talin가 막 근위 한 12-16을 대상으로하면서 kindlin은 막 말단 모티브에 바인딩합니다. 또 둘 Kindlins 및 talin 다른 FERM 단백질 11,17에서 발견되지 유비퀴틴 조 방면으로 N-말단 F0 도메인을 갖는다. kindlin-2의 F0 도메인에 대한 연구는 독립적으로 포스파티딜 이노시톨 - (4,5) - 비스 포스페이트 풍부한 막 (17)에 결합하는 것으로 나타났습니다.
kindlins는 paralogue 특정 조직 발현 패턴 및 비 중복 생리 기능을 나타낸다. Kindlin-1은 주로 표피에서 발현되고, 또한 작은 범위로 결장, 위, 신장은; kindlin-2 재하 표현 되나 집중되어 가로 무늬 매끄러운근육 및 배아 발달 4로 표현만을 kindlin이고, kindlin-3 거핵구 18에서 발견 kindlin-3의 높은 농도를 가진 조혈 조직에서 발현된다. 그러나, 최근의 연구는 기능성 단백질이뿐만 아니라 19 내피 조직에서 발현되는 것을 제안했습니다.
Kindlin-3에 의한 혈액의 중요한 생리 학적 역할에 심각한 의료 관심입니다. 그것은 혈소판 응집 및 감염과 염증 (21, 22)과 골 흡수 (23) 파골 세포 족 세포 형성에 대한 응답으로 혈전 형성 (20), 백혈구 압연시 확산을 위해 중요하다. 생명을 위협하는 출혈 장애 및 재발 성 세균성 감염 20,24,25 특징으로하는 질환 - 또한, kindlin-3 인간의 파괴는 접착 결핍 유형-III 백혈구에 이르게한다. 생쥐 Kindlin-3 녹아웃 연구에서 단백질의 중요한 기능을 밝혀세포 접착 KIND3 -. / - 마우스는 kindlin-3가 결여 인간의 증상을 닮은 인해 비활성 혈소판 인테그린, 심각한 골 화석 증, 및 손상 백혈구 접착 (20, 22)에 심한 출혈과 같은 별개의 표현형을 표시합니다.
kindlins에 대한 고해상도 구조 데이터는 날짜, 같은 pleckstrin 동성 kindlin-1 9 kindlin-2 (26, 27)의 (PH) 도메인 및 kindlin-1 11 kindlin의 F0 도메인으로 각각의 하위 도메인으로 제한되어 -2 17. 각 kindlin 폴리펩티드의 하위 도메인의 대부분은 그러나 복제 및 구조 분석 (예이츠와 길버트, 게시되지 않은 관찰) 저항했으며, 전체 길이 단백질의 연구는 E.을 사용하여 충분한 양을 표현하고 정화의 어려움에 의해 방해되었다 대장균 (게시되지 않은 관찰과 Harburger 등. 14). kindlin-3와 푸에 상당한 의료 관심이 있습니다nction, 다른 두 가족과 함께, 최근에 우리는 배큘로 바이러스 감염 (12)에 의해 구동 스포 도프 테라 프 루기 페르 세포에서의 재조합 발현에 의해 밀리그램 수량을 생성합니다. 따라서 우리는 여기에 광범위한 구조 연구, 생화학 적 분석에 적합한 곤충 세포 배양에서 재조합 마우스 kindlin-3 밀리그램 수량의 생산을위한 방법을 설명합니다.
이 프로토콜에서 우리는 설계 녹아웃 백 미드 (: 1629 BAC10 KO)를 사용합니다 즉, 혼자, 가능한 비리 (28)를 생성 할 수 없습니다. 바이러스 성 DNA 따라서이 경우도 kindlin-3 유전자 (FERMT3)과 재조합의 결과로 바이러스 높은 표현하지만 중복 늦게 유전자를 대체하는 FERMT3 유전자의 결과를 포함하는 전송 벡터와 재조합에 의해 구출 바이러스 라이프 사이클 (28)의 일부로서 마우스 kindlin-3을 표현 바이러스. 우리는이 문제를 확인kindlin-3의 제조 방법의 다른 표현 호스트로 표현하고 정화하려는 시도로 인해 우리는 복제에 사용하고 많은 표현에 배포 할 수있는 POPIN 벡터 제품군의 다양성에 (게시되지 않은 관찰) 엄청나게 어려운 증명뿐만 아니라 후 29 개최합니다.
프로토콜
IM 존스에 의해 개발 된 1629과 :이 프로토콜은 마우스 kindlin-3 유전자 (FERMT3가) 성공적으로 늦게 P10 프로모터의 하류와 벡터 백 미드 BAC10 KO로 재결합을 허용하는 배큘로 바이러스 시퀀스를 측면에서는 보유하는 벡터에 복제 된 것으로 가정합니다 동료 28. 이 프로토콜의 경우, kindlin-3 유전자는 pOPINE (29)에 클로닝 및 사용 프라이머 및 복제 전략은 다른 12 설명 찾을 수 있습니다. FERMT3 유전자 (kindlin-3) P10의 배큘로 바이러스 프로모터의 제어와 벡터 5 'UTR/ORF603 및 ORF 1629 포함하고 다운 스트림 정화 (29)의 C-말단 그의 6 태그를 인코딩 있도록 플라스미드 설계되었습니다.
1. 곤충 세포 배양 및 유지 관리
- 이전 재조합 배큘로 바이러스의 증폭, 스포 도프 테라 프 루기 페르 세포 현탁 배양 (인 Sf9 세포)에 적응한다성장과 유지된다. 곤충 세포는 항상 전용 조직 문화 흄 후드에서 무균 기술을 사용하여 처리해야한다.
- 곤충 세포의 현탁 배양 SF-900 II (SFM) 100 rpm에서 진탕 27 ° C에서 플라스크에 100 ㎍ / ml의 스트렙토 마이신 100 ㎍ / ㎖의 페니실린 및 보충 혈청 액체 배지에서 배양된다.
- 곤충 세포 배양 밀도 범위를 유지 6 10 x 1과 - 분할 및 신선한 SF-900 II 매체를 사용하여 세포 배양을 희석하여 세포 / ml 7 ~ 10 x 1.
주 : 건강 세포의 크기가 균일 한 모양입니다 모양의 구형해야한다. - 배양 세포의 밀도를 계산하는 혈구와 광 현미경을 사용하여 샘플 볼륨의 셀 개수.
2. 재조합 배큘로 바이러스의 생성
- 문화 및 100 ㎍ / ㎖의 penicilli 보충 SF-900 II 매체를 사용하여 현탁액 인 Sf9 세포를 유지n 및 100 ㎍ / ml의 스트렙토 마이신. 1 × 106 세포 / ml - 배큘로 바이러스 생성을위한, 곤충 세포를 5 × 105의 밀도 범위로 배양한다.
- 시드 약 1 × 10 100 ㎍ / ml의 스트렙토 마이신 100 ㎍ / ㎖의 페니실린 및 보충 SF-900 II 미디어 2 ㎖의 멸균 6 잘 조직 배양 플레이트의 웰 당 6 인 Sf9 세포. 플라스틱 우물의 바닥에 부착 흄 후드에서 실온에서 인 Sf9 세포를두고 따라서 단일 층을 형성한다.
- 단일 층 문화에 백 미드의 DNA와 플라스미드 DNA를 cotransfecting에 의해 재조합 배큘로 바이러스 생성을 수행합니다.
- 항생제 (솔루션)이없는 SF-900 II SFM 100 μL 1629 : 각 형질의 경우, 정제 BAC10 KO 0.5 μg과 ORF1629 배큘로 바이러스 요소를 가지고 정제 pOPINE-mFERMT3, 1 ~ 2 μg을 섞는다.
- 별도의 관으로, 안티없는 SF-900 II SFM 100 ㎕ 씩 Cellfectin II 시약 6 μl를 희석각 형질 전환 반응 (용액 B)에 대한 바이오 틱. 많은 형질이 필요한 경우 '마스터 믹스', 감소 된 액체의 처리를 위해, 여기에서 생성 할 수 있습니다.
- 두 용액 (A와 B, 약 200 μL)을 혼합하고 지질-DNA 복합체를 형성하기 위해 20 분 동안 RT에서 배양한다.
- 항생제없이 800 ㎕의 SF-900 II SFM과 지질-DNA 복합체를 희석. 조심스럽게 인 Sf9 세포 단일 층 미디어를 대기음 조심스럽게 인 Sf9 단일 층의 상부에 용액 A / B와 미디어를 피펫.
- 27 ° CO / N에 가습 인큐베이터에서 형질 전환 세포를 품어 다음 날 각 단층 문화 항생제없이 SF-900 II SFM의 추가 1 ML을 추가합니다. 추가로 5 일 동안 27 ° C에서 세포를 품어.
- 배양 배지 (총 약 2 ㎖)에서 직접 재조합 배큘로 바이러스를 수확하고 깨끗한 원심 분리 관 (예를 들어, 15 ML 팔콘 튜브)로 전송.
- 모든 인 Sf9 세포 debr을 명확히RT에서 5 분 1,000 XG에서 원심 분리하는 것입니다. 사용할 때까지 어둠 속에서 4 ° C에서 깨끗한 튜브 및 저장소에 상층 액에 있으며 P1을 표시 바이러스를, 전송합니다. 이 단계에서, 나머지 인 Sf9 단층은 곤충 세포에서 재조합 kindlin-3의 존재를 평가하여 재조합 바이러스 생성을 평가하기 위해 사용될 수있다.
- 0.5 ml의 PBS로 단일 층을 재현 탁하고 2X SDS-PAGE 로딩 버퍼의 동일한 볼륨과 10 ㎕의 샘플을 희석. 적어도 10 분 동안 95 ° C>에서 샘플을 가열한다.
- 그것은 적절한 젤 로딩이 너무 점성 경우 마이크로 소니 케이 팁을 사용하여 10 %의 진폭에서 1 초 동안 샘플을 초음파 처리.
3. 재조합 배큘로 바이러스의 증폭
- 현탁액 중의 바이러스의 증폭을 위해, 1.4 × 106 세포 / ㎖의 세포 밀도를 사용하고 이것은 (즉, 50 ㎖의 배양 플라스크에서 전체 부피의 1 / 20을 차지한다2 L 플라스크).
- 다음과 같은 공식을 사용하여 0.1의 감염의 다중성 (MOI)에서 P1 바이러스 재고와 곤충 세포 배양을 감염시켜 재조합 바이러스의 증폭을 달성;

주 : P1 바이러스 생성은 1 × 10 7 PFU / ㎖의 예상 바이러스 역가를 갖는 것으로 가정 할 수있다. 그러나, 플라크 분석은 이전에이 단계를 수행 할 수 있습니다. - 3 일 (72 시간) 100 rpm에서 진탕 27 ° C에서 P1에 감염된 곤충 문화를 품다.
- RT에서 5 분 1,000 XG에서 원심 분리하여 매체에서 세포를 분리하여 바이러스를 수확. SDS-PAGE 및 웨스턴 블로 팅에 의해 kindlin-3 단백질 발현을 평가하여 재조합 바이러스 생산의 확인이 단계에서 생성 된 세포 펠렛을 저장합니다.
- 4 ° C에서의 깨끗한 튜브 및 저장소에 명확히 바이러스 풍부한 미디어로 이동사용할 때까지 어둠. 이 바이러스 주식은 P2로 표시된다.
참고 : 2 × 10 8 PFU / ㎖ (100 PFU / 셀의 증폭 율)의 바이러스 역가를 예상 할 수있다.
4. 배큘로 바이러스에 감염된 인 Sf9에 kindlin-3의 발현
- 이전 정화 대규모 재조합 kindlin-3 발현에 100 ㎍ / ㎖의 페니실린 및 100 ㎍ / ml의 스트렙토 마이신, 보충 SF-900 II SFM에 현탁액 인 Sf9 세포 배양의 적절한 볼륨을 성장. 100 rpm에서 진탕 및 전체 문화 볼륨 27 ° C에서 서스펜션 문화를 품다 : 1:5의 부피비 플라스크.
- 2 × 106 세포 / ml의 밀도로 인 Sf9의 현탁 배양 물을 감염.
- 1 %의 최종 농도 인 Sf9 문화를 보완 (V는 / V) 태아 혈청 (FBS)이 1 MOI에게 제공하기 위해 증폭 된 재조합 바이러스 (P2-바이러스 주)로 하였다. 100 rpm에서 진탕 27 ° C에 감염된 문화를 품다.
- 수확 recombinaNT kindlin-3-CHIS 1,000 XG에서 원심 분리하여 인 Sf9 세포에게 72 시간의 감염 후 6 발현 그 결과 세포 펠렛은 사용할 때까지 -20 ° C에 저장, 또는 장기 저장을 위해 -80 ° C.
5. 재조합 Kindlin-3의 정제
- 얼음에 재조합 kindlin-3를 발현하는 해동 냉동 배큘로 바이러스에 감염된 곤충 세포 (인 Sf9) 펠렛.
- 용해 완충액으로 해동 된 세포 펠렛을 재현 탁 (50 mM 트리스-HCl, pH7.5, 500 mM의 염화나트륨, 1 % (v / v) 트윈 -20), EDTA 프리 프로테아제 저해제 칵테일 및 DNase1의 1,000-2,000 U로 보충.
주 : 염화나트륨 농도 내인성 인 Sf9 세포의 단백질 및 하류 정화에 사용 고정화 금속 친 화성 칼럼 (아래 참조) 간의 비특이적 인 상호 작용을 방지하기 위해 500 mm로 조정되는 경우 대안 적으로, 수정 된 인산 완충 식염수 (PBS)도 사용될 수있다. - 세제와 할머니와 함께 재현 탁 세포를 배양하여 세포를 Lysertexing. 초음파 처리 얼음 욕조에서 추가 세포 분열에 대한 샘플을 (40 % 진폭, 10 초 펄스의 10주기는 10 초 냉각 다음에) 또는 다른 방법 다운스 호모 게 나이저를 사용합니다.
- 4 ℃에서 1 시간 동안 48,000 XG에 원심 분리하여 해물을 명확히 1 ㎖ / 분의 속도로 4 ° C에서 HisTrap 열 (5 ㎖ 열 볼륨), 용해 완충액으로 미리 평형화에 상층 액을 넣습니다.
참고 : 또는 명확히 해물 사전 평형 니켈 세파 비즈의 1 ~ 5 ㎖의 침대 볼륨 1 ~ 2 시간 동안 4 ° C에서 (예를 들어, 니켈 파로스 6 빠른 흐름)와 함께 배양 할 수있다. 니켈 파로스의 열은 중력 유동 열을 이용하여 결합 단계 후에 형성 될 수있다. - 세척 버퍼의 10 열 볼륨과 열을 세척 (에 50 mM Tris-HCl, pH 7.5, 500 mM의 NaCl을, 10 mM의 이미 다졸)이 결합되지 않은 단백질을 제거합니다.
- 용출 악타 FPLC를 사용하여 10 월 / ㎖ (또는 10 열 볼륨에서)의 속도로 10 ~ 500 밀리미터에서 선형 이미 다졸의 그라데이션을 사용하여바인딩 재조합 kindlin-3-CHIS 6.
- 보통 300 mM의 이미 다졸의 농도에서 용출 kindlin -3 - CHIS 6을 함유하는 분획을 함께 악타 FPLC를 사용하여 0.5 ㎖의 분획으로 용출 분획.
- SDS-PAGE에 의해 용출액의 단백질 조성물을 평가하고 처음 않는 정화를 위해 항-6 그의 항체 또는 항 - 마우스 kindlin-3 항체를 사용하는 웨스턴 블롯에 의해 확인한다.
- 20 mM 트리스-HCl, pH7.5, 200 mM의 NaCl을 50 kDa의 분자량 컷 오프 (MWCO)로 원심 단백질 농축기를 사용하여 버퍼 및 시료 농도로 희석 시리즈 경유로 kindlin-3 및 버퍼 교환을 함유하는 분획을 풀링 4 ° C에서
참고 : 또한, 20 mM 트리스-HCl, pH7.5, 200 mM의 염화나트륨에 O / N로 4 시간 동안 4 ° C에서 30 kDa의의 MWCO로 슬라이드-A-lyzer 투석 카세트 테이프를 사용하여 단백질 용액을 dialyze.
참고 : 용 이온 교환 (아래 참조)의 NaCl의 낮은 농도, 일 수 있으며이 단계에 사용되었다. - 0.5 ㎖ / 분의 속도를 사용하는 FPLC 악타 미리 평형화 된 HiTrap 헤파린 HP 컬럼 (5 ㎖ 칼럼 부피)로 버퍼 교환 된 단백질 용액을 적용한다.
주 : 바운드 kindlin-3 10mM의 / ㎖의 속도로 증가하고, 동일한 완충액에서 선형 염화나트륨 구배 (1 M의 NaCl, 0.2 M의 NaCl)를 이용하여 용출한다. Kindlin-3-CHIS6은 ~ 0.6 M의 NaCl에서 용출 할 것으로 예상된다. - 0.5 ㎖의 분수에 용출을 분별하고 적절한 경우, SDS-PAGE 및 웨스턴 블로 팅에 의해 단백질 구성을 평가합니다.
참고 : SDS-PAGE에 의해 평가 하나에 가까운 95 %의 단백질 순도를 기대할 수 있습니다. - 0.5 ㎖의 최종 부피로 50 kDa의 MWCO를 가진 단백질 원심 농축기를 이용하여 kindlin-3 및 농축을 함유 분획을 풀.
- 최종 단계에서, 농축 된 단백질을 연마 및 크기 배제 크로마토 그래피 (SEC)를 사용하여 단백질을 교환 버퍼.
- 스와로 정제 된 단백질을 적용합니다perdex S200 (16 / 59) 또는 (11 / 30) 컬럼의 크기에 따라 1 ㎖ / 분, 0.5 ㎖ / 분의 속도로 20 mM 트리스-HCl, pH7.5, 200 mM의 염화나트륨, 1 mM의 DTT에 미리 평형화 사용.
주 : 필요한 경우, 크기 배제 크로마토 그래피는 또한 인산 완충 식염수 (PBS)에서 수행 될 수있다. - 사용되는 칼럼의 크기에 따라 / 분 1 ㎖, 0.5 ㎖ / 분의 속도로 컬럼에 적용하여 버퍼 사이즈에 의해 단백질을 정제 하였다.
주 : 컬럼으로부터 용출 단백질 분획하고 280 nm에서 흡광도를 사용하여 모니터링. - 단일 흡광도 피크 분획 및 균질성을 결정하기 위하여 SDS-PAGE에 의해 평가하고,이 단계 후에 일반적으로> 95 % 순수 SEC에서 예상되어야한다.
- 스와로 정제 된 단백질을 적용합니다perdex S200 (16 / 59) 또는 (11 / 30) 컬럼의 크기에 따라 1 ㎖ / 분, 0.5 ㎖ / 분의 속도로 20 mM 트리스-HCl, pH7.5, 200 mM의 염화나트륨, 1 mM의 DTT에 미리 평형화 사용.
- 집중 50 kDa의 MWCO에 ~ 15 ㎎ / ㎖, 109,320 M의 계산 된 흡광 계수 (ε)를 사용하여 분광 광도계 평가로 -1 cm -1 <와 원심 분리기 단백질 집중을 사용하여 kindlin-3-CHIS 6 정제/ SUP> (모든 시스테인 잔기가 감소한다고 가정).
- -80 ° C에서 -20 ° C 및 장기 기억에 저장을 위해, 나누어지는 PCR 튜브 및 플래시 동결 액체 질소에서 샘플에 단백질. 대안 적으로, 단백질은 생화학 및 생물 물리학 적 기법의 개수를 사용하여 조사를 위해 직접적으로 사용될 수있다.
결과
배큘로 바이러스에 감염된 인 Sf9 세포를 사용하여 재조합 마우스 kindlin-3의 대규모 발현 개략적도 1a에 도시 된 바와 같이, 밀리그램 양을 달성하기 위해 2 주 미만 취해 QIAprep 미니 프렙 키트에서 플라스미드 DNA의 소량 만이 필요위한 수 예. 도면에 도시 된 바와 같이, 5-7 일 후에 새로 형성된 비리 수확 : 재조합 바큘로 바이러스의 생성은 마우스 kindlin -3 (FERMT3) 설계 선형화 된 백 미드 (KO 1629 BAC10)와 함께 플라스미드를 함유하여 인 Sf9 세포를 cotransfecting함으로써 달성된다 1A. 기본적으로이 100 % 재조합 바이러스의 배큘로 바이러스 생성 결과의 방법과는 플라크 정제 (28, 30)에 대한 필요성을 분배합니다. 대표적인 소규모 (2 ㎖ 단층) 문화 ML의 당 1 × 10 7 플라크 형성 단위의 예상 바이러스 역가 (PFU)와 재조합 바이러스를 포함하는 솔루션을 생성합니다문화. 하나는 실제 바이러스 역가를 결정하는 플라크 분석을 수행 할 수 있지만, 구조의 높은 수의 구조 연구에 대한 검사를 할 때이 아마도 너무 노동 집약적이다. 바이러스 생성 단계의 성공은 일반적으로 설명되는 본 kindlin-3 구조를 들면 인 Sf9 단층은 PBS에 재현 탁 될 수 있고, SDS-PAGE 및 웨스턴 블롯에 의해 평가 병렬 EGFP 함유하는 플라스미드를 사용하여 평가 될 수있다, 또는 도 1b에 도시 된 바와 같이 75 kDa의 그의 표지 된 단백질에 대응하는 클리어 밴드. 바이러스이어서 대규모 (리터 부피) 곤충 세포의 감염 및 재조합 단백질의 분리를위한 충분한 양을 생성하기 위해 증폭된다. 제 2 통로 바이러스 (P2)은 0.1의 MOI 잡은 (프로토콜 참조) 현탁 배양 물을 감염시켜 생성된다. 그것은 증폭 인 Sf9 서스펜션 문화는 총 플라스크 볼륨 만 이십분의 일을 차지하는 것이 중요합니다. 이 추가 통풍이 보장하는 결과 비르3 일 (72 시간) 후 감염 후 수확 우리 함유 매체는, 이후의 식 문화에 kindlin-3의 충분한 양을 생성합니다. 증폭 된 바이러스 재고 (P2)가 2 × 10 6 세포의 세포 밀도를 사용하여 / ㎖ 증폭 31시 100 PFU / 셀의 보수적 인 추정치에 기초하여 2 × 10 8 PFU / ㎖의 예상 바이러스 역가를 가지고 간주됩니다. 도 1C에 도시 된 바와 같이 일반적으로, 최적의 배큘로 바이러스의 증폭 및 단백질 생산을 위해, 인 Sf9 세포 크기와 구형으로 균일해야한다. 또한, 바이러스의 증폭 및 단백질 발현은 72 시간 게시물 감염에서 최대이며, 둘은 크게 96 시간 게시물 감염에서 줄일 수 있습니다.
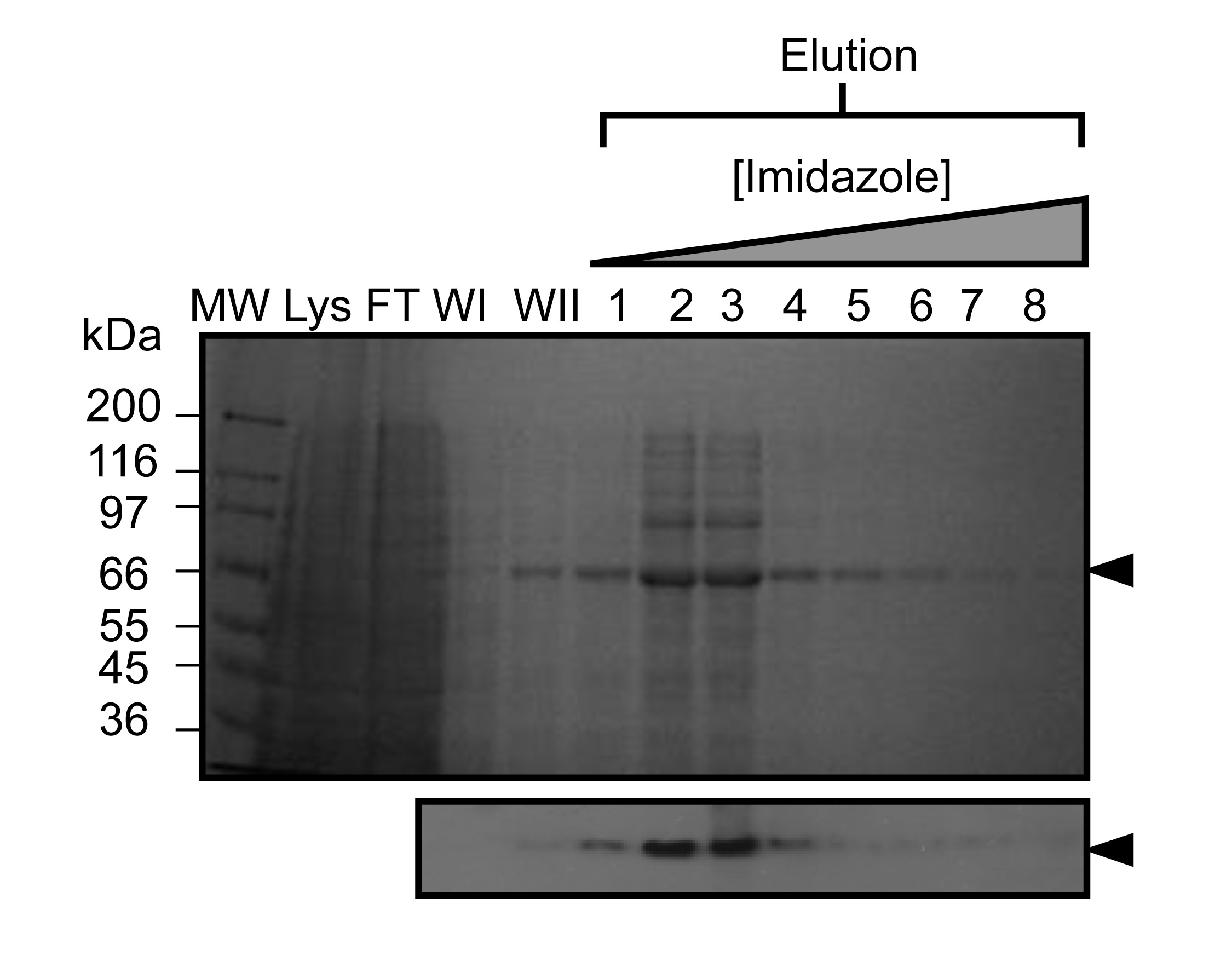
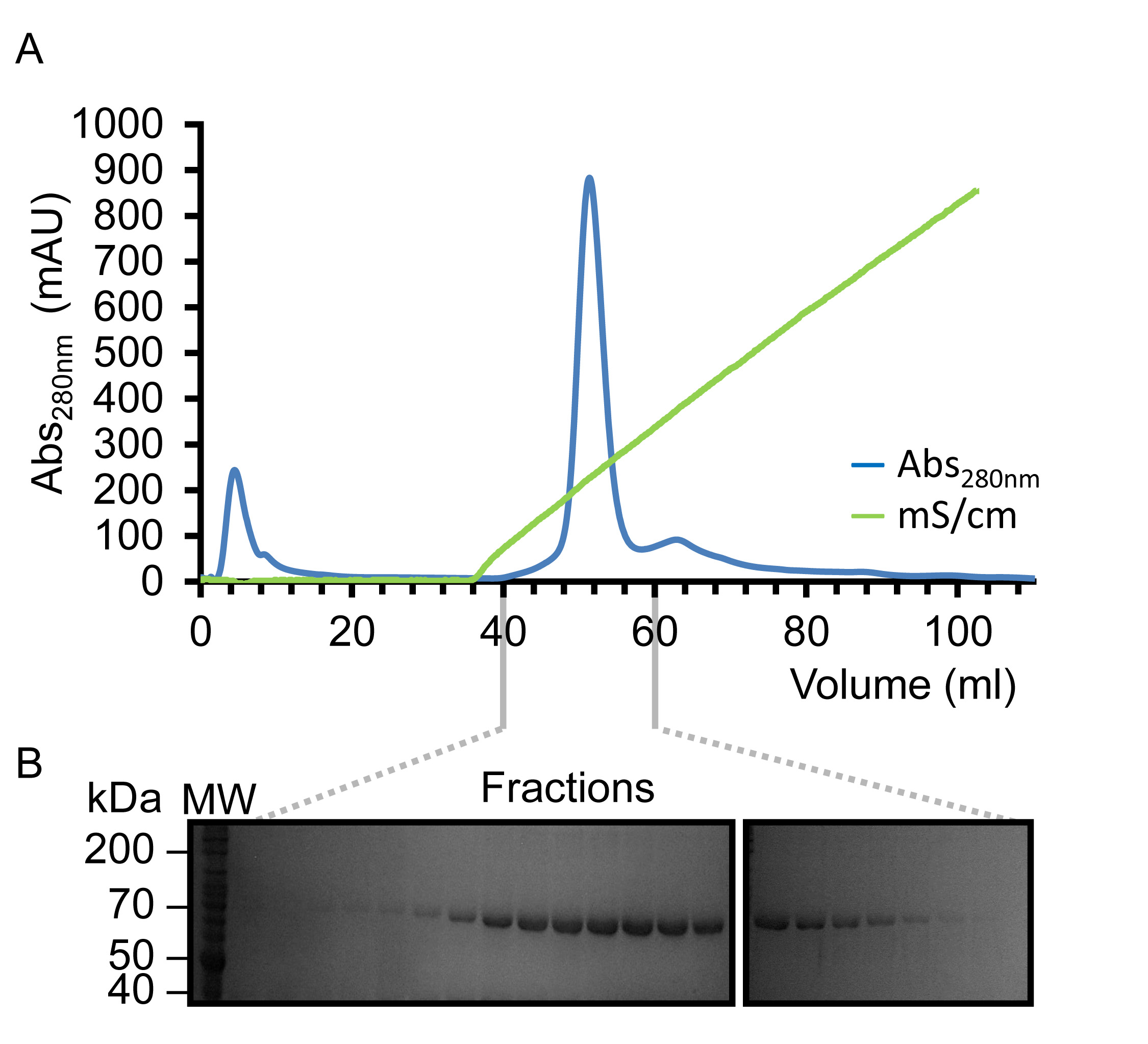
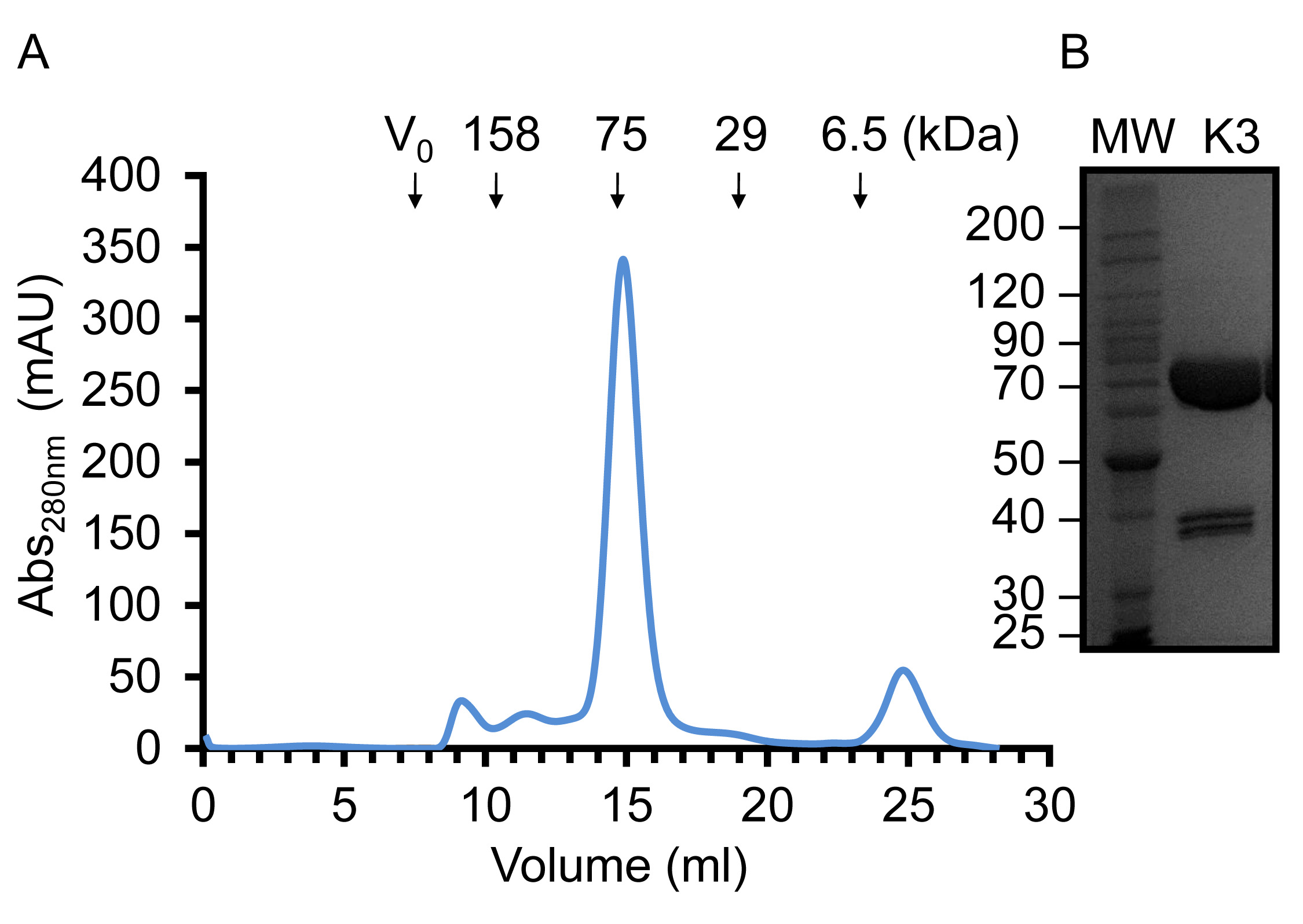
바이러스가 증폭되고 큰 규모의 실험 인 Sf9 세포를 감염하는 데 사용되면 재조합 kindlin-3은 부분적으로는 설계 C-말단 그의 6 태그를 사용하여 고정 된 금속 친 화성 크로마토의 미덕에 의해 정제그래피,도 2에 도시 된 바와 같이. 그것은 4 ° C에서 정화를 수행하는 것이 중요하고 충분한 단백질 분해 효소 억제제는 단백질 분해를 방지하기 위해 추가되었습니다. 도 3에 도시 된 바와 같이 부분적으로 정제 kindlin-3, 헤파린 컬럼을 이용한 이온 교환 크로마토 그래피 (IEC)에 의해 가까운 동질성을 추가로 정제한다. 우리는 대신에, 종래의 이온 교환 칼럼의 헤파린 컬럼의 사용을 채용 제외한 kindlin -3 F1 도메인 내의 폴리 리신 스트레치 등 염기성 잔기의 다수는, 음전하 황산염 그룹과 강하게 상호 작용할 것이라고 예측대로 열. 이 전략은 CID1 32 uridylyltransferase 예를 들어 RNA 결합 터미널, 기본 핵산 결합 패치 DNA 및 RNA 결합 단백질에 특히 유용합니다. 마지막 kindlin -3 순도 같이 응집체를 분리하고 균질성을 달성하기 위해, 크기 배제 크로마토 그래피에 의해 "연마"입니다그림 4. 프로토콜에 지정된 버퍼는 자주 구조 분석을위한 단백질 정제에 사용되는 표준 버퍼입니다. 일반적으로, 인산염 기반의 버퍼는, 구조 연구에 단백질의 정제에 의한 (특히 4 ° C에서 실험) 결정화에 떨어진다 인산 결정의 형성에 특히 결정화 검사를 피할 수있다. 도 5에 도시 된 바와 같이 그러나, 버퍼가 kindlin-3에 대해 안정화 된 결정 Thermofluor 기반 열 변화 분석법을 수행 하였다. 간단히, 정제 된 단백질 용액을 따라서 2 차원 화면을 형성, pH와 염화나트륨 농도의 범위를 커버 버퍼로 희석한다. 단백질의 용해는 온도 범위 20 ~ 95 °의 C (293-368 K)에 접힌 단백질 코어 내에서 소수성 잔기에 결합 Sypro 오렌지 염료 (분자 프로브)에서 형광을 관찰하여 측정한다. 온도 중간 지점 토륨하는E 단백질 (전이 온도 T의 m은)의 Opticon 모니터 소프트웨어를 사용하여 계산하고, 그 밖에 33 설명한 전개. Kindlin-3 일관성 전이 온도 (T의 m)로, pH 범위 7.0-9.0 내에 높은 염화나트륨 농도 (500 ㎜)에 안정한 것으로 관찰되었다 55 ° C의 또한 Kindlin-3 T의 m은 7.0-7.5 무관 염화나트륨 농도의 pH 범위 내에서 약 55 ° C 것을 관찰 하였다.
우리는 거기 kindlin-3의 제한으로 단백질 분해가 정화 중에했지만 고농축 단백질 (~ 15 ㎎ / ㎖)의 SDS-PAGE 분석은 동일한 강도의 두 개의 추가 폴리펩티드로 제한된 오염을 증명 것으로 나타났습니다. 도 4에 도시 된 바와 같이 이상한 그러나, 크기 배제 크로마토 그래피에 의해 추가로 종의 징후는 없다. 그것은 따라서 단백질은 단백질 분해 효소에 의해 다친 것을 생각하지만 접어 hydrodyn 유지전체 길이의 단백질에서 amically 구별. 흥미롭게도, 칼 페인 알려진 단백질 분해 효소입니다 pleckstrin 상동 (PH) 도메인 (34)의 β1-β2 루프에 우리의 관찰을 설명 할 수 Tyrosine373,에서 kindlin-3을 절단. 요약하면 또한, 서부 모래 바닥은 폴리 펩타이드 이중선 중 하나가 C-말단 그의 태그와 이중선의 겉보기 분자량을 보유하고 있음을 보여, 75 kDa의 네이티브 단백질과 같은 분자량은 같습니다. 정제 된 재조합 kindlin -3 리터당 인 Sf9 세포 (~ 2g 세포 무게)의 수율은 기껏 5 밀리그램이다.

그림 1. 배큘로 바이러스 generatio의 배큘로 바이러스에 감염된 곤충 세포 이종 단백질 발현의 개요. (A) 도식화 개요인 Sf9 세포에서 N과 kindlin-3의 발현. DNA 벡터 도식에서 'UTR / ORF603은 빨간색으로 색 ORF1629은 녹색으로 색과 관심 (POI)의 단백질의 유전자는 파란색으로 표시됩니다. (B) 재조합 배큘로 바이러스 재조합 쥐 kindlin-3를 생산 인 Sf9 세포의 6 개의 작은 규모 (2 ㎖ 단층) 문화 (차선은 문화에 따라 레이블)의 안티 그의 6 항체를 이용한 웨스턴 블롯,,. (C) SF-900 II SFM에 정지에서 자란 건강한 인 Sf9 세포의 광학 현미경 이미지는 항생제로 보충하고 35mm 잘 조직 배양 접시 (프로토콜 참조)로 전송. 이미지는 20 배 배율입니다.

그림 2. Represe니켈 친 화성의 고정화 금속 친 화성 크로마토 그래피 (IMAC)에 의해 kindlin-3 ntative 정화. SDS-PAGE 정제 된 재조합 kindlin-3 인 Sf9 세포 (~ 2g 세포 무게) 감염 배큘로 바이러스로 표현 된 쥐. 흡착 된 단백질 (겔 위에 표시된) 이미 다졸 구배를 사용하여 용출시켰다. 다음과 같이 차선이 표시되어, MW, 분자량 마커,리스, 전체 세포 용 해물, FT는 (언 바운드)을 통과, 위스콘신, 1 세척, WII, 2 씻는다. 웨스턴 블롯 분석 (아래) 또한 재조합 단백질의 설계 그의 태그의 존재를 확인하기 위해 용출 분수를 사용하여 수행 하였다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3. Reprekindlin -3 헤파린 친 화성 크로마토 그래피의 sentative 정제. (A) 용출 프로필은 0.05-1.0 M.의 염화나트륨 농도 범위를 사용하는 선형 염화나트륨 구배 (그린)하에 용출 단일 대칭 피크를 표시하는 파장 280nm (청색) (관찰 75 kDa의 단백질의 존재를 보여주는 분별 용출 B) SDS-PAGE 분석, kindlin-3 (표시 K3는). 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

사용도 4. 주제 겔 여과 크로마토 그래피 정제 kindlin-3의 농축 kindlin-3. (A) 겔 여과 용출 프로파일20 ° C에서 트리스-HCl, pH7.5, 150 mM의 염화나트륨 및 1 mM의 DTT의을 Superdex S200 (16 / 59) 용출 부피를 기준으로, kindlin-3을 이주는 그것이 주로 단량체임을 시사 75 kDa의 단백질에 대한 예상대로. (B)의 SDS-PAGE 고농도의 정제 된 재조합 kindlin-3 14.5 밀리그램 / ML. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

버퍼 상영. Kindlin-3도 5. Thermofluor 계 열 시프트 분석법 염화나트륨 농도 대 pH를 2 차원 화면을 포함하는 다양한 버퍼로 희석 하였다. 전이 온도 (온도 중간 점)의 형광으로 관찰소수성 결합 염료, Sypro 오렌지 (분자 프로브)과의 Opticon 모니터 소프트웨어를 사용하여 계산. (A) 전이 온도의 차원 히스토그램 (B) 50.4 ° C.의 계산 된 평균에서의 전이 온도의 변화와 함께 플롯 이해를 돕기 위해 바가을 해당되는 온도 범위에 따라 색상이 지정됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
토론
배큘로 바이러스 발현 시스템은 점점 더 인기를 X-선 결정학을 포함한 생물 물리학 연구를 사용하여 단백질의 특성에 대한 재조합 단백질의 밀리그램 양의 생산을위한 중요한 도구가되고있다. 더 실험적 요구에도 불구 배큘 발현 시스템은 E. 비해 여러 장점을 제공한다 대장균 중 하나는 적절한 보호자 및 번역 후 변형에 대한 기회의 존재 예를 들어, 진핵 세포 유래의 단백질에 대한 네이티브에 가까운 환경입니다. kindlin-3을 표현하는 우리의 노력에서 다른 표현 호스트는 포유 동물 세포 라인과 박테리아 발현 균주 (게시되지 않은 관찰)을 포함 하였다. 일반적으로, 많은 E. 대장균 시험 균주는 재조합 kindlin-3의 매우 적은 양 (;되지 않은 관측 ~ 0.5 ㎎ / 문화 L)를 생산했다. 그러나, 곤충 세포에서 배큘로 바이러스 중심의 표현은 공동으로, 특히 효과적이었다재조합 세포질 단백질 (게시되지 않은 관찰)의 밀리그램을 분리하는 데 필요한 큰 바이오 매스를 생성 할 의무가 더 포유 동물 세포에서 일시적 발현에 mparison. 우리는 존재 진핵 생물의 보호자가 kindlin-3의 효율적인 생산을 허용 할 수 있음을 추측.
baculoviridae는 곤충 세포를 감염 및 재조합 단백질 발현이 작품에 사용되는 배큘로 바이러스는 핵 polyhedrosis 바이러스 (AcNPV) 캘리 포니 오토 그라파에 기초한다. 자연 속에서 오토 그라파 캘리 포니 카 (알팔파 lopper) 곤충의 유충에 감염 AcNPV는 virons는 따라서 그들의 릴리스에 대한 필요한 보호를 제공하는 결정 성 단백질 매트릭스에 캡슐화되어있다 폐색을 형성하는 폴리 헤드린 단백질을 필요로한다. 배양 된 세포에 폐쇄기구의 형성은 복제에 필요하기 때문에 비용 소비하지 않습니다. 외래 단백질을 발현의 경우는 폴리 헤드린 단백질 유전자는 R 수있다관심의 단백질에 대한 유전자 재조합 AcNPV에 eplaced. AcNPV 다른 Lepidopteron 종을 감염시킬 수있는 재조합 단백질 발현 군대 웜 스포 도프 테라 프 루기 페르의 목적을 위해 번데기 난소 세포가 사용됩니다. 그것이 같은 것을 : 필수적인 바이러스 성 유전자, ORF1629가 (KO 1629 BAC10) 노크 아웃 백 미드의 결과 클로람페니콜 아세틸 트랜스퍼의 삽입에 의해 불 활성화 될 수 있도록 여기에 설명 된 방법에서는 AcNPV의 백 미드 (BAC10)가 설계된다 전염성 baculovirions (28)를 형성 할 수 없습니다. KO 1629 및 FERMT3 함유 전송 벡터 수리도 폴리 헤드린 프로모터 (28)의 제어하에 FERMT3 유전자를 포함하고 있습니다 가능한 게놈의 결과로 조옮김을 통해 비활성 ORF1629,, : 선형 BAC10와 인 Sf9 세포의 공동 형질.
우리는 THRE 통해 kindlin-3 고순도의 재조합 마우스의 격리를위한 정화 프로토콜을 설명E 단계 크로마토 그래피 방법. 여기에 사용되는 방법은 쉽게 다른 그의 태깅 된 단백질에 적용될 수있다. 우리는 더 kindlin-3를 정화 이온 교환 단계를 사용하지만 우리는 kindlin-3는 F1 도메인 내에서 폴리 라이신 스트레칭 등의 기본 잔류의 큰 숫자를 가지고 같은이 더 의사 선호도 단계라고 생각합니다. 또한 kindlin-3에 결합하고 함수 원형질막의 세포질 표면과 상호 작용하는 것으로 간주되고, 따라서 우리는 염기성 잔기의 클러스터링이 단백질은 음으로 하전 된 막에 대항 할 수 있다고 예측한다.
정제 프로토콜에 기술 된 버퍼는 표준 간주 자주 구조 생물학에서 사용된다. thermofluor 분석 (그림 5) kindlin-3는 pH가 6.0 이상이 가장 버퍼 조건에서 안정되어 있음을 보여줍니다. β1 : kinldin-3을 연구 할 때 우리의 실험을 알리는에 특히 유용하고 중요염화나트륨 (12)의 낮은 농도로 pH 6.1에서 뛰어난 스펙트럼을 산출 NMR에 의해 꼬리 상호 작용.
모든 생물 물리학 연구에 착수하기 전에, 그것은 관심의 정제 된 단백질이 실제로 제대로 접혀 기능적으로 활성화되고 있음을 입증하는 것이 중요합니다. 이전의 간행물에서 우리는 재조합 kindlin-3 발현 및 크기 - 배제 크로마토 그래피, 동적 광산란, 분석 용 초 원심 분리 및 소각 X 선 산란에 의해 평가 정제 된이 방법을 사용하면, 용액 중의 단량체와 단 분산 것을 입증하고 있었다 또한 이전의 세포 생리 학적 연구 14,20,22와 라인에 기본 단백질, 같은 동작하는지 확인하여, 바인딩 및 막 원심 NPxY 및 β의 1A 세포질 꼬리 (14)의 상류 세린 / 트레오닌 클러스터를 인식 할 수. 열 안정성 분석의 사용을 제안 번째의 추가 방법관심의 단백질의 전자 적절한 폴딩, 같은 잘못 접힌 단백질로 인해 노출 된 소수성 잔기에 높은 형광 배경에 발생합니다.
단백질의 kindlin 제품군은 생체 내에서 인테그린의 필수 coactivators 그들의 예기치 않은 역할이 발견 된 이후 많은 관심의 초점을했습니다. 이 재조합을 표현하고 그 구조를 해결하기 위해 많은 노력이 시작되었습니다. 지금까지 제한된 성공은 전체 길이 재조합 단백질의 밀리그램 수량을 표현에서보고되고있다 그러나 우리는 여기에 구조 연구가 가능하게 수준에서 큰 규모의 표현을 허용하는 배큘로 바이러스 시스템의 사용을 설명합니다. 재조합 kindlin-3의 대량 생성함으로써 우리는이 단백질의 추가 연구를 도움이 될 것으로 기대하고 있습니다. 재조합 쥐 kindlin-3 여기에 설명 된 배큘로 바이러스 구동 방법 및 정화 워크 플로우는 다른 kindlin 이소 형을 표현하고 정화하는 데 사용할 수어느 또한 표현하고 또한 폴리 라이신 펼쳐진을 소유하기 어려운, 또한 이러한 균주에 표현하지 핵산 결합 단백질과 같은 다른 세포질 단백질을 위해 구성 될 수있다.
공개
저자가 공개하는 게 없다.
감사의 말
우리는 문화 인 Sf9 세포 주식의 유지 보수 기술 지원을 Weixan 루 감사합니다. LAY는 의학 연구위원회 (MRC) 대학원 재학에 의해 지원되었다. RJCG는 왕립 학회 대학의 연구원이었다. 구조 생물학의 옥스포드 사업부는 인간 유전학, 웰컴 트러스트 (Wellcome Trust) 코어 상 부여 번호 090532/Z/09/Z에 대한 Wellcome 트러스트 센터의 일부입니다.
자료
| Name | Company | Catalog Number | Comments |
| Sf-900 II serum free media (SFM) 1x liquid | Life Technologies | 10902-096 | store at 4 °C and warm to RT before use |
| Cellfectin II Reagent | Invitrogen | 10362-100 | Alternatively, GeneJuice transfection (EMD) reagent can be used |
| Streptomycin sulphate (solid) | Melford | S0148 | Sterilize filter (0.22 μm filter) before use |
| Penicillin G, potassium salt (solid) | Melford | P0580 | Sterilize filter (0.22 μm filter) before use |
| CELLSTAR Sterile 6-well Culture Plate | Greiner Bio-One | 657160 | |
| Fetal Bovine Serum (FBS) | Life Technologies | 10100-147 | |
| Protease Inhibitor Cocktail | Sigma | P8849 | Caution: Protease inhibitors are dissolved in DMSO |
| Bovine pancrease deoxyribonuclease (Dnase) I | Sigma | D5025 | |
| HisTrap FF (5 ml) | GE Heathcare | 17-5286-01 | Requires an Äkta FPLC machine |
| HiTrap Heparin (5 ml) | GE Heathcare | 17-0407-01 | Requires an Äkta FPLC machine |
| Amicon Ultra-15 Centrifugal Filter Units (with Ultracel-50 membrane) | Millipore | UFC905024 | 15 ml capacity and a MWCO of 50 kDa protein concentrator |
참고문헌
- Moser, M., Legate, K. R., Zent, R., Fassler, R. The tail of integrins, talin, and kindlins. Science. 895, 899- (2009).
- Yu, Y., et al. Kindlin 2 forms a transcriptional complex with β-catenin and TCF4 to enhance Wnt signalling. EMBO Rep. 13, 750-758 (2012).
- Yu, Y., et al. Kindlin 2 promotes breast cancer invasion via epigenetic silencing of the microRNA200 gene family. Int J Cancer. , (2013).
- Meves, A., Stremmel, C., Gottschalk, K., Fassler, R. The Kindlin protein family: new members to the club of focal adhesion proteins. Trends Cell Biol. 19, 504-513 (2009).
- Siegel, D. H., et al. Loss of kindlin-1, a human homolog of the Caenorhabditis elegans actin-extracellular-matrix linker protein UNC-112, causes Kindler syndrome. Am. J. Hum. Genet. 73, 174-187 (2003).
- Hart, R., Stanley, P., Chakravarty, P., Hogg, N. The kindlin 3 PH domain has an essential role in integrin LFA-1-mediated B cell adhesion and migration. J. Biol. Chem. , (2013).
- Liu, J., et al. Structural basis of phosphoinositide binding to Kindlin-2 pleckstrin homology domain in regulating integrin activation. J. Biol. Chem. 286, 43334-43342 (2011).
- Qu, H., et al. Kindlin-2 regulates podocyte adhesion and fibronectin matrix deposition through interactions with phosphoinositides and integrins. J. Cell Sci. 124, 879-891 (2011).
- Yates, L. A., et al. Structural and Functional Characterisation of the Kindlin-1 Pleckstrin Homology Domain. J. Biol. Chem. 287, 43246-43261 (2012).
- Bouaouina, M., et al. A conserved lipid-binding loop in the kindlin FERM F1 domain is required for kindlin-mediated αIIbβ3 integrin coactivation. J. Biol. Chem. 287, 6979-6990 (2012).
- Goult, B. T., et al. The structure of the N-terminus of kindlin-1: a domain important for AlphaIIbBeta3 integrin activation. J. Mol. Biol. 394, 944-956 (2009).
- Yates, L. A., Fuzery, A. K., Bonet, R., Campbell, I. D., Gilbert, R. J. Biophysical Analysis of Kindlin-3 Reveals an Elongated Conformation and Maps Integrin Binding to the Membrane-Distal β-Subunit NPXY motif. J. Biol. Chem. 287, 37715-37731 (2012).
- Anthis, N. J., Campbell, I. D. The tail of integrin activation. Trends Biochem. Sci. 36, 191-198 (2011).
- Harburger, D. S., Bouaouina, M., Calderwood, D. A. Kindlin-1 and -2 directly bind the C-terminal region of beta integrin cytoplasmic tails and exert integrin-specific activation effects. J. Biol. Chem. 284, 11485-11497 (2009).
- Tadokoro, S., et al. Talin binding to integrin beta tails: a final common step in integrin activation. Science. 302, 103-106 (2003).
- Garcia-Alvarez,, et al. Structural determinants of integrin recognition by talin. Mol. Cell. 11, 49-58 (2003).
- Perera, H. D., Ma, Y. Q., Yang, J., Hirbawi, J., Plow, E. F., Qin, J. Membrane binding of the N-terminal ubiquitin-like domain of kindlin-2 is crucial for its regulation of integrin activation. Structure. 19, 1664-1671 (2011).
- Ussar, S., Wang, H. V., Linder, S., Fassler, R., Moser, M. The Kindlins: subcellular localization and expression during murine development. Exp. Cell Res. 312, 3142-3151 (2006).
- Bialkowska, K., et al. The integrin co-activator Kindlin-3 is expressed and functional in a non-hematopoietic cell, the endothelial cell. J. Biol. Chem. 285, 18640-18649 (2010).
- Moser, M., Nieswandt, B., Ussar, S., Pozgajova, M., Fassler, R. Kindlin-3 is essential for integrin activation and platelet aggregation. Nat. Med. 14, 325-330 (2008).
- Lefort, C. T., et al. Distinct roles for talin-1 and kindlin-3 in LFA-1 extension and affinity regulation. Blood. 119, 4275-4282 (2012).
- Moser, M., et al. Kindlin-3 is required for beta2 integrin-mediated leukocyte adhesion to endothelial cells. Nat. Med. 15, 300-305 (2009).
- Schmidt, S., Nakchbandi, I., Ruppert, R., Kawelke, N., Hess, M. W., Pfaller, K., Jurdic, P., Fassler, R., Moser, M. Kindlin-3-mediated signaling from multiple integrin classes is required for osteoclast-mediated bone resorption. J. Cell Biol. 192, 883-897 (2011).
- Malinin, N. L., et al. A point mutation in KINDLIN3 ablates activation of three integrin subfamilies in humans. Nat. Med. 15, 313-318 (2009).
- Svensson, L., et al. Leukocyte adhesion deficiency-III is caused by mutations in KINDLIN3 affecting integrin activation. Nat. Med. 15, 306-312 (2009).
- Liu, Y., Zhu, Y., Ye, S., Zhang, R. Crystal structure of kindlin-2 PH domain reveals a conformational transition for its membrane anchoring and regulation of integrin activation. Protein Cell. 3, 434-440 (2013).
- Liu, J., et al. Structural basis of phosphoinositide binding to kindlin-2 protein pleckstrin homology domain in regulating integrin activation. J. Biol. Chem. 286, 43334-43342 (2011).
- Zhao, Y., Chapman, D. A., Jones, I. M. Improving baculovirus recombination. Nucleic Acids Res. 31, (2003).
- Berrow, N. S., et al. A versatile ligation-independent cloning method suitable for high-throughput expression screening applications. Nucleic Acids Res. 35, 45 (2007).
- Nettleship, J. E., Assenberg, R., Diprose, J. M., Rahman-Huq, N., Owens, R. J. Recent advances in the production of proteins in insect and mammalian cells for structural biology. J Struct. Biol. 172, 55-65 Forthcoming.
- Wasilko, D. J., et al. The titerless infected-cells preservation and scale-up (TIPS) method for large-scale production of NO-sensitive human soluble guanylate cyclase (sGC) from insect cells infected with recombinant baculovirus. Protein Expr. Purif. 65, 122-132 (2009).
- Yates, L. A., Fleurdépine, S., Rissland, O. S., De Colibus, L., Harlos, K., Norbury, C. J., Gilbert, R. J. Structural Basis for the activity of a cytoplasmic RNA terminal uridylyl transferase. Nat. Struct. Mol. Biol. 19, 782-787 (2012).
- Sainsbury, S., Ren, J., Saunders, N. J., Stuart, D. I., Owens, R. J. Crystallization and preliminary X-ray analysis of CrgA, a LysR-type transcriptional regulator from pathogenic Neisseria meningitidis MC58. Acta Crystallogr. Sect. F. Struct. Biol. Cryst. Commun. 64, 797-801 (2008).
- Zhao, Y., et al. Regulation of cell adhesion and migration by Kindlin-3 cleavage by calpain. J. Biol. Chem. 287, 40012-40020 Forthcoming.
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유