Method Article
Um método para rastrear a evolução temporal dos potenciais evocados de estado estacionário
Neste Artigo
Resumo
Um protocolo para avaliar a evolução do tempo do arrastamento neural aos estímulos repetitivos externos é apresentado. As gravações de estado estacionário da mesma condição experimental são adquiridas e médias no tempo-domínio. A dinâmica do estado estacionário é analisada traçando a amplitude da resposta em função do tempo.
Resumo
A arrastamento neural refere-se à sincronização da atividade neural com a periodicidade dos estímulos sensoriais. Esta sincronização define a geração de respostas evocadas de estado estacionário (ou seja, oscilações na fase de eletroencefalograma-bloqueado para os estímulos de condução). A interpretação clássica da amplitude das respostas evocadas de estado estacionário assume uma resposta neural invariante do tempo estereotipada mais flutuações de fundo aleatórias, de tal forma que a média de apresentações repetidas do estímulo recupera a resposta estereotipada. Essa abordagem ignora a dinâmica do estado estacionário, como no caso da adaptação eliciada por exposições prolongadas ao estímulo. Para analisar a dinâmica das respostas de estado estacionário, pode-se supor que a evolução temporal da amplitude de resposta seja a mesma em diferentes execuções de estimulação separadas por pausas suficientemente longas. Com base nessa suposição, é apresentado um método para caracterizar a evolução temporal das respostas de estado estacionário. Um número suficientemente grande de gravações é adquirido em resposta à mesma condição experimental. As execuções experimentais (gravações) são médias em coluna (ou seja, as execuções são médias, mas a época dentro das gravações não é calculada com os segmentos anteriores). A média em coluna permite a análise de respostas de estado estacionário em gravações com proporções notavelmente elevadas de sinal-ruído. Portanto, o sinal de média fornece uma representação exata da evolução do tempo da resposta do estado estacionário, que pode ser analisada nos domínios de tempo e frequência. Neste estudo, uma descrição detalhada do método é fornecida, usando o estado estacionário visualmente evocado potenciais como um exemplo de uma resposta. As vantagens e as advertências são avaliadas com base em uma comparação com métodos de teste único projetados para analisar a arrastamento neural.
Introdução
Quando gravado a partir do couro cabeludo, atividade elétrica cerebral é observada como mudanças contínuas e regulares em tensões ao longo do tempo. Esta atividade elétrica é chamada eletroencefalograma (EEG) e foi descrita pela primeira vez por Hans Berger no final dos anos vinte do século passado1. Os estudos seminal subseqüentes descreveram o EEG como uma série composta do tempo, em que os testes padrões rítmicos ou repetitivos diferentes podem ser observados2,3,4. Atualmente, o EEG é tipicamente dividido em cinco bandas de frequências bem estabelecidas, Delta, Theta, alfa, beta e gama, que estão associadas ao processo sensorial e cognitivo diferente.
Durante anos, o estudo das oscilações cerebrais utilizando EEG foi restrito a qualquer análise do espectro na atividade em andamento ou mudanças na atividade oscilatória provocada por eventos sensoriais não periódicos. Nas últimas décadas, diferentes metodologias têm sido implementadas para a modulação das oscilações contínuas do EEG e para a exploração dos efeitos de tais modulações nos processos perceptuais e cognitivos, incluindo a apresentação da estimulação sensorial rítmica para induzindo o arrastamento neural. O termo arrastamento neural refere-se à sincronização da atividade neural com as propriedades periódicas dos estímulos sensoriais. Esse processo leva à geração de potenciais evocados de estado estacionário (i.e., oscilações do EEG bloqueadas às propriedades periódicas dos estímulos de condução). Os potenciais evocados de estado estacionário são os mais comumente elicitados pela estimulação visual, auditiva e vibrotátil, usando estímulos transitórios apresentados a uma taxa constante ou estimulação contínua modulada em amplitude na frequência de interesse. Considerando que os potenciais evocados somatosensory do estado estacionário (ssseps) são gravados em resposta à estimulação tátil repetitiva5,6, potenciais visualmente evocados de estado estacionário (ssveps) são geralmente eliciados pelo periódico apresentação de luminância Flickers, imagens e faces7,8. As respostas auditivas de estado estacionário (ASSRs) são geralmente geradas por trens de estímulos acústicos transitórios ou pela apresentação contínua de tons amplitude-modulados9,10.
A extração de potenciais evocados de estado estacionário do EEG medido baseia-se essencialmente na média dos epochs de EEG adquiridos subseqüentemente tempo-Locked ao estímulo11. Devido à periodicidade das respostas, elas podem ser analisadas nos domínios de tempo e frequência. Após a transformação de frequência-domínio, a resposta sensorial é observada como picos de amplitude na taxa de apresentação ou frequência de modulação dos estímulos externos, e seus correspondentes harmônicos. Estes procedimentos (média do tempo-domínio e a subsequente transformação do domínio de frequência) têm sido essenciais para o desenvolvimento de um teste auditivo baseado na detecção de métodos de ASSR com finalidades clínicas12,13,14 ,15,16.
Além disso, a média do tempo-domínio clássico de epochs de EEG tem sido extremamente útil para analisar processos fisiológicos tais como a geração e a extinção de ssvep17,18. Apresentando trens consecutivos de luzes de cintilação e média de épocas subseqüentes dentro de uma gravação, Wacker et al.19 observaram que o índice de bloqueio de fase do SSVEP aumentou rapidamente durante os primeiros 400 MS de estimulação e permaneceu alto depois . Igualmente relataram que o arrastamento visual robusto estêve estabelecido entre 700-1 100 ms após o início do estímulo. Um certo grau de arrastamento permaneceu efetivo após o deslocamento do trem de estimulação, que durou aproximadamente três períodos da resposta oscilatória17,18. Esses comportamentos têm sido interpretados como o efeito envolvente/desengajamento das oscilações observadas, o que é consequência do processamento de informações não lineares no sistema visual humano17. Alternativamente, sabe-se que determinadas condições experimentais, a estimulação da cintilação pode eliciar em-respostas no começo, e fora-respostas na extremidade de trens da estimulação em vez do arrastamento neural18.
A suposição principal a média consecutivamente adquiriu epochs de EEG é que o sinal de EEG representa uma combinação linear da resposta sensorial e do ruído de fundo20. Além disso, presume-se que a amplitude, a frequência e a fase da resposta oscilatória sejam estacionárias, enquanto o ruído de fundo é considerado como uma atividade aleatória. Entretanto, nos casos em que esta suposição não é satisfeita, a amplitude da resposta computada depois que diversos epochs não correspondem necessariamente à amplitude instantânea do potencial evocado.
Tem sido relatado recentemente que o ASSR gerado no tronco encefálico de ratos se adapta à apresentação contínua de tons modulados por amplitude (i.e., a amplitude de resposta diminui exponencialmente ao longo do tempo)21,22. A adaptação tem sido interpretada como um mecanismo neural que reflete a perda de novidade de um estímulo sensorial monótono repetitivo, aumentando a sensibilidade às flutuações relevantes no ambiente acústico23,24. Na via auditiva, a adaptação pode aumentar a compreensão da fala em ambientes ruidosos. Além disso, esse processo pode ser uma parte dos mecanismos existentes para monitorar o feedback auditivo da própria voz para controlar a produção da fala.
Analisando a evolução do tempo do 40 Hz ASSR em humanos, Van Eeckhoutte et al.25 observaram uma diminuição significativa mas pequena na amplitude de resposta ao longo do tempo (cerca de-0, 2 μV/s com base na análise do grupo, quando se assume uma diminuição linear ao longo do tempo). Conseqüentemente, estes autores concluíram que o 40 Hz ASSR em humanos não se adapta à estimulação. Nos seres humanos, comportamentos não estacionários têm sido observados ao analisar a estabilidade do SSVEP26. Esses autores observaram que a amplitude da frequência fundamental e a segunda harmônica do SSVEP foram estacionárias em apenas 30% e 66,7% dos sujeitos testados, respectivamente. As fases de ambos os componentes da freqüência de SSVEP, embora relativamente estáveis sobre o tempo, exibiram drifts pequenos26.
Conseqüentemente, embora a média clássica do tempo-domínio de épocas subseqüentemente adquiridas permita a exploração de propriedades estacionárias do arrastamento neural, esta metodologia precisa de ser revisada quando a dinâmica a longo prazo do arrastamento é o foco do pesquisa, ou quando a média da dinâmica de curto prazo é corrompida pela ocorrência de dinâmicas de longo prazo. Para caracterizar os comportamentos não estacionários das respostas do estado estacionário, a resposta evocada calculada em uma janela de tempo dada não deve ser comprometida por aquelas computadas nos segmentos precedentes de EEG. Em outras palavras, o potencial evocado deve ser extraído do ruído de fundo sem que o tempo-domínio seja em média com os segmentos de EEG precedentes.
Neste estudo, um método para avaliar a dinâmica da arrastamento neural é apresentado. As respostas de estado estacionário são gravadas repetitivamente em resposta à mesma estimulação, onde gravações consecutivas são intercaladas por um intervalo de repouso de três vezes o comprimento da corrida experimental. Considerando que, se a evolução temporal da resposta fisiológica for a mesma em diferentes ensaios experimentais independentes (gravações independentes), as gravações são médias em coluna. Em outras palavras, as epochs correspondentes ao mesmo local nas diferentes gravações são médias, sem a média de epochs dentro de uma gravação. Consequentemente, a amplitude de resposta calculada em qualquer intervalo de estimulação corresponderá à amplitude instantânea do potencial evocado. As respostas sensoriais podem ser analisadas no domínio temporal ou transformadas em domínio de frequência, dependendo do objetivo do experimento. Em qualquer caso, as amplitudes podem ser plotadas como uma função do tempo para analisar a evolução do tempo da resposta do estado estacionário. A geração e a extinção dos potenciais evocados de estado estacionário podem ser avaliadas restringindo a análise à primeira e última epochs das gravações.
A dinâmica do arrastamento neural pode ser analisada usando outras abordagens, como filtragem de banda estreita medições de teste único em torno da freqüência de interesse e computação do envelope do sinal de energia usando low-pass Filtragem25 e os Transformação de Hilbert27. Comparado a essas metodologias, a média em coluna de epochs permite computar parâmetros de estado estacionário com base em sinais com a maior relação sinal-ruído (SNR). Recentemente, a filtragem de Kalman surgiu como uma técnica promissora para a estimativa de 40-Hz ASSR amplitudes28,29,30. A implementação da filtragem de Kalman pode melhorar a detecção de respostas de estado estacionário mais perto do limiar eletrofisiológico e reduzir o tempo do teste auditivo29. Além disso, as respostas estacionárias não são necessárias para serem assumidas quando uma abordagem de filtragem de Kalman é usada para estimar a amplitude de ASSR30. No entanto, apenas um estudo analisou a evolução temporal dos ASSRs usando a filtragem de Kalman25. A conclusão do estudo é que a amplitude de 40-Hz ASSR é estável sobre o intervalo de estimulação. Conseqüentemente, a filtração de Kalman precisa de ser testada nas circunstâncias que o ASRR não é estacionário.
Embora demorada, o método de média de coluna é livre de modelo e não precisa de valores de inicialização e/ou a priori definições do comportamento de ruído. Além disso, desde que não envolva tempos da convergência, a média coluna-sábia pode fornecer uma respresentação mais de confiança do início do arrastamento neural. Conseqüentemente, os resultados obtidos com o método de média coluna-sábio podem ser considerados como a verdade à terra para analisar a dinâmica do arrastamento neural usando a filtração de Kalman.
Essa descrição do protocolo é baseada em um exemplo de SSVEP. No entanto, é importante notar que o método aqui apresentado é independente da modalidade, de tal forma que também pode ser usado para analisar a evolução do tempo de SSSEP e ASSR.
Protocolo
O presente estudo foi realizado aprovação do Comitê de pesquisa e ética da Universidad de Valparaíso, Chile (código de avaliação CEC170-18), confirmado pelas diretrizes nacionais de pesquisa com sujeitos humanos.
1. preparação
- Bem-vindos ao assunto.
- Explicar os objetivos e a relevância do estudo. Forneça uma descrição dos detalhes técnicos relevantes. Responda todas as perguntas completamente.
- Mencionar explicitamente que ela/ela pode interromper a sessão experimental a qualquer momento, se desejado.
- Peça ao voluntário que leia o termo de consentimento livre e esclarecido e assine o formulário correspondente. Interromper a sessão experimental se o consentimento informado não for obtido.
2. preparação do assunto
- Peça ao sujeito que se sente em uma cadeira de laboratório em uma posição confortável.
- Limpe o couro cabeludo com etanol (uma solução em 95%) para remover a camada de células mortas da pele e sebo que cobri-lo. Esta etapa é importante para reduzir a impedância entre os eletrodos e couro cabeludo.
- Meça a circunferência da cabeça com uma fita métrica para definir o tamanho da tampa do eletrodo a ser usada.
- Peça ao sujeito que use a tampa do eletrodo. Forneça as instruções para o posicionamento confortável mas correto da tampa.
- Meça a distância entre o násio (NZ, o ponto médio da sutura nasofrontal, que pode ser identificado pela depressão entre os olhos e a parte superior do nariz) e o inion (Iz, a proeminência do osso occipital) usando uma fita métrica.
- Meça a distância entre os pontos pré-auriculares esquerdo e direito (identificado como a depressão pouco antes da aurícula das orelhas) usando uma fita métrica.
- Corrija a posição da tampa do eletrodo, de modo que a interseção entre as linhas imaginárias definidas nas etapas anteriores corresponda ao vértice da cabeça. Certifique-se que o assunto está confortável após os ajustes.
- Coloque o gel condutor nos suportes do eletrodo, de acordo com os locais considerados para o experimento.
- Use 64 locais do couro cabeludo seguindo o sistema 10-20 internacional31 para usar o resultado do protocolo para executar a análise de localização de origem. O maior número de locais do eletrodo (128) no couro cabeludo pode ser usado se necessário.
- Implemente os ajustes clínicos ou ambulatórios (com somente alguns elétrodos) se a análise da localização da fonte não é planeada. Use locais occipital para gravar SSVEP, locais temporais para adquirir ASSR e locais parietal para gravar SSSEP.
- Empurre os eletrodos nos suportes dos eléctrodos. Certifique-se de que o rótulo do eletrodo coincide com o rótulo de localização na tampa.
- Acompanhe o voluntário para a sala experimental (de preferência, uma câmara blindada e atenuada pelo som). Peça ao sujeito que se sente em uma cadeira dentro da sala, em uma posição confortável.
- Coloque eletrodos externos no nariz e lóbulos da orelha se uma referência física (diferente dos eletrodos do couro cabeludo) for usada para re-referenciar a gravação do EEG (na etapa 3.8.1).
- Coloque eletrodos externos em locais perioculares.
- Coloque eletrodos na bochecha e na região frontal da cabeça, aproximadamente 1 cm acima da sobrancelha, para gravar piscando (no passo 2.6.1).
- Coloc elétrodos no canto exterior dos olhos, aproximadamente 1 cm acima de/abaixo da linha média, para gravar movimentos de olho (na etapa 2.6.1).
Nota: o electrooculograma (EOG) será usado na etapa 3.8.5 para remover os artefatos EGG induzidos por movimentos piscantes e oculares.
- Gire o sistema da aquisição de EEG sobre e verific a impedância do elétrodo se um sistema da baixo-impedância é usado gravando o EEG. Corrija a impedância, conforme necessário, de acordo com as direções do fabricante. A impedância deve ser mantida abaixo de 10 kΩ32.
- Peça ao sujeito que pisque e mova os olhos em direções diferentes para garantir que o EOG esteja sendo gravado corretamente.
- Para analisar a dinâmica do SSVEP, ajuste a localização da tela na direção vertical, para coincidir com o ângulo de visão do assunto. Escurecer as luzes do quarto até que um nível confortável seja conseguido. Ajuste o nível de luminância da tela para o limite superior do nível de conforto do participante.
- Para analisar a dinâmica do ASSR, insira os fones de ouvido usando as pastilhas de espuma corretas, para que os fones de ouvido caiam no canal auditivo. Verifique se os sons são entregues na intensidade desejada (por exemplo, um nível de conforto psicofísico33).
3. aquisição e pré-processamento de EEG
- Definir os parâmetros de estímulo definidos no experimento experimental. Consulte o manual do usuário fornecido pelo fabricante do sistema de estimulação para obter detalhes sobre o software.
Nota: para explicações abrangentes sobre o estímulo utilizado para a geração de SSVEP e ASSR, ver Norcia et al.8 e Rance34, respectivamente. - Instrua o assunto a prestar atenção à estimulação, no caso em que a arrastamento Visual é o tema do experimento.
- Apresente um filme legendado com o som desligado quando a arrastamento auditivo é o tema do experimento.
Nota: a apresentação de um filme silencioso permite a deflexão da atenção da estimulação acústica, mantendo o nível de excitação25.
- Apresente um filme legendado com o som desligado quando a arrastamento auditivo é o tema do experimento.
- Apresentam estímulos com mais de 90 s, como foi feito para investigar a evolução do tempo de ssveps e ASSR em humanos e modelos animais21,22,25,26.
Nota: apresentar estímulos mais curtos na duração se um estudo piloto tiver sido realizado. - Pause a estimulação por 2 min se apenas uma condição experimental estiver sendo testada. Interagir com o assunto para verificar a conscientização.
Nota: a duração da pausa depende da duração da estimulação. Pausa 3x mais do que os intervalos de estimulação irá garantir que uma resposta eliciada por um estímulo não é afetada pela estimulação anterior. Pausas mais longas são permitidas se o assunto assim o solicitar.- Pausar a estimulação por pelo menos 10 s quando diferentes condições experimentais são testadas desde a estimulação alternada com pausas de 10 s tem sido proposto para diminuir os efeitos de adaptação extra e reduzir o comprimento do experimento25.
- Repita as etapas da apresentação (etapas 3.3-3.4) pelo menos 30x para assegurar o SNR elevado das medidas após a média dos epochs (etapa 4,4).
- Registre o EEG usando procedimentos padrão35. Crie um arquivo EEG separado para cada execução experimental.
Nota: consulte o manual do usuário do sistema de aquisição para obter detalhes sobre o software. - Monitore a gravação de EEG para detectar períodos de sono baseados no nível de atividade alfa e na freqüência em que os artefatos piscantes aparecem. Pause o experimento quando níveis aumentados de alfa acompanhados por frequências intermitentes reduzidas são detectados, o que é indicativo de sonolência. Rejeite a execução experimental de uma análise mais aprofundada quando os períodos de sono são detectados.
- Calcule a amplitude da resposta de estado estacionário no final de cada execução experimental, seguindo as instruções fornecidas no manual do usuário do software de aquisição usado no experimento.
- Monitore o nível de atenção do sujeito comparando a amplitude das respostas de estado estacionário obtidas no final de cada ensaio experimental. Definir a amplitude do estado estacionário obtida nas primeiras execuções experimentais como uma amplitude de referência.
- Definir um limiar de rejeição (uma diminuição na amplitude de resposta de 5% em relação à amplitude de referência). Rejeite as execuções experimentais em que a amplitude da resposta do estado estacionário atende ao critério de rejeição.
- Termine a sessão experimental depois de adquirir o número de execuções definidas no experimento experimental.
- Pré-processe os dados de EEG off-line usando procedimentos padrão de EEG35 descritos nas próximas etapas por direções do fabricante.
- Rereferenciar a gravação usando uma referência média (média de todos os eletrodos de gravação) ou a média de um subconjunto de eletrodos. Alternativamente, use uma referência física (por exemplo, eletrodos externos colocados no nariz e lóbulos de orelha descritos na etapa 2,4).
- Converta as coordenadas do eletrodo para o sistema 10/20 internacional se o sistema de coordenadas radial foi usado durante as aquisições de EEG. Consulte o manual do fabricante para obter detalhes sobre a conversão.
- Band-pass filtre o sinal de EEG entre 0.5-300 hertz. Defina um filtro de entalhe (centralizado em 50 Hz ou 60 Hz), se necessário.
- Para baixo-prove o sinal de EEG para diminuir o tempo de execução do algoritmo selecionado removendo artefatos da ocular (etapa 3.8.5).
Nota: uma frequência de amostragem de 512 Hz é adequada para analisar oscilações cerebrais de frequência abaixo de 40 Hz35. - Remova os artefatos oculares.
Nota: para este fim, diferentes técnicas podem ser usadas (ver Urigüen e Garcia-Zapirain35 para uma extensa revisão sobre algoritmos de remoção de artefatos). Entre eles, a análise de componentes independentes é uma das metodologias mais estendidas e é implementada tanto em softwares comerciais como em análise gratuita37,38,39. - Segmente os dados de EEG nos epochs tempo-fechados à estimulação. Selecione o comprimento de epochs de acordo com o objetivo do experimento.
Nota: as epochs devem ser suficientemente longas para permitir a análise da resposta do estado estacionário no domínio de frequência com uma resolução espectral adequada. - Não execute algoritmos de rejeição de artefato neste estágio para detectar e remover epochs contendo artefatos.
Nota: a remoção de Epoch nesta fase induz erros quando o conjunto de dados é organizado para executar a média de coluna de epochs (passos 4,2 e 4,4). Os algoritmos de rejeição são implementados em uma etapa de processamento posterior (etapa 4.1.4). - Execute a função DC-Detrend para calcular DC-Trends em epochs de EEG individuais e corrigi-los.
- Execute a função de correção de linha de base para corrigir a linha de base da gravação. Selecione intervalos de tempo pré-estímulo maiores que 200 ms.
Observação: a correção de linha de base consiste em calcular a média dos dados no intervalo de tempo selecionado. A média é calculada para cada canal e subtraída de cada ponto de dados em cada época.
4. cálculo das amplitudes de resposta
- Entrar os parâmetros necessários para o cálculo das respostas de estado estacionário (Figura 1a).
Nota: o código em casa utilizado para o processamento dos dados está disponível gratuitamente em < https://figshare.com/projects/Steady-state_visually_evoked_potentials_SSVEP_elicited_in_humans_by_continuos_light_modulated_in_amplitude_at_10Hz/62573 >. Consulte o texto de ajuda dentro do código para obter mais instruções. Da mesma forma, um subconjunto dos dados utilizados neste estudo está disponível.- Insira o número de gravações (execuções experimentais) do experimento.
- Insira o comprimento dos epochs para segmentar as gravações individuais.
- Insira a frequência de amostragem do experimento.
- Selecione algoritmos de rejeição de artefato para detectar e remover epochs contendo artefatos. Os critérios de seleção disponíveis são 1) gradiente (diferença absoluta entre duas amostras consecutivas), 2) Max-min (a diferença entre a amplitude máxima e mínima na época) e 3) amplitude (amplitudes máximas e mínimas absolutas).
- Execute o código de processamento.
Nota: as etapas 4.2-4.7 são executadas automaticamente quando esta opção é selecionada. Execute as etapas manualmente se apropriado.
- Reorganize as epochs em uma matriz de dados de n linhas e colunas m , nas quais n representa o número de gravações (ensaios experimentais) e m o número de epochs (Figura 1b).
- Peso da época para atenuar o efeito do movimento e artefatos musculares.
Nota: os epochs ponderados de EEG são obtidos dividindo cada amostra da tensão pela variação da amplitude do Epoch que pertencem a, de modo que a variância seja usada como uma medida da variabilidade da amplitude e do fator de ponderação40. - Coluna-Wise Average o conjunto de dados. Para este fim, a média do tempo-domínio os epochs que correspondem à mesma janela de tempo nas gravações diferentes.
Nota: esta etapa permite a computação da amplitude do estado estacionário nas gravações com uma relação sinal-ruído notavelmente elevada (SNR). - Exporte a série temporal resultante da média para uma análise mais aprofundada da evolução temporal do arrastamento em software externo.
- Calcule a amplitude da resposta de estado estacionário em cada época resultante da média de coluna, usando a transformada rápida de Fourier (FFT).
Nota: o comprimento de FFT deve corresponder ao comprimento de um epoch. A implementação de uma técnica de Windowing não é obrigatória. A amplitude da resposta do estado estacionário é definida como a amplitude espectral obtida na frequência da modulação de amplitude dos estímulos sensoriais.
- Calcule a amplitude da resposta de estado estacionário em cada época resultante da média de coluna, usando a transformada rápida de Fourier (FFT).
- Vector a amplitude de um número ad-hoc de compartimentos FFT em cada lado da frequência da resposta para calcular o nível de ruído residual (RNL). O número de escaninhos de FFT deve corresponder com uma faixa de freqüência de aproximadamente 3 hertz, em cada lado da freqüência da resposta.
Nota: a alta frequência-especificidade das respostas do estado estacionário torna a amplitude de resposta independente daquelas oscilações de fundo com frequências semelhantes, que por sua vez distribui uniformemente em uma banda de frequência relativamente estreita41 , 42 , 43. - Plotar a amplitude da resposta do estado estacionário e o RNL como uma função do índice de coluna (ou seja, o número da época adquirida) para explorar a evolução da resposta do estado estacionário durante o intervalo de estimulação.
Resultados
O SSVEP foi eliciado por estímulos visuais contínuos de 40 s de comprimento, onde a intensidade da luz foi modulada por uma onda sinusoidal de 10 Hz (profundidade de modulação de 90%). Os estímulos foram entregues por quatro diodos emissores de luz (LEDs) situados no centro de uma tela preta de 50 cm x 50 cm, como vértices de um quadrado de 5 cm x 5 cm. Quando o participante sentou-se 70 cm da tela, a área do quadrado de LEDs subtende um ângulo visual de cerca de 4 °. A tela LED foi projetada usando um sistema de desenvolvimento microcontrolador baseado em USB e quatro LEDs brancos super brilhantes de 10 mm de diâmetro. A técnica de modulação por largura de pulso (PWM) foi utilizada para controlar a potência fornecida aos LEDs. Esta técnica controlou as intensidades dos diodos emissores de luz em uma freqüência dada e gera o envelope sinusoidal final. Uma freqüência de PWM de 40 kHz foi usada para evitar um efeito de cintilação perceptível.
Foram obtidas trinta gravações, segmentadas em epochs de 4 s. Portanto, foi obtido um conjunto de dados composto por 10 colunas (número de epochs de EEG dentro de gravações) e 30 linhas (número de gravações, número de ensaios experimentais).
O tempo de oscilação neural bloqueado para a estimulação tornou-se evidente à medida que a média de coluna foi realizada (Figura 2). Significativamente, o intervalo no qual o SSVEP é gerado pode ser observado em traços correspondentes à coluna 1. Nessa coluna, 0,2 s de linha de base pré-estímulo são plotados, além dos primeiros 0,8 s de arrastamento neural. Conseqüentemente, o procedimento descrito aqui permite a caracterização de 1) a dinâmica da resposta oscilatória uma vez que o arrastamento neural é estabelecido já e 2) o acoplamento de oscilações neural. Uma ou mais epochs registradas após o término da estimulação também podem ser incluídas na matriz de dados para estudar a extinção da resposta do estado estacionário após o deslocamento do estímulo.
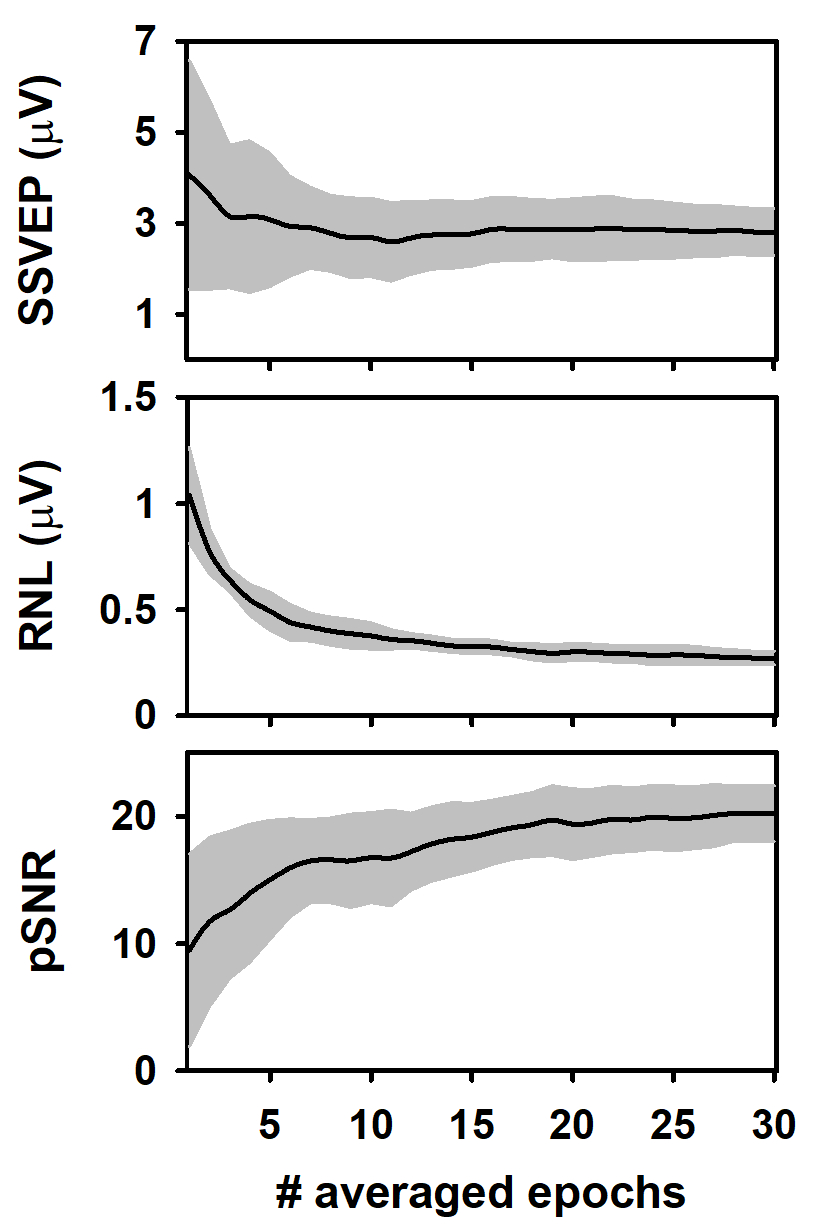
Durante a média da coluna de epochs, a amplitude média do SSVEP (amplitude espectral a 10 Hz, calculada pela aplicação do FFT) diminuiu durante a média das primeiras epochs das colunas e tendeu a estabilizar posteriormente (Figura 3a). Este resultado concorda com estudos prévios analisando a evolução do ASSR durante a média das epochs sequencialmente adquiridas21,22,40,43,44. O comportamento da amplitude da resposta durante a média é explicado geralmente pela contribuição relativamente elevada do ruído unaveraged à amplitude da resposta computada nos primeiros epochs, que é atenuado como a média é executado13, 44 , 45 , 46 , 47. notável, a variabilidade da amplitude do SSVEP diminuiu significativamente à medida que a média progrediu.
Também analisamos a RNL das medições durante a média de epochs em coluna (Figura 3B). O RNL foi computado em uma faixa de freqüência estreita (3 hertz) em ambos os lados da freqüência do SSVEP. Embora este procedimento não é comum quando ssvep são analisados, vector-média de um determinado número de caixas de freqüência em torno do que o arrastamento neural é o padrão para estimar o RNL em medidas de ASSR41,42, 43. como esperado, o RNL diminuiu progressivamente à medida que o número de épocas médias aumentou e atingiu o nível assíptico após cerca de 20 epochs foram processados. Diferentemente do observado quando a amplitude do SSVEP foi analisada, o desvio padrão da RNL permaneceu relativamente constante à medida que o número de épocas médias aumentava, sugerindo que as condições de registro eram estáveis ao longo da sessão experimental.
Os resultados apresentados acima determinaram as alterações na relação sinal-ruído de pico (pSNR) das medições durante a média de época da coluna (Figura 3C). Este termo é definido aqui como a relação (no dB) entre a amplitude quadrada da resposta (SSVEP) e a amplitude quadrada do RNL. Como a média progrediu, o pSNR aumentou enquanto o número de épocas médias aumentou até 18, aproximadamente. Uns aumentos mais adicionais no número de épocas médias não impactam significativamente a qualidade do sinal. A variabilidade do pSNR diminuiu à medida que mais epochs foram calculados.
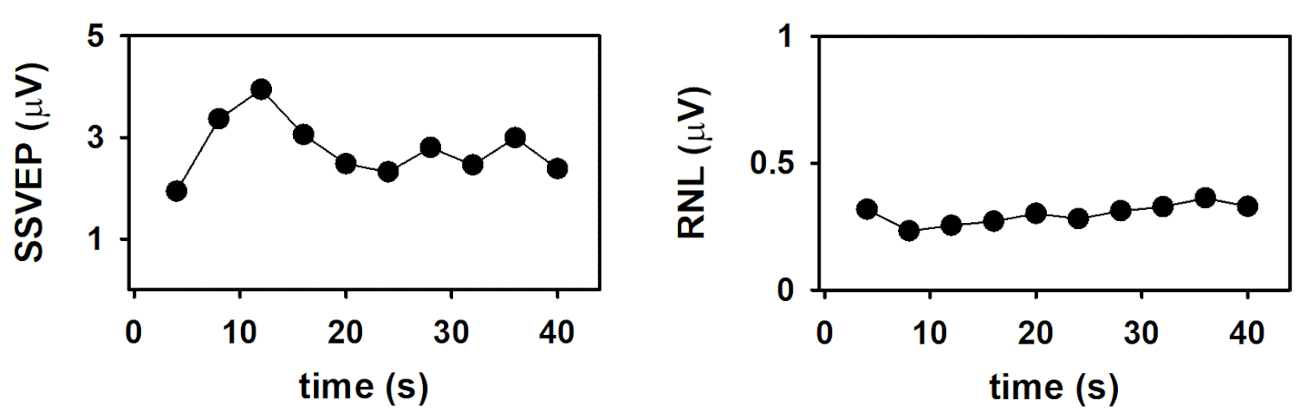
Finalmente, a dinâmica da amplitude do SSVEP e da RNL estão representadas na Figura 4. Essas evoluções de tempo foram obtidas por meio da plotagem dos parâmetros de resposta computados no final da média em coluna de epochs em função do número de colunas (em função do tempo). Como demonstrado por Labecki et al.26, a dinâmica do SSVEP pode variar significativamente entre os sujeitos. Como os resultados apresentados na Figura 4 correspondem a um único indivíduo, não é possível fazer generalizações. Neste assunto, a amplitude do SSVEP exibiu um comportamento relativamente complexo (figura 4a). A amplitude da resposta aumentou gradualmente durante os primeiros 12 segundos que seguem o início do estímulo (tempo que corresponde ao comprimento de 3 epochs). Enquanto o estímulo persistiu, o SSVEP diminuiu consistentemente durante os seguintes 12 segundos, e permaneceu relativamente constante mais tarde. Esses resultados não podem ser explicados pelo comportamento do RNL, uma vez que esse parâmetro foi relativamente constante durante o intervalo de estimulação (Figura 4B). O aumento da amplitude do SSVEP após o início do estímulo é evidente nos traços apresentados na Figura 2 e pode ser explicado pelos processos de integração, que resultam na estabilização do arrastamento neural. A diminuição subseqüente na amplitude sugere a adaptação de SSVEP à estimulação sustentada. No entanto, essas hipóteses precisam ser testadas em experimentos controlados com tamanhos de amostra apropriados.

Figura 1 : Etapas críticas para extrair a evolução do tempo da amplitude de respostas de estado estacionário. (A) captura de tela do código de processamento, onde os parâmetros de análise são definidos. (B) diagrama representativo ilustrando a organização do conjunto de dados. Uma matriz de dados composta por 30 gravações de 10 epochs é representada. A média em coluna de epochs é destacada na primeira coluna. A linha vertical representa a direção da média. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2 : Mudanças na forma de onda de potenciais evocados visualmente de estado estacionário (SSVEP) durante a média em coluna de epochs. As respostas foram eliciadas pela apresentação contínua da luz modulada em amplitude a 10 Hz. As linhas mostram as formas de onda obtidas após a média de todas as gravações anteriores (ou seja, a linha 1 é a primeira gravação, a linha 5 é a forma de onda obtida após a média das primeiras cinco gravações, e a última linha é a média de todas as gravações). As formas de onda mais confiáveis de SSVEP foram observadas em cada coluna, pois o número de médias de execuções aumentou. Para fornecer clareza (para tornar visíveis as oscilações do SSVEP), apenas o primeiro segundo dos epochs é representado. As exceções são rastreamentos na primeira coluna do conjunto de dados, para os quais 0,2 segundos de linha de base pré-estímulo são exibidos. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3 : Alterações nos parâmetros de resposta e gravação durante a média em coluna de epochs. (A) evolução da amplitude do SSVEP. (B) comportamento do RNL. (C) alterações no PSNR. As linhas pretas representam os valores médios obtidos para cada coluna (n = 10) e a sombra cinzenta representa a área coberta por ± um desvio padrão. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4 : Evolução do tempo do SSVEP eliciado pela apresentação da estimulação visual contínua, modulada em amplitude a 10 Hz. (A) tempo de curso da amplitude do SSVEP. (B) curso de tempo do RNL. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
Este trabalho descreve um procedimento experimental para analisar a dinâmica das respostas cerebrais oscilatórias. Tal metodologia consiste em adquirir um número suficiente de ensaios experimentais independentes da mesma condição experimental, e os tempos-domínio média epochs correspondente ao mesmo tempo janela nas diferentes gravações (colunas-sábio média em Figura 1B). A amplitude computada nos dados da média representa a amplitude instantânea da resposta oscilatória. Plotar essas amplitudes como uma função do tempo (ou o número de colunas no conjunto de dados) permite analisar a evolução do tempo do tempo de resposta oscilatória-bloqueado para a estimulação. Esta metodologia é uma modificação do proposto por Ritter et al.23 para a análise da adaptação de potenciais evocados corticais transitórios. O método tem sido utilizado para analisar a dinâmica dos potenciais evocados auditivos em ambos os seres humanos24 e modelos animais20,21.
Do ponto de vista metodológico, a combinação de parâmetros utilizados para provocar a resposta do estado estacionário e aquelas implementadas para extrair a resposta neural do ruído de fundo é fundamental para analisar a evolução temporal dos potenciais evocados de estado estacionário 22. o comprimento do estímulo utilizado no experimento aqui apresentado (40 s) foi selecionado com base nos resultados obtidos em um estudo piloto. Esse comprimento de estímulo foi suficiente para analisar a adaptação de ASSR gerada no tronco encefálico21,22. Além disso, o comprimento do estímulo deve exceder o tempo em que o poder de banda instantânea assíptica de SSVEPs é atingido (Figura 1 em Labecki et al.26). No entanto, o poder da faixa instantânea assíptica de SSVEPs pode ser alcançado além dos anos 60 em alguns casos (Figura 2 em Labecki et al.26). Portanto, a execução de um estudo piloto de pequena amostra é recomendada para definir o comprimento do estímulo da estimulação. Caso contrário, um comprimento de estímulo maior do que 90 s é recomendado para alcançar a representação completa da evolução do tempo da resposta. O uso de pausas longas adequadamente entre gravações consecutivas implica considerar execuções experimentais consecutivas como sendo estatisticamente independentes (ou seja, medidas diferentes e independentes da mesma variável). Ao melhor de nosso conhecimento, nenhuma experimentação foi executada para analisar a pausa a melhor entre funcionamentos (a pausa mínima exigida para fazer funcionamentos independentes de se). O critério de usar pausas pelo menos 3x mais longo do que o comprimento do estímulo é conservador bastante para assegurar-se de que a resposta do steady-state gravada em todo o funcionamento dado não seja afetada pela estimulação precedente.
