
Method Article
BtM, um Datalogger aberto de baixo custo para estimar o teor de água de criptógamas Nonvascular
* Estes autores contribuíram igualmente
Neste Artigo
Resumo
Apresentamos um método simples e de baixo custo para construir um datalogger aberto que mede a condutância de criptógamas nonvascular juntamente com a temperatura ambiental e umidade. Descrevemos o projeto de hardware do datalogger e fornecer instruções de montagem passo a passo, a lista de software de registro necessária do código-fonte aberto, o código para executar o datalogger e um protocolo de calibração.
Resumo
Comunidades de criptógamas nonvascular, tais como musgos ou líquenes, são uma parte importante da biodiversidade terrestre, contribuindo para o Regulamento do carbono e nitrogênio ciclos em muitos ecossistemas. Sendo organismos poikilohydric, eles não ativamente controlar seu conteúdo de água interna e precisam de um ambiente úmido para ativar o seu metabolismo. Portanto, estudar relações de água de criptógamas nonvascular é crucial entender tanto seus padrões de diversidade e suas funções nos ecossistemas. Apresentamos o datalogger BtM, uma plataforma open-source de baixo custo para o estudo do teor de água dos nonvascular criptógamas. O datalogger é projetado para medir a temperatura, a humidade e condutância de até oito amostras simultaneamente. Nós fornecemos um desenho para uma placa de circuito impresso (PCB), um protocolo detalhado para montar os componentes e o código-fonte necessários. Tudo isso faz com que a montagem do datalogger BtM acessível a qualquer grupo de investigação, mesmo para aqueles sem conhecimento especializado anterior. Portanto, o projeto apresentado aqui tem o potencial para ajudar a popularizar o uso deste tipo de dispositivo entre ecologistas e biólogos de campo.
Introdução
Comunidades de nonvascular criptógamas são uma onipresente e uma parte muitas vezes negligenciada de ecossistemas terrestres1. Eles são constituídos por um agregado de organismos de pequeno porte muito diferentes, entre os quais briófitas e líquenes são proeminentes produtores primários. Estes dois grupos de organismos compartilham uma característica fisiológica que os torna únicos: poikilohydry, ou a incapacidade de controlar ativamente seu conteúdo de água interno. Isto tem profundas implicações para os processos fisiológicos desde o metabolismo cessa quando as células são secas em resposta a baixos níveis de umidade e recomeça quando o ambiente é úmido novamente2. Como consequência, criptógamas nonvascular evitar a seca em vez de lidar com isso2, que permite que essas comunidades a sobreviver em uma ampla gama de ambientes de desertos quentes e frios para os trópicos3,4.
Além disso, eles também mostram estruturas relativamente simples e têm baixas exigências nutricionais. Estas características tornam altamente sensíveis às condições microclimáticas. Na verdade, criptógamas nonvascular muitas vezes ocupam um espaço de nicho que é indisponível para plantas vasculares de um tamanho maior, formando ecossistemas em miniatura que constituem uma parte importante da diversidade do mundo. Briófitas e líquenes sozinhos incluem quase 40.000 espécies (ca. 20.000 briófitas lato sensu5,6 e ca. 20.000 líquenes7). Além disso, sua contribuição para a biodiversidade da terra é ainda maior, uma vez que suas comunidades oferecem abrigo para um grande número de espécies de fungos, incluindo uma flora diversificada de fungos micorrízicos e vida livre, cianobactérias N-fixando crescem como epífitas e uma miríade de microinvertebrados, tais como tardigrades, collembola, myriapods, insetos e ácaros que aproveitam da retenção na fonte de água, as condições de capacidade e tamponadas dentro destes ecossistemas em miniatura.
Comunidades de criptógamas avasculares também contribuam para a regulamentação dos ciclos biogeoquímicos do carbono. Em ecossistemas secos, as crostas do solo biológicos chamados cobrem até 40% de sua superfície8 em desempenham um papel importante como sumidouros de carbono. Uma análise recente estima-se que crostas de solo biológica de ambientes secos podem ser fixação 7% de todo o carbono fixado pela vegetação terrestre. Além disso, em outros ecossistemas onde briófitas ou líquenes ou uma combinação dos dois são produtores primários - como alguns sistemas de floresta boreal ou turfeiras - eles produzem entre 30% e 100% da produtividade primária líquida total10,11 . Eles também são importantes nos ecossistemas em que esses organismos não são dominantes, tais como: florestas temperadas. Na verdade, briófitas de chão de floresta tinham um equivalente de absorção de carbono anual de cerca de 10% floresta respiração de andar em uma floresta temperada da Nova Zelândia. Além disso, eles também são importantes para fixação do nitrogênio, desde as cianobactérias vivem como epífitas nessas comunidades poderiam ser concentrado quase 50% do montante global de nitrogênio biológico4.
Devido a dependência de sua atividade fisiológica, a disponibilidade de água no ambiente circundante, tanto a diversidade das comunidades nonvascular Criptógama e suas funções nos ecossistemas dependem fortemente de conteúdo de água2. Observe que, desde ativamente, eles não podem controlar o teor de água em seus tecidos, seus papéis no balanço de carbono e fixação de nitrogênio são acoplados com os ciclos de hidratação e desidratação e, portanto, dependem o intervalo e a periodicidade dos ciclos de seca-molhada. Assim, saber que o status de conteúdo de água desses organismos em tempo real é a chave para entender as funções executadas por criptógamas nos ecossistemas.
Apesar de sua importância, o desenvolvimento de métodos para medir a água atividade conteúda e fisiológica em organismos poikilohydric tem sido relativamente lenta. Em 1991, Coxson12 fez uma primeira abordagem para medir diretamente o teor de água dos líquenes. Depois disso, havia uma lacuna neste tipo de estudo até um desenvolvimento recente, quando vários trabalhos forneceram métodos para aproximar as medidas do estado fisiológico das criptógamas nonvascular13,14,15, 16. No entanto, tal conhecimento é ainda escasso e dispersos, e estas obras concentram-se sobretudo em solo crostas4,8. No entanto, briófitas e líquenes também desempenham um papel relevante em muitos outras ecossistemas, particularmente em regiões polares, temperadas e boreais1, e sua importância é significativa não só em comunidades de solo, mas também por comunidades epífitas que crescem em saxicolous comunidades em rochas e árvores. Esta falta de pesquisa está parcialmente ligada à ausência de dataloggers de medição disponíveis comercialmente, que obriga os grupos de pesquisa para construir seu próprio equipamento. Desenvolver um datalogger requer conhecimento específico que a maioria dos ecologistas não têm, portanto, aumenta substancialmente o custo de implementação de redes de medição relativamente grandes, necessárias para obter dados representativos sobre o desempenho do nonvascular criptógamas ao longo ambiental e gradientes de habitat.
Neste trabalho, apresentamos um método simples e de baixo custo para construir um datalogger capaz de medir a condutância de organismos criptogâmica nonvascular simultaneamente com a temperatura e a umidade. Ele está programado para gravar de forma autônoma para relativamente longos períodos de tempo (até dois meses) e é robusto o suficiente para suportar as condições severas de campo ao ar livre. Devido à sua simplicidade, será uma ferramenta útil para ecologistas e biólogos de campo sem formação especializada no desenvolvimento de dataloggers ou os grupos de pesquisa que faltam de pessoal especializado. Portanto, este datalogger tem potencial para ajudar a popularizar o uso deste tipo de dispositivo.
Desenvolvemos um datalogger de baixa potência e baixo custo capaz de medir a condutância de até oito diferentes fontes e gravar a temperatura ambiental e umidade relativa em simultâneo. O dispositivo é projetado depois projeto12 do Coxson e implementado sobre uma plataforma open-source (Tabela de materiais). O objetivo foi para priorizar a facilidade de montagem e poder eficiência e facilitar a manutenção das instalações a longo prazo. O design é derivado de um artigo por Sensores de ciência de prédio de fonte aberta (OSBSS)17. Este projeto foi modificado por incorporando circuitos adicionais para ler para fora a impedância de criptógamas e tornando-o mais compacto e fácil de fabricar.
O resultado é BtM (placa Bryolichen temperatura umidade), placa de circuito impresso aberto18. Cada placa é controlada por um microcontrolador de alto eficiência energética (Tabela de materiais). Dados ambientais de temperatura e umidade relativa são reunidos através de um sensor de temperatura e umidade que vem mergulhar e, além de seu baixo consumo de energia, tem uma relação preço / qualidade adequada.
A placa usa um protocolo de comunicação digital (padrão SPI serial) para gerenciar o ciclo de medição. Um relógio de tempo real (DS3234) montado em cada placa fornece sincronismo exato. Para reduzir o consumo de energia, o processador permanece no modo de espera na maioria das vezes. Precisam de cada vez dados a serem coletados, o relógio de tempo real ativa o processador e desencadeia o processo de registro. Relógio de tempo real também é usado para gravar com precisão a data e hora de cada caso de dados.
Até oito musgo e/ou Líquen amostras podem ser registradas em paralelo usando uma única placa de BtM. Quando o experimento é configurado, duas sondas de eletrodo de crocodilo-clip são aplicadas para cada amostra de musgo/líquen. Em seguida, um divisor de tensão entre cada eletrodo e uma referência de resistor com um valor conhecido (330 KΩ neste caso) são usados. Esse valor de resistor foi selecionado por meio de calibração e com base em medidas anteriores das criptógamas. Fornece uma resolução de uma ordem de magnitude em torno do valor de referência (100-1.000 KΩ). A queda de tensão é armazenada em buffer e então leia com o microcontrolador usando suas portas analógicas (A0 - A7)18. A tensão é calculada aplicando a seguinte fórmula.
Vi = (ADCi x VCC) / 1023
Aqui, ADCi é matéria-prima o valor do ADC (conversor analógico-Digital) do canal, VCC é a tensão de alimentação (3.3 V neste caso) e 1023 é o intervalo de saída o ADC. A tensão resultante Vi é usada em combinação com a lei de Ohm para calcular a resistência (Ri, Ω) e condutância (G, S) de cada amostra de musgo.
Ri = (VCC-x RL) / Vi - RL
G = 1 / Ri
Aqui, RL é o valor da referência do resistor (330 KΩ neste caso). Software de bordo do microcontrolador incorpora todas estas equações, para que ele se pode registrar diretamente os valores de resistência e condutância.
O Conselho também recolhe as medições da temperatura ambiente e umidade usando sensores. Em seguida, cada ponto de dados é gravado em um arquivo de log em um cartão microSD. Um microSD TransFlash placa de fuga foi montado em cada placa de BtM para essa finalidade. Finalmente, o cartão microSD pode ser coletado manualmente após a experiência. Todos os pontos de dados podem ser transferidos para um computador para posterior análise.
Protocolo
1. montagem do Datalogger
- Prepare um ferro de solda e um carretel de arame de solda. Espere para aquecer o ferro de soldar e umedeça a esponja de limpeza.
- Cortar as tiras de cabeçalho do pino até o comprimento desejado e solda-los nos encaixes para os módulos de fuga de relógio e microSD RTC, o microcontrolador e o sensor de temperatura e umidade.
- Para solda, pré-aqueça a junção desejada com a ponta do ferro de soldar.
- Em seguida, aplique uma pequena quantidade de material desde o fio de solda, suficiente para encher a junção.
- Finalmente, retire o ferro de soldar e espere a junção refrigerar para baixo.
- Montar os componentes para a placa de circuito, usando o mesmo procedimento como na etapa 1.2, seguindo as marcações de PCB e as referências do componente especificadas na Tabela de materiais (veja a Figura 1 para um esquema de montagem).
- Primeiro, solde os resistores. Em seguida, solde os soquetes para os amplificadores operacionais, o sensor de SHT7X e os módulos de fuga de relógio e microSD RTC.
- Em seguida, solde os dois transistores. O Conselho também precisa ser soldados agora, usando cabeçalhos de pino. Finalmente, solde os conectores para a placa.
- Solde o sensor de umidade/temperatura de SHT7X em um cabo de extensão ou cabeçalho de pin para reforçar os contatos.
- Prepare um multímetro no modo de teste de continuidade teste ou condutividade. Use o multímetro para verificar que não há nenhum curto-circuito entre qualquer um dos pinos ou conexões.
- Doublecheck fornecem os polos positivo e negativos do poder. Além disso, verifique se que cada junta de solda cria uma conexão estável entre os pinos do componente e as faixas de cobre do circuito.
Nota: Este passo é muito importante; não ignorá-lo.
- Doublecheck fornecem os polos positivo e negativos do poder. Além disso, verifique se que cada junta de solda cria uma conexão estável entre os pinos do componente e as faixas de cobre do circuito.
- Conecte os terminais da bateria e grampos de cabo para a placa usando uma chave de fenda.
- Primeiro, use qualquer ferramenta de corte para decapar ~ 4 mm de cada extremidade do fio, expondo o núcleo condutor. Em seguida, introduzir o terminal apropriado a cada cabo e aperte o parafuso com a chave de fenda Phillips.
- Assegurar e doublecheck a polaridade dos cabos, especialmente aqueles do poder fornecer. Testar a força da conexão, puxando os cabos ligeiramente, verificar que tudo está conectado firmemente.
- Para reduzir ainda mais o consumo de energia, retire o LED de alimentação da placa microcontrolador ou desoldering ou cortando o diodo LED da placa.
- Finalmente, monte a placa de BtM em um compartimento à prova de intempéries para manter a umidade longe da eletrônica.
- Encaixe o recinto com a bateria, conectando-o aos terminais positivos e negativos. Monte o sensor de umidade/temperatura fora da caixa, deixando-o conectado à placa do BtM.
- Rota a oito pares de crocodilo grampeia necessário para medições de condutância para o exterior do compartimento à prova de intempéries. Ontem, clip cada vertente de musgo com os clipes de crocodilo.
2. carregar o Software
- Baixe e instale o ambiente de desenvolvimento integrado (IDE) 1.0.6 o site19. O microcontrolador usado é uma plataforma de computação física open-source e vem com sua própria IDE. É importante baixar a versão adequada, uma vez que existem problemas de compatibilidade conhecidos com alguns das bibliotecas necessárias.
- Baixar as bibliotecas necessárias do repositório GitHub18: DS3234, DS3234lib3, economia, SdFat e Sensirion.
- Baixe o código fonte principal para o datalogger o GitHub repositório18.
- Abra o arquivo clock.ino para configurar a data e hora atual. Edite os parâmetros para a função RTC.setDateTime , com o tempo atual e a data usando o seguinte formato:
RTC.setDateTime(DD,MM,YY,hh,mm,ss); Data: DD/MM/AA hh
Aqui, DD é o dia, MM é o mês, YY é o ano, hh é a hora, mm é minutos e ss é segundos. - Em seguida, enviar o programa do relógio à diretoria BtM, conectando o adaptador USB-para-Serial (fuga FTDI) os portos de programação de microcontrolador e usando um cabo mini-USB-para-USB para ligar a câmara ao computador. Finalmente, prima primeiro verificar e, em seguida, fazer o Upload no IDE.
- Abra o projeto do datalog no IDE e modificar o arquivo de datalog.ino. Configurar a hora de início para o agente de log edição as seguintes variáveis:
int dayStart = DD, hourStart = hh, minStart = mm
Aqui, DD é o número do dia, hh é a hora inicial das medições e mm o minuto do início.
Nota: O código para marcar um horário específico deve ficar assim:
RTC.setDateTime(DD,MM,YY,hh,mm,ss); / / data de 12/01/17 12:00 - Defina o intervalo entre medições (em segundos), modificando o valor do intervalovariável.
3. configuração das sondas de medição
- Coloque os clipes de crocodilo em uma posição central das comunidades nos casos de briófitas, líquenes fruticose e líquenes (Figura 2). Fruticose líquenes, anexe os clipes no talo e, musgos, diretamente na haste de um indivíduo. No caso de líquenes, coloque os clipes na fronteira do talo.
- Mantenha uma distância mínima de ca. 5 mm entre os eletrodos. Certifique-se de que os grampos não são facilmente destacados antes de iniciar as medições.
4. calibração para medições de condutância
- Para garantir que os espécimes estão secos, execute a calibração ao meio-dia, em um dia com umidade relativa de ar baixa, precedido pelo menos um e de preferência dois, dias de secos.
- Selecione uma comunidade de musgo ou líquenes que é saudável e bem estruturado.
- Conecte o datalogger para o musgo ou um líquen, seguindo os passos na secção 3 do presente protocolo.
- Começar as medições (Ligue o datalogger) e deixar a placa de BtM correndo por aproximadamente 3 min estabilizar os valores registrados.
- Execute um teste precalibration para estimar a quantidade de água necessária em cada evento de irrigação. Conecte os clipes para a amostra e adicionar água até a condutância atinge um valor que não aumenta com a adição de água. Este é o valor de condutância máxima de que a amostra. Esse valor será usado para estabelecer as etapas de regas da calibração (ver passo 4.7.1).
- Espere até que as medidas de condutância retornam aos valores iniciais (as amostras estão secas).
- Em seguida, adicione água sequencialmente com um pulverizador pequeno.
- Umedeça as amostras com uma quantidade de água equivalente a 1/10 da quantidade de água necessária para atingir a máxima condutância (consulte a etapa 4.5) na amostra.
- Espere até que o musgo ou Líquen totalmente absorve a água e as medições de condutância são estáveis antes de molhar novamente (~ 1 min entre cada evento de irrigação).
- Repita até a condutância atinge o valor máximo (saturação) e o musgo ou Líquen é totalmente hidratado.
Nota: Cada teste de calibração deve levar em torno de 15 min, dependendo do intervalo entre as regas, que devem ser 1-2 min.
- Após terminar a calibração, levar o cartão microSD do tabuleiro BtM e copie o arquivo de dados para um computador.
Nota: Os valores registrados em seguida podem ser usados como base para os experimentos. Também é necessário fazer esta etapa para verificar que a afinação está corretamente registrando a condutância das amostras, apenas antes de executar a experiência.
5. alternativa calibração para experimentos de laboratório
- Totalmente hidratar a comunidade de musgo ou Líquen até observa-se um excesso de água externa. Para garantir que a Comunidade está totalmente hidratada, manter a Comunidade úmida por 30 min.
- Conecte o datalogger para o musgo ou um líquen, seguindo os passos na secção 3 do presente protocolo.
- Começar as medições e deixar a placa de BtM correndo por aproximadamente 3 min estabilizar os valores registrados.
- Espere até que a condutância atinge o valor mínimo (dessecação) e o musgo ou Líquen já não conduz eletricidade.
Nota: Cada calibração poderia durar pelo menos 1 h, mas a duração é altamente variável, dependendo da espécie. As medidas devem ser tomadas até um valor mínimo de condutância é alcançado.
Resultados
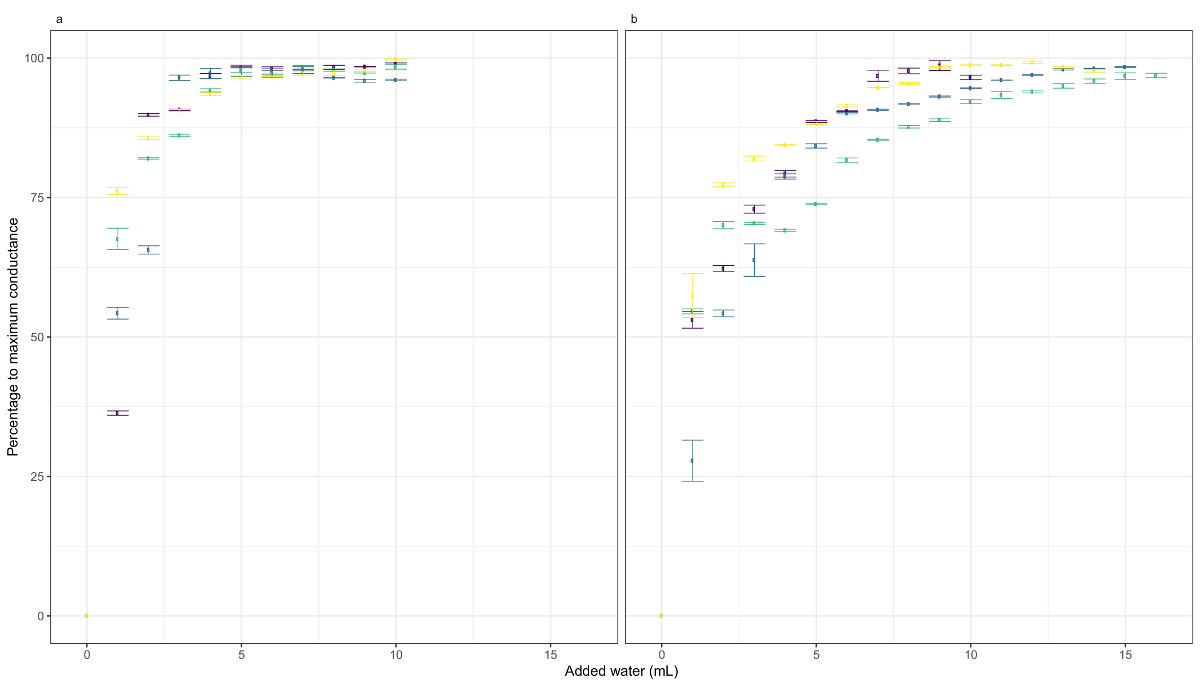
Analisamos as alterações na condutância em duas espécies de musgos, Dicranum scoparium Hedw. e Homalothecium aureum (Spruce) H. Rob. (Figura 3), durante o processo de calibração em condições de laboratório. Tapetes de musgos os dois foram mantidos por 24 h em sílica gel e colocados em um substrato artificial (ou seja, pasta (ouate)) que manteve sua estrutura original (Figura 2). Em seguida, as amostras foram regada 15 x 20 x com um spray em intervalos de 1 min. Cada evento de irrigação consistia de aprox. 0,1 mL de água. Em ambas as espécies, uma alta correlação entre a água adicionada e a condutância de amostra (d. scoparium rS = 0,88, p < 0,001; H. aureum rs = 0,87, p < 0,001) foi observada. Houve um aumento elevado a condutância (de 0% a 25% pelo menos) apenas na primeira água adição e as medidas atingiu sua máxima condutância em 4 mL de d. scoparium e 10 mL de H. aureum. É importante observar que a relação entre a quantidade de água e condutância é logarítmica. Portanto, os valores de condutância precisam ser transformada para ter uma relação linear entre ambas as variáveis, e sua relação deve ser modelada usando regressão não-linear.
Encontramos alguns variabilidade entre as amostras (ver as diferentes cores nas Figs. 3a e 3b), apesar de todas as amostras pertencentes à mesma espécie desenhou uma curva semelhante. A variação entre amostras pode ser atribuída às diferenças de biomassa e morfologia dos patches. Amostras no campo são muito prováveis mostrar este tipo de variabilidade, portanto tomar várias medidas de cada tipo de comunidade é recomendado. Não surpreendentemente, a maior variabilidade foi encontrada entre as espécies, desde espécies diferem em vários traços fundamentais (por exemplo, a agregação dos tapetes ou morfologia). Para controlar para intrae a variabilidade entre espécies, recomendamos calibrar cada clipe até alcançar os valores de condutância máxima e, em seguida, rescaling os resultados para cada clipe para que os valores vão de 0 a 100. Considere que absoluta condutância valores dependem da distância entre os grampos e a condutância basal das hastes, então os valores fornecem não são directamente comparáveis.
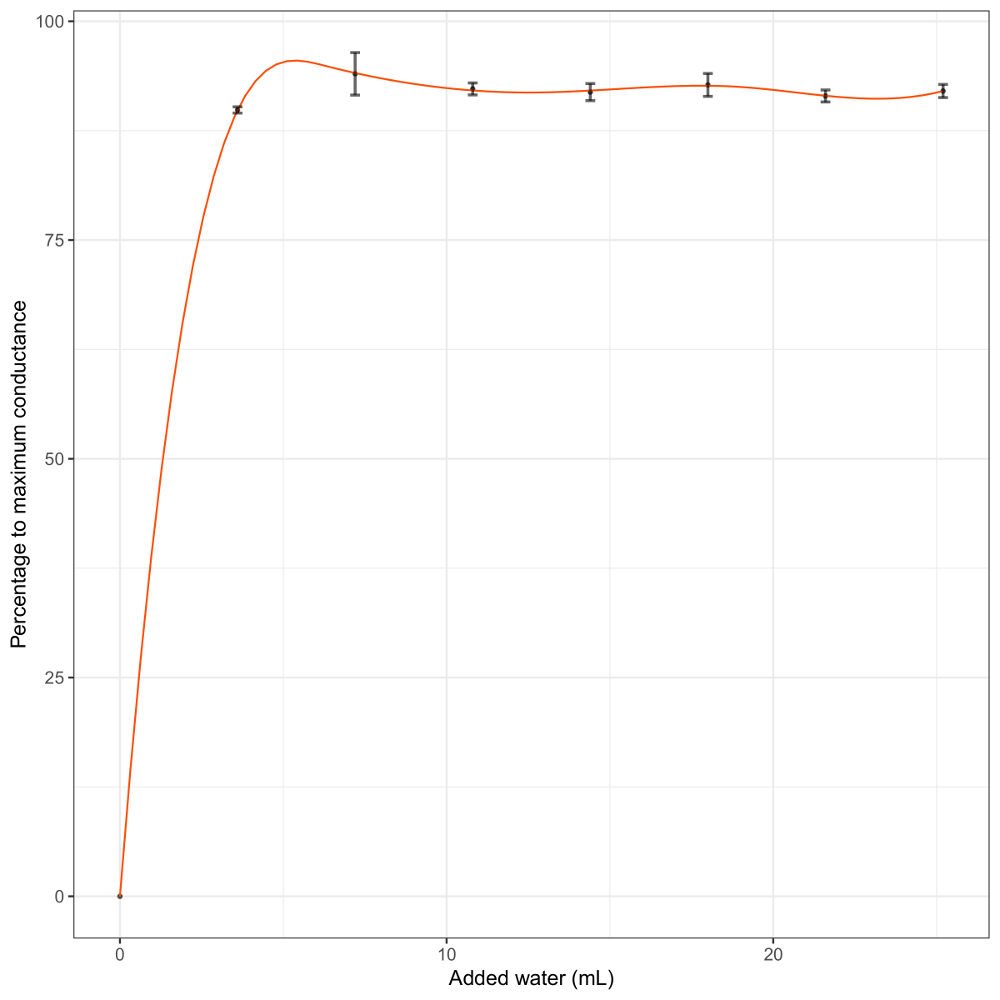
A quantidade de água adicionada em cada evento de irrigação do processo de calibração é crucial e fortemente afetará os resultados. Aqui, o objectivo era ter vários eventos molhando na faixa de precisão máxima do BtM. Nós apresentamos um exemplo de uma curva de calibração quando muita água é adicionada em cada etapa (Figura 4). Se a amostra é overwatered o primeiro evento de irrigação, o aumento da condutância não pode ser apreciado e a calibração será imprecisa. Isso pode levar a vieses na faixa onde nonvascular criptógamas são ativos, que são as mais interessantes as medições feitas com o BtM.
Também analisamos a curva de dessecação das mesmas duas espécies (H. aureum e d. scoparium), para fornecer um procedimento alternativo de calibração. Tapetes de musgos os dois eram regadas durante a noite para garantir que eles foram totalmente saturados. Em seguida, uma haste representativa de cada esteira foi extraída e colocada em um ambiente estável e controlado e a condutância registou-se continuamente. Quanto a outra medida de calibração, os valores de condutância precisam ser transformada para ter uma relação linear entre ambas as variáveis, e sua relação deve ser modelada usando regressão não-linear.
Figuras 5a e 5b mostram as curvas de dessecação de H. aureum e a d. scoparium variabilidade entre amostras da mesma espécie. O intra e interespécie variabilidade encontrada eram muito grandes e, como o procedimento de calibração, poderiam ser atribuídas às diferenças de biomassa e morfologia de cada haste. Para controlar, por isso, recomendamos a realização de pelo menos três medições por espécie. Valores de condutância absoluta não são directamente comparáveis neste procedimento de calibração, como eles também dependem da distância entre os clipes e condutância basal das hastes.
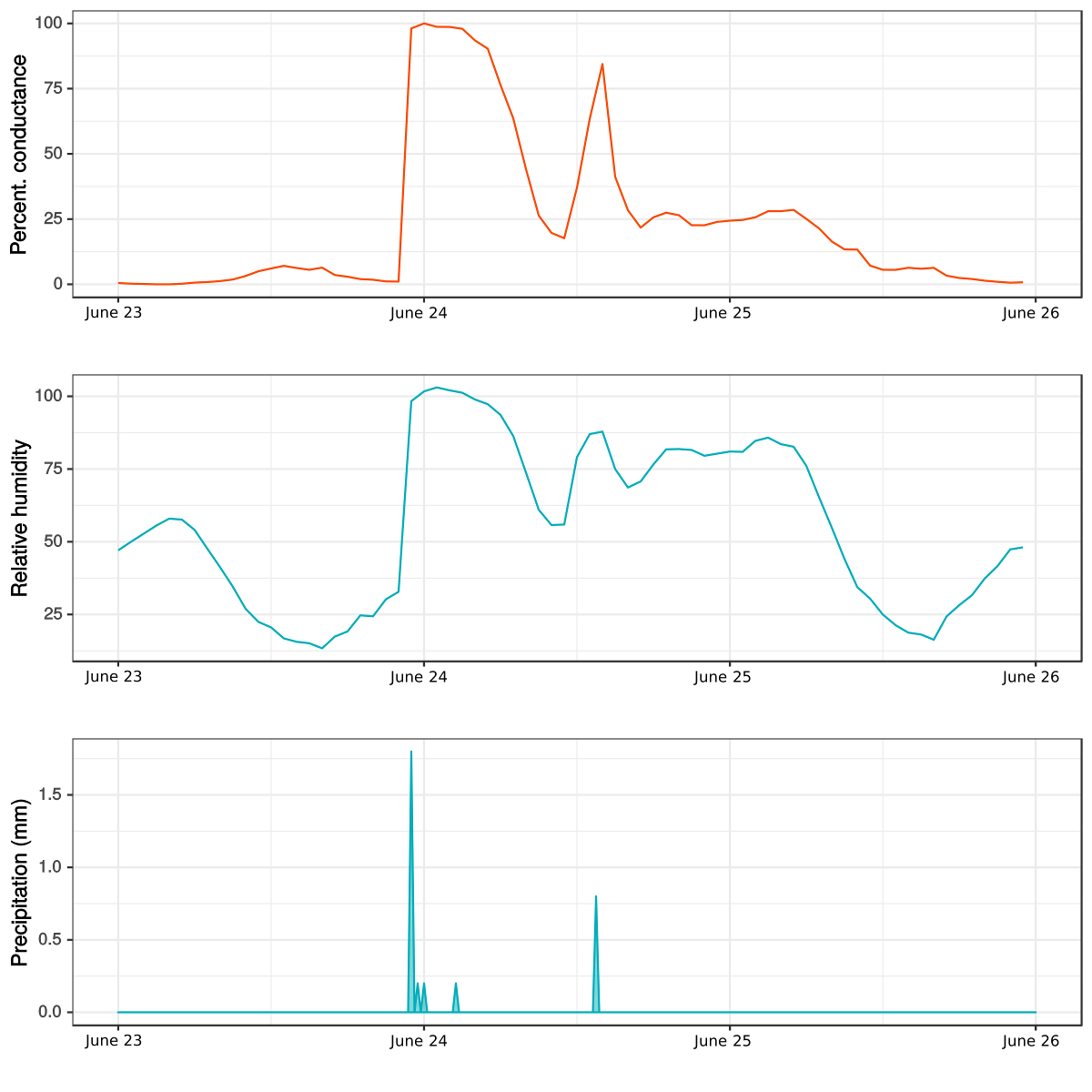
Nós apresentamos um exemplo de dados de campo após um evento de chuva ocorreu entre 23 a 26 de junho de 2014. Mostramos a variação diária na percentagem de condutância (Figura 6um), (Figura 6b) umidade relativa do ar e precipitação (Figura 6c) para uma espécie de musgo (Syntrichia ruralis (Hedw.) F. Weber & Mohr d.). Havia uma forte relação entre a condutância de musgo, os eventos de precipitação e umidade relativa do ar. Durante o período analisado, houve dois picos na condutância e umidade em consequência de dois eventos de precipitação. O primeiro deles ocorreu pouco antes da meia-noite de 23 de junho e o segundo depois do meio-dia de 24 de junho. Cerca de 8 h após o primeiro evento de chuva, observamos uma diminuição da umidade relativa do ar, seguido por uma queda repentina na condutância de musgo que vai abaixo de 25%. O segundo evento de chuva era menor e, consequentemente, produziu um pico menor na condutância. Após este evento de chuva, o musgo não secar imediatamente mas ficou hidratado, enquanto a umidade foi superior a 75%.

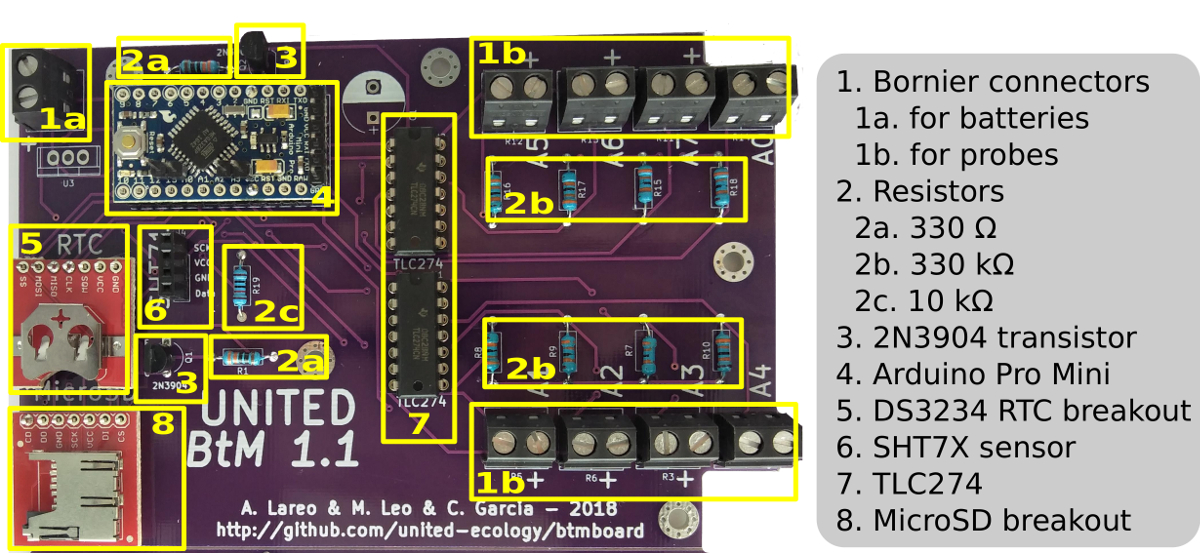
Figura 1 : Esquema de montagem do datalogger BtM. O esquema inclui uma foto da placa BtM e a colocação de cada componente na placa. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2 : Colocação correta dos grampos em um musgo (Homalothecium aureum). A imagem mostra como colocar os clipes para manter uma distância mínima entre os clipes sem danificar os briófitos. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3 : A resposta de condutância para adição de água. Estes painéis mostram a resposta de condutância à água adição em (uma) Dicranum scoparium e (b) H. aureum. As cores mostram as diferentes repetições. Os pontos de dados são a média da condutância log-transformado em um intervalo entre 10 e 30 s após o evento de regando. As barras de erro representam o desvio padrão dos dados nesse intervalo. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4 : Resposta de condutância log-transformado para adição de água em scoparium d. quando a quantidade de água adicionada é grande demais para permitir a calibração. As barras de erro representam o desvio padrão dos dados nesse intervalo. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5 : Curvas de dessecação. Estes painéis mostram as curvas de dessecação de (um) d. scoparium e (b), H. aureum. Os pontos de dados são a média da condutância log-transformadas medido cada show 30 pontos preto s. a média das três repetições e as barras de erro representam o desvio padrão dos dados nesse intervalo. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6 : Variação diária em de um musgo condutância (Syntrichia ruralis), precipitação e umidade relativa do ar. As medidas foram tomadas em comunidades de solo do Cantoblanco, Campus da Universidad Autónoma de Madrid, Espanha. A condutância e a umidade relativa do ar foram medidos com o protótipo de BtM, enquanto os dados de precipitação vem de uma estação meteorológica colocada a poucos metros de distância do local de medição. Clique aqui para ver uma versão maior desta figura.
{kind=link}
| Data/hora | Temp(C) | RH(%) | Conductance(KMho) |
| 03/11/18 12:00 | 26,6 | 66,6 | 139.53 |
| 03/11/18 12:00 | 26,6 | 66,4 | 167.92 |
| 03/11/18 12:00 | 26,8 | 66,4 | 199.14 |
| 03/11/18 12:00 | 26,9 | 66,4 | 212.75 |
| 03/11/18 12:00 | 26,6 | 66,6 | 217.15 |
| 03/11/18 12:01 | 26,9 | 66,7 | 218.93 |
| 03/11/18 12:01 | 27 | 66,8 | 139.53 |
| 03/11/18 12:01 | 27.1 | 66,9 | 164.28 |
| 03/11/18 12:01 | 27.1 | 67,3 | 194.21 |
| 03/11/18 12:01 | 27.3 | 67,3 | 209.28 |
Tabela 1: Exemplo da saída BtM.
Discussão
A nosso conhecimento, isto é a primeira vez que um datalogger para medir temperatura, umidade e condutância simultaneamente como um proxy do teor de água dos organismos poikilohydric foi concebido baseado em uma plataforma de acesso aberto. O datalogger BtM é fácil de construir e de baixo custo e também fornece medições de qualidade de dados de impedância usando o mínimo de energia, temperatura e umidade do ar.
A montagem simples é uma das principais vantagens deste datalogger. Como é um projeto open-source, nós fornecemos o software de registro de dados e um esquema detalhado de sua estrutura, juntamente com um manual de técnica para a construção de um datalogger de BtM ready-to-use. Isto torna o método acessível a qualquer grupo de investigação, mesmo para aqueles que não trabalham com um engenheiro ou técnicos especializados. Além disso, a montagem de cada datalogger quase requer 1 hora se o circuito impresso placa é usado e cerca de 4 horas se o circuito é montado pelos pesquisadores. Além disso, o datalogger BtM é altamente rentável. O custo estimado dos componentes de cada unidade é de cerca de 100 euros, um preço bastante baixo que pode ser reduzido ainda mais em projetos de grande escala, pela montagem de lotes de vários dataloggers.
Embora tenha havido vários recentes desenvolvimentos metodológicos vistos implementar dispositivos que medem diferentes aspectos relacionados com a atividade fisiológica das comunidades nonvascular Criptógama, a BtM preenche uma lacuna de conhecimentos importantes. Raggio et al 15 empregam Moni-Da, um sistema de monitoramento que obtém informações fisiológicas e microclimáticas. A atividade fisiológica é coletada através de uma fluorescência de clorofila, um método amplamente utilizado no laboratório para estimar a atividade de organismos fotossintéticos. Embora este método seja altamente preciso, é significativamente mais caro que o datalogger BtM. Além disso, o sistema de monitoramento é um produto de empresa privada, o que reduz a autonomia do grupo de pesquisa.
Os dois outros métodos que recentemente foram publicados baseiam-se também a estimar o teor de água de nonvascular criptógamas. O primeiro é baseado em medições térmicas (um método de pulso (DPHP) dual-sonda térmica). Apesar de resultados promissores recentemente foram mostrados por jovem et al 16, a falta de qualquer regime específico no jornal faz montá-lo sem conhecimento especializado altamente desafiador. Por fim, Weber et al 14 apresentou um sensor chamado a sonda de umidade de biocrust (BWP), que é muito parecida com o datalogger BtM. No entanto, eles não fornecem qualquer esquema para sua construção, o que dificulta a possibilidade de construir o datalogger sem a ajuda de um especialista. Podemos superar esse problema, fornecendo não só o regime de construção, mas também a placa de circuito para montar o datalogger. Curiosamente, a BtM pode ser facilmente modificado para medir biocrusts, indivíduos isolados ou almofadas, apenas mudando os clipes de crocodilo (para indivíduos/coxins de líquen ou briófitos) para pinos de eletrodo de liga de cobre (para o biocrusts). Se necessário, pode ser substituída apenas parte dos crocodilos, permitindo comparações diretas entre os tipos de sonda de medição de dois.
Ao interpretar os resultados, a relação entre atividade e conteúdo de água deverá ser cuidadosamente dirigida, porque o BtM não mede diretamente da fotossíntese. Fotossíntese e atividade estão intimamente relacionados em criptógamas nonvascular desde que um organismo poikilohydric seco em cessar metabólica e molhado é ativo. No entanto, o grau de atividade fotossintética não pode ser inferido diretamente o conteúdo de água, apesar de uma maior atividade metabólica - e, assim, uma maior atividade fotossintética - podem ser esperados em um organismo bem hidratado.
Passos críticos:
Apesar da facilidade de montagem, existem alguns passos críticos no protocolo que deve ser cuidadosamente abordado pelos pesquisadores durante a montagem do sensor. Primeiro, como enfatizado no protocolo, é muito fácil produzir curtos-circuitos quando solda, que, na pior das hipóteses, pode resultar em sérios danos para o microcontrolador. É muito importante para verificar sua presença com um multímetro e removê-los antes de ligar as baterias. Nós recomendamos usar o design de PCB fornecido desde significativamente simplifica o processo e pode ser a melhor opção para superar esse problema. Em segundo lugar, nem todas as versões do IDE são compatíveis com as bibliotecas necessárias para este datalogger. É importante baixar o apropriado um (1.0.6) para evitar qualquer compatibilidade emite. Em terceiro lugar, é importante observar a polaridade das pilhas. Uma inversão de polaridade pode resultar em sérios danos ao hardware. Em quarto lugar, a calibração é uma etapa crítica. O datalogger BtM é projetado para que a resolução maior coincide com o momento em que a Criptógama vai do seco-húmido estado. Isto implica que os valores de condutância saturam muito antes que a amostra está saturada em água. No entanto, se o estudo em questão requer uma exatidão mais elevada em torno de outros valores, ele pode ser modificado. Medidas, para além de uma ordem de magnitude desta referência requerem o resistor a ser alterada e um processo de recalibração (veja abaixo). Como a temperatura ambiental pode afetar a precisão das medições, recomendamos tendo em conta este factor quando calibrando. Para isso, a calibração deve ser feita em temperaturas baixas para verificar se há alterações na precisão e estabilidade (ver Coxson12 para efeitos de temperatura).
Modificações:
Embora a maioria dos componentes do BtM é fixos, alguns podem ser facilmente modificado no campo sem resoldagem. A modificação mais simples é substituir os clipes de crocodilo para outros sistemas de medida ou sonda. Por exemplo, em vez dos clipes de crocodilo, uma sonda com dois pinos, como o sugerido no Weber et al . 14, pode ser usado.
Em ambientes remotos, onde mudar as pilhas pode não ser possível dentro a frequência necessária, baterias poderiam ser complementadas com um painel solar para alimentar o datalogger BtM por períodos mais longos.
Alterando os resistores de referência utilizados para medir a condutância, o posto de maior resolução pode ser facilmente modificado para valores superiores ou inferiores. Se modificado, recomendamos uma recalibração precisa. Também, no código-fonte, a variável de RValue , que está programada para um valor de resistor de 330 KΩ, deve ser atribuída para o novo valor correspondente (datalog.ino).
Conclusão:
Criptógama nonvascular comunidades são altamente diversificadas e jogam um número de diferentes papéis ecológicos fundamentais, para que compreender as suas relações com o ambiente abiótico é uma questão crucial. O datalogger BtM tem vários aplicativos que ajudarão a avançar o conhecimento dessas relações. Por exemplo, isso ajudará a aprofundar conhecimentos sobre as condições onde estes organismos estão agindo como sumidouros de carbono ou fontes de carbono. As flutuações entre essas duas funções são fortemente relacionadas às condições abióticas tais como temperatura e umidade3, mas grandes quantidades de dados são necessários para descrever e compreender as variações dessa relação em escala global. Isto exige redes de sensores densa que são possíveis apenas se eles contam com equipamentos de baixo custo e fácil de implementar.
Para resumir, este dispositivo é uma ferramenta útil para grupos de pesquisa ecológica e supera as limitações técnicas de concepção e construção de um datalogger. A combinação destes dois factores pode levar a uma popularização do uso de dataloggers para medir as relações de água do nonvascular criptógamas em situ. Isto, por sua vez, pode aumentar o estabelecimento de redes de monitoramento de médio e longo prazo. O desenvolvimento destas redes é essencial para avaliar a resposta de criptógamas nonvascular locais e regionais factores ambientais, bem como a determinar seu papel nos processos do ecossistema (por exemplo, ciclos de nutrientes, montagem de comunidade) e sua resposta mais provável, tendo em conta as alterações em fatores climáticos e antrópicos associados com a mudança global.
Divulgações
Os autores não têm nada para divulgar.
Agradecimentos
Os autores são gratos a Manuel Molina (UAM) e Cristina Ronquillo (MNCN-CSIC) pela ajuda fornecida durante os ensaios de calibração e a Estébanez de Belén (UAM) por sua ajuda durante as campanhas de amostragem.
Materiais
| Name | Company | Catalog Number | Comments |
| BtMboard circuit (PCB) | 1 | ||
| Arduino Pro Mini 328 3.3 V (APM) | Arduino | 1 | |
| FTDI Basic Breakout | SparkFun | 1 | |
| MiniUSB to USB cable adapter | 1 | ||
| TLC274 operational amplifier | Texas Instruments | 2 | |
| 2.54 mm breakout pin strip | 1 | ||
| 330 KOhm resistor | 8 | ||
| 330 Ohm resistor | 2 | ||
| 10 KOhm resistor | 1 | ||
| 2N3904 Transistor | 2 | ||
| Bornier connector, 2x1 5.08 mm | 9 | ||
| 1.5 V AA battery | 3 | ||
| 3xAA battery holder with switch | 1 | ||
| Sensirion SHT71 | Sensirion | 1 | |
| DS3234 RTC Breakout (clock) | SparkFun | 1 | |
| CR1225 3 V Coin-cell battery | 1 | ||
| MicroSD Transflash breakout | SparkFun | 1 | |
| Crocodile clip connector | 16 | ||
| Weatherproof enclosure box | 1 | ||
| 12 AWG stranded cable spool | 1 | ||
| Cutting pliers | 1 | ||
| 30 W soldering iron | 1 | ||
| Solder wire spool | 1 | ||
| Arduino IDE 1.0.6 | Arduino | 1 | |
| Arduino library DS3234 | Arduino | 1 | |
| Arduino library DS3234lib3 | Arduino | 1 | |
| Arduino library Powersaver | Arduino | 1 | |
| Arduino library SdFat | Arduino | 1 | |
| Arduino library Sensirion | Arduino | 1 |
Referências
- Fontaneto, D., Hortal, J., Ogilvie, L. A., Hirsch, P. R. Microbial Biogeography: Is Everything Small Everywhere. Microbial Ecological Theory: Current Perspectives. , 87-98 (2012).
- Proctor, M. C. F., et al. Desiccation-tolerance in bryophytes: a review. The Bryologist. 110 (4), 595-621 (2007).
- Lindo, Z., Gonzalez, A. The Bryosphere: An Integral and Influential Component of the Earth's Biosphere. Ecosystems. 13 (4), 612-627 (2010).
- Elbert, W., et al. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nature Geoscience. 5, 459-462 (2012).
- Magill, R. E. Moss diversity: New look at old numbers. Phytotaxa. 9 (1), 167-174 (2014).
- Söderström, L., et al. World checklist of hornworts and liverworts. PhytoKeys. (59), 1-828 (2016).
- Lücking, R., Hodkinson, B. P., Leavitt, S. D. The 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota - Approaching one thousand genera. The Bryologist. 119 (4), 361-416 (2016).
- Bowker, M. A. Biological Soil Crust Rehabilitation in Theory and Practice: An Underexploited Opportunity. Restoration Ecology. 15 (1), 13-23 (2007).
- Wilske, B., et al. The CO2 exchange of biological soil crusts in a semiarid grass-shrubland at the northern transition zone of the Negev desert, Israel. Biogeosciences Discussions. 5 (3), 1969-2001 (2008).
- Wardle, D. A., et al. Linking vegetation change, carbon sequestration and biodiversity: insights from island ecosystems in a long-term natural experiment. Journal of Ecology. 100 (1), 16-30 (2012).
- Lindo, Z., Nilsson, M. -. C., Gundale, M. J. Bryophyte-cyanobacteria associations as regulators of the northern latitude carbon balance in response to global change. Global Change Biology. 19 (7), 2022-2035 (2013).
- Coxson, D. S. Impedance Measurement of Thallus Moisture Content in Lichens. The Lichenologist. 23 (1), 77-84 (1991).
- Raggio, J., et al. Continuous chlorophyll fluorescence, gas exchange and microclimate monitoring in a natural soil crust habitat in Tabernas badlands, Almeria, Spain: progressing towards a model to understand productivity. Biodiversity and Conservation. 23 (7), 1809-1826 (2014).
- Weber, B., et al. Development and calibration of a novel sensor to quantify the water content of surface soils and biological soil crusts. Methods in Ecology and Evolution. 7 (1), 14-22 (2016).
- Raggio, J., et al. Metabolic activity duration can be effectively predicted from macroclimatic data for biological soil crust habitats across Europe. Geoderma. 306, 10-17 (2017).
- Young, M. H., Fenstermaker, L. F., Belnap, J. Monitoring water content dynamics of biological soil crusts. Journal of Arid Environments. 142, 41-49 (2017).
- . GitHub - united-ecology/btmboard Available from: https://github.com/united-ecology/btmboard (2018)
- . Arduino - Software Available from: https://www.arduino.cc/en/Main/Software (2018)
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados