
Method Article
무세포 재구성을 위한 재조합 셉틴 복합체의 정제 및 품질 관리
요약
세포골격 단백질의 체외 재구성은 이러한 단백질의 기본 기능적 특성을 이해하는 데 필수적인 도구입니다. 본 논문은 세포 분열 및 이동에 중심적인 역할을 하는 재조합 셉틴 복합체의 품질을 정제하고 평가하는 프로토콜을 설명합니다.
초록
Septins는 헤테로 올리고머 복합체로부터 세포 골격 필라멘트와 고차 구조를 형성 할 수있는 보존 된 진핵 생물 GTP 결합 단백질 계열입니다. 그들은 다른 세포 골격 구성 요소 및 세포막과 상호 작용하여 이동 및 세포 분열과 같은 중요한 세포 기능에 참여합니다. 셉틴의 많은 상호 작용의 복잡성, 많은 수의 셉틴 유전자(인간의 경우 13개) 및 셉틴이 다른 서브유닛 구성을 가진 헤테로-올리고머 복합체를 형성하는 능력으로 인해 무세포 재구성은 셉틴 생물학의 기초를 이해하는 데 중요한 전략입니다. 본 논문은 먼저 2단계 친화성 크로마토그래피 접근법을 사용하여 재조합 셉틴을 그의 헤테로올리고머 형태로 정제하는 방법을 설명한다. 그런 다음 셉틴 복합체의 순도와 무결성을 확인하는 데 사용되는 품질 관리 프로세스가 자세히 설명되어 있습니다. 이 공정은 네이티브 및 변성 겔 전기 영동, 음성 염색 전자 현미경 및 간섭계 산란 현미경을 결합합니다. 마지막으로, 음성염색전자현미경과 형광현미경을 이용하여 셉틴 복합체의 중합능을 확인하는 과정에 대한 설명이 주어진다. 이것은 초파리 셉틴 헥사머뿐만 아니라 다양한 이소 형태의 septin_9 포함하는 고품질의 인간 셉틴 헥사 머 및 옥타 머를 생산할 수 있음을 보여줍니다.
서문
세포 골격은 고전적으로 액틴 필라멘트, 미세 소관 및 중간 필라멘트 1로 구성된 3 성분 시스템으로 설명되었지만 최근에는 셉틴이 세포 골격1의 네 번째 구성 요소로 인정되었습니다. 셉틴은 진핵생물2에서 보존되는 GTP 결합 단백질 계열입니다. 셉틴은 세포 분열3, 세포-세포 부착4, 세포 운동성5, 형태형성6, 세포 감염7, 세포 극성의 확립 및 유지8와 같은 많은 세포 기능에 관여합니다. 중요한 기능에도 불구하고 셉틴이 그러한 과정에 어떻게 관여하는지는 잘 이해되지 않습니다.
단백질의 셉틴 계열은 단백질 서열 유사성2에 따라 여러 하위 그룹 (분류에 따라 4 개 또는 7 개)으로 세분됩니다. 상이한 서브패밀리의 구성원은 회문 헤테로-올리고머 복합체를 형성할 수 있으며, 이는 필라멘트의 빌딩 블록이며, 이는 차례로 번들, 링 및 메쉬웍스 1,9,10,11,12와 같은 고차 구조로 조립됩니다. 더 많은 분자 복잡성은 상이한 스플라이스 변이체의 존재로부터 발생하며, 예를 들어 인간 SEPT9가 있으며, 여기서 상이한 스플라이스 변이체13,14,15의 특정 기능에 대한 증거가 존재한다. 추가적으로, 헤테로올리고머의 길이는 종 및 세포 유형에 의존한다. 예를 들어, Caenorhabditis elegans septins는 사량 체16을 형성하고, Drosophila melanogaster septins는 헥사 머 17 (그림 1A), 사카로 미세스 cerevisiae 셉틴은 옥타 머 18을 형성하고, 인간 셉틴은 헥사 머 및 옥타머19 (그림 1A)를 형성합니다. 동일한 서브패밀리로부터의 셉틴 이소형, 스플라이스 변이체 및 번역 후 변형된 셉틴이 복합체에서 서로를 대체하는 능력 및 상이한 크기의 헤테로-올리고머의 (공존)존재는 상이한 헤테로-올리고머 복합체12의 세포 기능을 묘사하는 것을 어렵게 만들었다.
셉틴의 또 다른 흥미로운 능력은 세포의 많은 결합 파트너와 상호 작용하는 능력입니다. Septins는 간기 및 세포 분열20,21,22 동안 원형질막과 막 세포 소기관에 결합합니다. 분열하는 세포에서 셉틴은 세포질 분열23,24,25 및 액틴 및 미오신 26,27과 협력합니다. 세포질 분열의 후기 단계에서, 셉틴은 중체 이탈28에 대한 수송 (ESCRT) 시스템에 필요한 엔도솜 분류 복합체를 조절하는 것으로 보인다. 또한, 액틴 피질에 위치한 셉틴과 간기 세포 29,30,31에서 세포의 액틴 스트레스 섬유의 증거도 있습니다. 특정 세포 유형에서, 셉틴은 또한 미세소관 세포골격(32,33)에 결합하고 조절한다.
이러한 모든 기능으로 인해 셉틴은 연구하기에 매우 흥미로운 단백질 시스템이지만 도전적인 시스템이기도 합니다. 많은 수의 셉틴 서브 유닛 (스플 라이스 변이체를 계산하지 않고 인간의 13 개 유전자2)과 동일한 서브 패밀리의 셉틴 서브 유닛이 서로를 대체하고 다른 크기의 헤테로 올리고머를 형성 할 가능성의 조합은 유전자 조작에 의한 특정 셉틴의 세포 기능에 대한 결론을 도출하는 것을 어렵게 만든다. 또한, 셉틴의 다중 상호 작용은 세포 골격 또는 막 구성 요소를 대상으로 하는 약물34 와 같은 일반적인 연구 도구의 효과를 해석하는 것을 어려운 작업으로 만듭니다.
이러한 상황을 극복하는 방법은 셉틴의 시험관 내 (무 세포) 재구성으로 세포 연구를 보완하는 것입니다. 시험관 내 재구성은 특정 서브 유닛 조성 및 길이 18,35,36,37을 갖는 단일 유형의 셉틴 헤테로 올리고머의 분리를 허용한다. 그런 다음이 복합체는 셉틴38,39,40의 기본 구조적 및 물리 화학적 특성을 발견하기 위해 단독으로 또는 모델 생물막 11,41,42, 액틴 필라멘트 10,27 또는 미세 소관32,36과 같은 원하는 파트너와 함께 제어 된 환경에서 연구하여 특성을 해독 할 수 있습니다. 상호 작용.
따라서 다양한 셉틴 복합체를 효율적으로 정제하는 신뢰할 수 있는 방법은 셉틴 연구에 필수적입니다. 그러나 동일한 프로토콜을 사용하더라도 다른 정제는 다른 활성/기능 또는 무결성을 가진 단백질을 제공할 수 있습니다. 효소와 같은 상업적으로 이용 가능한 단백질의 경우, 기능성 및 효소 활성이 신중하게 검증된다43. 세포골격 또는 셉틴과 같은 구조 단백질에 대한 신중한 품질 관리를 구현하는 것은 어려울 수 있지만 실험실 간 실험을 비교할 수 있도록 하는 것이 중요합니다.
이 논문은 대장균 세포에서 단일 또는 이중 시스트로닉 구축물을 포함하는 두 벡터의 동시 발현(표 1)을 기반으로 고품질 재조합 셉틴을 이종 올리고머 형태로 정제하는 강력한 방법을 설명합니다. 이 방법은 그의6-태그된 셉틴과 Strep-II-태그된 셉틴을 모두 포함하는 셉틴 헤테로올리고머를 포획하기 위한 2단계 친화성 크로마토그래피 접근법으로 구성됩니다(그림 1B,C). Iv et al.10에 처음 기술된 이 프로토콜은 초파리 셉틴 헥사머 11,27,35, 인간 셉틴 헥사 머 10 및 상이한 천연(이소형 1, 3 및 5)10,32 또는 돌연변이된 SEPT9 이소폼 32를 함유하는 여러 인간 셉틴 옥타머를 정제하는 데 사용되었습니다. . 또한, 정제된 셉틴의 품질을 평가하기 위한 일련의 기술에 대한 설명이 제공된다. 먼저, 셉틴 서브 유닛의 완전성과 정확한 화학 양 론은 변성 전기 영동 및 투과 전자 현미경 (TEM)을 사용하여 확인됩니다. 그런 다음 정확한 분자량의 헤테로 올리고머의 존재와 복잡한 불안정성을 나타내는 단량체 또는 더 작은 올리고머의 존재를 간섭계 산란 현미경(iSCAT)을 통한 기본 전기영동 및 질량 광도법으로 검사합니다. 마지막으로, 마지막 단계는 형광 현미경 및 TEM을 사용하여 셉틴의 중합 활성을 평가하는 것으로 구성됩니다.

그림 1: 정제 전략 . (A) 인간(왼쪽)과 초파리 (오른쪽) 세포에 존재하는 셉틴 헤테로올리고머의 도식. 숫자는 표시된 그룹의 셉틴 하위 단위를 나타내고 P는 땅콩을 나타냅니다. 인간 SEPT9는 그의 이소형일 수 있다. 셉틴 서브 유닛은 비대칭 모양을 가지며 인간 육각 체 위에 각각 NC 및 G로 표시된대로 두 개의 별개의 인터페이스 인 NC : NC 및 G : G 인터페이스와 길이 방향으로 연결됩니다. (ᄃ,씨) (B) 인간 셉틴 헥사머 및 (C) 옥타머에 대해 표시된 2단계 크로마토그래피 전략의 개략도. H는 그의 태그를 나타내고 S는 스트렙-II 태그를 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
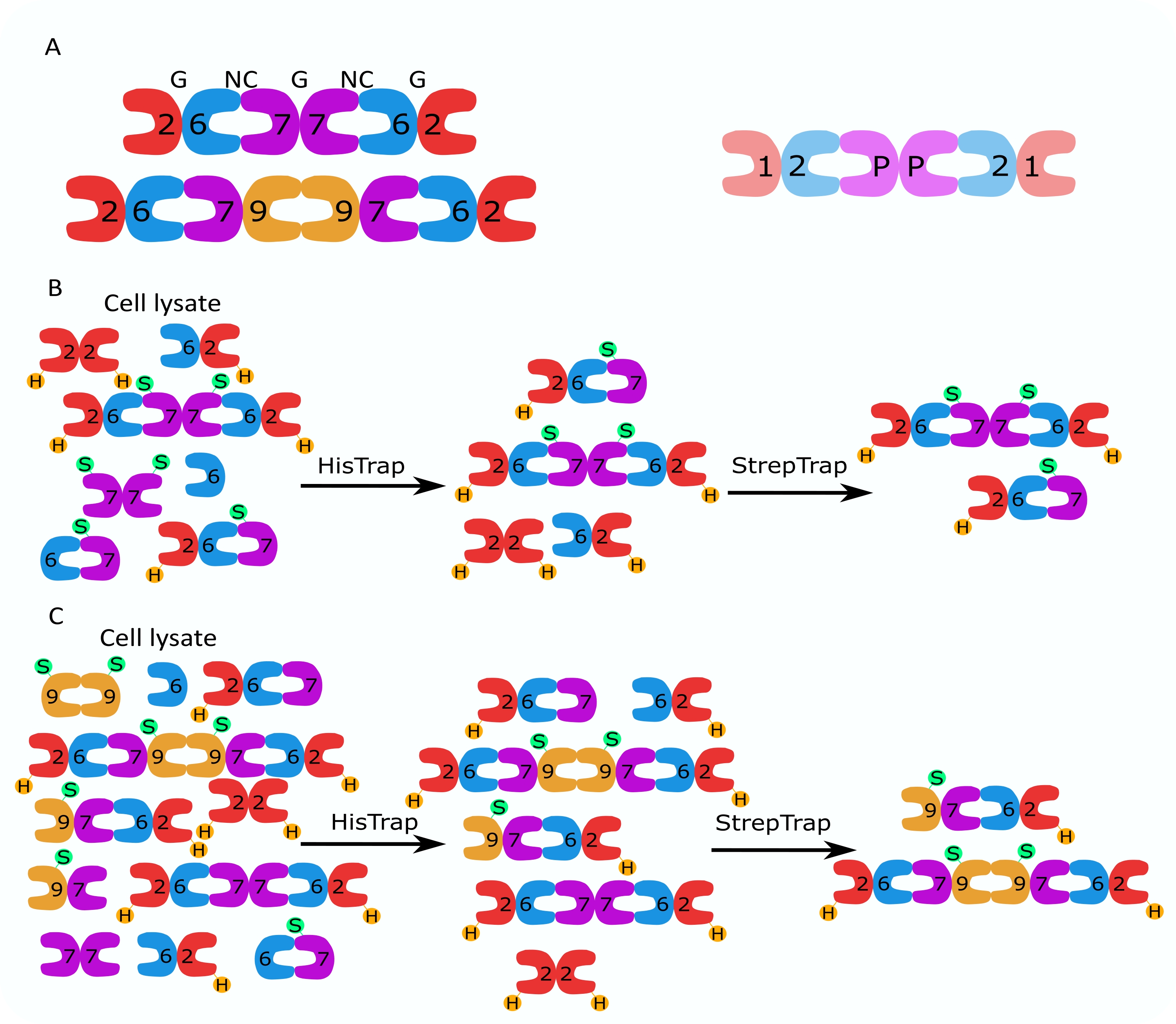{kind=link}
프로토콜
1. 셉틴 헤테로올리고머의 정제
- 박테리아 세포와 발현 벡터의 공동 형질전환
- 발현에 사용될 하나의 pnEA 및 하나의 pnCS 플라스미드44의 조합을 선택한다. 셉틴 헤테로-올리고머10,35의 원하는 서브유닛 조성과 형광 태깅이 필요한지 여부에 따라 조합을 선택하십시오.
참고: C-말단 태그가 지정된 단량체 슈퍼폴더 GFP(msfGFP)-태그가 지정된 SEPT2(인간 셉틴의 경우) 또는 msfGFP- 또는 단량체 강화 GFP(mEGFP)-DSep2( 초파리 셉틴의 경우)가 여기에 사용됩니다(표 1). - 각 플라스미드 1μL(~1ng/μL)를 유능한 BL21 대장균 세포 100μL에 피펫팅하고 얼음에서 20분 동안 배양합니다.
- 세포를 42 ° C의 수조에 40 초 동안 놓은 다음 즉시 얼음 위에서 3 분 동안 배양합니다.
- 0.9mL의 리소제니 브로스(LB) 배지를 세포 현탁액에 추가하고 37°C에서 1시간 동안 세포를 성장시킵니다. 100μg/mL 암피실린과 100μg/mL 스펙티노마이신을 포함하는 따뜻한 LB-한천 플레이트에 세포 100μL를 플레이트하고 37°C에서 밤새 배양합니다.
- 발현에 사용될 하나의 pnEA 및 하나의 pnCS 플라스미드44의 조합을 선택한다. 셉틴 헤테로-올리고머10,35의 원하는 서브유닛 조성과 형광 태깅이 필요한지 여부에 따라 조합을 선택하십시오.
- 박테리아 사전 배양 성장
- 250mL 삼각 플라스크에 100 μg/mL 앰피실린과 100 μg/mL 스펙티노마이신이 함유된 훌륭한 액체(TB) 또는 LB 배지를 채웁니다.
- 멸균 접종 루프가 있는 LB-한천 플레이트에서 단일 콜로니를 선택하고 1.2.1단계에서 새 배지로 옮깁니다.
- 로터리 셰이커 인큐베이터에서 37°C에서 밤새 또는 최소 6시간 동안 배양합니다.
참고: 이 배양물로부터, 박테리아 현탁액을 글리세롤과 1:1로 혼합하고 -80°C에서 저장함으로써 글리세롤 스톡을 제조할 수 있다. 이 스톡은 1.2.2단계에서 사용할 수 있습니다. 갓 변형 된 식민지 대신.
- 세균 배양 및 단백질 발현 유도
- 성장한 박테리아 100mL를 50μg/mL 암피실린과 50μg/mL 스펙티노마이신이 함유된 5L의 TB 또는 LB에 옮깁니다.
- 이 배양액을 쉐이커 인큐베이터에서 37°C에서 비표지 셉틴의 경우 2-3 또는 msfGFP/mEGFP 표지 셉틴의 경우 0.6-0.8 범위의 파장 600nm에서 측정된 광학 밀도(OD)에 도달할 때까지 성장시키고 0.5mM IPTG의 최종 농도를 추가하여 단백질 발현을 유도합니다. 표지된 셉틴에 대한 더 낮은 OD는 다음 단계에서 상세히 설명되는 바와 같이, 그들의 더 긴 발현 시간에 사멸 단계에 도달하는 것을 피하기 위한 것이다.
- 표지되지 않은 셉틴 헤테로올리고머를 발현하는 세포를 37°C에서 3시간 동안 배양하거나 msfGFP-표지된 헤테로올리고머를 발현하는 세포를 17°C에서 밤새 배양합니다.
참고: 단백질 분해를 방지하기 위해 더 풍부한 TB 배지를 사용하여 촉진되는 표지되지 않은 복합체의 짧은 단백질 발현 시간이 선택됩니다. 표지된 복합체에 대해 더 낮은 온도와 결합된 더 긴 발현 시간은 msfGFP 태그의 올바른 폴딩을 허용하도록 선택됩니다.
- 박테리아 용해 및 용해물 정화
참고: 정제 절차의 이 시점부터 단백질 분해 단백질 분해 또는 활성 손실을 방지하기 위해 단백질 함유 용액을 항상 얼음 위에 또는 4°C에 보관하십시오.- 배양된 세포를 수집하여 4,000 x g 에서 4°C에서 20분 동안 원심분리한다. 상청액을 폐기하십시오.
- 선택적으로 이 단계에서 펠릿을 스냅 동결하고 -80°C에서 최대 6개월 동안 보관합니다. 이 옵션을 선택한 경우 계속하기 전에 펠릿을 얼음 위에서 해동하십시오.
- 펠릿을 용해 완충액 100mL(표 2)에 용해하고 세포를 용해시킵니다. 아래 두 가지 옵션 중 하나를 선택하십시오.
- 30% 진폭을 사용하는 팁 초음파 처리기로 30초 ON 및 59초 OFF의 7 주기로 초음파 처리합니다(설정은 초음파 처리기에 따라 다름).
- 프렌치 프레스의 셀을 최소 3x 통과시켜 분해합니다.
- 4°C에서 30분 동안 20,000 x g 에서 원심분리하여 세포 용해물을 명확히 하고 상청액을 유지한다. 1.5.1 단계부터 시작하는 것이 좋습니다. 이 원심 단계 동안.
- 선택적으로, 섹션 2에 설명된 대로 변성 전기영동을 위한 샘플을 채취한다.
- 배양된 세포를 수집하여 4,000 x g 에서 4°C에서 20분 동안 원심분리한다. 상청액을 폐기하십시오.
- His-tagged 단백질에 대한 친화성 크로마토그래피
참고: 이 단계는 니켈 컬럼을 사용하여 인간 SEPT2 또는 초파리 Sep1을 포함하는 복합체를 생성합니다(그림 1B).- 사전 충전된 니켈 세파로스 고성능 크로마토그래피 컬럼을 셉틴 완충액으로 평형화합니다(표 2).
- 정제된 상청액을 1mL/min의 속도로 컬럼에 로드하고 결합된 단백질을 최소 3개의 컬럼 부피의 셉틴 완충액으로 세척합니다.
- 셉틴 복합체를 50% HisTrap 용리 완충액(표 2)으로 1mL/분으로 용리하면서 0.5mL 분획을 수집하여 250mM의 이미다졸 농도를 수득했습니다.
- 빠른 단백질 액체 크로마토그래피(FPLC) 시스템으로 온라인으로 모니터링하거나 마이크로볼륨 분광광도계로 정제한 후 280nm에서 용리액의 광학 흡광도로 표시된 대로 셉틴 복합체를 포함하는 분획을 선택합니다.
참고: 이미다졸은 280nm에서 빛을 흡수합니다. 이것은 아마도 단백질 피크가 셉틴 용리 후 흡광도 0으로 돌아 가지 않는 이유를 설명합니다 (그림 2A).
- 스트렙-II 태그된 단백질에 대한 친화성 크로마토그래피
참고: 이 단계에서는 Strep-Tactin 컬럼을 사용하여 인간 SEPT7(헥사머), 인간 SEPT9(옥타머) 또는 초파리 땅콩을 포함하는 복합체를 생성합니다(그림 1B). 크로마토그래피 컬럼은 변형된 비오틴-스트렙타비딘 시스템을 기반으로 합니다. 단백질에는 변형된 비오틴(Strep-II-tag)이 태그되어 있으며 컬럼에는 조작된 스트렙타비딘(Strep-Tactin)이 포함되어 있습니다. 비오틴-스트렙타비딘 시스템에서 변형되었음에도 불구하고 스트렙-택틴-스트렙-II-태그 시스템과 비오틴-스트렙타비딘 시스템 사이에는 간섭이 없습니다. 설명 된 시스템은 비오틴 및 스트렙타 비딘을 사용하는 재구성 분석에 대한 간섭을 피하기 위해 사용됩니다.- 사전 충전된 스트렙탁틴 세파로즈 고성능 크로마토그래피 컬럼을 셉틴 완충액으로 평형화합니다(표 2). 니켈 컬럼에서 회수한 셉틴 함유 분획을 1mL/분으로 로딩하고 결합된 단백질을 최소 3개의 컬럼 부피의 셉틴 완충액으로 세척합니다.
- 셉틴 복합체를 100% StrepTrap 용리 완충액(표 2)으로 1mL/분으로 용리하면서 0.5mL 분획을 수집하여 2.5mM 데스티오비오틴 농도를 수득했습니다.
참고: StrepTrap 용출 완충액의 데스티오비오틴은 신선하게 용해되어야 합니다. - FPLC 시스템으로 온라인으로 모니터링하거나 마이크로볼륨 분광광도계로 정제한 후 280nm에서 용리액의 광학 흡광도에 의해 표시된 바와 같이 셉틴 복합체를 포함하는 분획을 선택합니다.
참고: 변성 전기영동은 일반적으로 이 시점에서 컬럼 세척 및 셉틴 분획 샘플을 사용하여 수행됩니다. 컬럼의 순서는 구별할 수 없는 결과, 즉 1.4단계 이후에 명확한 용해물로 반전될 수 있습니다. Strep-Tactin 친화성 크로마토그래피에 이어 니켈 친화성 크로마토그래피를 실시할 수 있습니다.
- 투석 및 보관
- 최종 저장 용액에서 데스티오비오틴을 제거하려면 셉틴 복합체를 밤새 1mM DTT가 보충된 셉틴 버퍼(표 2)에 대해 ~1:300 샘플 대 버퍼 부피비로 투석하거나 30kDa MWCO 투석막을 사용하여 4°C에서 최소 4시간 동안 투석합니다.
- 임의로, 30 kDa MWCO 원심 농도 컬럼을 사용하여 원하는 농도까지 셉틴을 농축한다. 용액 280nm에서 광학 흡광도를 통해 측정하고 ProtParam을 통해 계산된 이론적 흡광 계수를 사용하여 5-7μM의 농도를 목표로 합니다(표 3).
- 단백질 복합체를 원하는 분취량 크기로 분취하고, 분취량을 스냅 동결하고, -80°C에서 보관한다.
알림: 단백질을 6 개월 이상 보관하지 않는 것이 좋습니다. 또한 특히 단백질을 권장 시간보다 오래 보관하는 경우 정기적 인 품질 관리 실험을 수행하는 것이 좋습니다.
2. 셉틴 헤테로올리고머의 순도 및 무결성의 품질 관리
참고: 헤테로-올리고머 품질 관리는 용액에 존재하는 셉틴 복합체의 질량과 무결성을 검출할 수 있는 일련의 생화학적 및 이미징 기술로 구성됩니다.
- 올바른 성분으로 셉틴 헤테로올리고머의 형성을 확인하기 위한 변성 전기영동
- 선택된 분획 10μL를 2x SDS 샘플 버퍼 10μL와 혼합하고 프리캐스트 4%-15% TGX 겔에 로드하고 트리스/글리신/SDS 실행 버퍼로 시스템을 채웁니다.
- 200V에서 35분 동안 전기영동을 실행하고 겔(재료 표)을 염색하여 결과를 시각화합니다. 개별 셉틴 단백질 및 셉틴 헤테로-올리고머 복합체의 분자량은 표 3에서 확인할 수 있습니다.
- 대조 반전 이미지에서 정제된 셉틴을 포함하는 각 레인 내부의 각 밴드의 상대적 강도를 측정합니다. 이렇게 하려면 각 밴드 주위의 동일한 크기의 사각형과 동일한 레인에 밴드가 없는 영역에서 동일한 크기의 사각형의 평균 강도를 계산합니다. 그런 다음 각 밴드의 강도를 밴드가 없는 영역의 강도로 나누어 값을 정규화합니다.
참고: 강도가 포화된 경우(예: 대비 반전 이미지의 8비트 이미지에 대한 값 255) 차선을 건너뜁니다.
- 네이티브 전기영동을 통한 앙상블 평균 네이티브 크기 분포
- 전날 양극 완충액 800mL와 하늘색 음극 완충액 200mL를 준비하여 냉장고에 보관한다. 양극 완충액을 준비하려면 40mL의 20x 실행 완충액을 760mL의 I형 탈이온수(I-water)로 희석합니다. 하늘색 음극 완충액을 준비하려면 10mL의 20x 실행 완충액과 1mL의 20x 음극 첨가제를 189mL의 I-water로 희석합니다. 러닝 버퍼 및 음극 첨가제는 키트(재료 표)와 함께 제공됩니다.
- ~500ng의 셉틴을 필요한 양의 시료 완충액(이 경우 2.5μL, 4x 시료 완충액 사용으로 인해 4μL, 재료 표 참조) 및 10μL의 부피에 도달하기에 충분한 I-water를 혼합하여 시료 10μL를 준비합니다.
- 샘플을 젤에 로드하고 얼음처럼 차가운 양극 및 음극 버퍼로 시스템을 채웁니다.
- 낮은 전류에서 멈추지 않는 전원 공급 장치로 150V에서 약 115분 동안 전기영동을 실행하고 젤(재료 표)을 염색하여 결과를 시각화합니다. 서열에 기초하여 계산된 단일 단백질 및 복합체의 분자량은 표 3에서 확인할 수 있다.
- 간섭계 산란 현미경을 통한 질량 측광법을 사용한 단일 분자 질량 분포
- #1.5 유리 슬라이드를 초음파 세척기에서 I-water에서 5분, 이소프로판올에서 5분, 마지막으로 I-water에서 5분 동안 초음파 처리하여 세척합니다.
- 두 개의 유리 슬라이드를 부드러운 질소 가스 흐름으로 건조시키고 슬라이드 중 하나의 중앙에 0.01% Poly-L-Lysine(PLL) 용액 7μL 방울을 놓습니다. 그런 다음 다른 슬라이드의 가운데를 PLL 드롭 위에 놓고 쉽게 분리할 수 있도록 두 슬라이드의 방향을 직각으로 지정합니다. 30 초 동안 배양하십시오.
- I-water 1x로 비커에 담그고 I-water 2x의 스트림을 직접 적용하여 세척합니다. 그런 다음 질소 가스의 흐름으로 두 슬라이드를 건조시킵니다. 이 슬라이드는 건조한 상태의 실온에서 약 6주 동안 보관할 수 있습니다.
참고: 실험을 올바르게 실행하기 위해 PLL로 처리된 슬라이드의 측면에 레이블을 지정합니다.
- I-water 1x로 비커에 담그고 I-water 2x의 스트림을 직접 적용하여 세척합니다. 그런 다음 질소 가스의 흐름으로 두 슬라이드를 건조시킵니다. 이 슬라이드는 건조한 상태의 실온에서 약 6주 동안 보관할 수 있습니다.
- 실험 직전에 2 x 2, 3 x 2 또는 3 x 3 개스킷(각각 4, 6 또는 9개의 이미징 챔버/슬라이드 생성) 조각을 자르고 유리 슬라이드와 개스킷이 더러운 표면에 닿지 않도록 유리 슬라이드의 PLL 처리 부분에 붙입니다. 슬라이드를 경량 와이퍼 티슈에 놓고 피펫 팁으로 개스킷을 눌러 개스킷에 있는 보호 플라스틱에 붙입니다.
- 셉틴 완충액(표 2)을 실온으로 데우고 손에 든 단백질을 해동합니다(나중에 얼음에 보관).
참고: iSCAT은 단백질 신호45와 유사한 일부 세제 및 소분자의 신호를 보여줍니다. DTT는 작은 분자 중 하나이므로 이 실험에 사용되지 않습니다. 저장된 셉틴에서 나오는 DTT의 흔적만 있습니다. - 19μL의 셉틴 버퍼가 포함된 상업용 질량 측광 시스템에 개스킷이 있는 슬라이드를 놓고 자동 초점 옵션을 사용하여 현미경의 초점을 맞춥니다. 제조업체의 지침에 따라 찾은 초점이 올바른지 확인하십시오. 설정의 일부인 표준 100x 대물렌즈가 여기에 사용됩니다.
- 새 프로젝트의 파일 또는 프로젝트 로드>> 사용하여 데이터를 저장할 프로젝트 폴더를 만들거나 로드합니다.
- 19μL의 셉틴 버퍼 드롭(단계 2.3.5)에 1μL의 샘플을 피펫팅하여 초점을 맞추고 혼합하는 동안 아무 것도 건드리지 않음으로써 슬라이드의 움직임을 최소화합니다. 그런 다음 녹화를 클릭하여 6,000프레임 비디오를 녹화합니다.
- 올바른 분석을 위해 셉틴 버퍼, 신호 대 질량 비율 교정을 위한 단백질 질량 표준(최근 보정이 가능하고 환경 조건이 변경되지 않은 경우 이 샘플을 건너뛸 수 있음) 및 DTT 없이 셉틴 버퍼에 희석된 셉틴 복합체 250nM(최종 농도는 ~12.5nM임)을 기록합니다.
- 제조업체의 소프트웨어를 사용하여 비디오를 분석하여 단백질 질량 분포를 얻습니다. 다음과 같이 양질의 데이터를 확인하십시오.
- 서로 다른 셉틴 헤테로-올리고머 크기의 피크가 너무 많이 겹치거나 너무 많은 이벤트가 감지되는 경우(10.8μm x 2.9μm에 걸쳐 128픽셀 x 34픽셀의 일반 시야각을 가진 6,000프레임 비디오의 경우 >3,500개 이벤트), 최종 셉틴 농도를 줄이고 다시 측정합니다.
- 측정된 단일 분자 수가 충분하지 않은 경우(일반 시야가 있는 6,000프레임 비디오의 경우 최소 2,500-3,500개) 셉틴 농도를 높이고 다시 측정합니다.
- 음성 염색 투과 전자 현미경을 통한 셉틴 복합체의 직접 이미징
- 샘플을 셉틴 완충액 중 약 50 nM의 농도로 희석하고, 염색 용액 (2% 우라닐 포르메이트 또는 우라닐 아세테이트I-water)을 준비한다.
알림: 우라닐 포르메이트는 신선하게 준비해야 합니다. - 희석된 셉틴 4μL를 글로우 방전된 전자 현미경 그리드에 피펫팅하고 30초 동안 배양합니다.
- 여과지를 사용하여 대부분의 단백질 용액을 제거하고 그리드를 셉틴 버퍼로 2x, I-물로 1x 세척하여 느슨하게 흡착된 셉틴을 제거합니다.
- I-water에서 2% 우라닐 아세테이트 또는 우라닐 포르메이트 용액으로 1분 동안 염색하고, 염색 용액을 여과지로 흡수하고, 그리드를 몇 분 동안 자연 건조시킨다.
- 적절하게 정렬된 투과 전자 현미경을 사용하여 그리드를 스크리닝하여 강화된 염색 영역을 검색하고 이러한 선택된 영역 내에서 약 100개의 이미지를 수집합니다.
- 최소 50,000x 배율로 이미지를 수집하여 약 2Å/픽셀의 픽셀 크기와 -1μm에서 -2μm까지 다양한 디포커스를 얻습니다. 200kV의 가속 전압을 사용하십시오. 바람직하게는, 데이터를 수집하기 위해 자동화된 절차를 사용하며, 이는 이용 가능한 획득 소프트웨어에 의존할 것이다.
- 전용 소프트웨어를 사용하여 2D 이미지 처리 수행
- 전용 소프트웨어(46)를 사용하여 적어도 2,000개의 입자를 박스아웃한다.
- 추가 개선 없이 클래스를 얻을 때까지 2차원 정렬 및 분류를 반복적으로 수행합니다. 첫 번째 정렬 및 분류 단계는 분류의 편향을 피하기 위해 참조가 없어야 합니다.
- 참조가 없는 첫 번째 분류에서 얻은 평균을 새 참조로 사용하여 추가 분류 라운드를 수행합니다. 더 이상 개선되지 않을 때까지 이 프로세스를 반복적으로 반복합니다. 각 클래스가 50-100개의 선택된 파티클을 기반으로 하고 개별 서브유닛이 명확하게 보이는지 확인합니다. 다른 소프트웨어 도구를 사용할 수 있습니다 (스파이더, 이만 또는 Relion)46,47,48.
- 샘플을 셉틴 완충액 중 약 50 nM의 농도로 희석하고, 염색 용액 (2% 우라닐 포르메이트 또는 우라닐 아세테이트I-water)을 준비한다.
3. 중합 분석을 통한 Septin 기능성 품질 관리
참고: 기능 품질 관리는 중합된 셉틴 복합체의 검출을 허용하는 일련의 이미징 기술로 구성됩니다. 아래에서 표지되지 않은 셉틴을 "다크" 셉틴이라고 하며, 표지되지 않은 셉틴을 중합하는 데 사용되는 완충액을 "어두운" 셉틴 중합 버퍼(SPB)라고 합니다.
- 형광 현미경을 통한 셉틴 번들 이미징
- 셉틴 완충액 + 1mM DTT 중 원하는 최종 농도보다 6배 높은 농도로 5x fluoSPB(표 2) 및 90% 다크 셉틴 및 10% msfGFP-셉틴으로 구성된 셉틴 혼합물을 준비합니다. 이 분석에 대한 전형적인 농도는 300 nM이고, 따라서, 농도는 이 혼합물에 대해 1,800 nM이다.
- 이 특정 순서로 I- 물 (최종 원하는 부피까지 채우기에 충분), 20 % 5xfluoSPB (1 : 5의 최종 희석), 0.05 μM PCD 및 16.67 % 셉틴 믹스 (1 : 6의 최종 희석)를 혼합하여 셉틴을 중합합니다. 10 μL의 경우, 6.23 μL의 I- 워터, 2 μL의 5xfluoSPB, 0.1 μL의 PCD (5 μM의 재고 포함) 및 1.67 μL의 셉틴 믹스를 혼합한다. 이 혼합물을 실온에서 최소 30분 동안 배양합니다.
- fluoSPB로 세척된 이미징 챔버에 샘플을 추가하고(표 2) 셉틴 다발을 이미지화합니다. 이전 연구10,32에서 설명한 PLL-PEG-부동태화 유동 채널은 이 실험에 적합합니다.
- 음성 염색 투과 전자 현미경을 통한 셉틴 다발 상상
- 셉틴 완충액 + 1mM DTT에서 원하는 최종 농도보다 6배 높은 농도로 5x darkSPB(표 2) 및 100% 다크 셉틴으로 구성된 셉틴 혼합물을 준비합니다. 이 분석에 대한 전형적인 농도는 300 nM이고, 따라서, 농도는 이 혼합물에 대해 1,800 nM이다.
- 이 특정 순서로 I- 물 (최종 원하는 부피까지 채우기에 충분함), 20 % 5xdarkSPB 및 16.67 % 셉틴 혼합을 혼합하여 셉틴 복합체를 중합합니다. 5μL의 경우 I-워터 3.16μL, 5x darkSPB 1μL, 셉틴 혼합물 0.83μL를 혼합합니다. 이 혼합물을 실온에서 최소 30분 동안 배양합니다.
- 3-5 μL의 샘플을 글로우 방전 전자 현미경 그리드에 추가하고 1분 동안 배양합니다. 그런 다음 여과지로 액체를 흡수하고 darkSPB 버퍼 한 방울을 추가하여 darkSPB(표 2)로 그리드를 2배 세척하고, I-water로 1x 세척하고, 2% 우라닐 아세테이트로 ~30초 동안 배양하고, 얼룩을 제거하고, 샘플을 몇 분 동안 자연 건조시킵니다.
- 120kV에서 셉틴 번들을 이미지화하고 1-2μm의 디포커스로 5,000x에서 60,000x 사이의 배율을 이미지화합니다.
결과
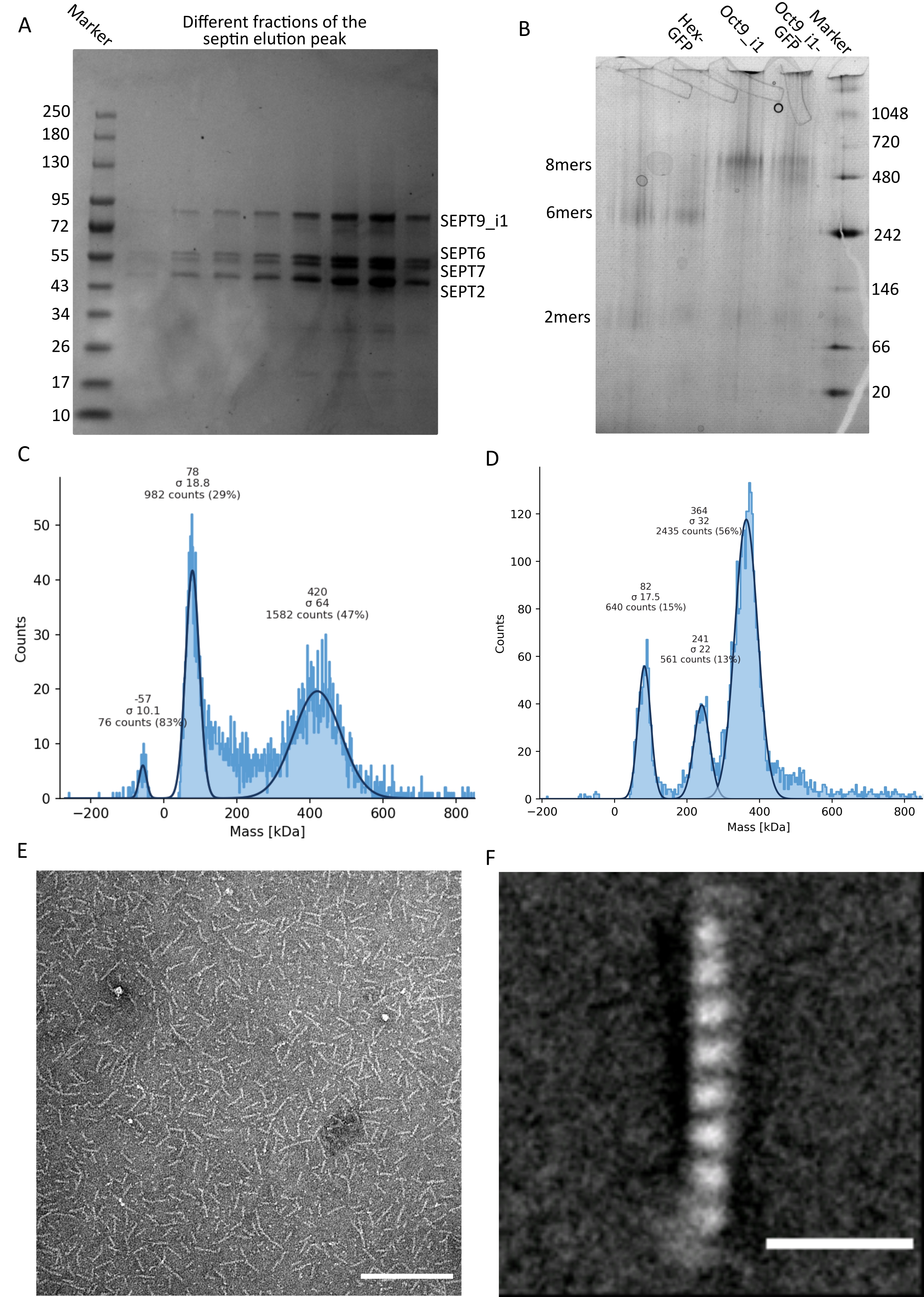
프로토콜에서 언급된 바와 같이, 2개의 셉틴 발현 플라스미드와 공동 형질전환된 5L의 대장균 세포를 성장시키고, IPTG를 첨가하여 셉틴의 발현을 유도하였다. 3 시간 후, 세포를 원심분리에 의해 수집하고, 용해 완충액에 재현탁시키고, 초음파 처리에 의해 용해시켰다. 이어서, 용해물을 원심분리에 의해 정화하고, 정제된 용액을 HisTrap 컬럼에 적용하였다(도 2A). 첫 번째 정제 후, 셉틴 함유 분획을 풀링하여 StrepTrap 컬럼에 적용했습니다(그림 2B). 이것은 일반적으로 약 3-5mL의 ~1μM 셉틴 복합체를 생성합니다. 셉틴 함유 분획을 풀링하기 전에, 변성 겔 전기영동을 사용하여 셉틴 서브유닛의 완전성 및 복합체를 형성하는 상이한 셉틴 서브유닛 사이의 등몰 화학량론적 비율을 확인할 수 있다. (그림 3A). 겔이 셉틴 서브유닛의 분자량(표 3)에 상응하는 유사하게 강렬한 밴드를 나타내면, 프로토콜은 계속될 수 있다. 그렇지 않은 경우 프로토콜을 다시 시작하는 것이 좋습니다. SEPT9_i1 가진 인간 셉틴 옥타머에 대해 도시된 예에서, 그림 3A 는 유사한 강도를 갖는 SEPT9_i1, SEPT6, SEPT7 및 SEPT2(위에서 아래 순서로)에 해당하는 대역을 명확하게 보여줍니다. 정규화된 강도의 99% 신뢰 구간은 SEPT2의 경우 1.128 ± 0.048, SEPT6의 경우 1.092 ± 0.034, SEPT7의 경우 1.108 ± 0.040, SEPT9의 경우 1.067 ± 0.029였습니다. SEPT2에 msfGFP 태그가 지정된 경우 SEPT9_i1 아래에서 매우 가깝게 이동합니다. 사용 된 전기 영동 시스템과 SEPT7에 대한 C- 말단 TEV-Strep 태그의 존재 (태그가없는 SEPT7보다 더 느리게 이동)에 따라 SEPT7 및 SEPT6 밴드는 비슷한 분자량으로 인해 때때로 병합됩니다. 다음 단계는 분획을 풀링하고 DTT로 셉틴 완충액에 대해 투석하는 것입니다. 투석 후, 농도가 너무 낮거나(<2μM) 더 높은 농도가 실험에 필요한 경우, 프로토콜에 기재된 바와 같이 농축 단계가 포함될 수 있다. 1μM 미만의 농도는 일반적으로 셉틴의 나쁜 기능적 품질을 나타냅니다. 3.5 μM에서 7 μM 사이의 최종 셉틴 복합체 농도는 대부분의 시험관 내 분석에 적합합니다. 이러한 농도는 일반적으로 농도 후 부피가 0.5-1 mL에 도달 할 때 얻어집니다.

그림 2: 어두운 인간 셉틴 octamers_9i1의 정제에 해당하는 크로마토그램의 예. (A) HisTrap 컬럼 크로마토그램. 셉틴 용리 피크 후, 흡광도는 0으로 돌아 가지 않는데, 이는 완충액에 이미 다졸이 존재하기 때문일 수 있습니다. 풀링된 분획은 용리 피크의 시작부터 흡광도가 약 250mL에서 안정화될 때까지 진행되었습니다. (B) 스트렙트랩 컬럼 크로마토그램. 풀링된 분획은 용리 피크의 시작부터 흡광도가 50mL에서 약 0으로 돌아올 때까지 진행되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
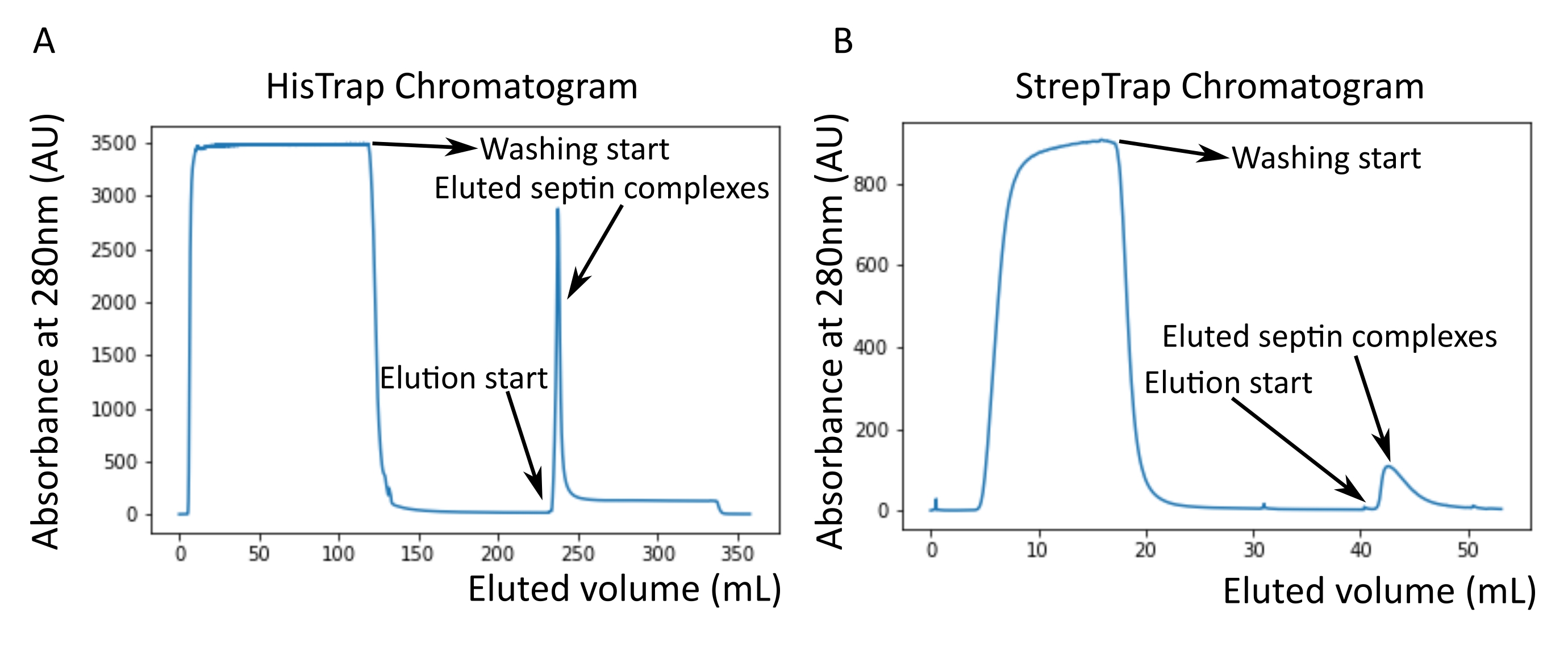{kind=link}
품질 관리를 계속하기 위해, 프로토콜에 기재된 바와 같이 천연 전기영동을 수행하였다(도 3B). 겔에서, 무손상 헤테로올리고머에 상응하는 메이저 밴드 및, 보통, 이량체에 대응하는 마이너 밴드가 관찰될 수 있다. 인간 헥사머는 242kDa 마커 밴드보다 약간 위에 있는 반면, 옥타머는 계산된 분자량보다 높은 480kDa 밴드 위에서 발견됩니다. 이들 밴드의 위치는 진핵세포 추출물32의 웨스턴 블롯 분석에 의해 확인되었다. msfGFP로 태그를 지정하면 각 SEPT2가 msfGFP 단백질과 결합됩니다. 이로 인해 셉틴 복합체의 분자량이 53.4kDa(26.7kDa/msGFP 분자)로 증가합니다. 그럼에도 불구하고, 천연 전기영동 겔 상에서, msfGFP-태그된 복합체의 겉보기 분자량은 태그되지 않은 복합체의 겉보기 분자량과 구별할 수 없다.
셉틴 복합체가 손상되지 않았는지 여부를 테스트하는 보완 기술은 iSCAT 현미경에 의한 질량 측광법입니다. iSCAT은 기준광과의 간섭에 의해 증폭된 유리 슬라이드에 착륙하는 분자의 광산란(일반적으로 유리 슬라이드 바닥에 레이저가 반사됨)을 모니터링합니다. 그런 다음 배경 빼기 방식을 사용하여 입자에 대비를 제공합니다. 이러한 보정으로 인해, 신호는 입자가 유리 상에 떨어지는지 또는 유리로부터 멀어지는지에 따라 양수 및 음수 값을 나타낸다(49). 검출 된 신호는 단백질50의 분자량에 정비례합니다. 따라서 질량 표준물질을 사용한 신호 대 질량 교정은 샘플 단백질의 질량을 결정할 수 있습니다. 도 3C 는 SEPT9_i1 함유하는 인간 셉틴 옥타머의 예를 나타낸다. 검출된 단일 입자의 대부분(~50%)은 SEPT9_i1(423kDa)를 포함하는 완전한 옥타머에 대해 예상되는 분자량입니다(그림 3C). 150-300 kDa 사이의 질량을 가진 입자도 있지만 명확한 피크는 관찰되지 않아 다른 셉틴 종의 존재 가능성을 나타냅니다. 유사하게, mEGFP 태그가 부착된 초파리 헥사머에 대해 검출된 대부분의 단일 입자는 원형 헥사머(361kDa)에 대해 예상되는 분자량입니다(그림 3D). 241 kDa에서의 추가적인 명확한 피크는 2 개의 땅콩 단백질, 하나의 DSep1 및 1 개의 mEGFP-DSep2를 함유하는 안정한 사량 체의 존재를 나타낸다. 마지막으로, 인간 및 파리 셉틴 복합체는 모두 단량체와 이량 체의 혼합 일 수있는 약 80 kDa의 피크를 나타내며, 아마도 DTT 또는 응집하는 다른 소분자의 흔적에 의해 증폭되어 플롯(45)의 양성 측면에서 피크를 나타낸다.

그림 3: 올리고머 품질 관리 결과 의 예. (a) 어두운 인간 셉틴 octamers_9i1의 정제로부터의 용리 피크의 상이한 분획을 나타내는 변성 겔의 예. (B) 상이한 셉틴 복합체의 천연 전기영동의 예. (씨,디) 셉틴 복합체의 12.5 nM에서의 질량 측광의 히스토그램 결과의 상이한 예: (C) 어두운 인간 셉틴 octamers_9i1 및 (D) DSep1-msfGFP 초파리 셉틴 헥사머. 선은 가우스 피팅입니다. (e) 셉틴 완충액 중 25 nM 어두운 인간 셉틴 octamers_9i1의 TEM 이미지. 스케일 바 = 200nm. (F) SEPT2-msfGFP 인간 셉틴 octamers_9i1의 클래스 평균 이미지. msfGFP 태그는 양쪽 끝에 퍼지 밀도로 표시됩니다. 스케일 바 = 10nm. 패널 (E)와 (F)는 The Company of Bioologists의 저작권이며 Iv et al.10 에서 허가를 받아 각색되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
네이티브 겔과 iSCAT 모두 앙상블 평균 정보만 제공한다는 점을 감안할 때, 단일 셉틴 올리고머의 투과 전자 현미경 이미지의 클래스 평균화를 사용하여 직접 시각화를 통해 복합체의 무결성과 순도를 확인했습니다. 셉틴 완충액에서 셉틴 복합체의 TEM 이미지에서 길이가 24nm(헥사머) 또는 32nm(옥타머)인 막대가 관찰될 수 있습니다. SEPT9_i1 함유하는 인간 셉틴 옥타머의 예는 도 3E에서 알 수 있다. 이들을 평균화할 때, 도 3F에서 SEPT9_i1 갖는 msfGFP-태그된 인간 옥타머에 대해 볼 수 있는 바와 같이, 각각의 서브유닛을 관찰하고 계수할 수 있다. 올리고머가 형광 표지된 경우 막대 끝에서 SEPT2-msfGFP에 해당하는 추가 밀도를 관찰할 수 있습니다(그림 3F).
상기 기술의 조합은 정확한 화학량론적 비율 및 고순도를 갖는 옥타머(또는 헥사머)가 설명된 프로토콜을 사용하여 정제될 수 있음을 증명한다. 마지막으로, 마지막 품질 관리 검사는 중합 능력 측면에서 셉틴 복합체의 기능에 대한 것입니다. 낮은 염 농도 (<기재된 완충액9를 갖는 150 mM KCl)의 존재 하에서, 셉틴이 다른 단백질 또는 음으로 하전 된 지질 막의 존재 하에, 이들은 번들(9)로 자체 조립된다. 셉틴은 높은 (300 mM) KCl 농도를 갖는 저장 완충액에 보관함으로써 중합을 방지합니다. 그런 다음 셉틴 헤테로-올리고머를 KCl이 없는 동일한 조성의 완충액에서 1:6 부피비로 희석하여 50mM의 최종 KCl 농도를 달성합니다. 형광 이미징을 수행하기 위해 이 완충액은 광퇴색으로부터 보호하기 위한 산소 제거 시스템과 깜박이는 억제기로 보완됩니다. TIRF 현미경 검사에서 얕은 TIRF 필드 (~ 100 nm; 그림 4A, B). 컨포칼 현미경에서 필라멘트 구조의 큰 클러스터가 용액 위로 떠 있는 것을 볼 수 있습니다(그림 4C). 마지막으로, TEM을 사용하면 TIRF에 의해 관찰 된 클러스터에 해당하는 작은 셉틴 다발 (그림 4D)과 공초점 현미경으로 관찰 된 구조에 해당하는 큰 다발 (그림 4E)이 관찰 될 수있다. 그림 4D, E 의 삽입물은 두 가지 유형의 구조가 평행하게 실행되는 길고 얇은 필라멘트로 구성되어 끝이 가늘어지는 번들을 형성한다는 것을 보여줍니다. 형광 및 TEM 이미지는 함께 정제된 셉틴 복합체가 필라멘트로 중합될 수 있음을 증명하며, 필라멘트는 차례로 번들로 자체 조립됩니다.

도 4: 중합능 품질 관리의 결과의 예. (A) fluoSPB 중 300nM 인간 셉틴 헥사머(10% msfGFP 표지 헥사머)의 TIRF 이미지. (B) fluoSPB 중 SEPT9_i1 (10 % msfGFP 표지 octamers9_i1)를 함유하는 300 nM 인간 셉틴 옥타머의 TIRF 이미지. (C) fluoSPB에서 300nM 인간 셉틴 octamers_9i3의 0.5μm 간격으로 ~30μm에 걸친 Z-스택의 공초점 최대 강도 투영. (A-C) 스케일 바 = 10 μm 및 반전 된 그레이 스케일. (D,E) (D) darkSPB에서 인간 셉틴 octamers_9i1의 작은 및 (E) 큰 다발의 TEM 이미지 예. 인셋은 번들 내에서 평행하게 실행되는 투명한 필라멘트가 관찰될 수 있는 영역을 보여줍니다. 스케일 바 = 500nm. 패널(C-E)은 The Company of Bioologists의 저작권이며 Iv et al.10에서 허가를 받아 각색되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
표 1: 플라스미드 목록. 이 프로토콜에 따라 셉틴 올리고머를 정제하는 플라스미드. 모든 플라스미드는 Addgene (첫 번째 컬럼)에 기탁되었습니다. 이 표를 다운로드하려면 여기를 클릭하십시오.
표 2: 버퍼 목록. 셉틴 올리고머의 정제 및 품질 관리에 사용되는 완충액 조성물. 이 표를 다운로드하려면 여기를 클릭하십시오.
표 3: 분자량 및 흡광 계수. 복합체의 서열을 기반으로 ProtParam으로 계산된 파장 280nm에서의 분자량(MW) 및 광학 흡광 계수(ε) 목록, 셉틴 서브유닛, 상이한 셉틴 복합체 및 표 1에 나열된 플라스미드로 정제할 수 있는 고유한 셉틴 서브유닛(MW만)의 선형 융합을 가정합니다. 이 표를 다운로드하려면 여기를 클릭하십시오.
토론
여기에 설명된 방법은 사전 형성된 셉틴 헤테로올리고머의 강력한 정제 및 품질 관리를 가능하게 합니다. 이 방법의 올바른 적용을 위해 고려해야 할 몇 가지 주요 문제는 다음과 같습니다. 크로마토그래피 분리의 용리 단계 중에 셉틴 복합체의 희석을 최소화하기 위해 권장(또는 더 낮은) 유속을 사용하는 것이 중요합니다. 또한 최종 농축 단계에서 단백질 회수를 최대화하기 위해 농축기 컬럼은 용액이 필터에 밀려나지 않는 방식으로 배향됩니다(한쪽에만 필터가 있는 경우). 용액이 필터로 직접 이동하면 단백질이 필터에 훨씬 더 달라 붙어 최종 수율이 크게 감소합니다. 농축 단계가 항상 필요한 것은 아니라는 점을 고려하는 것도 중요합니다. 크로마토그램의 피크 주변의 좁은 범위에서만 분획을 선택하는 것은 일반적으로 많은 재구성 응용 분야(일반적으로 10-300nM 사이에서 작동)에 대해 충분히 높은 스톡 농도(>3,000nM)를 제공합니다. 마지막으로, 형광 현미경에 의한 셉틴 복합체의 기능성을 품질 제어하기 위해서는 셉틴 복합체가 유리에 열렬히 달라 붙기 때문에 현미경 슬라이드의 표면을 올바르게 부동 태화하는 것이 중요합니다. 유리 슬라이드의 패시베이션은 PLL-PEG 기능화 또는 중성(100% DOPC) 지지된 지질 이중층(11,32)의 형성에 의해 수행될 수 있다.
Iv et al.10에 처음 기술된 원래의 정제 프로토콜과 비교하여, 완충액 조성에 변화가 있다(표 2). MgCl2의 농도는 5mM에서 2mM로 감소하였고, Tris-HCl의 농도 및 pH는 각각 50mM에서 20mM 및 8.0에서 7.4로 감소하였다. 이러한 변경은 완충 조건을 인간 셉틴과 지질 이중층, 액틴 필라멘트 및 미세 소관10,11,32의 상호 작용에 대한 연구와 양립 할 수 있도록하기 위해 이루어졌습니다. 이는 저자들이 F- 완충액에 ATP가 존재하는 것을 제외하고는 darkSPB의 조성과 동일한 F- 완충액에서 지지된 지질 이중층과 중합 된 액틴을 형성했기 때문입니다. 완충액 변화는 원래 완충액과 비교하여 정제된 셉틴의 품질 또는 수명에 어떠한 변화도 일으키지 않았다.
이 정제 방법에는 여전히 몇 가지 한계가 있습니다. 첫째, 상이한 정제 시도는 정제된 셉틴 복합체의 번들 형성 능력에 의해 확인된 바와 같이 수율(2-5μM 셉틴 복합체의 0.5-1mL) 및 기능적 품질에서 다양할 수 있다. 그렇기 때문에 이 백서에 설명된 품질 검사를 일관되게 수행하는 것이 매우 중요합니다. 박테리아 배양의 발현 시간과 광학 밀도를 매우 잘 제어하면 수율 차이를 완화하는 데 도움이 될 수 있습니다. 둘째, 이 정제 파이프라인은 삼량체와 헥사머 또는 사량체와 옥타머를 구별할 수 없습니다(그림 1B). 그러나, 품질 관리 실험은 셉틴 복합체의 대다수가 그들의 긴 올리고머 형태임을 증명하기 위해 사용될 수 있다. 더 좁은 올리고머 크기 분포가 필요한 경우 1.6단계 사이에 크기 배제 크로마토그래피를 삽입할 수 있습니다. 및 단계 1.7. 정제 프로토콜의. 그러나 이 선택적 단계는 수율을 크게 감소시키며 꼭 필요한 경우가 아니면 권장되지 않습니다. 마지막으로, 보다 근본적인 한계는 재조합 셉틴 복합체의 발현 시스템으로 대장균을 사용하는 것에서 비롯됩니다. 당연히이 시스템은 인산화, 아세틸 화 및 수모 일화 6,51,52,53과 같은 동물 세포에서보고 된 번역 후 변형 (PTM)을 허용하지 않습니다. 이러한 번역 후 변형은 곤충 또는 인간 세포에서 유사한 정제 전략을 구현함으로써 추가 될 수 있습니다. 또한, 이 논문은 셉틴의 재구성 자체에 대해서만 논의했지만, 세포에 대한 연구에 따르면 Borg 계열 54,55 및 아닐린24,25,56의 단백질과 같은 조절 단백질은 셉틴의 조립 및 기능에 실질적이지만 잘 이해되지 않은 영향을 미칠 수 있으므로 결국 체외에서 통합하는 것이 중요합니다. 연구. Borg 단백질 및 아닐린의 정제를 위한 프로토콜은54,57로 보고되었다.
여기에 보고된 셉틴 정제 프로토콜은 올바른 서브유닛 화학량론을 사용하여 올리고머 형태의 셉틴을 정제하는 표준화된 방법을 제공하여 단일 셉틴 서브유닛에 의존하는 많은 초기 시험관 내 연구에 비해 중요한 발전을 제공합니다. 특정 상황에서 일부 셉틴이 단일서브유닛2으로 작용할 수 있지만, 현재의 문헌은 동물 세포에서 셉틴이 대부분 복합체 9,58에서 기능한다는 것을 강력하게 시사합니다. 따라서, 이 논문 및 기타 10,11,18,32,35,36,37에 기술된 것과 같은 미리 형성된 헤테로-올리고머의 사용은 in vitro를 통해 셉틴의 구조적 및 생물물리학적 특성을 연구하는 데 매우 중요합니다. 세포에서 기능을 해부하기위한 재구성. 또한, 셉틴은 막과 세포 골격을 포함한 많은 상호 작용 파트너를 가진 자기 조립 단백질이므로 상향식 합성 생물학59,60,61 및 곡률42,62,63과 같은 막 생물 물리학 적 특성의 단백질 유도 변화에 대한 연구에 큰 관심을 끌고 있습니다.
공개
저자는 경쟁 또는 재정적 이익을 선언하지 않습니다.
감사의 말
질량 측광(iSCAT) 실험에 도움을 준 Cecilia de Agrela Pinto, Tomás de Garay, Katharina Häußermann에게 감사드립니다. 아르옌 야코비와 윌 에버스가 TEM에 도움을 주었습니다. TIRF에 대한 그녀의 도움을 주신 루시아 발다우프; 파스칼 베르디에 - 피나르 (Pascal Verdier-Pinard)는 네이티브 전기 영동에 관한 조언을 해주었습니다. Agata Szuba와 Marjolein Vinkenoog는 Drosophila septin 정제 노력과 프랑스 국립 연구 인프라 France-BioImaging (ANR10-INBS-04)의 회원인 Institut Curie의 Cell and Tissue Imaging (PICT-IBiSA)을 설정하는 데 도움을 주었습니다. 이 연구는 'BaSyC-합성 세포 구축' 중력 보조금(024.003.019)을 통해 네덜란드 과학 연구 기구(NWO/OCW)와 Agence Nationale pour la Recherche(ANR 보조금 ANR-17-CE13-0014: "SEPTIMORF"; ANR-13-JSV8-0002-01: "9월"; 및 ANR-20-CE11-0014-01: "SEPTSCORT").
자료
| Name | Company | Catalog Number | Comments |
| 488nm laser combiner iLAS2 | Gataca | TIRF microscope | |
| 488nm Sapphire laser lines | Coherent | Confocal microscope | |
| 4k X 4k F416 CMOS camera | TVIPS | For JEM-1400plus | |
| 4x sample buffer nativePAGE | Thermo Fisher scientific | BN2003 | |
| 6-Hydroxy-2,5,7,8-tetramethylchromane-2-carboxylic acid (TROLOX) | Sigma-Aldrich | 238813 | To prevent blinking |
| AKTA pure 25 M1 | GE healthcare | 1680311 | |
| Ampicillin | Sigma-Aldrich | A9518-25G | |
| Carbon Type-B, 300 mesh EM grid | Ted pella | 01813-F | |
| Carbon Type-B, 300 mesh EM grid | Electron micoscopy sciences | CF300-Cu | |
| Cover glass #1.5H | Thorslabs | CG15KH | |
| CSU-X1-M1 confocal unit | Yokogawa | Confocal microscope | |
| Desthiobiotin | Sigma-Aldrich | D1411-1G | |
| Dithiothreitol (DTT) | Sigma-Aldrich | D9779 | |
| DNAse | Sigma-Aldrich | 10104159001 | |
| DOPC | Avanti Polar Lipids | 850375C | |
| Eclipse Ti2-E | Nikon instruments | Confocal microscope | |
| EDTA-free protease inhibtor cocktail | Roche | 481761 | |
| HisTrap HP, 5 mL | GE healthcare | 29-0588-3 | |
| iLAS2 azimuthal TIRF illumination system | Gataca | TIRF microscope | |
| Imidazole | Sigma-Aldrich | 1202-1KG | |
| InstantBlue Protein Gel Stain | Westburg Life Sciences | EP ab119211 | |
| Isopropyl β-D-1-thiogalactopyranoside (IPTG) | Thermo Fisher scientific | 10849040 | |
| iXon Ultra 888 EMCCD camera | Andor | Confocal microscope | |
| iXon Ultra 897 EM-CCD | Andor | TIRF microscope | |
| JEM-1400plus | JOEL | TEM microscope TUDelft | |
| kappa-cassein | Sigma-Aldrich | C0406 | |
| LB broth | Sigma-Aldrich | L3022-6X1KG | |
| Lyzozyme | Sigma-Aldrich | 62971-10G-F | |
| Magnesium Chloride | Sigma-Aldrich | M8266-100G | |
| Magnesium sulfate | Sigma-Aldrich | 746452-1KG | |
| Methylecllulose | Sigma-Aldrich | 8074844 | |
| MilliQ system (Integral 10) | Merck-Millipore | I-water dispenser | |
| Mini protean TGX gels | BIORAD | 4561086 | |
| NativeMark unstained protein standard | Invitrogen | LC0725 | For iSCAT and Native gels |
| NativePAGE 4-16% GELS | Thermo Fisher scientific | BN1002BOX | |
| NativePAGE Running Buffer kit | Thermo Fisher scientific | BN2007 | |
| Nikon Ti2-E | Nikon instruments | TIRF microscope | |
| Nr. 1 Menzel coverslips | Thermo Fisher scientific | 11961988 | |
| parafilm | Sigma-Aldrich | P7668 | |
| Plan Apo ×100/1.45 NA oil immersion objective | Nikon instruments | Confocal microscope | |
| PMSF | Sigma-Aldrich | 10837091001 | |
| Poly(L-lysine)-graft-biotinylated PEG (PLL-PEG) | SuSoS | CHF560.00 | |
| Poly-L-lysine solution 0.01% | Sigma-Aldrich | P4832 | For iSCAT glass slides |
| Pottassium Chloride | Sigma-Aldrich | P9541-1KG | |
| Power supply for native gels | CONSORT | S/N 71638 | |
| POWERPAC UNIVERSAL | BIORAD | 042BR31206 | |
| Protocatechuate 3,4-Dioxygenase (PCD) | Sigma-Aldrich | P8279-25UN | oxygen scavenger - enzyme |
| Protocatechuic acid (PCA) | Sigma-Aldrich | 03930590-50MG | oxygen scavenger - reagent |
| Q500 Sonicator | Qsonica | Q500-110 | |
| Quemesa camera | Olympus | For Tecnai Spirit | |
| Refeyn OneMP | Refeyn | ||
| Sample buffer, laemmli 2x concentrate | Sigma-Aldrich | S3401-10vl | |
| Silicon gaskets | Sigma-Aldrich | GBL103250-10EA | |
| Slide-A-Lyzer Dialysis cassettes 30k MWCO 3mL | Thermo Fisher scientific | 66381 | |
| Spectinomycin | Sigma-Aldrich | PHR1441-1G | |
| StrepTrap HP, 1 mL | GE healthcare | 28-9075-46 | |
| Tecnai Spirit microscope | Thermo Scientific, FEI | TEM microscope Institute Curie | |
| Terrific broth | Sigma-Aldrich | T0918-1KG | |
| Tris/Glyine/SDS buffer | BIORAD | 1610772 | |
| Tris-HCl | Sigma-Aldrich | T5941-1KG | |
| Ultrasonic cleaner | Branson | CPX2800H-E | |
| Vivaspin 6, 30,000 MWCO PES | Sartorius | VS0622 |
참고문헌
- Mostowy, S., Cossart, P. Septins: The fourth component of the cytoskeleton. Nature Reviews Molecular Cell Biology. 13 (3), 183-194 (2012).
- Shuman, B., Momany, M. Septins from protists to people. Frontiers in Cell and Developmental Biology. 9, 3802(2022).
- Bridges, A. A., Gladfelter, A. S. Septin form and function at the cell cortex. Journal of Biological Chemistry. 290 (28), 17173-17180 (2015).
- Smith, C., et al. Septin 9 exhibits polymorphic binding to F-actin and inhibits myosin and cofilin activity. Journal of Molecular Biology. 427 (20), 3273-3284 (2015).
- Gilden, J. K., Peck, S., Chen, Y. C. M., Krummel, M. F. The septin cytoskeleton facilitates membrane retraction during motility and blebbing. Journal of Cell Biology. 196 (1), 103-114 (2012).
- Marquardt, J., Chen, X., Bi, E. Architecture, remodeling, and functions of the septin cytoskeleton. Cytoskeleton. 76 (1), 7-14 (2018).
- Van Ngo, H., Mostowy, S. Role of septins in microbial infection. Journal of Cell Science. 132 (9), (2019).
- Fung, K. Y. Y., Dai, L., Trimble, W. S. Cell and molecular biology of septins. International Review of Cell and Molecular Biology. 310, 289-339 (2014).
- Kinoshita, M., Field, C. M., Coughlin, M. L., Straight, A. F., Mitchison, T. J. Self- and actin-templated assembly of mammalian septins. Developmental Cell. 3 (6), 791-802 (2002).
- Iv, F., et al. Insights into animal septins using recombinant human septin octamers 2 with distinct SEPT9 isoforms. Journal of Cell Science. 134 (15), (2021).
- Szuba, A., et al. Membrane binding controls ordered self-assembly of animal septins. eLife. 10, 63349(2021).
- Kinoshita, M. Assembly of mammalian septins. Journal of Biochemistry. 134 (4), 491-496 (2003).
- Connolly, D., et al. Septin 9 isoform expression, localization and epigenetic changes during human and mouse breast cancer progression. Breast Cancer Research. 13 (4), 76(2011).
- Connolly, D., et al. Septin 9 amplification and isoform-specific expression in peritumoral and tumor breast tissue. Biological Chemistry. 395 (2), 157-167 (2014).
- Estey, M. P., Di Ciano-Oliveira, C., Froese, C. D., Bejide, M. T., Trimble, W. S. Distinct roles of septins in cytokinesis: SEPT9 mediates midbody abscission. Journal of Cell Biology. 191 (4), 741-749 (2010).
- John, C. M., et al. The Caenorhabditis elegans septin complex is nonpolar. EMBO Journal. 26 (14), 3296-3307 (2007).
- Field, C. M., et al. A purified Drosophila septin complex forms filaments and exhibits GTPase activity. Journal of Cell Biology. 133 (3), 605-616 (1996).
- Bertin, A., et al. Saccharomyces cerevisiae septins: Supramolecular organization of heterooligomers and the mechanism of filament assembly. Proceedings of the National Academy of Sciences of the United States of America. 105 (24), 8274-8279 (2008).
- Sellin, M. E., Sandblad, L., Stenmark, S., Gullberg, M. Deciphering the rules governing assembly order of mammalian septin complexes. Molecular Biology of the Cell. 22 (17), 3152-3164 (2011).
- Akil, A., et al. Septin 9 induces lipid droplets growth by a phosphatidylinositol-5-phosphate and microtubule-dependent mechanism hijacked by HCV. Nature Communications. 7, 12203(2016).
- Tanaka-Takiguchi, Y., Kinoshita, M., Takiguchi, K. Septin-mediated uniform bracing of phospholipid membranes. Current Biology. 19 (2), 140-145 (2009).
- Omrane, M., et al. Septin 9 has two polybasic domains critical to septin filament assembly and Golgi integrity. iScience. 13, 138-153 (2019).
- Carim, S. C., Kechad, A., Hickson, G. R. X. Animal cell cytokinesis: The rho-dependent actomyosin-anilloseptin contractile ring as a membrane microdomain gathering, compressing, and sorting machine. Frontiers in Cell and Developmental Biology. 8, 575226(2020).
- El Amine, N., Kechad, A., Jananji, S., Hickson, G. R. X. Opposing actions of septins and Sticky on Anillin promote the transition from contractile to midbody ring. Journal of Cell Biology. 203 (3), 487-504 (2013).
- Renshaw, M. J., Liu, J., Lavoie, B. D., Wilde, A. Anillin-dependent organization of septin filaments promotes intercellular bridge elongation and Chmp4B targeting to the abscission site. Open Biology. 4 (1), 130190(2014).
- Vogt, E. T., et al. The ultrastructural organization of actin and myosin II filaments in the contractile ring: new support for an old model of cytokinesis. Molecular Biology of the Cell. 28 (5), 613-623 (2017).
- Mavrakis, M., et al. Septins promote F-actin ring formation by crosslinking actin filaments into curved bundles. Nature Cell Biology. 16 (4), 322-334 (2014).
- Karasmanis, E. P., et al. A septin double ring controls the spatiotemporal organization of the ESCRT machinery in cytokinetic abscission. Current Biology. 29 (13), 2174-2182 (2019).
- Hagiwara, A., et al. Submembranous septins as relatively stable components of actin-based membrane skeleton. Cytoskeleton. 68 (9), 512-525 (2011).
- Calvo, F., et al. Cdc42EP3/BORG2 and septin network enables mechano-transduction and the emergence of cancer-associated fibroblasts. Cell Reports. 13 (12), 2699-2714 (2015).
- Salameh, J., Cantaloube, I., Benoit, B., Poüs, C., Baillet, A. Cdc42 and its BORG2 and BORG3 effectors control the subcellular localization of septins between actin stress fibers and microtubules. Current Biology. 31 (18), 4088-4103 (2021).
- Kuzmić, M., et al. Septin-microtubule association via a motif unique to isoform 1 of septin 9 tunes stress fibers. Journal of Cell Science. 135 (1), (2022).
- Shindo, A., et al. Septin-dependent remodeling of cortical microtubule drives cell reshaping during epithelial wound healing. Journal of Cell Science. 131 (12), (2018).
- Hu, Q., Nelson, W. J., Spiliotis, E. T. Forchlorfenuron alters mammalian septin assembly, organization, and dynamics. Journal of Biological Chemistry. 283 (43), 29563-29571 (2008).
- Mavrakis, M., Tsai, F. C., Koenderink, G. H. Purification of recombinant human and Drosophila septin hexamers for TIRF assays of actin-septin filament assembly. Methods in Cell Biology. 136, 199-220 (2016).
- Nakos, K., Radler, M. R., Spiliotis, E. T. Septin 2/6/7 complexes tune microtubule plus-end growth and EB1 binding in a concentration- and filament-dependent manner. Molecular Biology of the Cell. 30 (23), 2913-2928 (2019).
- Kaplan, C., et al. Absolute arrangement of subunits in cytoskeletal septin filaments in cells measured by fluorescence microscopy. Nano Letters. 15 (6), 3859-3864 (2015).
- Castro, D. K. S. V., et al. A complete compendium of crystal structures for the human SEPT3 subgroup reveals functional plasticity at a specific septin interface. IUCrJ. 7, Pt 3 462-479 (2020).
- Jiao, F., Cannon, K. S., Lin, Y. -C., Gladfelter, A. S., Scheuring, S. The hierarchical assembly of septins revealed by high-speed AFM. Nature Communications. 11 (1), 1-13 (2020).
- Bertin, A., et al. Phosphatidylinositol-4,5-bisphosphate promotes budding yeast septin filament assembly and organization. Journal of Molecular Biology. 404 (4), 711-731 (2010).
- Bridges, A. A., Jentzsch, M. S., Oakes, P. W., Occhipinti, P., Gladfelter, A. S. Micron-scale plasma membrane curvature is recognized by the septin cytoskeleton. Journal of Cell Biology. 213 (1), 23-32 (2016).
- Beber, A., et al. Membrane reshaping by micrometric curvature sensitive septin filaments. Nature Communications. 10, 420(2019).
- Zhou, R., Shi, Y., Yang, G. Expression, purification, and enzymatic characterization of intramembrane proteases. Methods in Enzymology. 584, 127-155 (2017).
- Diebold, M. L., Fribourg, S., Koch, M., Metzger, T., Romier, C. Deciphering correct strategies for multiprotein complex assembly by co-expression: Application to complexes as large as the histone octamer. Journal of Structural Biology. 175 (2), 178-188 (2011).
- Lebedeva, M. A., Palmieri, E., Kukura, P., Fletcher, S. P. Emergence and rearrangement of dynamic supramolecular aggregates visualized by interferometric scattering microscopy. ACS Nano. 14 (9), 11160-11168 (2020).
- Ludtke, S. J., Baldwin, P. R., Chiu, W. EMAN: Semiautomated software for high-resolution single-particle reconstructions. Journal of Structural Biology. 128 (1), 82-97 (1999).
- Zivanov, J., et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife. 7, 42166(2018).
- Frank, J., et al. SPIDER and WEB: Processing and visualization of images in 3D electron microscopy and related fields. Journal of Structural Biology. 116 (1), 190-199 (1996).
- Young, G., Kukura, P. Interferometric scattering microscopy. Annual Review of Physical Chemistry. 70, 301-322 (2019).
- Young, G., et al. Quantitative mass imaging of single biological macromolecules. Science. 360 (6387), 423-427 (2018).
- Hernández-Rodríguez, Y., Momany, M. Posttranslational modifications and assembly of septin heteropolymers and higher-order structures. Current Opinion in Microbiology. 15 (6), 660-668 (2012).
- Ribet, D., et al. SUMOylation of human septins is critical for septin filament bundling and cytokinesis. Journal of Cell Biology. 216 (12), 4041-4052 (2017).
- Sinha, I., et al. Cyclin-dependent kinases control septin phosphorylation in Candida albicans hyphal development. Developmental Cell. 13 (3), 421-432 (2007).
- Sheffield, P. J., et al. Borg/Septin interactions and the assembly of mammalian septin heterodimers, trimers, and filaments. Journal of Biological Chemistry. 278 (5), 3483-3488 (2003).
- Joberty, G., et al. Borg proteins control septin organization and are negatively regulated by Cdc42. Nature Cell Biology. 3 (10), 861-866 (2001).
- Chen, X., Wang, K., Svitkina, T., Bi, E. Critical roles of a RhoGEF-anillin module in septin architectural remodeling during cytokinesis. Current Biology. 30 (8), 1477-1490 (2020).
- Kučera, O., et al. Anillin propels myosin-independent constriction of actin rings. Nature Communications. 12 (1), 1-12 (2021).
- Hsu, S. C., et al. Subunit composition, protein interactions, and structures of the mammalian brain sec6/8 complex and septin filaments. Neuron. 20 (6), 1111-1122 (1998).
- Olivi, L., et al. Towards a synthetic cell cycle. Nature Communications. 12 (1), 1-11 (2021).
- Hürtgen, D., Härtel, T., Murray, S. M., Sourjik, V., Schwille, P. Functional modules of minimal cell division for synthetic biology. Advanced Biosystems. 3 (6), 1800315(2019).
- Jia, H., Schwille, P. Bottom-up synthetic biology: Reconstitution in space and time. Current Opinion in Biotechnology. 60, 179-187 (2019).
- Cannon, K. S., Woods, B. L., Crutchley, J. M., Gladfelter, A. S. An amphipathic helix enables septins to sense micrometer-scale membrane curvature. The Journal of Cell Biology. 218 (4), 1128-1137 (2019).
- Lobato-Márquez, D., Mostowy, S. Septins recognize micron-scale membrane curvature. Journal of Cell Biology. 213 (1), 5-6 (2016).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유