Recentemente, estímulos alternados (condições experimentais) têm sido propostos como uma escolha para reduzir a pausa entre ensaios experimentais, evitando o efeito de adaptação extra25. Da mesma forma, o número de ensaios experimentais (30) implementados neste protocolo experimental é conservador, uma vez que o RNL e o pSNR assíticos são tipicamente alcançados após uma média de 20 ensaios experimentais, aproximadamente. Quando os estímulos caem dentro da região meio-superior do intervalo dinâmico da resposta (níveis de sensação alta), um menor número de execuções provavelmente é necessário para analisar a dinâmica da resposta evocada. No entanto, nos casos em que diferentes condições experimentais são testadas, ter o mesmo número de ensaios experimentais é crucial para fazer comparações entre as condições (ou seja, diferentes níveis de sensação).
Além do que a média coluna-sábia dos epochs, a dinâmica de potenciais evocados oscilatórios foi analisada filtrando as medidas do único-teste em uma faixa de freqüência estreita em torno da freqüência do interesse e computando o envelope do poder sinal usando filtragem passa-baixa26. Da mesma forma, a análise experimental única foi implementada para caracterizar o período de transição que precede a região estável do SSVEP48, e as alterações na amplitude e na fase do ssvep durante a região estável da resposta49. Embora as análises de avaliação única permitam a discriminação de flutuações relativamente rápidas na amplitude de resposta, os designs experimentais para analisar a resposta média em blocos separados por um determinado intervalo de interblocos só respondem por variações de longo prazo no amplitude do potencial evocado50,51. A média de coluna-Wise de epochs está entre estas duas opções. Convertendo o sinal de média para o domínio de frequência usando o FFT implica analisar a dinâmica da resposta com uma resolução igual ao comprimento da época. No exemplo apresentado aqui, o SSVEP foi relatado cada 4 s. Embora 4 s de resolução seja adequada para descrever a dinâmica que ocorre em intervalos de tempo superando dezenas de segundos, como a do SSVEP26, as epochs parcialmente sobrepostas nas gravações originais permitem descrever a evolução do tempo do resposta de estado estacionário de forma mais refinada25.
A dinâmica das respostas do estado estacionário obtida após a média em coluna de epochs representa principalmente a evolução da atividade oscilatória que é sincronizada entre os segmentos médios de EEG (aqueles que sobrevivem à média). Portanto, uma questão importante sobre a viabilidade da metodologia é a possível atenuação das amplitudes de resposta devido a variações na fase de oscilações neurais de uma corrida experimental independente para outra (ou seja, entre as gravações). Este tópico precisa ser abordado experimentalmente. No entanto, evidências indicam que a fase de respostas oscilatórias cerebrais é menos variável do que o esperado. De fato, vários estudos relataram uma regularidade na fase esperada do ser humano 80 Hz ASSR47,48,49. Quando as latências são estimadas com base na fase da atividade oscilatória, observou-se o efeito previsível da intensidade e da frequência portadora dos estímulos acústicos sobre a latência das respostas auditivas (i.e., a diminuição da latência como a intensidade e aumento de frequência do portador)52,53,54. Além disso, alterações maturacionais típicas na amplitude e na assimetria da esquerda para a direita nos níveis auditivos também foram observadas quando as latências são estimadas a partir da fase do ASSR47,55,56 , 57 , 58. ao descrever a evolução do tempo do SSVEP utilizando uma análise de teste único, Labecki et al.26 observaram que, embora a variabilidade interexperimental das amplitudes de resposta dentro do mesmo assunto fosse consideravelmente alta, a variabilidade da fase foi significativamente menos pronunciada.
Com base em suas observações, Labecki et al.26 sugeriram que um mínimo de 50 ensaios deve ser em média para obter uma estimativa confiável do envelope de potência média da resposta. Esses resultados indicam que, mesmo quando a amplitude da resposta é computada em ensaios únicos, a média (de envelopes nesse caso) é necessária para relatar resultados confiáveis. Além disso, a variabilidade interexperimental na amplitude do SSVEP relatada por Labecki et al.26 sugere que o cálculo desse parâmetro em ensaios individuais pode ser altamente influenciado pelo ruído de fundo. Considerando a evolução da relação sinal-ruído apresentada na Figura 2, o cálculo da resposta no sinal de média em vez de ensaios simples reduz significativamente o número de segmentos de EEG necessários para serem processados para obter Medidas. Adicionalmente, a baixa variabilidade na fase obtida por Labecki et al.26 sustenta a ideia de que a média em coluna de epochs aqui apresentada é um procedimento válido para computar a dinâmica dos potenciais evocados oscillatórios.
A média dos dados em diferentes níveis leva a uma interpretação diferente dos resultados. Em relação aos potenciais evocados oscilatórios, computando a amplitude de resposta após a média do tempo de execução de execuções independentes implica analisar apenas oscilações de tempo bloqueado (ou seja, aquelas que sobrevivem à média). Este procedimento pode filtrar informações relevantes sobre a dinâmica da resposta em ensaios individuais. Entretanto, garantias uma relação sinal-à-ruído suficientemente elevada das medidas. Esse aspecto pode ser de significância quando as respostas estão próximas ao limiar eletrofisiológico, condição na qual a detecção do arrastamento pode ser comprometida devido à baixa relação sinal-ruído da medida.
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Os autores reconhecem graciosamente Lucía Zepeda, Grace A. Whitaker, e Nicolas Nieto por suas contribuições para a produção de vídeo. Este trabalho foi apoiado em parte pelos programas CONICYT BASAL FB0008, MEC 80170124 e bolsista de doutorado 21171741, bem como o Instituto Nacional de surdez e outros distúrbios da comunicação dos institutos nacionais de saúde o número de premiação P50DC015446. O conteúdo é unicamente da responsabilidade dos autores e não representa necessariamente os pontos de vista oficiais dos institutos nacionais de saúde.
Materiais
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
Referências
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G., Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. . An introduction to the event-related potential technique. Second Edition. , 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J., Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados