
Method Article
Organoides epiteliais mucociliais de células embrionárias xenopus: geração, cultura e imagem ao vivo de alta resolução
Neste Artigo
Resumo
Descrevemos um protocolo simples para desenvolver organoides epiteliais mucociliais a partir de células ectoderm profundas isoladas dos embriões Xenopus laevis. Os progenitores multipotentes regeneram precursores de células de cálice epitelial e permitem o acompanhamento ao vivo da iniciação e progressão das transições celulares na superfície dos organoides.
Resumo
O epitélio mucociliary fornece a primeira linha de defesa removendo partículas estranhas através da ação da produção de muco e liberação mediada por cílios. Muitos defeitos clinicamente relevantes no epitélio mucociliary são inferidos à medida que ocorrem profundamente dentro do corpo. Aqui, introduzimos um modelo 3D tratável para epitélio mucociliário gerado a partir de progenitores multipotentes que foram microsurgicamente isolados dos embriões Xenopus laevis. Os organoides epiteliais mucociliais são cobertos com epitélio recém-gerado a partir de células ectoderme profundas e posteriormente decorados com distintas células multiciliadas padronizadas, células secretas e células de cálice produtoras de muco que são indistinguíveis da epiderme nativa dentro de 24 horas. As sequências completas de transições celulares dinâmicas de mesenquimal para epitelial que emergem na superfície apical dos organoides podem ser rastreadas por imagens ao vivo de alta resolução. Esses organoides epiteliais mucociliais in vitro, auto-organizados, oferecem vantagens distintas no estudo da biologia do epitélio mucocilial com alta eficiência na geração, condições de cultura definidas, controle sobre número e tamanho e acesso direto à imagem ao vivo durante a regeneração do epitélio diferenciado.
Introdução
Lesões, infecções e doenças do epitélio mucociliário estão associadas à produção prejudicada e ao despejo de muco, que muitas vezes é encontrado em doenças pulmonares pulmonares crônicas, asma, fibrose cística, bronquiectase e diskinesia ciliar primária1,2,3,4. Um recente avanço na tecnologia organoide, por exemplo, o organoide pulmonar derivado de células basais chamado traqueosfera que recapitula a regeneração do epitélio mucociliary surge como um modelo promissor com potencial terapêutico1,5,6. No entanto, seu uso é atualmente limitado, em parte devido à falta das condições de cultura definidas e à baixa eficiência nas produções organoides. Epitélio mucociliary nas vias aéreas humanas e epiderme de sapo são notavelmente semelhantes em morfologia tecidual, composição celular, e sua função7,8,9,10,11,12. Em ambos os organismos, o epitélio mucociliary fornece defesa de primeira linha, secretando substâncias mucosas e antimicrobianas e limpa partículas e patógenos nocivos através da ação sincronizada de cílios.
Aqui, descrevemos um protocolo simples para gerar organoides epiteliais mucociliais usando os progenitores multipotentes dos embriões Xenopus laevis 13,14. Anteriormente, relatamos14 que, na ausência de fatores de crescimento exógenos e da matriz extracelular, as células profundas microsurgicamente isoladas do estágio gastrula precoce se reúnem espontaneamente em agregados, regeneram o epitélio em sua superfície e amadurecem em epitélio mucociliário, intercalando células multicilitárias e outras células acessórios dentro de 24h. Além do rápido desenvolvimento, este protocolo oferece uma oportunidade distinta para acessar diretamente as transições de células ectoderme profunda multipotentes em progenitores de células de cálice epitelial que recapitulam as etapas de regeneração de um epitélio14 interrompido que não estão disponíveis a partir de embriões intactos e ectoderme (também conhecido como cape animal)15,16,17. O número e o tamanho dos organoides produzidos são escaláveis com alta eficiência, controlando os materiais iniciais dos embriões Xenopus. Organoides na cultura flutuante podem ser facilmente classificados e transferidos na fase desejada para análises posteriores, incluindo imagens de alta resolução, testes mecânicos, tratamento medicamentoso e caracterização genética14. Esta regeneração espontânea e baseada em mecânica tecidual do epitélio na superfície dos agregados de células embrionárias resulta em organoides epiteliais mucociliais e fornecem um novo modelo tridimensional (3D) para estudar a biologia do epitélio mucociliário.
Protocolo
Os protocolos experimentais de uso e uso animal foram aprovados pelo Comitê Institucional de Cuidados e Uso de Animais (IACUC) do Instituto de Ciência Básica (IBS 18-01) e do Instituto Avançado de Ciência e Tecnologia da Coreia (KA2017-22).
1. Embriões
- Obter embriões X. laevis usando um procedimento padrão: coletar manualmente ovos de sapos fêmeas estimuladas e realizar fertilização in vitro18,19.
- Desgarrar os embriões fertilizados com agitação suave por cerca de 5 min em 2% de cisteína em 1/3x modificado salina de Barth (MBS; veja a receita de 1X MBS abaixo) em pH 819.
- Passo opcional para imagens ao vivo: rotular fluorescentemente proteínas específicas e observar sua dinâmica no organoide, proceder à seção 5.
- (Opcional) Para monitorar a contaminação de células de ectoderme superficiais dentro dos organoides, rotule a superfície apical dos embriões com NHS-rhodamina no estágio 914. Incubar embriões em 1 mg/mL NHS-Rhodamine em 1/3x MBS (pH 9.0) por 30 min com porca suave. Lave os embriões três vezes transferindo-se para uma placa de Petri cheia de 1/3x MBS por 15 min.
- Cultura o embrião em 1/3x MBS na temperatura preferida (14-26 °C) até que os primeiros sinais do estágio 10 sejam detectados (ou seja, o aparecimento de células pigmentadas escuras ao redor do blastopore na visão vegetal).
2. Preparação de ferramentas microcirúrgicas, soluções e vasos culturais
- Prepare as ferramentas necessárias, incluindo um par de fórceps de grau cirúrgico e ferramentas de cabelo (laço de cabelo e faca de cabelo) para microcirurgia20.
- Prepare as seguintes mídias culturais para embriões: 1/3X MBS, onde 1X MBS é feito com NaCl (88 mM), KCl (1 mM), NaHCO3 (2,4 mM), MgSO4 (0,82 mM), Ca(NO3)2 (0,33 mM), CaCl2 (0,41 mM) e HEPES (10 mM). Ajuste o pH para 7,4 com 10 M NaOH.
NOTA: Opcional: adicione gotas de fenol vermelho para indicar pH. - Prepare as seguintes mídias culturais para tecidos embrionários e organoides: Danilchik's for Amy (DFA)21 complementado com antibiótico fresco de 1% e solução antimíctica. Prepare DFA com NaCl (53 mM), Na2CO3 (5 mM), gluconato de potássio (4,5 mM), gluconato de sódio (32 mM), CaCl2 (1 mM) e MgSO4 (1 mM). Ajuste o pH para 8.3 com bicine granular. Filtrar DFA (filtro de 0,2 μm de tampa de garrafa), aliquot e armazená-lo a -20 °C.
- Prepare o DFA sem cálcio e magnésio para separar células profundas do ectoderme usando a receita acima e omitindo CaCl2 e MgSO4. Aliquot e armazenar a -20 °C.
- Prepare tubos PCR não adesivos para agregação de células embrionárias.
- Para induzir a agregação espontânea das células embrionárias isoladas, prepare tubos PCR não adesivos, revestindo tubos PCR de fundo redondo com 200 μL de 1% BSA (1 g de BSA em 100 mL de água destilada) durante a noite a 4 °C ou por 2h à temperatura ambiente. Cada tubo será usado para montar um organoide.
- Enxágüe os tubos PCR revestidos de BSA com DFA três vezes para remover qualquer BSA residual.
- Encha tubos PCR com 200 μL de DFA.
3. Isolamento de células de ectoderme profundo
- Selecione e reúna embriões à medida que chegam ao estágio inicial 10 usando ferramentas capilares sob um estereoscópio.
- Usando uma pipeta de transferência descartável, transfira os embriões selecionados para uma placa de Petri preenchida com DFA.
- Remova a membrana vitelline dos embriões usando fórceps afiados do lado vegetal sem interromper o lado animal do embrião.
NOTA: Tenha cuidado para evitar expor embriões ao ar. Introduzir bolhas de ar na solução ou trazer embriões à superfície fará com que o embrião exploda. - Para isolar a tampa animal, posicione o lado animal do embrião para cima.
- Estimar visualmente a extensão da tampa animal a ser extirpada e fazer a primeira incisão ao longo da borda da tampa animal com uma faca de cabelo. Puxe a faca de cabelo para fora para fazer um corte.
- Repita o passo 3.5 para criar uma cadeia de pequenos cortes para extirpar a tampa animal.
- Apare a borda grossa da tampa animal usando uma faca de cabelo para evitar a inclusão de precursores de mesoderm.
NOTA: Para evitar a cicatrização e agregação de tampas animais isoladas, proceda ao próximo passo dentro de 10 minutos. Normalmente, isolamos de 5 a 10 tampinhas de animais por vez para montar vários organoides. - Para separar as células de ectoderme profundo da tampa animal, transfira as tampas de animais excisadas para uma placa de Petri cheia de DFA sem cálcio e magnésio com uma pipeta de transferência descartável. Tenha cuidado para não introduzir bolhas de ar durante a transferência.
- Para manter espaço suficiente para os próximos passos, usando as ferramentas de cabelo, posicione as tampas dos animais para enfrentar o lado animal para cima e mantenha uma distância generosa de outras explantas.
- Aguarde de 5 a 10 minutos e monitore as explants sob um estereoscópio. Uma vez que células profundas soltas tenham sido encontradas a partir da borda da camada superficial pigmentada escura, comece a levantar a camada superficial para longe das células de ectoderme profundas de cor clara usando uma faca de cabelo sob o estereoscópio.
- Desprender cuidadosamente (peel-off) a camada superficial com uma faca de cabelo, começando na borda.
- Colete as células de ectoderme profundas com aspiração mínima (10\u201215 μL) para limitar a quantidade de DFA livre de cálcio e magnésio que é transferida para a mídia de agregação na próxima etapa.
NOTA: Células superficiais separadas podem ser removidas da mídia para evitar contaminar as células de ectoderme profundas restantes.
4. Geração de organoides epiteliais mucociliais
- Transfira células de ectoderme profundas coletadas para um tubo PCR não adesivo contendo 200 μL de DFA. Pipeta suavemente a mídia (2\u20123 vezes) para dispersar células transferidas no tubo PCR.
NOTA: O tempo designado como hora de agregação pós(hpa) começa nesta etapa. O tamanho dos organoides resultantes é controlado pelo número de células de ectoderme profundas adicionadas a um tubo PCR. Podem ser utilizadas células de ectoderme profundas de uma ou mais tampas de animais, dependendo do tamanho desejado do organoide. - Feche o tubo PCR. Mantenha os tubos PCR eretos para induzir a agregação espontânea na parte inferior.
- Monitore o processo de agregação sob um estereoscópio. As células normalmente se reúnem na parte inferior do tubo PCR dentro de uma hora e se reúnem em agregados esféricos dentro de 2-3 h, dependendo do tamanho.
- Para realizar imagens vivas ou testes de drogas durante o desenvolvimento dos organoides epiteliais mucociliary, colete agregados a 2 hpa usando uma pipeta de 200 μL equipada com pontas ampliadas (cortadas com tesoura esterilizada) para evitar causar danos aos agregados durante a coleta.
- Para permitir que os agregados se desenvolvam no organoide epitelial mucociliary na cultura, colete agregados esféricos do tubo PCR a 5 hpa e transfira-os para uma placa de Petri preenchida com DFA.
- Posicione os agregados longe dos outros para evitar que se fundam. Dentro de 24h de cultura à temperatura ambiente sem quaisquer fatores adicionados, organoides epiteliais mucociliais maduros podem ser observados como rotativos com a ação de bater cílios cobrindo a superfície do epitélio diferenciado
5. (Opcional) Imagem ao vivo de alta resolução de organoides em desenvolvimento
- Prepare mRNA para microinjeção.
- Para visualizar a epitelialização que ocorre no estágio inicial da formação organoide, prepare o mRNA para a proteína ocluidora de zonula occludens específica epitelial-1 (ZO-1) e para delinear as membranas celulares amplificando pCS2-ZO1-RFP e plasmídeos pCS2-mem-GFP (um presente de Lance Davidson).
- Extrair e linearizar o DNA plasmídeo e, em seguida, transcrever o mRNA tampado usando um kit de transcrição in vitro SP6/T7.
- Aliquot o mRNA transcrito e armazene-o a -80 °C.
- Microinjete o mRNA em um embrião fertilizado
- Coloque embriões fertilizados em 3% Ficoll em 1x MBS.
- Carregue 3-4 μL de mRNA usando uma ponta de micro-carregador em uma agulha de vidro puxada (uma ponta de agulha longa e fina com diâmetro interno de 10\u201230 μm).
- Conecte a agulha a um microinjetor e ajuste o tempo e a pressão para fornecer um volume constante de mRNA para microinjeção.
- Injete o mRNA logo abaixo da superfície apical do polo animal. Um patch circular de cor pálida distinto que é causado pela expansão do córtex é visível no momento da microinjeção.
- Transfira os embriões injetados para 1/3X MBS e cultue-os para o estágio 9.5.
- Colete os embriões fluorescentes rotulados sob um estereóscópio de fluorescência (configurações de excitação/emissão para GFP (488/510) e RFP (532/588)).
- Prossiga com a etapa 1.3.
- Montar e cultivar o organoide (seções 3 e 4) até o estágio desejado de desenvolvimento.
- Faça imagens ao vivo.
- Prepare uma câmara de imagem de fundo de vidro colando um vidro de cobertura em uma câmara de acrílico personalizada usando graxa de silício.
NOTA: Feche firmemente a câmara para evitar vazamentos da mídia cultural. - Encha a câmara de imagem com DFA.
- Escolha uma rede de microscopia eletrônica de transmissão hexagonal (TEM) (75 malha) de um recipiente usando fórceps e aplique uma pequena quantidade de graxa na borda da grade.
NOTA: O tamanho da malha deve ser menor do que o diâmetro do agregado para que o agregado fique na grade. - Pressione levemente para baixo para fixar a grade TEM na parte inferior da câmara de imagem.
- Transfira os agregados para a câmara de imagem e posicione-os dentro da grade.
NOTA: Evite posicionar os agregados ao lado da graxa. Durante todo o experimento, os agregados devem sentar-se na grade TEM sem entrar em contato com a parte inferior da câmara para evitar a compressão física. - Encha a câmara de imagem com DFA e sele-a com um copo de cobertura e graxa.
NOTA: A câmara deve ser hermética sem bolhas de ar para evitar qualquer turbulência ou movimento durante a imagem. - Para acompanhar a progressão da formação organoide epitelial mucociliary, colete imagens de pilha z-stack de tempo de agregados (a partir de 2 hpa) usando um microscópio confocal.
NOTA: Normalmente coletamos pilhas z de ~120 μm de espessura a cada 15 minutos usando um objetivo de 20X para seguir os comportamentos dinâmicos das células, mas essas especificações devem ser otimizadas para o objetivo de experimentos.
- Prepare uma câmara de imagem de fundo de vidro colando um vidro de cobertura em uma câmara de acrílico personalizada usando graxa de silício.
6. (Opcional) Imagem desenvolvendo organoides por fixação e imunostaining
- Corrija organoides na fase desejada de desenvolvimento transferindo-os para um frasco de vidro cheio de uma solução fixa.
NOTA: Adicione um volume de solução fixativa que é >20 vezes o das amostras para garantir a fixação completa. Execute os seguintes processos em um nutator, a menos que seja observado de outra forma. Em geral, os organoides são fixados com 4% de paraformaldeído (PFA) em PBS. No entanto, diferentes fixações podem ser necessárias para detectar proteínas específicas. Por exemplo, usamos 4% de PFA com glutaraldeído de 0,25% no PBS para detectar f-actina e tubulina acetilada. Para detectar intelectina (ITLN) e ZO-1, os organoides são fixados com a solução de Dent gelada (4:1 metanol:sulfoxida de dimetila) durante a noite a -20 °C. Os organoides fixos da Dent devem ser desidratados em série antes da lavagem (etapa 6.3). A duração da incubação e lavagem de anticorpos pode ser otimizada para necessidades específicas. - Fixar organoides por 15 min em temperatura ambiente (RT) ou durante a noite a 4 °C.
- Lave 3 vezes com PBST (PBS com Triton X-100) de 0,1% por 15 min na RT.
- Bloqueie a ligação inespecífica com soro de cabra de 10% em PBST (PBSGT) por 1 h na RT.
- Incubar com anticorpo primário (1:200) no PBSGT durante a noite a 4 °C.
- Lave 3 vezes com PBST por 15 min no RT.
- Incubar com anticorpo secundário (1:200) no PBSGT durante a noite a 4 °C.
- Lave 3 vezes com PBST por 15 min no RT.
- Transfira os organoides fixos e imunossuados para uma câmara de imagem e proceda com imagens confocal.
Resultados
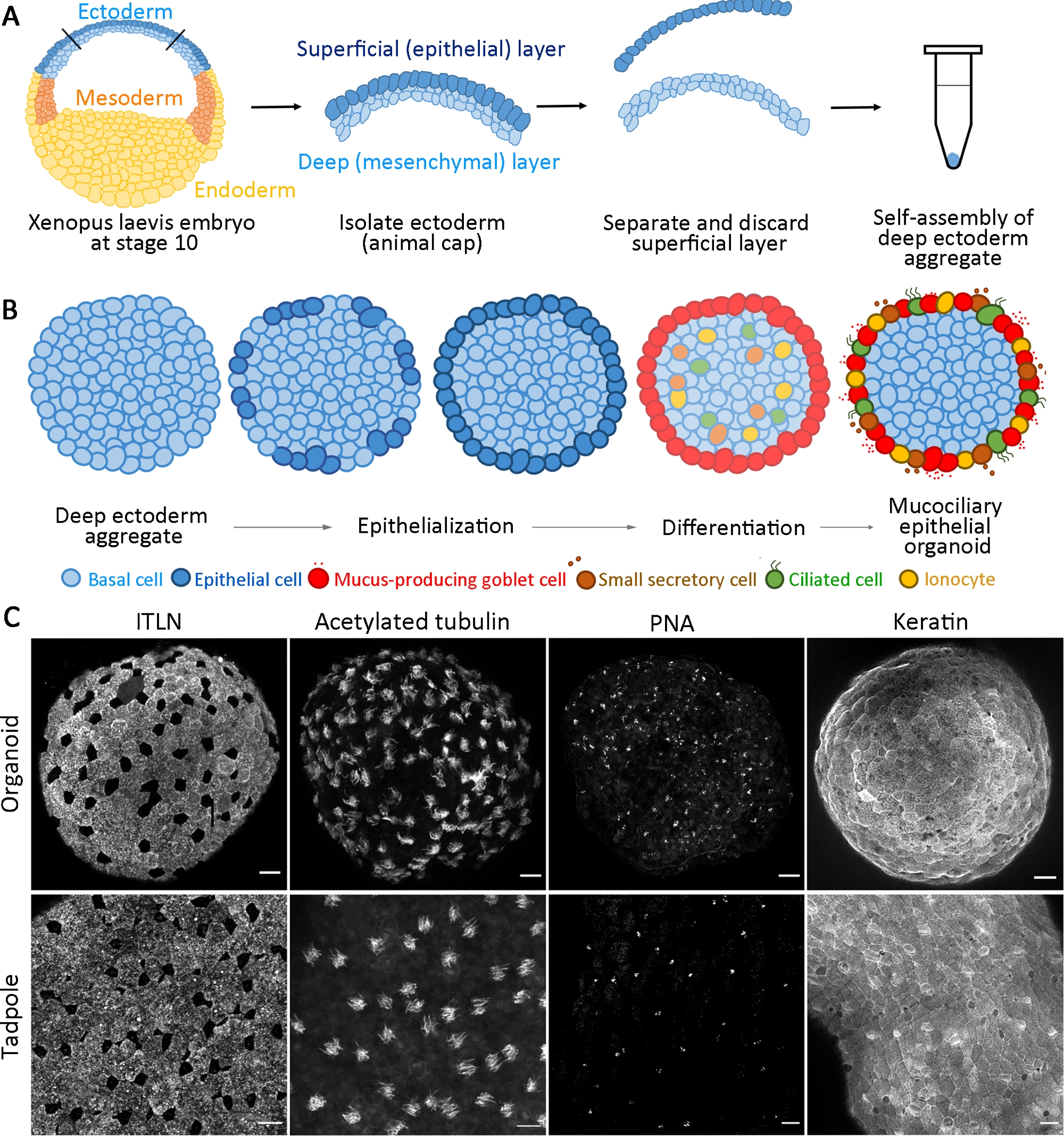
Este protocolo padronizado gera um organoide epitelial mucociliary de progenitores multipotentes isolados do estágio gastrula inicial X. laevis embriões dentro de 24 horas do cultivo14. Coletâneos células de ectoderme profundo se auto-montam para formar um agregado em um tubo PCR não adesivo e sofrem epitelialização superficial e diferenciação de células de cálice. A superfície recém-epitelializada dos agregados fornece um substrato semelhante ao epitélio nativo encontrado in vivo para intercalar células internas (por exemplo, multiciliárias e outras células acessórios) e desenvolve-se para formar organoides epiteliais mucociliais(Figura 1A,B). Dentro de 24 horas após a agregação, organoides epiteliais mucociliary auto-organizados regeneram uma epiderme madura que é indistinguível da epiderme de um girino. Os organoides compreendem epitélio totalmente diferenciado (queratina), células gúrias secretantes de muco (ITLN), células multiciliadas (tubulina acetilada) e pequenas células secretary (aglutinina de amendoim, PNA)(Figura 1C).
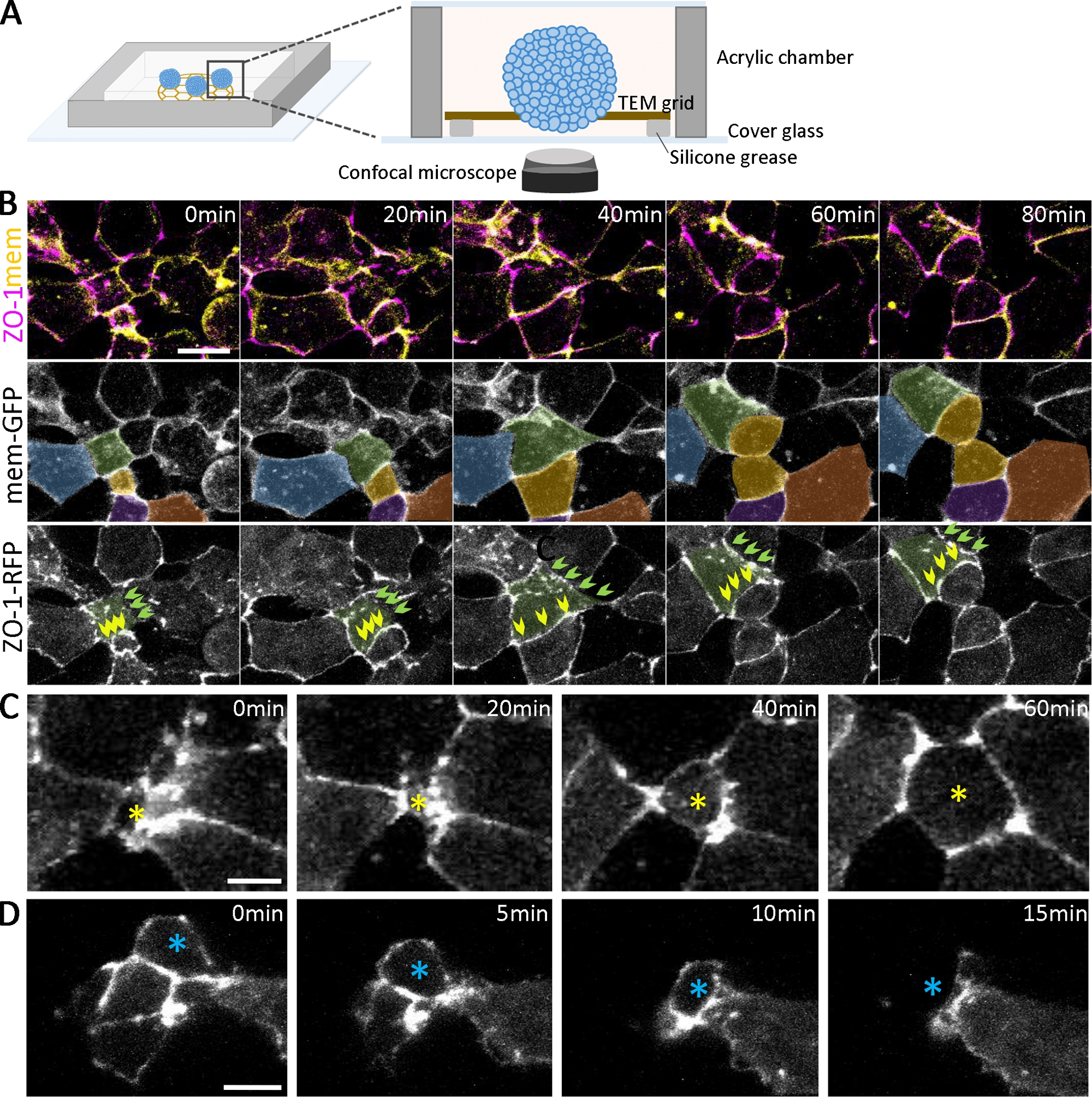
Além de confirmar o desenvolvimento de diferentes tipos celulares com imunossuagem, a dinâmica do desenvolvimento organoide pode ser seguida por imagem viva(Figura 2A). Para examinar a epitelialização que emerge no estágio inicial da formação organoide(Figura 1B),rotulamos embriões expressando proteínas de junção apertadas marcadas fluorescente (ZO-1-RFP) e proteínas de localização de membrana (mem-GFP). Com rotulagem dupla, os passos sequenciais da formação de junção apertada ZO-1 positivo podem ser marcados e analisados quantitativamente durante a epitelialização(Figura 2). Por exemplo, para células (Figura 2B, de cor verde) em diferentes estágios da epitelialização (em 0 min), algumas regiões de adesão celular têm espalhado puncta de ZO-1 (Figura 2B, setas verdes). Em contraste, outras áreas têm expressão ZO-1 totalmente montada (Figura 2B, setas amarelas). Com o tempo, o puncta coalesce e conecta-se para formar junções apertadas contíguas(Figura 2B, setas verdes), e junções apertadas contíguas mantêm sua morfologia mesmo durante a divisão celular(Figura 2B, setas amarelas). À medida que as junções apertadas amadurecem, as células se movem dinamicamente para dentro e para fora da superfície ao longo dos planos apical dos organoides(Figura 2C,D). Além disso, rastreando células espacialmente na superfície de organoides diferenciadores(Figura 2B, células codificadas por cores), a análise em várias escalas é possível, variando de puncta individual a junções apertadas contíguas, limites celular-células e subconjuntos de populações celulares dentro de organoides.

Figura 1: Geração de organoides epiteliais mucociliais.
(A) Um esquema mostrando o protocolo para montar agregados profundos de ectoderm de embriões X. laevis. (B) Um esquema para um modelo de formação organoide epitelial mucociliary originário de células ectoderme profunda multipotentes (visão transversal). Células posicionadas na superfície transitam em células epiteliais e se diferenciam em células de cálice. Diferenciando células ciliadas, células secretas e ionócitos intercalam radialmente na superfície e regeneram uma epiderme madura. (C) Projeção máxima z de epitélio mucociliary imunostidido para ITLN (células gúrias produtoras de muco), tubulina acetilada (células ciliadas), PNA (pequenas células secretary) e queratina (células epiteliais) em organoides a 24 hpa (painel superior) e epiderme de girinos (painel inferior). Barra de escala = 30 μm. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Imagem ao vivo de organoides em desenvolvimento.
(A) Um esquema da câmara de imagem para organoides vivos (para não escalar). (B) Sequências de lapso de tempo de pilhas confocal coletadas de agregados de células ectoderm profundas expressando ZO-1-RFP e mem-GFP de 2,5 hpa. Barra de escala = 20 μm. As células são pseudocoloridas para rastrear ao longo do tempo. As células de cor verde têm diferentes status de adesão celular, incluindo uma adesão positiva ZO-1 progressivamente em desenvolvimento (setas verdes) e uma mantendo a adesão positiva contígua ZO-1 (setas amarelas) ao longo do tempo. (C, D) Imagens confocalas de tempo de zo-1-RFP expressando agregados de células profundas ectoderm mostram as células radialmente intercalando para a superfície (C, estrela amarela) e movendo-se dentro dos agregados (D, estrela azul). Barras de escala = 10 μm. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
Organoides epiteliais mucociliais gerados a partir de células de ectoderme profundos do embrião X. laevis são uma ferramenta poderosa para estudar a epitelialização e diferenciação de progenitores multipotentes in vitro. Em contraste com o ensaio da tampa animal amplamente adotado16 utilizado para organogênese in vitro13 e o desenvolvimento da epitélio mucociliaria15,17,22 que utilizam o ectoderme intacto, os organoides derivados do ectoderme profundo apresentados neste protocolo oferecem uma oportunidade distinta de monitorar as fases de regeneração orientadas pela mecânica tecidual do epitélio superficial14. Em torno de 2 hpa, as células epiteliais positivas zo-1 recém-geradas(Figura 2) começam a aparecer na superfície apical dos organoides e aumentam sua população para cobrir todo o organoide à medida que o tecido solidifica ou reduz a conformidade14. A regeneração do epitélio e as especificações subsequentes de linhagem para células de cálice produtoras de muco prosseguem espontaneamente em uma mídia cultural quimicamente definida dentro de um dia. Estes organoides epiteliais mucociliais em rápido desenvolvimento fornecem uma plataforma para examinar comportamentos celulares dinâmicos em tempo real, em alta resolução, durante etapas progressivas de regeneração epitelial. Também permitem a investigação de questões fundamentais que surgem durante o desenvolvimento do epitélio mucociliary, homeostase e doenças associadas2,9,23. Em particular, a sensibilidade mecânica das células progenitoras profundas durante a transição para precursores de células de cálice epitelial identificadas nos organoides14 pode servir para ligar doenças respiratórias associadas à diferenciação basal anormal onde as células de cálice que se secretam muco estão sobre ou sub-produzidas23.
