
Method Article
Micromanipulation 기법 전체적 역동성의 분석 및 Cytoskeletal 레 귤 레이 터의
요약
어떻게 마이크로 및 photomanipulation 기술이 FRAP photoactivation 활성화 운동 매개 변수 결정 및 마이그레이션 내 단백질의 spatiotemporal 역학 셀 설명 합니다. 실험 정보 subcellular 역학과 운동 성 레 귤 레이 터 또는 기본 말라 골격의 회전율을 포함합니다.
초록
단백질의 spatiotemporal 역학 조사 하는 것은 다양 한 문맥에서 그들의 기능적인 중요성을 밝힐 수 있다. 이 문서에서 그것은 photobleaching (FRAP) 및 photoactivation 기술을 후 논의 어떻게 형광 복구 단백질 subcellular 위치에의 spatiotemporal 역학 공부를 사용할 수 있습니다. 우리는 또한 이러한 기술을 간단한 결정 걸 cytoskeletal 레 귤 레이 션 및 세포 운동에 연결 하는 다양 한 매개 변수를 사용 하는 방법을 보여줍니다. 또한, 세포의 microinjection 또한 설명 (잠재적으로 앞 또는 상기 photomanipulation 기술을 보완) 대체 치료로 셀에 translocated 단백질의 트리거 즉각적인 효과를 형태와 기능 Micromanipulation 단백질 주입 또는 원형질 막 투과 약물 또는 cytoskeletal 억제제의 로컬 응용 프로그램과 같은 단일 세포 및 subcellular 세포 행동에 주어진된 치료의 즉각적인 결과 기록 하는 강력한 도구 역할을 할 수 있습니다. 수준입니다. 이것은 궁 행 여기 lamellipodial 셀 가장자리 돌출의 즉시 유도 의해 재조합 Rac1 단백질의 주입에 의해 4 분의 1 세기 전에 설립. 또한, 향상 된 녹색 형광 단백질 (EGFP)의 매출을 결정 하기 위한 프로토콜 제공-VASP는 걸 필 라 멘 트 중 합 효소 두드러지게 B16-f 1 셀의 lamellipodial 끝 축적 FRAP 고용 하 고 포함 하 여 관련 데이터 분석 및 커브 피팅입니다. 우리 또한 EGFP 태그 β-걸 표현 하는 세포에 의해 exemplified lamellipodial 말라 네트워크 중 합 속도 추정 하는 것에 대 한 지침을 제시. 마지막으로, 지시 급속 한 필 라 멘 트, lamellipodia, 돌기의 끝에와 같은 사용 하 여 어셈블리 photoactivation의 사이트에서 말라 설립 뒤 셀 세포질 내에서 말라 단위체 이동성의 비율을 조사 하는 방법에 대 한 주어 집니다. 접근 한다. 이러한 프로토콜의 구성 요소 또는 말라 골격의 레 귤 레이 터에 제한 하지만 유사한 패션 spatiotemporal 역학 및 다양 한 다른 subcellular 구조 또는 기능적 단백질의 기능에 찾아보기 쉽게 확장 될 수 있습니다. 컨텍스트입니다.
서문
단백질 및 살아있는 세포에서 다른 분자의 spatiotemporal 역학 모니터링 세포와 분자 생물학의 여러 분야에서 필수적인 도구가 되고있다. 고급 형광 현미경 검사 법 기술로 형광 공명 에너지 전달 (무서 워) 무서 워-형광 일생 (FLIM 무서 워), 이미징 또는 FRAP, photobleaching (플립)과 photoactivation 형광 손실 뿐만 아니라 많은 다른 사람 허용 시간적 및 공간적 확산의 속도 론 및 지역화 셀1,2에 있는 다른 단백질의 결정 뿐 아니라 단백질-단백질 상호 작용, 구조적 변화의 추적. FRAP 및 photoactivation 기술을, 특히 말라 골격 및 세포 이동의 레 귤 레이 터를 조사 하기 위한 널리 적용 됩니다. 이 기술은 또는 microinjection3, 같은 추가 micromanipulation 기술로 적용 될 수 있다 고 붙일 표시 된 단백질의 표현을 포함. 그들은 걸 부자 구조 단백질 협회의 활동의 추정 filopodia 또는 lamellipodia, 초점 접착4, 단백질의 회전율 등 셀 마이그레이션에 관련 된 또는 말라 네트워크5분기 있습니다. 그들은 또한 lamellipodial 걸 중 합 속도, 결정 단위체 말라 cytosol, subcellular 말라 단위체 전 polymerizing에서 튀어나온 걸 필 라 멘 트에의 속도 내에서 분산의 평가 가능 하 게 lamellipodia6, 그리고 다른 매개 변수입니다.
FRAP 시각화 및 원래 악 셀7에 의해 1970 년대에 개발 된 살아있는 세포 내에서 단백질의 이동성을 측정 하는 방법 이다. 붙일 표시 된 단백질으로 채워진 셀, 내 관심 (ROI) 영역 정도 높은 강도, 시간의 짧은 시간 동안이 지역에 존재 fluorophore 분자의 표백을 충분 한 레이저에 노출 됩니다. 표백, 붙일 표백, 하는 동안 투자 수익 밖에 있는 단백질 분류 확산 되며 시간이 지남에 photobleached 분자의 변위를 일으키는 그들의 spatiotemporal 역학에 따라 표백된 지역에 침투. 형광 표백된 지역에 복구 속도가 주어진된 분자의 확산의 속도 및 크기를 포함 하 여 다양 한 요인에 따라 달라 고 내는 상 상속의 회전율이 표백된 구조를 연결 하는 코스. 따라서, 수용 성 단백질 단백질 초점 유착 등 구조와 밀접 하 게 관련 된 동안 보급을 통해 급속 하 게 표백된 ROI 내에서 형광의 복구 중재할 것 이다, 그들의 형광 복구 것입니다 회전율 시간이 더 오래 있을 것 이다 모두 구조 관련 분수의 단백질과 분리 협회 활동의 녹는 분수의 보급에 따라 달라 집니다. 형광 복구는 일반적으로 취득 하 고 계량 형광의 사전 표 백제 농도의 초기 수준에 도달할 때까지. 그러나,이 경우 초기 형광 강도의 일부 확산에 의해 보충 될 수 있는 소위 움직이지 분수에 속한다 또는 모바일을 구성 하는 분자의 대부분에 비해 매우 느린 속도로 보충은 발생 하지 않습니다. 분수입니다. 단백질 회전율의 비율을 확인 하려면 FRAP 곡선 시간이 지남에 형광 복구의 범위를 나타내는 생성 됩니다. 이러한 복구 곡선에서 단백질 복구의 평균 반 시간을 계산할 수 있습니다. 생성 함으로써 평균 FRAP 데이터 및 따라서 수학적 분석의 곡선에 맞는, 그것은 또한 모바일 분수의 평균 이직 률 구성 분자의 한 균질 인구의 합성 또는 그것은으로 구성으로 추정할 수 차동 속도에서 선회 하는 분자의 두 개 이상의 모집단 양적 접근에 의해 단백질 이직 률을 추정, 뿐만 아니라 lamellipodia에 photobleached 지역의 복구를 추적 허용할 수 있다 또한 역행 흐름, 돌출, lamellipodial 운동 성 매개의 정확한 정량화 그리고 말라 중 합 속도입니다. 따라서, FRAP 살아있는 세포의 구조 내에서 다양 한 매개 변수를 평가 하기 위한 적용 되는 다양 한 도구를 구성 합니다.
Photoactivation는 단백질 또는 지정 된 세포 위치에서 발생 하는 분자의 확산을 추적 하는 데 사용 되는 방법입니다. 기술을 사용 하 여, 예를 들어, 야생-타입 녹색 형광 단백질 (GFP), 패터 슨, 리 핀 코트 요 슈워츠8, 그것의 형광에 노출 시 매우 증가 될 수 있는 방식에 돌연변이 의해 처음에 개발의 변종 자외선 (UV) (약 400 nm; 여기, 405 nm). 패터 슨 외에 의해 설명된대로., 중립 페 놀 및 생산 약 397에 주요 흡 광도 피크 음이온 phenolates의 혼합된 인구로 서 존재 하는 야생-타입 GFP chromophores nm와 475에 사소한 한 nm, 각각. 자외선과 단백질의 조사, 시는 인구 photoconversion, 음이온 양식으로 변화를 겪 습. 488 흥분 때 nm, photoconverted/photoactivated 단백질 형광, 활성화 사이 구별 및 비-활성화 GFP 높은 본질적인 배경 형광 때문에 연습 부족에 3 증가 전시. 그러나, 배경 강도 감소 GFP 시퀀스 (히스티딘 대체 위치 203)에 단일 아미노산 돌연변이 도입 하 여 달성 되었습니다. Photoactivatable-GFP (PA-GFP)는 자외선과 방사선 조사 시 이후에 488 nm 빛에 의해 흥분 하는 때 거의 약 증가 작은 피크의 흡 광도 있는 뜻깊은 감소 특징 이다로 알려져는 결과 T203H 돌연변이 체 따라서, PA GFP 태그 단백질의 overexpression 유포의 결정 및 분자의 운동 성 세포 내에서 허용 하는 널리 사용 되 접근 이다. 우리는 이전 적용 PA GFP 태그 말라 말라 단위체 cytosolic 영역에서의 분산의 비율을 결정 하는 돌기에 cytosol, 뿐만 아니라 그들의 설립 속도 내에서 그들의 이동성의 뿐만 아니라 탐사를 허용 lamellipodial 걸 네트워크6. 최근 문학 소설, 사진-컨버터블 단백질 원리에서 사용 될 수 있는 유사한 방식, 설명 합니다 하지만 사진 변환 하기 전에 이미 볼 수 있도록 잠재적인 이점을 은닉. 형광 단백질의이 그룹에 대 한 예로 Dendra2 및 mEos29,10,,1112.
이 문서에서는, 우리는 단백질 microinjecting 셀 방법론 설명. 우리 더 어떻게이 기술은 결합 될 수 있다 FRAP, 말라 골격 규제 및된 운동 성, photobleaching 단백질에 의해 어떻게 FRAP 곡선 및 하프 타임 모바일 분수의 복구의 파생 될 수 있습니다 설명 합니다. 또한, 우리는 FRAP 기술을 사용 하 여 lamellipodial 네트워크의 걸 중 합 속도 결정 하는 방법을 예를 제공 합니다. 우리는 또한 지침 및 단위체 말라의 cytosolic 이동성 및 lamellipodia로 말라 설립의 결정 하는 데 사용할 수 있습니다 photoactivation 실험을 수행 하는 방법에 대 한 도움말을 제공 합니다. 이러한 기술은 물론, 하지 않습니다만 제한 걸 골격 구성 요소를 추적 하지만 잠재적으로 필요한 적당 한 적응 또는 최적화, 널리 적용할 수 다른 세포 유형 또는 다른 단백질, 구조, 조사 하 고 매개 변수입니다.
프로토콜
1. Coverslip 세척 및 살 균
- 15 mm (직경) 커버 유리 (1 호) 40 mL 37 %HCl 60 mL 100%의 혼합물을 포함 하는 500 mL 플라스 크에 담가 EtOH (이상의 100 coverslips 솔루션을 세척 하는 100 mL 당).
참고: 갓 구입 하는 경우에 coverslips 청소 되어야 한다 엄격 하 게 그들의 표면에 세포를 뿌리기 전에. 이 때문에 그들은 거시적 표시 되지 않습니다 하지만 효율적으로 접착 및 적절 한 라이브 세포의 확산을 방해 수 있는 윤활제의 박막을 포함 될 수 있습니다. 반면 같은 영화는 산 이나 염기를 포함 하는 솔루션을 효율적으로 제거 될 수 있다 (참조 피셔 외. 13) 우리가 일상적으로 사용 하 여 위에서 설명한 산/알코올 혼합. - 회전 통에 30 분 동안 커버 유리를 포함 하는 플라스 크를 흔들어. 자유롭게, swirled 하지만 자주 위반을 피하기 위해 충분히 천천히 커버 유리를 허용 하는 속도 선택 하십시오. 재사용 하는 경우 깨진된 유리 조각을 제거 하는 솔루션을 여과.
- 살 균 물 200 mL 이상 포함 하는 플라스 크를 커버 유리를 전송 하 고 산 성 냄새는 멸종 될 때까지 반복 해 서 물을 교체 하는 동안 회전 통에 품 어. 몇 시간 동안 여러 세척은 HCL EtOH 흔적의 완전 한 제거 하는 것이 좋습니다.
- 필터 종이에 개별 커버 유리를 건조.
- 10 cm (직경) 페 트리 접시의 하단에 커버 유리 필터 종이, 고 열으로 덮여 장소 건조 소독. 이 함께 커버 유리 하면 압력가 마로 소독을 하지 마십시오.
2. 셀, Transfection, 및 Coverslips에 뿌리기의 치료
- B16-F1 마우스 흑색 종 세포에 따라 표준 셀 문화 DMEM (4.5 g/L 포도 당)에 10% 태아 종 아리 혈 청, 2mm 글루타민, 그리고 1% 페니실린-37 ° C, 7% CO2에서 스 성장.
- DMEM (4.5 g/L 포도 당) 포함 10% 태아 둔감 한 혈 청, 1 mM 나트륨 pyruvate, 1 x MEM 비필수 아미노산에 따라 표준 셀 문화 (37 ° C, 7% CO2에서 조직 문화 인큐베이터) microinjections에 대 한 NIH3T3 fibroblast 세포 성장 2 mM 글루타민, 및 1% 페니실린-스.
- Transfections, 3 cm (직경) 플라스틱 접시에 10 cm 요리와 1:5 비율로 통로에 100% 합류를 B16-F1 셀을 성장.
- 같은 날, B16-F1 셀 적어도 6 h에 대 한 준수 허용 되었다, 후 500 ng/photoactivatable PA-GFP-말라의 요리 또는 β-말라 EGFP 태그 플라스 미드 DNA transfect. MCherry 인코딩 벡터와 PA-GFP-말라의 공동 transfections, 3 cm 접시 당 플라스 미드 DNA의 1 µ g의 총을 섞는다.
- Transfection 시 약 (자료 테이블) B16-F1 셀 transfect 3 cm 요리 150 m NaCl 포함 500 m의 200 µ L를 섞어 DNA의 ng 150mm transfection 시 약의 1 µ L을 포함 하는 NaCl의 200 µ L로 구성 (즉, DNA (µ g): 시 (µ L) 비율 1: 2의 사용 되었다).
- 실 온 (RT)와 drop-wise 셀을 포함 하는 3 cm 접시에 피펫으로 20 분 transfection 혼합물을 품 어. 부드럽게 혼합 하 고 37 ° C, 7% CO2에서 밤새 품 어 접시를 소용돌이 친다.
- 50 mM Tris, pH 7.4와 150 m m NaCl 포함 된 laminin 코팅 버퍼를 준비 합니다.
- B16-F1 셀에 대 한 15 m m 커버 안경 laminin (laminin 코팅 버퍼에 25 µ g/mL)의 150 µ L를 확산 하 여 코트와 실시간에서 1 시간에 대 한 품 어 NIH3T3 세포 fibronectin 솔루션 (인산 염 버퍼 식 염 수 (PBS)에 25 µ g/mL)와 커버 안경 코트와 실시간에서 1 시간에 대 한 품 어
- PBS, 커버 유리 laminin 또는 fibronectin incubated 세척 후 PBS를 발음 하 고 transfected 세포의 2 개 mL를 추가.
- Transfected B16-F1 셀 씨앗 (1시 30분에 confluent 접시에서 비율), transfection laminin 코팅 coverslips에 다음날에. NIH3T3 fibroblasts 씨앗 (1시 20분에 confluent 접시에서 비율) fibronectin 코팅 coverslips에.
- 현미경 검사 법 전에 37 ° C에서 조직 문화 인큐베이터에서 하룻밤 laminin 또는 fibronectin 코팅 커버 유리에 확산에 셀 수 있습니다. 또한, 현미경 실험 세포는 적어도 2-3 h에 대 한 확산 수는 같은 날에 시작할 수 있습니다.
3입니다. 현미경 이미징 챔버의 조립
- 열 전도성 RC-26 알루미늄 챔버 (그림 1a) 현미경 이미징 사용 하 여. 주사기 (그림 1b)를 사용 하 여 플라스틱 마감재 개통의 컨투어 주위 실리콘 윤활제 얼룩.
- 그림 1(c) 상공에 셀 사이드 최대 커버 글라스를 놓습니다.
- 플라스틱 마감재는 coverslip와 챔버 사이 보안 씰을 커버 글라스 위에 놓습니다. (그림 1의d)를 유출 하는 매체를 피하기 위해 챔버에 슬라이딩 클램프를 조이는 여 (대각선)을 피하기 위해 coverslip 파손 플라스틱 마감재를 수정.
- 중앙 지역으로 피 펫 37 ° C 사전 온수 현미경 매체. Autofluorescence에 따라서 현미경 검사 법에 대 한 최적화 된 매체에 대 한 설명 위에 있지만 F12-햄 대신 DMEM, 또한 포함 된 20 mM HEPES CO2 (그림의 부재에 있는 세포의 배양과 문화 매체로 동일한 제조 법을 사용 1e).
- 열 검출기 챔버의 지정 된 슬롯에 삽입 하 고 37 ° C (그림 1f)의 일정 한 온도 유지 하는 TC 324B 자동 온도 조절기 챔버의 전극 연결.
- 목표에 침수 기름의 작은 방울을 배치 하 고 챔버 상단에 배치.
- 설치 하는 동안 온도에서 복구 하 고 현미경 매체에 적응할 수 있도록 적어도 10-30 분 동안 셀 챔버를 품 어.
- 현미경 검사 법을 시작 하기 전에 중간 구성 요소와 중간 증발 때문에 혈 청의 부적절 한 농도 피하기 위해 챔버 (약 800 µ L)의 중앙 저수지에서 문화 매체를 교체 합니다. 개방 챔버와 함께 장기간된 현미경 세션 증발 매체의 일상적인 변화 필요 합니다.
4. microinjection 절차
- 코트는 coverslips, 셀, 준비 하 고 위에서 설명한 대로 이미징 챔버를 조립.
- 녹여 주입을 순화 된 단백질의 약 수 (일반적으로 10 µ L 이하로) 적절 한 microinjection 버퍼와 희석.
참고: 버퍼 구성 단백질 및 셀 형식에 따라 달라질 수 있습니다 하지만 대부분 셀 PBS와 주입을 좋아하지 않아 하는 6.95 사이 8.00 pH를 사용 하 여 PBS를 사용 하지 않도록 알아서. - Rac1 microinjection에 대 한 100 m m NaCl, 50 mM Tris HCl pH 7.5, 5 mM MgCl2, 1mm DTT 포함 하는 버퍼를 준비 합니다. 마그네슘2 + 이온 작은 GTPase 안정성을 위해 필수적입니다.
주: 단백질 농도 일반적으로 0.1-1 사이 다 mg/mL (최대 2 mg/mL), 단백질에 따라, 실험, 및 세포의 유형 유형. - 해당 하는 경우 주사 하기 전에 바늘 흐름의 존재를 확인할 수 있습니다 하 고 실험 후 성공적인 주사의 설명서는 단백질 해결책에 형광 염료, 불활성 dextran (0.5 µ g/mL, 70 kDa) 등을 추가 합니다.
참고: 여기에 실험 다음 주입된 Rac1의 역학을 직접 형광 단백질의 라벨에 따라 가능한 것만 겨냥 하지. 형광 염료 나 형광 단백질 융합 단백질의 결합은 가능 하지만 피할 여기 특히 가족 Rho GTPase Rac1 같은 작은 단백질의 신호 전달 기능 방해의 위험을 비호 (20 kDa). - 바늘 microinjection 모 세관에 있는 경우를 막힘으로 이어질 수 있는 단백질 집계 제거 하려면 적어도 30 분 동안 10000 x g에 단백질 해결책을 원심.
- 유연한 피 펫 팁/microloader 팁을 사용 하 여 뒷면에서 주입 혼합물의 1 µ L 함께 microinjection 바늘 (모 세관 microinjection)를 로드 합니다.
- 공기 방울 바늘 팁에 있다면, 그들을 제거 하기 위해 부드럽게 바늘 기지를 누릅니다. 니 들 막힘을 일으킬 수 바늘 팁의 건조를 피하기 위해 빠르게 진행 합니다.
- 신중 하 게 조정 하는 micromanipulation 장치에 바늘 홀더. 위상 대비 영상, 바늘 선적의 앞에 거꾸로 한 현미경을 사용 하 여 확인 하는 경우 현미경 콘덴서를 방해 하지 않고 바늘을 위아래로 이동 충분 한 공간이 있다.
- 바늘 홀더에 microcapillary를 속이 고, 시 세포 배양 매체에 바늘 끝을 translocating 전에 microinjection 압력 장치를 사용 하 여 바늘에 압력 (20-50 hPa 배경 압력) 적용 됩니다.
참고: 때 바늘은 매체에 압력을 활성화 것 이다 매체 모 세관 힘에 의해 빨려 고 따라서 관심의 솔루션의 주입을 금지. - (낮은 배율 목표를 사용 하 여 촉진) 보기의 필드에 바늘을 위치. Microinjection 실험에 대 한 40 X 건조 목표는 여기 사용 되었다.
- 거시적 수직 위치에서 가운데를 기준으로 목표 렌즈의 (이 가속화 될 것으로 바늘 끝을 찾는) 바늘 팁을 배치 합니다. 단계 대조 광학 현미경을 사용 하 여 보기의 필드, 광학 평면에 상대적인 수평 평면에 바늘 팁 잘-셀 레이어 위에 이동.
참고: 바늘은 처음 보기의 필드에 그림자로 나타나고 포커스 면 다음 팁을 시각화 하기 위해 조정 될 수 있다. 바늘의 팁 발견 되 면 점차적으로 광학 평면 뒤에 셀 레이어 가까운 위치까지 바늘 팁 낮은. - 형광 dextran을 사용 하 여 형광 채널을 전환 하 여 바늘 흐름을 확인 하 고 지속적인 "배경" 흐름을 얻기 위해 압력 장치를 사용 하 여 흐름을 조정.
참고:이 문서에서 설명 셀을 터치를 통해 플라즈마 멤브레인을 통해 침입 하 여 중재 수동 주입, 표면 및 지속적인 바늘 흐름 중 바늘 팁의 움직임. 이 더 높은 셀 숫자 나중 세포 인구로의 주입에 대 한 적절 한 주입 이벤트 기간 동안 프로그램된 바늘 바늘을 낮추고 압력 증가 함께 자동 주입 장치에서 고유 해야 합니다. 분석입니다. 여기 설명 하는 방법은 하기 전에, 도중, 그리고 후에 microinjection 시간 경과 현미경 검사 법에 의해 단일 세포 분석을 위해 최적화 되어 있습니다. - 관심의 셀 찾기 및 점차적으로 셀 위에 바늘.
- 때 microinject 준비, 낮은 초점에서 세포를 유지 하면서 micromanipulator 조이스틱의 정밀한 피니언을 사용 하 여 셀 perinuclear 지역으로 점차적으로 바늘.
- Microinjection, 셀, 침투 또는 현미경 설치에 매우 부드러운 탭 과도 막 파열 원조 충분 할지도 모른다 세포의 원형질 막에 터치 부드럽게 합니다.
참고: 바늘 끝에 흰 점이 원형질 막;와 접촉 시간이 표시 됩니다. 막 파열, 다음 바늘 팁은 다시 봉인, 셀에 주입 솔루션의 부드러운 흐름을 동반. - 셀에 표시 되 자 마자 주입 프로세스를 중지 (0.3 이내 s) 매체에 바늘 끝을 이동 하 여. 형광 dextran을 사용 하 여, 성공적인 주사 형광에 의해 즉시 문서화 될 수 있다.
- 원하는 경우 시간 경과 이미지 수집 전이나 후에 microinjection를 시작 합니다.
참고: 약물 이나 억제제의 로컬 응용 프로그램 실행할 수 있습니다, 여기에 모든 단계에서 제외 하 고는 microinjection 이벤트 당. 로컬 응용 프로그램에 대 한 활성 분자의 확산 유량 압력에 의해 제어 되 고 형광, 문서화 될 수 고 바늘 팁 원하는 높이에 배치 될 수 있습니다. 로컬 응용 프로그램의 참조 하십시오 예를 들어, Rottner 및14 또는 Kaverina 외. 15 - Microinjection에 따라 단백질의 효과 발생 될 때까지 기다립니다. 다른 단백질 및 예상된 결과 따라 보육 시간 달라질 수 있습니다. 작은 GTPase Rac1 lamellipodium 형성의 응답 수 1 분 이내 시작 또는 더 적은, 하지만 완벽 하 게 (그림 1g, h)를 개발 하는 것을 평균 약 10-15 분 걸립니다.
- 판사는 microinjection 다음 세포의 생존 능력.
참고: 부적절 하거나 유해한 주사 자주 동반 하는 비 특정 셀 지 철회 또는 원형질 막 파열 세포 손상을 일으킬 수 있습니다.- 평면 세포 지역에서 주사에 대 한 발생할 수 있는 위쪽 및 아래쪽 플라즈마 멤브레인 통해 파고 하지 마십시오.
참고: 사출 볼륨은 최소한으로 유지 되어야 한다 (이상적으로 < 세포 볼륨의 5%)와 femtoliter 범위에 일반적으로 있을 것입니다. 필요한 주입 볼륨 또한 변화 농도에 의해 통제 될 수 있다 하지만 단백질, 농도 대 한 유의 > 2 mg/mL 자주 바늘 막힘으로 인해 허무 하 게 될 수 있습니다. 그러나,이 또한 품질과; 순화 된 단백질의 동작에 따라 다릅니다. 예를 들어, 걸 붙일 결합의 주입 농도 의존 하 고 피할 수 없는 합 바늘 팁에 의해 복잡 하 게 하 고 그래서 그것 드물게 오늘 실행 됩니다 (참조 작은 외. 16)입니다.
- 평면 세포 지역에서 주사에 대 한 발생할 수 있는 위쪽 및 아래쪽 플라즈마 멤브레인 통해 파고 하지 마십시오.
- 전에, 동안, 나는 microinjection의 효과 후, FRAP 또는 photoactivation에서 수행할 수 있습니다 같은 셀 (섹션 5 및 6 참조).
5. 졸라 절차
- Transfect 세포 유형의 관심 (B16-F1 셀에 여기) 플라스 미드 DNA 관심 붙일 태그가 단백질 인코딩 (여기, β-말라의 EGFP 태그 버전 사용 되었다). 시드 laminin 코팅 coverslips (단계 2.10)에 셀입니다.
- 조립 영상 실 (제 3).
- 다음 설정을 사용 하 여 lamellipodial 지역 photobleaching에 대 한: 65 mW 레이저 전원 (실험 설치 및 레이저 소스에 따라 변수); 10 픽셀 레이저 빔 직경; 1 ms 표 백제 망설임 시간/픽셀; 500 GFP 노출 시간; 1, 500 밀리초 시간 간격입니다. 이 논문에서 실험 결과 100 X 1.4NA apochromatic 목표와 수행 했다.
- Photobleached 지역의 치수에 정확성을 보장 하기 위해 레이저 캘리브레이션을 수행 합니다. 교정, 이전 어떤 셀/형광 신호를 부족 한 영역을 보기의 필드를 이동 하 고 디스플레이에 그림을 관찰 합니다.
- 객관적인 확대 각각 확대 버튼을 클릭 하 여 선택 하 고에서 레이저 전력 (3-5 mW)를 줄일 수는 "패널 | 강도"메뉴입니다. Visiview 소프트웨어 (v2.1.4)에 수동 보정을 시작 하려면 선택 합니다 있는 "구성 | FRAP"메뉴를 클릭 합니다 있는" 보정 | 수동 조정"메뉴입니다. 레이저는 날카로운 점으로 구별 될 수 있다 확인 하십시오. 만약에 아닙니다, 촛점 하거나 레이저 하드웨어를 조정 합니다.
- 수동으로 미리 결정된 소프트웨어 X Y 좌표에는 레이저를 안내 하 여 보정을 수행 합니다. 이 소프트웨어 특히 현재 배율에 대 한 사용자 정의 영역에 레이저를 표적 하는 방법을 지시 합니다.
- 레이저를 트리거링 하기 전에 GFP로 전환 하 고 이미지/시간 경과 수집을 시작 합니다.
- 수동으로 영역에 디스플레이 보면서 GFP 채널에 photobleached 됩니다 그립니다.
- 적어도 3-4 프레임 이미지 수집의 개시 후 405 nm 레이저의 수동 방 아 쇠에 의해 photobleaching을 시작 합니다. Photobleaching 이전 프레임을 획득 하는 것이 나중에 데이터 분석에 이미지의 필요 합니다.
6. Photoactivation 절차
참고: 소프트웨어, 현미경 설치 및 설정, 레이저 파워 제외한 비슷합니다 FRAP에 대 한. Photoactivation, FRAP, 비교는 중요 한 차이점에에서는 photobleaching, 동시에 photobleaching 없이 PA-GFP를 활성화를 사용 해야 합니다에 대 한 고용 405nm 레이저 전력 보다 크게 낮은 그것.
- 공동 transfect 세포 유형의 관심 (B16-F1 여기 세포; 단계 2.5 참조) 플라스 미드 DNA와 인코딩 PA-GFP-말라와 다른 붙일 표시 된 단백질 (예: mCherry 또는 mCherry-Lifeact).
참고: 대부분의 경우, mCherry-긍정적인 세포도 됩니다 PA-GFP-말라 벡터, photoactivation 이전 GFP 채널에서 볼 일반적으로 후자에 대 한 긍정적인. MCherry-긍정적인 세포는 또한 PA GFP에 대 한 긍정적인 기회를 강화 하려면 mCherry:PA의 1:2 transfection 비율-GFP-말라. 이 프로토콜, mCherry을 표현 하는 세포의 90% 이상 PA-GFP-걸의 성공적인 활성화 표시. - 시드 B16-F1 셀 laminin 코팅 coverslips (단계 2.10)에.
- 조립 영상 실 (제 3).
- Photoactivation 실험을 시작 하기 전에 필요한 경우, 선택 된 목표 (5.4-5.6 단계)에 대 한 레이저 캘리브레이션을 수행 합니다.
- GFP/488 nm 이미지 수집 (실험 설계)에 따라 500 ms 노출 및 1, 500 밀리초 시간 간격을 설정 합니다.
-
듀얼-채널 또는 3 채널 시간 경과 영화 "파장 시리즈" 사각형을 표시 하 고 채널에서 원하는 수를 선택 하 여 인수에 대 한 소프트웨어 설정을 조정는 "취득 | 파장"메뉴입니다. 시간 경과 영화 위상 대조와 GFP 채널 인수 하는 것이 좋습니다.
- 선택적으로, 또한 포함 mCherry 채널; 그러나, 너무 많은 빛으로 세포를 노출 photodamage를 유도 수 있습니다. 이것은 Oxyrase17, 등 산소 청소부와 효과적인 치료 셀 챔버 씰링을 필요로 하지만 피할 수 있었다.
- MCherry 채널에서 transfected 세포를 찾아.
- 레이저를 실행 하기 전에 이미지/시간 경과 수집을 시작 하 고 수동으로 디스플레이 보면서 단계 대비 채널에 photoactivated 수 영역을 그립니다.
- Photoactivation 405 nm 레이저의 수동 트리거에 의해 시작 (강도 설정에서 5 ~ 15 mW 사이 "패널 | 강도"메뉴) 이미지 수집 개시 후 최소한 3-4 프레임.
7. 데이터 분석 및 FRAP 결과의 프레 젠 테이 션
참고: 제시 메서드는 VASP는 접착 사이트와 lamellipodia를 돌기의 끝이이 경우에 동적 말라 어셈블리의 사이트에 축적 하는 단백질의 매출 조사를 위해 사용 됩니다. 우리는 lamellipodium 끝에 그것의 회전율 분석 하지만 VASP 또는 다른 단백질 및 다른 subcellular 구획의 회전율을 조사 분석의 동일한 원리를 적용할 수 있습니다.
- 오픈 시간 경과 영화에서에서 파생 된 Visiview Metamorph 소프트웨어에. 이 문서에서는, Metamorph v7.8.10 사용 되었다.
- 수동으로 Metamorph 각각 영역을 표시 하 여 photobleached 지역에 대 한 강도 값을 파생 합니다. Photobleached 면적의 일부 또는 전체를 커버 하 고 프레임에 그것의 위치를 수동으로 조정 하는 lamellipodium의 끝에 모양에서 (즉, 가장자리 돌출 하는 경우), 필요한 경우의 lamellipodial 농도 변경 내용을 추적 하기 위해서는 그리기 팁 변위 중 해당 구성 요소입니다.
- 배경 및 photobleaching 수집의 교정, 지역 내부 및 외부 셀을 분석 합니다. 그림 2는 측정된 농도의 대표 지역에 대 한 참조 하십시오.
- 동안 ROI 선택 메뉴를 사용 하 여 Metamorph에 그것의 강도 값을 추출 "측정 | 지역 측량 "입니다. "구성" 메뉴에서 "경과 시간" 및 "평균 강도" 옵션이 선택 되어 확인 하십시오. "로그 열기"를 클릭 하 고 "동적 데이터 교환"을 선택 합니다. 열고 Excel 스프레드 시트를 Excel에 붙여 Metamorph 값을 다시 "로그 열기" 버튼을 클릭 "OK"를 클릭 합니다.
참고: 이러한 값은 형광 복구 곡선을 생성 하는 데 사용 됩니다. - Photobleached 지역 (photobleaching 전에 지역 강도를 정규화)의 lamellipodium 끝에 형광 복구 곡선 생성, 다음 식을 적용 됩니다.
 공식 1
공식 1
어디: 졸라Tn 은 다음 photobleaching; 관심의 각 프레임에 대 한 photobleached 지역 강도 테네시 밖으로 photobleaching; 다음 관심의 각 프레임에 대 한 셀 (배경) 외부 촬영 지역 강도 기능테네시 는 photobleaching (시간이 지남에 수집 photobleaching에 대 한 정상화 하는 데 사용); 다음의 각 프레임에 대 한 지역 농도 안에 평균 2 FRAPT-1 photobleaching; 전에 photobleached 지역 강도 T-1 이다 photobleaching; 하기 전에 셀 (배경) 외부 촬영 지역 강도 그리고 기능T-1 photobleaching 전에 관심의 각 프레임에 대 한 지역 농도 안에 평균 2. - 관심의 각 시간 프레임에 대 한 조사를 모든 시간 프레임을 포함 하는 형광 복구 커브를 공식 1 을 사용 합니다. 시간의 길이 엄격 하 게 조사 단백질에 의존 합니다. 때 알 수 없는, 단백질의 이직 률을 얻으려고 예비 실험을 수행 합니다.
- 복구의 하프 타임을 계산, 시그마 플롯 (12 절), 형광 복구 곡선에는 해당 시간 (초)의 값을 붙여 및 수행을 사용 하 여 적합 한 곡선에서 "동적 맞는 마법사 | 최대 지 수 상승"도구입니다. 모노 지 수 선택 (단일, 매개 변수 3) 또는 (더블, 매개 변수 4) bi 지 수에 맞는 최고의 곡선 따라 기능.
- 모노 지 수 함수에 대 한 다음 수식을 사용 하 여:
 공식 2
공식 2
- 이중 지 수 함수에 대 한 다음 수식을 사용 하 여:
 식 3
식 3
- 매개 변수 "b"와 "d" 복구의 하프 타임을 계산 하기 위해 Excel로 시그마 플롯공식 2 ( 식 3)에서 파생 된 붙여 넣습니다. 다음 방정식을 적용:
 방정식 4
방정식 4
또는 방정식 5
방정식 5
- 모노 지 수 함수 결과 맞는 정확한 곡선, 방정식 4만 적용.
- 모노 지 수 함수에 맞게 좋은 곡선 귀착되 지 않는다, 식 4 와 방정식 5모두 해결 하 여 bi 지 수 수식을 적용. 결과 두 반으로 대표 하는 두 개의 서로 다른 단백질 분수 회복의 시간을 고려:는 급속 하 게 그리고 천천히 교환 분수, 각각.
8. 졸라 Lamellipodial 걸 중 합 속도 의해 결정
- Lamellipodial 걸 중 합 속도 결정, transfect EGFP 태그 β-말라, B16-F1 셀 photobleach lamellipodial 지역 (단계 5.9) 1.5 s 시간 간격 및 500 ms GFP 노출 사용 하 고.
- Metamorph에서 Visiview에서 취득 시간 경과 영화 열고 사용 목적에 따라 픽셀 / µ m 비율 보정은 "측정 | 거리 보정"도구입니다.
- 시간 경과 영화를 재생 하 고 선으로 얇은 판자 쪽으로 거꾸로 흐르는, lamellipodial 형광 복구는 얇은 판자에 도달 했습니다 때 후방 흐름을 추적할 수 있습니다 더 이상 프레임에 중지.
- µ M는 lamellipodium의 팁과 복구 된 형광의 뒤에 거리를 측정 합니다. 이 거리는 역행 흐름 및 돌출 거리의 합에 해당합니다.
- 또는 역행 흐름에서 돌출을 구분 하는 photobleaching 전에 선 한 프레임 lamellipodial 팁을 표시 합니다. 사용 기준으로 선 포인트 프레임 photobleaching; 시 lamellipodium 팁의 원래 위치를 참조 하 참조점 돌출 거리와 역행 흐름을 측정 하기 위해 사용할 수 있습니다.
- 시간 (초) 형광 복구 photobleaching 후 발생 하는 데 필요한 note 시간 또는 프레임 속도에서 수동으로 계산 수 있습니다 통해 Metamorph에 의해 시각화는 "측정 | 지역 측정"도구입니다.
- (8.4-8.6 단계에서 Metamorph 측정에 따라 일부 방정식 매개 변수)와 다음과 같은 방정식을 사용 하 여 걸 중 합 속도 파생:
 공식 6
공식 6
어디 걸 중 합 속도가 µ m/분에서, 역행 흐름 거리 µ m에는, lamellipodial 돌출 거리는 µ m, 이며 시간 초.
9. 단백질 보급 및 Photoactivation에 따라 이동성 분석
참고: 여기에 제시 된 방법을 시각화와는 cytosol 통해 단백질 보급의 정량화에 의해 볼 수 있듯이 PA-GFP를 융합 하는 걸의 photoactivation를 채용 하 여 말라 단위체 이동성의 분석을 설명 합니다.
- Lamellipodial 지역 내의 축적 뿐 아니라 cytosolic 영역에서 photoactivatable 걸의 보급을 측정를 사용 하 여 Metamorph 지역은 시간이 지남에 강도 결정 ( 그림 3는 ): cytosolic photoactivated 지역 (PA); lamellipodial 지역, 어떤 photoactivated에 단백질 (Lam); 시간이 지남에 축적 하는 것으로 예상 된다 (밖으로) 배경 형광의 정규화에 사용 되는 셀 외부 지역.
- 말라는 cytosol 내에서 이동성을 결정할 때 (참조 그림 3는, 지역 R1-R5) 별개의 cytosolic 영역을 측정 합니다. 참고 photobleaching 수집 활성화 시 초점-그리고 결국 세포 형광 증가로 인해 비슷한 방식 FRAP,으로 확인할 수 없습니다.
- 모든 지역에 대 한 강도 값에서에서 전송 Metamorph를 Excel 스프레드시트에 단계 7.4에에서 설명 된 대로.
- Photoactivatable 걸 photoactivation의 cytosolic 영역 또는 lamellipodial 지역 (둘 다 시 0% 강도의 cytosolic photoactivated 지역으로 표현 하는) 내에서 법인의 그것의 속도의 변위 속도 검사 단계 9.3 데이터에서 형광 커브를 생성. 다음 방정식을 적용:
 공식 7
공식 7 공식 8
공식 8
어디: PA테네시 cytosolic photoactivated 지역의 photoactivation; 다음의 각 프레임에 대 한 강도 램테네시 는 lamellipodial 지역의 photoactivation; 다음의 각 프레임에 대 한 강도 테네시 밖으로 photoactivation; 다음 셀 (배경)의 각 프레임에 대 한 외부 촬영 영역의 강도 PAT-1 photoactivation; 전에 cytosolic photoactivated 영역의 강도 램T-1 photoactivation; 전에 lamellipodial 지역의 강도 T-1 지역 photoactivation; 하기 전에 셀 (배경) 외부 촬영의 강도가 이다 PAT0 는 cytosolic photoactivated 영역의 강도 시 0 (즉, 첫 번째 프레임 photoactivation 후에); 그리고t 0 0 (즉, photoactivation 후 첫 번째 프레임) 시 셀 (배경) 외부 촬영 영역의 강도 이다. - 필요한 경우 데이터의 더 나은 시각화에 대 한 각 후속 프레임에서 photoactivation 후 첫 번째 프레임의 강도 빼서 0 강도 곡선 정상화.
참고: 다음 분석 방법 (단계 9.6-9.8) 또한 photoactivated 걸 내는 cytosol의 cytosolic 분산을 계산할 수 있습니다. - 연속적으로 활성화 지역에서 distally 위치 cytosolic 영역에 대 한 농도 측정 합니다.
- 시간 0에 % 강도의 photoactivated 지역으로이 지역의 농도 나타내는 방정식 8, lamellipodial 농도 각 cytosolic 투자 수익에 대 한 강렬으로 대체 됩니다 어디를 적용 합니다. 크기와 개수는 지역 측정 셀 크기와 분산 거리에 따라 달라질 수 있습니다.
- Photoactivated 단백질 각 cytosolic 영역 침투의 속도 대 한 가능한 값을 파생, 시그마 플롯 (섹션 7에서 FRAP 분석 비슷합니다) 시간 및 각 지역에 대 한 형광 강도 증가의 곡선의 값을 붙여, 를 사용 하 여 방정식 2 및 파생 고원에 도달 형광 강도의 하프 타임 식 4 . 다른 실험적인 그룹 간의 t1/2 값을 비교 합니다.
결과
그림 1 g h 단계 이전 NIH3T3 fibroblast 세포 이미지와 10 분 후 microinjection은 복잡 한 파와의 상호 작용을 통해 lamellipodia 형성 유도 능력이 작은 Rho 가족 GTPase Rac1의 표시. 셀은 먼저 그것의 생존 능력 및 형태, 예를 들어, lamellipodia의 부족을 확인 microinjection (그림 1g), 전에 시각화 됩니다. 10 분 후 microinjection에서 셀 명확 하 게이 치료에서 예상 되며 성공적인 주입 (그림 1h) 나타냅니다의 형태를 변경 했습니다.
단순 하 고 선명도, 우리는 다음 또한 microinjected 하지가지고 셀에서 FRAP 및 photoactivation 분석을 위한 모범적인 결과 제공.
Lamellipodium 끝에 VASP EGFP 태그의 회전율의 분석 그림 2a ~ f에 표시 됩니다. Note VASP 또한 초기 및 초점 유착, 셀 내부18,19에 작고 길쭉한 점을 대상. 끝에 분명 VASP 누적 된 lamellipodial 영역의 형광 강도 표백 이었고, 중, 전과 후는 lamellipodium 돌출 전달으로 표백 투자 수익의 윤곽에 따라 각 기간에 대 한 측정. 표백된 EGFP VASP 단백질 비 표백 분자가이 사이트에서 재생 되는,으로 형광의 점진적인 회복 (그림 2b) 관찰 된다. 이 방식 하 고 사전 표 백제 농도 (1로 표현)으로 정규화 FRAP 복구 곡선 그림 2c에서 볼 수 있습니다. Photobleaching 효율과 다를 수 있습니다 표백이 예제에서 t 0에 있는 값에서 결정 하기 전에 값의 약 20%는 (photobleaching 후 첫 번째 프레임). 형광의 증가 고원 예 전에 표백 형광의 약 80%에 도달 합니다. 초점 접착 등 실험의 시간 과정 동안 정적 구조 사전 표 백제 강도와 고원 형광의 차이 도달 후 복구 움직이지 분수 (IF, 빨간색 화살표 그림 2에서로 정의 됩니다. c, e), 형광의 양을 사이 표백 하 고 완전 한 회복의 시간 복구 하는 반면 모바일 분수 (녹색 양방향 화살표 그림 2c, e)로 정의 됩니다. Lamellipodium 팁 여기 분석 등 동적으로 변화 하는 구조에서 IF의 범위 수 있습니다 뿐만 아니라 움직이기 힘 드는 분자를 대표 하지만 EGFP VASP 강도이 의존 알려져 있다 또한 돌출 속도의 감소에서 파생 매개 변수18. 복구의 하프 타임을 계산 하려면 시그마 플롯 (그림 2d)에 맞는 곡선을 만들었습니다. 해결 하는 방정식 2 에서 추출 된 "b" 매개 변수 값은이 경우에, 0.0754 때 적용 로그 함수 (식 4) 9.19의 복구의 결과 약된 반 시간에 (그림 2d , 맨 오른쪽 패널), 이전5에 게시는 상대적으로 평균에 비해 특정 셀이에 빨리. 그것은 그 복구-시간 달라질 수 있습니다 때로는 크게 셀에서 동일한 인구 내의 주목 해야 합니다. 따라서, 대표적인 결과 얻기 위해 적어도 15-20 셀에서 평균으로이 매개 변수를 확인 것이 좋습니다. 분산, 15 셀에서 평균 EGFP VASP 복구의 산술 방법의 정도 설명 하기 위해 각 시간-포인트 생성된 (그림 2e), 그리고 평균 곡선에 맞는 만든 유사한 패션 (그림 2 에 표시 f)입니다.
Lamellipodial 걸 네트워크의 중 합 속도 앞으로 네트워크 돌출 및 역행 흐름의 합으로 구성 됩니다. FRAP β-말라 EGFP 태그와 photobleaching 튀어나온 lamellipodial 지역 (그림 2g)와 (이 경우 F1 B16)에서 transfecting 셀 여 걸 중 합 속도 측정 하는 데 적용할 수 있습니다. Lamellipodial 걸 네트워크 중 합의 분석, 대 한 β-말라 EGFP 태그의 표백 시 형광 회복 시간이 지남에 평가. (이 모든 전면20쪽으로 포인트) lamellipodial 걸 필 라 멘 트의 가시 끝에 말라 단위체 진행의 합으로 네트워크 이며 끊임없이 translocated rearwards 전달, 진행 속도 쉽게 될 수 있습니다. photobleaching에 따라 형광의 편광된 복구 통해 얻은. lamellipodium의 형광 복구 완료 되 면 최대한 빨리 표백된 영역에는 lamellipodium의 뒤쪽 부분과 더 수평으로 배열 된 필 라 멘 트 번들의 낮은 밀도 의해 특징입니다 얇은 판자 사이 전환 영역에 도달 했습니다. 선회는 lamellipodium에서 무엇을 관찰 하는 것 보다 훨씬 더 느리게. 그림 2g에서 볼 수 있듯이 형광 복구 수평 가장자리를 돌출 고 (개별적으로 나타나는 퇴행 성 흐름의 거리 측정을 허용 하는 얇은 판자 쪽으로 거꾸로 흐르는 선으로 구상 될 수 있다 주황색과 빨간색 양방향 화살표로 그림 2g 의 맨 오른쪽 패널에서 각각).
우리는 또한 photoactivation PA-GFP-말라 말라 단위체는 cytosol 내에서 이동성 및 lamellipodia 튀어나온 내에서 그들의 설립의 속도 추적 하을 transfected B16-F1 셀에 적용 했습니다. 그림 3a, b에서 볼 수 있듯이 cytosolic 영역 405 nm 레이저에 노출에 의해 photoactivated, 이미지는 GFP에 인수 했다 하는 동안 채널 모든 1.5 GFP 태그, photoactivated 말라의 분포를 시각화에 대 한 s. 그림 3bcytosolic 지역의 확산 Photoactivated GFP-걸을 볼 수 있습니다. Photoactivated cytosolic 영역에서 형광 강도 감소의 속도 (t0)에서 초기 휘도의 비율로 표시 됩니다 (첫 번째 photoactivation; 후 프레임 그림 3 c). Photoactivated 말라 또한 새로운 걸 단위체의 돌출 하는 동안 걸 필 라 멘 트를 elongating 성장 가시 끝에 추가 됩니다 lamellipodia의 끝에 통합. Lamellipodial 설립의 속도 예상 하려면 우리는 약 5 µ m 폭에서 및 높이; 1 µ m의 2 차원 윤곽선/지역 시간이 지남에 형광 강도 측정 지역 끊임없이 다시 protruded 그것으로 lamellipodium의 끝에 위치 했다. 말라 설립 (t0)에서 photoactivated cytosolic 영역의 형광 강도의 비율로 표현 되었다 (그림 3d). 말라 필 라 멘 트의 연신 율 진행, 새로운 걸 단위체 lamellipodial 앞에 통합 했다. 이러한 걸 단위체의 분수 확률론 단위체 photoactivated을 했다 cytosolic 풀에서 파생 되었다. 첫 번째 20에 lamellipodia에 형광의 급속 한 증가에이 결과 후 photoactivation s. 로 새로운 단위체 퇴행 성 흐름으로는 얇은 판자 쪽으로 필 라 멘 트와 함께 이전 법인된 걸 단위체 흐름 lamellipodial 앞에 추가 되 고 있습니다. 시간, ROI 완전히 채워집니다 형광 단위체 및 형광에 대 지에 도달 하면 (그림 3d). 형광에 있는 점차적인 하락 다음 때, 셀에 걸쳐 photoactivated 단위체의 확산에 따라 관찰, 비 photoactivated 걸 단위체는 점점 추가 되 고 다시 lamellipodial 앞에. 형광이이 감소는 새로운 고원, photoactivated 및 photoactivated 비 단위체 사이 전체 셀에 균형 (데이터 표시 되지 않음)에 도달할 것입니다 발견할 것 이다.
cytosol에 걸쳐 걸 단위체의 이동성 distally ( 그림 3는 색상 코드 영역 표시에 exemplified photoactivated 지역에서 위치 하는 같은 크기의 영역에서 형광 강도 측정 하 여 파생 되었다 R1-R5). 이 지역의 각각에서 형광 강도에서 점차 감소e 그림 3볼 수 있듯이 cytosolically photoactivated 지역, photoactivated 말라 단위체의 일부분으로 된다 점점 희석 비 활성화 (즉, 비-형광 성) 단위체. 또한, 형광의 피크에 도달 하면 나중: 더 먼 측정된 영역은 위치 photoactivated 지역에서이 지역으로 확산 말라 단위체에 필요한 더 이상 시간. 각 지역에 말라 단위체 침투의 정도 대 한 대표적인 값 형광 고원 도달의 하프 타임을 측정 하 여 파생 될 수 있습니다. 더 먼 지역, 더 오래 걸리는 photoactivated 말라, 그것으로 무마 하 고 따라서 더 많은 시간 필요 도달 될 형광 고원 궁극적으로 더 높은 t1/2 값 (그림 3e)로 이어지는.

그림 1 : 챔버 어셈블리 및 microinjection 절차를 이미징. (한) 이미징 약 실 구성 요소입니다. (b) 실리콘 그리스는 신중 하 게 플라스틱 마감재의 오프닝의 주위 발 랐 습. (c)는 coverslip 셀-면 열고 이미징 챔버의 중앙에 배치 됩니다. (d)는 보안 물개는 coverslip 위에 플라스틱 날 인자를 배치 하 여 고 사이드 클램프를 강화 하 여 설정 됩니다. (e) 현미경 중간 챔버 슬롯에 pipetted입니다. (f) 이미징 챔버가 현미경 스테이지에 배치, 열 탐지기 및 전극 미리 37 ° C로 설정 하는 난방 장치에 연결 되 고 셀 현미경 시작 하기 전에 적어도 30 분에 대 한 적응을 사용할 수 있습니다. 이 예제에서 현미경 단계 또한 갖춘 한 micromanipulator microinjections, 수행 하 고 microinjection 바늘 이미징 챔버에 셀 레이어를 취재 하는 매체에 담근. (g)는 NIH3T3 fibroblast 세포 단계 대조 현미경 검사 법에 의해 microinjection 전에 시각 이다. Perinuclear 구획에 적십자 부피가 큰 핵에 가까운 근접 때문에 높은 세포질 영역에 해당 하는 미래 microinjection의 위치를 나타냅니다. (h) 다음 10 분 Rac1와 microinjection, 셀 (녹색 화살표로 표시 됨) 전체 셀 주위 lamellipodia의 저명한 형성에 의해 반응. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 2: FRAP 단백질 회전율 또는 lamellipodial 걸 중 합 속도 결정할 수 있습니다. B16-F1 셀 표시 lamellipodial 지역의 photobleaching 전에 EGFP VASP를 표현 (한) 대표 예. 서로 다른 색된 윤곽 또는 모양 어떤 지역 시간이 지남에 형광 강도 측정을 위한 고려 되었다 나타내는 레이블이 지정 됩니다. 참고 빨간색 윤곽선 레이블 cytosolic 영역 여러 소포 및 셀 표면 주름을 포함 하는 지역에 위치 하는 느낌표와 함께 표시. 이 같은 동적 영역으로 그들은 잠재적으로 부정확 한 결과가 발생 하는 형광의 강한 단기 변동에 의해 특징 이다 형광 참조의 영역을 선택 하는 데 피해 야 한다. (b) Lamellipodial photobleaching 전후 EGFP VASP 표현 세포의 영역입니다. 형광 신호 지역 내에서 photobleaching 자주색으로 표시 된 후의 복구 시간이 지남에 시각 이다. 화살표 걸 필 라 멘 트의 높은 밀도 인해 가능성이 VASP 위한 농축 한 microspike의 끝을 나타냅니다 polymerizing 거기19. (c)는 FRAP의 예를 들어 복구 곡선으로 b 레드 photobleached lamellipodium (보라색 컨투어)의 형광 강도 측정 하는에서 파생 하 고 오른쪽에 녹색 선, 각각, 부동 및 모바일 분수를 나타냅니다. (d) A c (왼쪽된 패널) FRAP 복구 곡선 및 복구 30 시간 (오른쪽 패널)를 파생 하는 데 사용 하는 계산 방법의 예의에 맞게. (e) FRAP 복구의 예 곡선 샘플 인구 내의 가변성의 정도 나타내는 SEM 바 15 셀의 형광 복구 곡선을 평균에서 파생. (f)는 곡선 맞추기 FRAP 복구 곡선 맞는 복구 반 시간 (오른쪽 패널)를 파생 하는 데 사용 하는 계산 방법의 예를 들어 15 셀 (왼쪽된 패널)의 평균에서 파생. (g) lamellipodium lamellipodial 지역의 표백으로 뒤는 lamellipodium에서 형광 복구 시간이 지남에 전후 EGFP 태그 β-걸 표현 B16-f 1 셀의 돌기의 시간 경과 패널. 맨 오른쪽 패널에서 값 돌출에 대 한 측정 및 역행 거리 제공 됩니다 (오렌지와 레드, 각각). 이미지 패널 아래 계산 돌출 및 역행 거리의 합 lamellipodial 말라 네트워크의 중 합 속도 파생 시키는 데 사용 하는 방법 공개. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
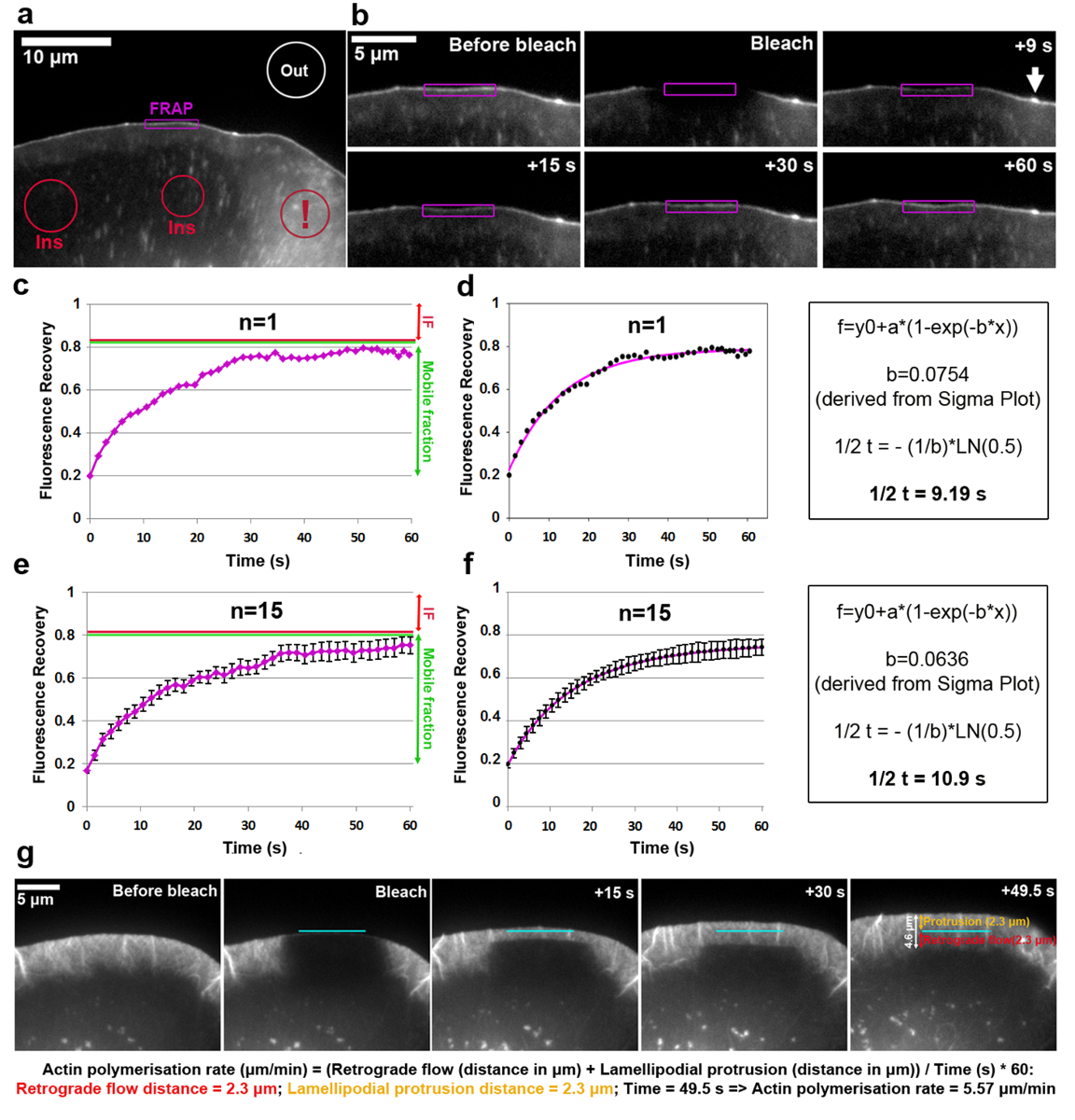{kind=link}

그림 3: 의 PA-GFP-말라 단위체 셀 전체 추적에 대 한 Photoactivation. 빨간색 동그라미 (PA)에 표시 된 대로 cytosolic 영역에서 photoactivation를 실행 하기 전에 PA-GFP-걸 표현 B16-f 1 셀의 (a) A 대표적인 예. 서로 다른 색된 윤곽 어느 지역 시간이 지남에 형광 강도 측정을 위한 고려 되었다 나타내는 표시 됩니다. (b) PA-GFP-걸 다음의 시간적 분포의 그림 photoactivation. Photoactivated 걸 멀리 확산으로 photoactivated, cytosolic 영역 (빨간색 동그라미)에 형광의 점진적 감소를 note. 앞을 네트워크에 그들의 보급으로 인해 photoactivated 걸 단위체 lamellipodia (사이안 지역) 및 거리 및 시간에 따라 패션에 cytosol (다른 색으로 구분 된 지역)에 걸쳐 점차적으로 강화 된다. (c) 대표, 일시적인 감소 photoactivated cytosolic 영역 (빨간색 윤곽선 b에서) 형광의. (d) 측 변화 lamellipodial 지역 (시안색 컨투어 b에서) 형광 강도. (e) 형광 강도의 cytosolic 영역 (b에서 색) photoactivation의 지역에서 가변 거리에 위치 때문에 시간적 변화의 대표적인 곡선. 어떻게 반-시간 형광의 고원 참고 (오른쪽에 범례에 표시)의 거리 증가 지역 photoactivation의 영역에, 주어진 증가와 연관 가능성이 시간에 말라 단위체의 보급에 필요한는 해당 지역입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
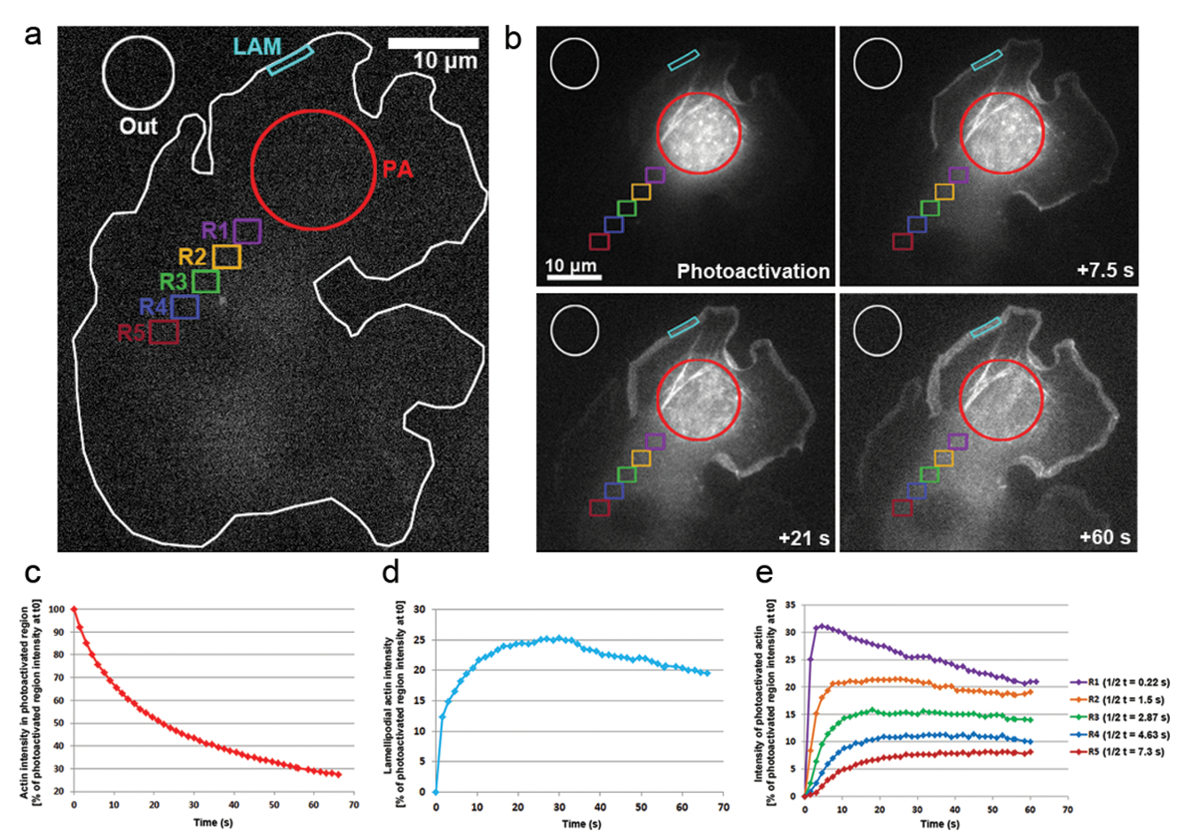{kind=link}
토론
여기 우리는 어떻게 그들이 다른 실험 조건에서 응용 프로그램에 대 한 최적화할 수 있습니다이 문서에 설명 된 기술에 중요 한 단계를 논의.
Microinjection 셀에 외 인 단백질, 억제제, 또는 약물 소개에서 인스턴트 효과 모니터링에 적용 될 수 있는 방법입니다. 그것은 특히 어려운에 단백질의 기능을 결정 하기 위한 유리한 수 transfect 세포 종류를 또는 장기 식 하지 원하는 상황에서. 그것은 특정 종류의 세포 생존에 씨를 뿌리고 기질에 따라 주목 해야 합니다. 가장 내 피, 상피, 또는 섬유 모양의 세포 유형, 심지어 작은 것 들 같은 생선 keratocytes (참조 댕 외. 21 고 앤더슨과 크로스22) 성공적으로 주입 될 수 있습니다. 그러나, 세포 이동의 우수한 모델 시스템을 구성 하지만 알 수 없는 이유로이 유형의 하 층에 주사와 호환 되지 않는 B16-F1 셀 laminin에 시드 같은 예외가 있다. NIH3T3 fibroblast 세포, 우리가 일상적으로 수행 주사 fibronectin 층, 및 추가적인 photomanipulation 기술은 졸라 (photoactivation도, 여기 B16-F1 셀에 대 한 표시) 등이 섬유 아이 세포 (참조에서에서 동일 하 게 수행할 수 있습니다. 예를 들어, Köstler 외. 3). 그것은 또한 고려 되어야 한다 그들의 기능 특성 및 실험 목표에 따라 다른 단백질 다른 양의 변경, 시간을 초에서 변화 하는 시간을 걸릴 수 있습니다. 기법의 장점은 외 인 에이전트의 복용량/농도 플라스 미드 transfection를 사용할 때 보다 예를 들어, 단일 셀 수준에서 더 정확 하 게 제어할 수는 이다. 또한, 단백질의 형광 태그 아니다 붙일 태그가 다른 단백질의 동시 다중 채널 시각화가 필요한 경우 유연성을 증가 시킬 수 있는 셀의 존재를 보장 하는 필요. Microinjection 셀 형태학 또는 (예를 들어, 당 외 골격의 동적 변화에 특정 단백질 또는 단백질 혼합물의 인스턴트 효과 분석 하는 데 특히 유용할 수 있습니다. 21 Arp2/3 복합물 억제제 Arpin에 의해 마이그레이션에 인스턴트 효과의 예). 기술의 단점은 세포 손상 또는 세포 형태에 영향을 미칠 수 있는 그것의 침해입니다. 따라서, microinjections를 수행할 때 중요 한 고려 사항은 세포 생존 능력을 모니터링입니다. 여기 소개 하는 방법은 수동 조작에 의존 합니다. Fibronectin 층에 성장 하는 섬유 아 세포 등 성공적인 주사와 호환 되는지 테스트 하는 조건에서 여기에 설명 된 수동 주입 프로토콜을 사용 하는 근처 100% 성공률; 3이전 게시 비디오 현미경 또는 FRAP를 포함 하 여 정교 하 고 시간이 걸리는 후속 실험으로이 접근을 결합할 때이 필수적입니다. 이 때때로 그를 제외 하지 않습니다, 그리고 개별 셀의 핵 및 세포질, 세포 가장자리 철회 뒤 대비의 갑작스러운 변화에 의해 안전 하 게 인식 될 수 있다 microinjection 이벤트에서 저하 될 수 있습니다. 이러한 드문 경우에 실험은 제외 하 고 따라서 추가 분석에 대 한 간주 되지 않습니다.
그러나, 반 자동 접근은 또한 일반적으로 사용, 예를 들어 빠른 채용 (< 300 ms) 컴퓨터 제어 바늘 낮추는 사출 압력 증가, 일치는 바늘만 각각 이전 각 셀 위에 위치 해야 사출입니다. 반-자동 주사의 성공률은 정의상 단순히 위에서 설명한 수동 방식 보다 낮은 속도 위해 최적화 때문에 뒤에;이 치료를 성공적으로 살아남은 여러 셀의 분석 따라서 개별 셀의 성공적인 주입에 의존 하지 않습니다. 따라서, 단일 세포 분석, 반대 반 자동 주사는 더 나은 적합 몇 백 셀, 예를 들어, 비디오 현미경 낮은 확대율에서 또는 셀 고정 및 얼룩에 의해 주입 효과 분석 합니다. 고용 하는 상세한 접근 방식에 관계 없이 microinjection 분석 결과, 끝점을 구성 하지 않습니다 하지만 다양 한 기법, FRAP 또는 photoactivation3를 포함 하 여 결합 될 수 있다.
현미경 이미징 및 설치 조건에 따라 레이저의 강도, 낙관 한다 FRAP에 의해 단백질 이직 률을 결정할 때 (확대, 목표, 등, 셀 형식, 구조, 및을 위한 형광 단백질 photobleaching)입니다. Note는 최적의 레이저 전력 효율적인 표백와 결합 가능한 최소 photodamage 수축 방지 또는 분석 (예: lamellipodia 또는 filopodia) 또는 세포 수준에서도 피해 구조 철회 완료. 이상적으로, 표백 효율의 70-80% 이상 달성 되어야, 비록 완전 한 표백 있는 경우, 50% 이상의 아무것도 또한 받아들일만 할 수는 단백질의 매우 빠른 회전율에 의해 방해 받을 수 있습니다. 주어진된 구조와 형광 염료에 대 한 최적의 표백 전원 실험적으로 시험 되어야 한다, 그것의 점진적 증가 다음 낮은 레이저 전원에서 시작. 물론, 모든 형광 염료 수 정의 표백 흥분의 절정에 가까운 빛 레이저 (488 nm FITC 또는 EGFP 같은 자주 사용된 녹색 염료에 대 한). 그러나, 근처 UV 레이저와 같은 더 짧은 파장을 가진 레이저 높은 힘을 전달 하 고 따라서 또한 사용할 수 있습니다 일반적으로 사용 되는 염료의 효율적인 표백. 우리가 일상적으로 사용 하는 405 nm 다이오드 레이저 (120 mW)와 후자 (데이터 표시 되지 않음)의 경우 약간 낮은 효율 EGFP와 (예: mCherry), 빨간색 형광 염료의 탈색. 로 405 nm 다이오드 PA-GFP (아래 참조)의 photoactivation도 사용할 수 있습니다, 그것은 최대한 유연성이 시스템을 endows.
B16-F1 셀 구조와 형광 단백질 photobleached 여기에 대 한 65-100 사이 405 nm 레이저 힘은 적용 되었다. 분석할 때 photobleached 지역, 그것은 보존 여부 주어진된 구조는 그것의 원래 모양에서 분석을 통해 기간을 고려 하는 것이 중요. 예를 들어, lamellipodia 끝 단백질의 매출을 분석할 때 주의 해야 lamellipodia의 곡률 크게 시간이 지남에 변경 여부로 곡률에 변화 지역/컨투어 분석 하지 않으면 부정확 한 결과으로 이어질 수도 있습니다. 완전히 각 측정 프레임에서 구조 전체를 포함 합니다. 또한, lamellipodia, microspikes, 등에 포함 된 번들 형광 강도 편차를 발생할 수 있습니다 주목 해야한다. 그림 2b (9 s 시간 프레임에 흰색 화살표)에서 볼 수 있듯이 microspike 같은 구조 측정된 photobleached 지역 옆에 위치 하 고 있습니다 하지만 외부 측정의 기간에 걸쳐 있고 따라서 모든 발생 하지 않습니다. 부정확입니다. 단백질 회전율의 분석, 지역 분석의 위치와 크기를 선택할 때 중요 한 고려는 시간이 지남에 그들의 형광 해야 하지 수 크게 영향을 변화 세포 형태학 또는 요인에 의해 다른 하드을 피하기 위해 수집 photobleaching입니다. 예를 들어, 구조 분석된 구조에 상당한 양적 기여를 제공 이동 해서는 안됩니다 측정 지역의 분석; 중 또한, 나노미터 구조 단백질을 유치 하는 등 관련 없는, 형광 엔터티 분석 중 관심 분야를 입력 하지 해야 합니다. Lamellipodial 걸 중 합 속도 결정 한다 주의 없음 제거 또는 구겨지지 (즉, 위쪽으로 접는) lamellipodia는 분석으로이 강하게 결과의 정확도 영향을 미칠 것입니다. 또한, lamellipodial 말라 합의 과대평가로 이어지는 잠재적으로 lamellipodial 지역 철회 급속 한 후방 전으로 나타날 수 있습니다. 추가 고려는 세포내 정규화 영역 (인수 photobleaching의 교정에 대 한 참조 위치로 촬영 한 것)의 거리에서 photobleaching의 실제 위치는 충분히 커야 한다 피하기 위해 직접 photobleached 영역에 의해 영향을.
PA GFP 태그 구문 photoactivation에 대 한 최적의 조건을 설정할 때, photoactivation 동안 인스턴트 표백 방지 하기 배려를 취해야 한다. 우리의 작업에서 최상의 결과 얻으려면 레이저 힘으로 가져온 EGFP의 표백에 대 한 일반적으로 고용 보다 5-10 배 낮은. Photoactivated 분자의 이미지 수집에 대 한 노출 시간 프레임 사이의 시간 간격 해야 지역 및 photoactivated 될 구조물의 크기를 고려 하 여 최적화와 photoactivated의 잠재적인 이동성 뿐만 아니라 분석 다른 subcellular 위치에 단백질입니다. 형광 이미징의 모든 유형에 서 세포 생존 능력의 유지 보수 순수 관련 결과 얻기 위해 결정적 이다.
원칙적으로, mEos 등 Dronpa 변종12 형광 단백질의 그린-레드 photoconversion 다음 역학의 동등 하 게 강력한 방법 및 subcellular 구조는 lamellipodium 등의 매출 구성 ( 예를 들어, 참조 Burnette 외. 23). PA-GFP 반대로 후자의 방법의 장점은 공동 추가 빨간색 형광 단백질을 표현 하는 필요 없이 두 가지 색상으로 변환 전후 단백질 역동성을 따라 가능성 것입니다. 그러나, 우리의 예비 실험에 대비 변경의 범위와 형광 신호 PA GFP의 photoactivation에 따라 달성의 강도 photoconverted 프로브에 비해 더 큰 아마도 레드와 그린의 우수한 스펙트럼 기능 형광 조사 (데이터 표시 되지 않음) 합니다. 어떤 경우에, lamellipodia 또는 Vaccinia 바이러스 유도 말라 꼬리 같은 셀 가장자리 돌출 말라 필 라 멘 트 매출에 대 한 상세한 연구 지금까지 출판 되었습니다 PA GFP 파생 상품5,,624를 사용 하 여.
Photoactivation에 따라 분석 하는 셀 영역을 고려할 때 여러 가지 요인 취해야 한다 계정에는 (활성화는 cytosol에 따라 셀 가장자리에 말라 단위체의 설립), 여기에 표시 된 특정 한 예를 사용 하 여 설명 하지만 다양 한 유사 과학적인 문제를 확실히 추정 될 수 있습니다. 첫째 때 측정 cytosolically photoactivated 단백질의, 예를 들어, lamellipodial 설립의 고유 실험 조건에서 (같이 Dimchev 그 외 여러분 6) 크기의 cytosolic 영역 및 lamellipodial 가장자리에 그들의 거리 실험 그룹 사이 비교 해야 합니다. 그것은 또한 photoactivating cytosolic 영역, 셀 두께 크면 위치 가까이에 핵을 고려 하는 것이 중요입니다. 두꺼운 세포 지역 활성화 활성화 될 단백질의 분포 homogenously는 cytosol에서 배포 되는 활성화 된 단백질의 더 높은 양을 발생할 수 있습니다. 마지막으로, 활성화 될 단백질의 표정 수준을 확실히 수 높은 변수 개별 셀에. 이러한 모든 고려의 다양성으로 인해 특정 지역에서 활성화 시 얻은 총 형광을 기준으로 셀에 다른 cytosolically 활성화 단백질의 통합 수준을 비교 결정적 이다.
우리는 설명 어떻게 microinjection 세포 형태학에 단백질의 효과 조사 도구로 사용할 수 있다 exemplified이 보여주는 NIH3T3 fibroblast 세포와 microinjected에서 lamellipodial의 강력한 유도 작은 GTPase Rac1. 우리 이전 셀 흉터/웨이브3의 C-터미널 WCA 도메인 microinjected Arp2/3 기능 방해에이 기술을 적용 했습니다. FRAP 또는 photoactivation 같은 다른 분석 실험에 의해 microinjected 세포에 다양 한 매개 변수를 분석할 수 있습니다. 우리는 어떻게 FRAP 및 photoactivation 조사 subcellular 역학과 말라 단위체의 이동성에 대 한 사용할 수 있습니다 설명 했습니다. FRAP 우리의 그룹에 의해 사용 되었습니다 이전5 VASP, Abi, cortactin, cofilin, lamellipodia를 지역화 하 고 상한 단백질, 단백질 또는 elucidating 존재에 초점 접착에서 부품의 매출에 대 한 매출을 조사 하 그리고 Rac의 부재 신호4. 또한, photobleaching EGFP 태그 β-말라5에 의해 달성 될 수 있다 걸 중 합 속도 측정 하지만 존재 하는 다른 방법. 라이브 셀 이미징 호환 프로브 셀룰러 걸 필 라 멘 트, Lifeact25, 같은 라벨 보는 형광 이질성을 추적 고용된6,26일 수도 있습니다. 여기 장점은 β-말라의 overexpression 피할 수 있다, 셀 가장자리 돌출 및 마이그레이션, 증가 수 그리고 이렇게 잠재적으로 방해 실험 질문 또는 특정 분석 결과 (참조 예를 들어, 케이지 외. 26. Peckham 외. 그러나 27). Lifeact 프로브의 뚜렷한 단점 있도록 프로브 매출에만 정보를 제공 하는 셀에 Lifeact 이라고 표시 된 걸 필 라 멘 트 구조의 표백/걸 필 라 멘 트에 바인딩의 활동 그것의 급속 한을 구성 하는, 하지만 하지 말라 필 라 멘 트, 있는 그것은 바인딩25의 회전율. 형광 이질성의 추적 고용 이전6,26 실용적인 타협에서는, 많은 유사한 형광 speckles의 널리 사용 되는 추적에 통합 filamentous cytoskeletal 구조 ( 예: 연어와 워터 맨28참조), 하지만 사용 하는 만큼 정직 하 고 F-말라 EGFP 태그 구조의 FRAP 처럼 정확 하지 않을 수 있습니다. Photoactivation으로 돌기 하는 그것의 이동성 cytosol, 실험적으로 조정된 cytosolic F 걸 레벨6의 맥락에서 전체로 서 lamellipodia, 단위체 말라 설립의 속도 추정을 위한 회사에 의해 적용 되었습니다. 기술은 cytosolic 영역 등 비교적 큰 지역에서 파생 된 이동성 및 단백질의 분포를 검사 하는 경우 유용 합니다. 그러나, 상대적으로 작은 photoactivated 구조;에서 파생 된 단백질의 분포 조사 예를 들어, 성장 콘 형광 분자 활성화의 낮은 숫자, 약한 신호를과 따라서 감도의 부족 될 수 있습니다. Photoactivation 또는 photoconversion 형광 (위 참조)의 잠재적인 대체 기술을 역 투자 수익, 형광 분자에서의 이동성을 추적 하 여 다음을 제외 하 고 전체 셀 photobleaching에 의존 FRAP 포함 될 수 있습니다. 이 지역. 기술은은 단백질의 photoactivatable 버전 overexpressing 필요 하지 않습니다 하지만 항상 잠재적으로 photodamage 등 원치 않는 부작용을 일으키는 레이저 힘의 비정상적으로 높은 복용량에 노출 포함 됩니다.
명확 하 게, photoactivation 및 FRAP 단백질 단위체, 이합체, 또는 심지어 작은 올리고, 이동 하는 그들은 추가 바인딩 파트너와 함께에서 이동 여부 및 구별할 수 없습니다. 형광 상관 분광학 기법29 또는에서 또는, FLIM 무서 워30대신 그런 종류의 정보를 얻을 수 있습니다. 그럼에도 불구 하 고, FRAP photoactivation 직접 관심, subcellular 위치, 또는 세포 유형 공부의 단백질에 관계 없이 셀에 지역 및 글로벌 단백질 역동성을 평가 하기 위해 간단한 접근 구성.
공개
저자는 공개 없다.
감사의 말
우리는 감사 독일 연구 재단 (DFG)에 재정 지원 (Nr. RO2414/5-1 선급을 부여).
자료
| Name | Company | Catalog Number | Comments |
| B16-F1 mouse skin melanoma cells | American Type Culture Collection, Manassas, VA | CRL-6323 | |
| NIH-3T3 cells | American Type Culture Collection, Manassas, VA | CRL-1658 | |
| DMEM 4.5g/L glucose | Life Technologies, Thermno Fisher Scientific, Germany | 41965-039 | |
| Ham’s F-12 medium | Sigma-Aldrich | N8641 | |
| Fetal calf serum (FCS) | PAA Laboratories, Linz, Austria | A15-102 | |
| Fetal bovine serum (FBS) | Sigma-Aldrich, Germany | F7524 | Lot054M3396 |
| MEM Non essential amino acids | Gibco, ThermoFisher Scientific, Germany | 11140035 | |
| L-Glumatine 200mM (100x) | Life Technolgies | 25030-024 | |
| Pen-Strep 5000 U/mL | Life technologies | 15070063 | |
| Sodium Pyruvate (100 mM) | Gibco, ThermoFisher Scientific, Germany | 11360-039 | |
| Laminin | Sigma-Aldrich | L-2020 | |
| Laminin coating buffer | Self-made: 50mM Tris ph7.4, 150mM NaCl | ||
| Fibronectin from human plasma | Roche Diagnostics, Mannheim, Germany | 11 051 407 001 | |
| Jetpei | Polyplus Transfection, Illkirch, France | 101-10N | |
| JetPei buffer | Polyplus Transfection, Illkirch, France | 702-50 | 150mM NaCl |
| PA-GFP-actin plasmid DNA | described in Koestler et al.2008 | ||
| pEGFP-actin plasmid DNA | Clontech, Mountain View, CA, USA | ||
| Rac1 protein for microinjection | Purified as GST-tagged version, and cleaved from GST prior to injection | ||
| Microinjection buffer | Self-made: 100mM NaCl, 50mM Tris-HCl ph7.5, 5mM MgCl2, 1mM DTT | ||
| Dextran, Texas Red, 70,000 MW, Lysine Fixable | Molecular Probes, Thermno Fisher Scientific, Germany | D1864 | |
| Microscope circular cover glasses 15mm, No.1 | Karl Hecht, Aisstent, Sondheim, Germany | 1001/15 | |
| Eppendorf Femtotips Microloader Tips | Eppendorf, Hamburg, Germany | 5242 956 003 | |
| Eppendorf Femtotip Microinjection Capillary Tips | Eppendorf, Hamburg, Germany | 930000035 | |
| Silicone Grease | ACC Silicones, Bridgewater, England | SGM494 | |
| Aluminium Open Diamond Bath Imaging Chamber | Warner instruments | RC-26 | |
| Automatic temperature controller | Warner Instruments | TC-324B | |
| Microscope: Axio Observer | Carl Zeiss, Jena, Germany | ||
| CoolSnap-HQ2 camera | Photometrics, Tucson, AZ | ||
| Lambda DG4 light source | Sutter Instrucment, Novato, CA | ||
| Laser source | Visitron Systems | ||
| Eppendorf FemtoJet microinjector | Eppendorf, Hamburg, Germany | With built-in compressor for pressure supply | |
| Nikon Narishige Micromanipulator system | Nikon Instruments, Japan | ||
| Visiview software v2.1.4 | Visitron Systems, Puchheim, Germany | ||
| Metamorph software v7.8.10 | Molecular Devices, Sunnyvale, CA | ||
| Sigma Plot v.12 | Systat Software Inc. |
참고문헌
- Day, R. N., Davidson, M. W. The fluorescent protein palette: tools for cellular imaging. Chem Soc Rev. 38 (10), 2887-2921 (2009).
- Ishikawa-Ankerhold, H. C., Ankerhold, R., Drummen, G. P. Advanced fluorescence microscopy techniques--FRAP, FLIP, FLAP, FRET and FLIM. Molecules. 17 (4), 4047-4132 (2012).
- Koestler, S. A., et al. Arp2/3 complex is essential for actin network treadmilling as well as for targeting of capping protein and cofilin. Mol Biol Cell. 24 (18), 2861-2875 (2013).
- Steffen, A., et al. Rac function is crucial for cell migration but is not required for spreading and focal adhesion formation. J Cell Sci. 126, 4572-4588 (2013).
- Lai, F. P., et al. Arp2/3 complex interactions and actin network turnover in lamellipodia. EMBO J. 27 (7), 982-992 (2008).
- Dimchev, G., et al. Efficiency of lamellipodia protrusion is determined by the extent of cytosolic actin assembly. Mol Biol Cell. 28 (10), 1311-1325 (2017).
- Koppel, D. E., Axelrod, D., Schlessinger, J., Elson, E. L., Webb, W. W. Dynamics of fluorescence marker concentration as a probe of mobility. Biophys J. 16 (11), 1315-1329 (1976).
- Patterson, G. H., Lippincott-Schwartz, J. A photoactivatable GFP for selective photolabeling of proteins and cells. Science. 297 (5588), 1873-1877 (2002).
- McKinney, S. A., Murphy, C. S., Hazelwood, K. L., Davidson, M. W., Looger, L. L. A bright and photostable photoconvertible fluorescent protein. Nat Methods. 6 (2), 131-133 (2009).
- Gurskaya, N. G., et al. Engineering of a monomeric green-to-red photoactivatable fluorescent protein induced by blue light. Nat Biotechnol. 24 (4), 461-465 (2006).
- Lippincott-Schwartz, J., Patterson, G. H. Photoactivatable fluorescent proteins for diffraction-limited and super-resolution imaging. Trends Cell Biol. 19 (11), 555-565 (2009).
- Kremers, G. J., Piston, D. Photoconversion of purified fluorescent proteins and dual-probe optical highlighting in live cells. J Vis Exp. (40), (2010).
- Fischer, A. H., Jacobson, K. A., Rose, J., Zeller, R. Preparation of slides and coverslips for microscopy. CSH Protoc. 2008, 4988 (2008).
- Small, J. V., Rottner, K., Carlier, M. F. . Actin-based Motility. , (2010).
- Kaverina, I., et al. Enforced polarisation and locomotion of fibroblasts lacking microtubules. Curr Biol. 10 (12), 739-742 (2000).
- Small, J., Rottner, K., Hahne, P., Anderson, K. I. Visualising the actin cytoskeleton. Microsc Res Tech. 47 (1), 3-17 (1999).
- Mikhailov, A. V., Gundersen, G. G. Centripetal transport of microtubules in motile cells. Cell Motil Cytoskeleton. 32 (3), 173-186 (1995).
- Rottner, K., Behrendt, B., Small, J. V., Wehland, J. VASP dynamics during lamellipodia protrusion. Nat Cell Biol. 1 (5), 321-322 (1999).
- Svitkina, T. M., et al. Mechanism of filopodia initiation by reorganization of a dendritic network. J Cell Biol. 160 (3), 409-421 (2003).
- Small, J. V., Isenberg, G., Celis, J. E. Polarity of actin at the leading edge of cultured cells. Nature. 272 (5654), 638-639 (1978).
- Dang, I., et al. Inhibitory signalling to the Arp2/3 complex steers cell migration. Nature. 503 (7475), 281-284 (2013).
- Anderson, K. I., Cross, R. Contact dynamics during keratocyte motility. Curr Biol. 10 (5), 253-260 (2000).
- Burnette, D. T., et al. A role for actin arcs in the leading-edge advance of migrating cells. Nat Cell Biol. 13 (4), 371-381 (2011).
- Humphries, A. C., et al. Clathrin potentiates vaccinia-induced actin polymerization to facilitate viral spread. Cell Host Microbe. 12 (3), 346-359 (2012).
- Riedl, J., et al. Lifeact: a versatile marker to visualize F-actin. Nat Methods. 5 (7), 605-607 (2008).
- Kage, F., et al. FMNL formins boost lamellipodial force generation. Nat Commun. 8, 14832 (2017).
- Peckham, M., Miller, G., Wells, C., Zicha, D., Dunn, G. A. Specific changes to the mechanism of cell locomotion induced by overexpression of beta-actin. J Cell Sci. 114, 1367-1377 (2001).
- Salmon, E. D., Waterman, C. M. How we discovered fluorescent speckle microscopy. Mol Biol Cell. 22 (21), 3940-3942 (2011).
- Machan, R., Wohland, T. Recent applications of fluorescence correlation spectroscopy in live systems. FEBS Lett. 588 (19), 3571-3584 (2014).
- Becker, W. Fluorescence lifetime imaging--techniques and applications. J Microsc. 247 (2), 119-136 (2012).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유