Embora este protocolo ofereça uma abordagem simples para gerar esses organoides, existem várias etapas críticas para o sucesso em experimentos. Para evitar a contaminação de células epiteliais superficiais durante o isolamento de células de ectoderme profundo da tampa animal, deve-se monitorar a tampa animal colocada em DFA livre de cálcio e magnésio sob um estereoscópio e detectar o momento certo para iniciar a separação da camada superficial pigmentada escura da tampa animal. Se o tecido for mantido em DFA livre de cálcio e magnésio por muito tempo, todo o tecido se dissociará e distinguir entre células profundas e superficiais seria então impossível para agregados profundos de ectoderme. Para confirmar a ausência de células superficiais em agregados de ectoderme profundo, recomendamos rotular fluorescentemente a superfície apical do embrião com NHS-rhodamina (passo 1.414) antes da microcirurgia; isso permitiria fácil identificação de células superficiais se existissem nos organoides resultantes. Uma vez que a regeneração epitelial é regulada pela mecânica tecidual14, é essencial evitar a geração de força não intencional para organoides auto-organizados. Em particular, sugerimos evitar o contato com o fundo de vidro da câmara de imagem durante a imagem ao vivo, colocando agregados nas bordas das grades TEM, pois isso permite o contato livre com a janela de imagem dos agregados ao vivo (passo 5.1.2.). Este modelo 3D in vitro-cultivado e auto-organizado para epitélio mucociliário servirá como uma ferramenta tratável para responder às questões fundamentais que surgem durante a regeneração do epitélio e a especificação de linhagem das células de cálice.
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Agradecemos aos membros do laboratório Kim e Lance Davidson por seus comentários e apoio. Este trabalho foi apoiado pela Young Scientist Fellowship para hyk do Institute for Basic Science (IBS-R0250Y1).
Materiais
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Dual-stage Glass Micropipette Puller | Narishige | PC-100 | |
| Picoliter microinjector | Warner Instruments | PLI-100A | |
| Confocal Laser Microscope | |||
| Stereoscope | |||
| Tools | |||
| Forcep | Dumont | Dumont #5 | |
| Hair knife | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| Hair loop | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| hCG injection | |||
| human chorionic gonadotropin | Sigma | cg10-10vl | |
| MBS solution | |||
| 10M Sodium hydroxide | Sigma | 72068 | |
| Calcium chloride | Sigma | C3881 | |
| Calcium nitrate | Sigma | C1396 | |
| HEPES | Sigma | H4034 | |
| Magnesium sulfate | Sigma | 230391 | |
| Phenol-red | Sigma | P0290 | |
| Potassium chloride | Sigma | 7447-40-7 | |
| Sodium bicarbonate | Sigma | S6014 | |
| Sodium chloride | Sigma | S9625 | |
| Sodium hydroxide reagent grade, 97%, powder-25g | Sigma | 655104 | |
| dejellying solution | |||
| L-Cysteine hydrochloride monohydrate | Sigma | C7880 | |
| Sodium hydroxide 10M | Sigma | 72068 | |
| Ficoll solution | |||
| Ficoll | Sigma | F4375 | |
| DFA solution | |||
| Sodium chloride | Sigma | S9625 | |
| 0.22mm Filter | Millipore | S2GPT05RE | |
| Antibiotic Antimycotic Solution | Sigma | A5955 | |
| Bicine | Sigma | B3876 | |
| Calcium chloride | Sigma | C3881 | |
| Magnesium sulfate | Sigma | 230391 | |
| Potassium gluconate | Sigma | G4500 | |
| Sodium carbonate | Sigma | 222321 | |
| Sodium gluconate | Sigma | G9005 | |
| mRNA in vitro transcription | |||
| SP6/T7 in vitro transcription kit | Invitrogen | AM1340 | |
| mRNA microinjection | |||
| Borosilicate glass capillary tubes | Harvard Apparatus | GC100-10 | |
| Eppendorf microloader pipette tips | ThermoFisher | A25547 | |
| Mineral oil | Sigma | M5904 | |
| PCR tube coating | |||
| BSA | Thermofisher | 26140079 | |
| PCR tubes | SSI | SSI-3245-00 | |
| Imaging | |||
| Custom-milled acrylic chamber | |||
| Coverglass 24mmX50mm | Duran | B01_001650 | |
| SPI Hexagonal TEM Grids, Gilded Nickel (50mesh) | SPI | 275HGN-XA | |
| SPI Hexagonal TEM Grids, Gilded Nickel (75mesh) | SPI | 2775GN-XA | |
| Silicone grease | Shinetsu | HIVAC-G | |
| Fixation | |||
| 20ml screw top-cap vial | Wheaton | WH.986580 | |
| 2ml screw top-cap vial | |||
| Benzyl alcohol | Sigma | 305197 | |
| Benzyl benzoate | Sigma | B6630 | |
| Dimethyl sulfoxide (DMSO) | Sgima | D4540 | |
| Glutaraldehyde 10% EM GRADE | Electron Microscopy Sciences | 16120 | |
| Goat serum | Jackson | 005-000-121 | |
| Methanol | Sigma | 322415 | |
| Paraforlamdehyde | Sigma | P6148 | |
| Phosphate-buffered saline (PBS) | LPS Solution | CBP007B | |
| Triton X-100 | Sigma | T8787 | |
| Primary antibody (1:200) | |||
| acetylated tubulin | Sigma | clone 6-11B-1 | |
| Itln1 | Proteintech | 11770-1-AP | |
| Keratin | Developmental Studies Hybridoma Bank | 1h5 | |
| ZO1 | Invitrogen | 402200 | |
| Vectors | |||
| pCS2-mem-GFP | Gift from Dr. Lance Davidson | ||
| pCS2-ZO1-RFP | Gift from Dr. Lance Davidson |
Referências
- Barkauskas, C. E., et al. Lung organoids: current uses and future promise. Development. 144 (6), 986-997 (2017).
- Puchelle, E., Zahm, J. M., Tournier, J. M., Coraux, C. Airway Epithelial Repair, Regeneration, and Remodeling after Injury in Chronic Obstructive Pulmonary Disease. Proceedings of the American Thoracic Society. 3 (8), 726-733 (2006).
- Tilley, A. E., Walters, M. S., Shaykhiev, R., Crystal, R. G. Cilia dysfunction in lung disease. Annual Review of Physiology. 77, 379-406 (2015).
- Vareille, M., Kieninger, E., Edwards, M. R., Regamey, N. The Airway Epithelium: Soldier in the Fight against Respiratory Viruses. Clinical Microbiology Reviews. 24 (1), 210-229 (2011).
- Rock, J. R., et al. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proceedings of the National Academy of Sciences of the United States of America. 106 (31), 12771-12775 (2009).
- Sachs, N., et al. Long-term expanding human airway organoids for disease modeling. The EMBO Journal. 38 (4), 100300(2019).
- Dubaissi, E., et al. A secretory cell type develops alongside multiciliated cells, ionocytes and goblet cells, and provides a protective, anti-infective function in the frog embryonic mucociliary epidermis. Development. 141 (7), 1514-1525 (2014).
- Hayes, J. M., et al. Identification of novel ciliogenesis factors using a new in vivo model for mucociliary epithelial development. Developmental Biology. 312 (1), 115-130 (2007).
- Walentek, P., Quigley, I. K. What we can learn from a tadpole about ciliopathies and airway diseases: Using systems biology in Xenopus to study cilia and mucociliary epithelia. Genesis. 55 (1-2), (2017).
- Werner, M. E., Mitchell, B. J. Understanding ciliated epithelia: The power of Xenopus. Genesis. 50 (3), 176-185 (2012).
- Dubaissi, E., Papalopulu, N. Embryonic frog epidermis: a model for the study of cell-cell interactions in the development of mucociliary disease. Disease Models & Mechanisms. 4 (2), 179-192 (2011).
- Walentek, P., et al. A novel serotonin-secreting cell type regulates ciliary motility in the mucociliary epidermis of Xenopus tadpoles. Development. 141 (7), 1526-1533 (2014).
- Asashima, M., et al. In vitro organogenesis from undifferentiated cells in Xenopus. Developmental Dynamics. 238 (6), 1309-1320 (2009).
- Kim, H. Y., Jackson, T. R., Stuckenholz, C., Davidson, L. A. Tissue mechanics drives regeneration of a mucociliated epidermis on the surface of Xenopus embryonic aggregates. Nature Communications. 11 (1), 665(2020).
- Haas, M., et al. DeltaN-Tp63 Mediates Wnt/beta-Catenin-Induced Inhibition of Differentiation in Basal Stem Cells of Mucociliary Epithelia. Cell Reports. 28 (13), 3338-3352 (2019).
- Green, J. Molecular Methods in Developmental Biology: Xenopus and Zebrafish. Guille, M. , Humana Press. 1-13 (1999).
- Stubbs, J. L., Davidson, L., Keller, R., Kintner, C. Radial intercalation of ciliated cells during Xenopus skin development. Development. 133 (13), 2507-2515 (2006).
- Nieuwkoop, P. D., Faber, J. Normal table of Xenopus laevis (Daudin) : a systematical and chronological survey of the development from the fertilized egg till the end of metamorphosis. , Garland Pub. (1994).
- Sive, H. L., Grainger, R. M., Harland, R. M. Early development of Xenopus laevis : a laboratory manual. , Cold Spring Harbor Laboratory Press. (2000).
- Joshi, S. D., Kim, H. Y., Davidson, L. A. Microscopy tools for quantifying developmental dynamics in Xenopus embryos. Methods in Molecular Biology. 917, 477-493 (2012).
- Sater, A. K., Steinhardt, R. A., Keller, R. Induction of neuronal differentiation by planar signals in Xenopus embryos. Developmental Dynamics. 197 (4), 268-280 (1993).
- Sedzinski, J., Hannezo, E., Tu, F., Biro, M., Wallingford, J. B. Emergence of an Apical Epithelial Cell Surface In Vivo. Developmental Cell. 36 (1), 24-35 (2016).
- Rock, J. R., Randell, S. H., Hogan, B. L. M. Airway basal stem cells: a perspective on their roles in epithelial homeostasis and remodeling. Disease Models & Mechanisms. 3 (9-10), 545-556 (2010).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados