Method Article
Два различных в режиме реального времени Место Предпочтение Парадигмс использованием оптогенетики в вентральной tegmental области мыши
В этой статье
Резюме
Здесь мы представляем два простых в последующей пошаговые протоколы для парадигмы предпочтения места с использованием оптогенетики у мышей. Используя эти две различные установки, предпочтения и поведение избегания могут быть надежно оценены в одном аппарате с высокой пространственной и временной избирательностью, и в простой манере.
Аннотация
Понимание того, как активация нейронов приводит к специфическим поведенческим выходу, имеет основополагающее значение для современной неврологии. Сочетание оптогенетики у грызунов с поведенческим тестированием в проверенных парадигмах позволяет измерять поведенческие последствия при стимуляции различных нейронов в режиме реального времени с высокой пространственной и временной избирательностью, и, таким образом, установление причинно-следственных связей между активацией нейронов и поведением. Здесь мы описываем пошаговый протокол для парадигмы предпочтений местоположения в реальном времени (RT-PP), модифицированную версию теста предпочтения кондиционированных мест (CPP). RT-PP выполняется в трех-отсекном аппарате и может быть использован для ответа, если оптогенетическая стимуляция конкретной нейронной популяции является полезным или отрицательным. Мы также описываем альтернативный вариант протокола RT-PP, так называемый протокол предпочтения нейтрального отсека (NCP), который может быть использован для подтверждения неприятия. Эти два подхода основаны на расширении классической методологии, основанной на поведенческой фармакологии и недавней реализации оптогенетики в области неврологии. Помимо измерения предпочтений места в режиме реального времени, эти установки могут также дать информацию об условном поведении. Мы предоставляем простые в последующей пошаговые протоколы наряду с примерами наших собственных данных и обсуждаем важные аспекты, которые следует учитывать при применении такого рода экспериментов.
Введение
Внедрение оптогенетики, современный нейробиологический экспериментальный инструмент, в котором свет используется для контроля нейронной активности, в последние годы привело к крупным достижениям в понимании того, как конкретные нейронные популяции влияют на поведение1,2,3. Выдающаяся пространственная и височная избирательность оптогенетики позволяет установить причинно-следственные связи между возбуждением или ингибированием клеточных групп, представляющих интерес, и поведенческим выходом2,3. Пространственная селективность в оптогенетике обычно обеспечивается через систему Cre-Lox, в которой активность рекомбины Cre приводит к рекомбинации любых последовательностей ДНК, присутствующих между участками Локса, так называемых флексированных аллелей (в окружении локс-сайтов)4. Цель с использованием системы Cre-Lox в оптогенетике заключается в достижении экспрессии аллелей, кодирующих оптогенетические опсины в специфических нейронах, представляющих интерес, оставляя окружающие нейроны лишенными экспрессии. Опсины являются светочувствительными белками, которые при светостимуляции конкретной длины волны позволяют ионный поток, который влияет на нервную возбудимость или влияет на клеточные функции, модулируя вниз по течению эффектор пути. Новые варианты опсинов, которые отличаются по действию (возбуждающие, ингибирующие, модуляторные), механизм, активация световой длиной волны и кинетическими свойствами5 постоянно разрабатываются для удовлетворения потребностей конкретных экспериментальных подходов. Что касается возбудимости, с помощью деполяризации или гиперполяризационизации opsin диктует активность нейронов (возбуждение или ингибирование, соответственно) на свет-стимуляции на определенной длине волны доставлены в мозг3.
Селективная активность промоутера направляет выражение рекомбиназы Cre к понам интереса. Путем снабжать floxed аллель opsin интереса, Cre-опосредованный recombination обеспечит что opsin селективно выражено в невенарах которые co-выразить Cre recombinase3,6. Это использование двойной трансгении для прямого пространственной избирательности оказалось очень эффективным в оптогенетике. Таким образом, в то время как свет-стимуляция для активации опсины широко поставляется через внутримозгово имплантированных оптических волокон, подключенных к источнику света (LED или лазер)3, только нейроны, выражающие как Cre recombinase и floxed opsin аллель будет реагировать на эту стимуляцию. Система Cre-Lox у грызунов может быть достигнута по-разному, используя только трансгении (как Cre recombinase и floxed opsin конструкции кодируются в трансгенных животных), только вирусные инъекции (ДНК конструкции для Рек рекомбиназа и флоксированный опсин оба поставляются через вирусный перевозчик), или сочетание двух (например, Крем рекомбиназы кодируется трансгенного животного, которое вводится с вирусом, несущим floxed opsin конструкции)5. Конструкция ДНК флокса opsin обычно клонируется в кадре с геном репортера, чтобы обеспечить визуализацию рекомбинации Cre-опосредоченной в секциях тканей. В то время как оптогенетика также может быть выполнена у крыс, представленные протоколы были созданы для мышей. Для простоты, мышей, несущих как Рек рекомбиназы и флоксированный опсин будет называться "оптогенетика мышей". В протоколах, описанных ниже, оптогенетика мышей были созданы смешанные трансгенные (Cre рекомбиназы под контролем двух различных промоутеров) и вирусных (с использованием адено-ассоциированного вируса, AAV, чтобы доставить флоксированный опсин ДНК конструкции - в нашем случае ChR2/H134R) подход. Получение и поддержание трансгенных линий мыши является неотъемлемой частью методологии. Cre-драйвер и флоксированный опсин трансгенных мышей могут быть произведены для каждой цели, или приобрели, если коммерчески доступны, как и целый ряд вирусов, несущих последовательности ДНК кодирования Cre recombinase и floxed opsins в различных формах.
Оптогенетика в сочетании с поведенческим тестированием оказалась ценным инструментом для изучения роли различных областей мозга, или дискретных нейронных популяций, в частности типов поведения. В контексте поведения, связанного с вознаграждением, оптогенетика позволила проверить предыдущие выводы в области поведенческой фармакологии и экспериментальной психологии, а также позволила новый уровень пространственно-временно релевантного рассечения в том, как некоторые нейроны влияют на поведение. Одним из методов, который был использован в нескольких исследованиях для оценки поведения, связанного с вознаграждением, является модифицированная версия классического метода, известного как Условные предпочтения места (CPP). Классическая CPP была использована для оценки награждения или aversive свойства наркотиков злоупотребления через их способность вызывать Павловские ассоциации с сигналами окружающей среды7,8. С точки зрения Павлова, препарат является безусловным стимулом, поскольку он может вызвать подход или вывод, если он является полезным или неблагоприятным, соответственно. Непрерывное спаривание препарата с различными нейтральными стимулами, которые сами по себе не вызывают никакой реакции, может привести к приближению или выводу только при предъявлении ранее нейтральных, но после спаривания, так называемых условных стимулов9. Анализ CPP обычно выполняется в аппарате, содержащем два отсека одного размера, но где каждый отсек определяется различными характеристиками, такими как текстура пола, настенные узоры и освещение (нейтральные стимулы). Два отсека соединены либо коридором, либо отверстием между отсеками. Во время кондиционирования, субъект, как правило, небольшой грызун, получает пассивные инъекции препарата в то время как ограничивается одним из двух основных отсеков и сольственный при ограничении в другом отсеке. Награждение эффекты препарата впоследствии оцениваются в свободной от наркотиков сессии, когда субъект имеет право свободно исследовать весь аппарат. Количество времени, затрачиваемого в ранее снадобье-парной отсек (условный ответ) считается отражением Павловских механизмов обучения, опосредованных между полезными эффектами препарата и сигналами отсека, связанных с его администрированием(условные стимулы). Если животное проводит больше времени в отсеке для спаривания наркотиков, препарат индуцировал условные предпочтения место, что означает, что он имеет полезные последствия для поведения. С другой стороны, если препарат воспринимается как аверсивный, животное будет избегать наркотиков парной отсек и проводить больше времени в солевой паре отсека, указывая условное место отвращение (CPA)8,9,10,11.
Так как оптогенетика может быть реализована для контроля нейронной активности в "реальном времени", использование поведенческой парадигмы, аналогичной, но отличается от установки CPP позволяет измерять предпочтения места при прямой активации нейронов. Оптогенетика-управляемый анализ предпочтений места поэтому часто refer to как в реальном времени парадигма анализа предпочтений места (RT-PP). В парадигме RT-PP оптогенетическая стимуляция различных нейронов через систему Cre-Lox заменяет системную доставку препарата, выполняемого в классическом CPP, так что парадигма RT-PP вместо этого измеряет, если оптогенетически индуцированная нейронная стимуляция воспринимается как награждение или aversive. Текущее описание будет сосредоточено на оптогенетики мышей, но и оптогенетики крысы могут быть протестированы с помощью аналогичных протоколов.
Вместо того, чтобы кондиционирования в одном отсеке в то время, как в классической парадигме CPP, оптогенетика мыши в парадигме RT-PP разрешено свободно перемещаться во всем аппарате и поведение записывается на протяжении всего сеанса. Вход в один из отсеков сопряжен с внутричерепной светостимуляцией. При соответствующих параметрах стимуляции света, нейроны, которые выражают возбуждающее опсин, тем самым будут активированы. Если светостимуляция воспринимается как полезная, мышь оптогенетики останется в светло-парном отсеке, в то время как если свет-стимуляция воспринимается как аверзивная, мышь выйдет из отсека, чтобы избежать стимуляции. Этот тип анализа позволяет оценить условное обучение: субъект может вызвать светостимуляцию и, следовательно, активацию нейронов, введя в отсек, и остановить стимуляцию, выйдя из отсека, подобно рычагу нажатия во время инструментальной задачи. Кроме того, механизмы ассоциативного обучения могут быть оценены во время последующих сессий, где время, проведенное в каждом отсеке измеряется в отсутствие стимуляции. Таким образом, исследователь может разъединяться между немедленно полезным и воздействием на стимуляцию нейронов, представляющих интерес, и ассоциативной формирования памяти, связанные с ним12.
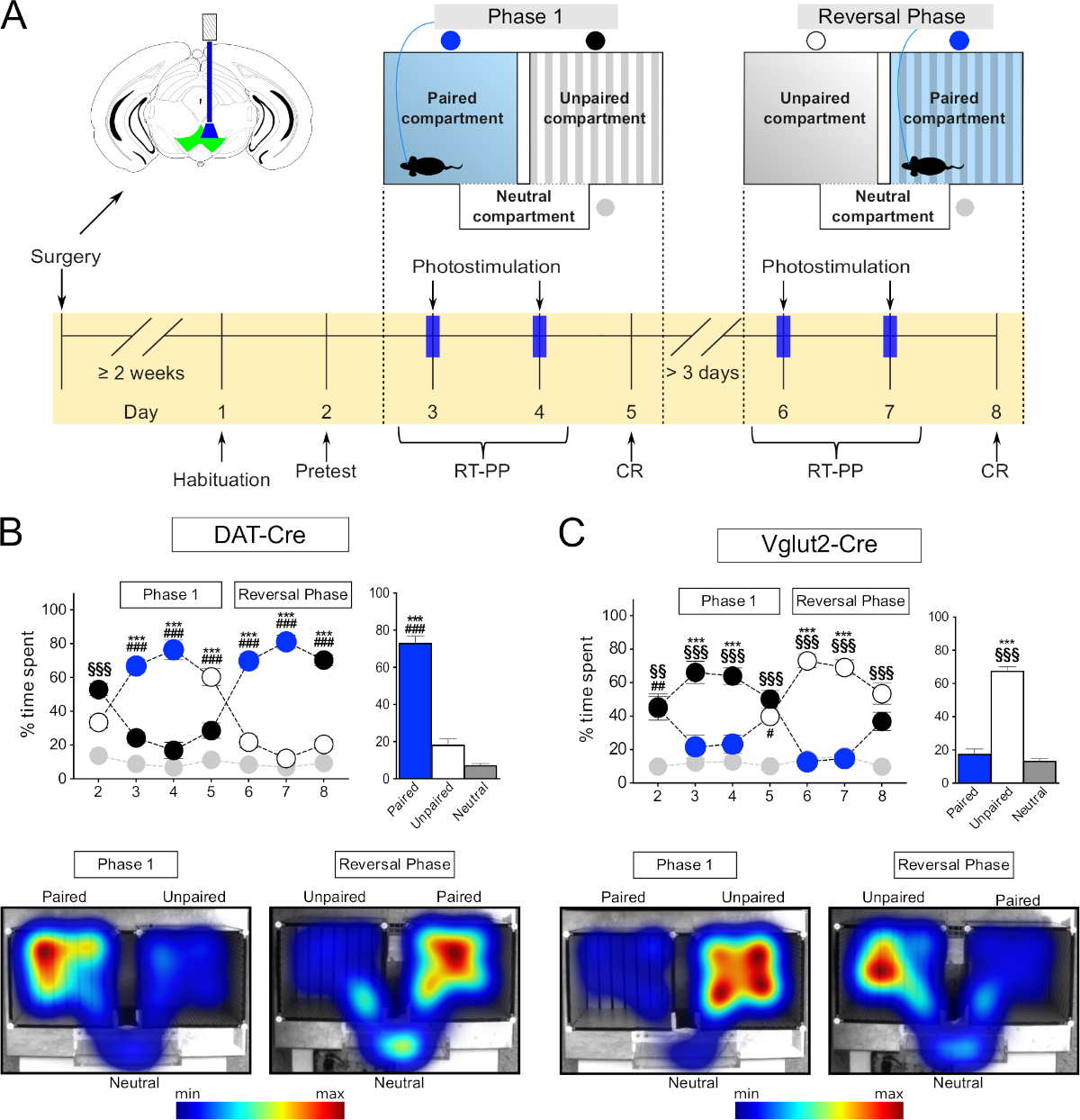
В текущем исследовании мы описываем два протокола пошаговой настройки для оптогенетики инициативе место предпочтения поведения свободно движущихся мышей. Первый протокол описывает RT-PP в трехкомпонентном аппарате и был изложен на основе протоколов, недавно представленных Root и коллегами 13 и другими авторами12,14,15,16,17,18. Эксперимент состоит из двух этапов, включающих несколько ежедневных сеансов (показано на рисунке 1A). Каждая сессия предназначена для различных целей и параметры стимуляции соединения с отсеком меняются соответствующим образом. Первая сессия,«Претест»,используется для оценки первоначального предпочтения субъекта к одному из отсеков. При подключении к патч шнур, субъект имеет право свободно исследовать аппарат в отсутствие стимуляции в течение 15 минут. Если начальное предпочтение одному отсеку составляет более 80%, мышь исключается из анализа, так как начальная боковая предвзятость может исказить анализ. После"Претеста"начинаетсяфаза 1. Первая часть состоит из двух последовательных, ежедневных, 30 минут сессий "RT-PP". Во времяфазы 1", оптогенетика мышь подключена к источнику лазера через патч шнур и помещены в арене, чтобы свободно исследовать его. Мышь получает внутричерепную лазерную стимуляцию при входе в один из основных отсеков. Пилотные эксперименты могут быть выполнены, чтобы определить, какой отсек будет назначен в качестве лазерного пара, а как неспаренный. Если будет показано, что стимуляция вознаграждает, лазер будет соединен с наименее предпочтительным отсеком во время«Претеста»и с наиболее предпочтительным, если стимуляция является неприятной. Таким образом, представленный протокол RT-PP следует предвзятой конструкции в том смысле, что лазерная стимуляция не случайным образом назначается любому из двух основных отсеков (непредвзятый дизайн), но выбран, чтобы избежать каких-либо первоначальных предпочтений мыши. Вход в другой основной отсек или нейтральный отсек, соединяющий два основных отсека, не приводит к стимуляции внутричерепного света и, таким образом, не имеет световой пары. Эти сеансы позволяют в режиме реального времени оценивать полезные или аверсивные свойства стимуляции конкретных нейронных популяций. В последний день1-го этапапроводится 15-минутная сессия без стимуляции. Эта сессия служит для решения условных ответов("CR"), которые являются результатом ассоциативного обучения между стимуляцией и окружающей средой, где она была получена. По крайней мере, через три дня после «Фазы 1», происходит«Фаза разворота»,которая следует той же структуре, что и «Фаза 1», но ранее непарный основной отсек теперь в паре со световой стимуляцией. Как и в случае с фазой1, за двумя сеансами стимуляции следует сессияCR. «Вспять»используется для подтверждения того, что поведение мыши зависит от оптогенетической стимуляции и не связано со случайными параметрами. Каждая сессия эксперимента RT-PP должна быть отдельно запрограммирована в рамках программного обеспечения для отслеживания. Текущий протокол описывает такие настройки в рамках конкретного программного обеспечения, но любое другое программное обеспечение отслеживания, способное отправлять сигналы модуляции транзистора-транзистора (TTL) в источник света.
Второй протокол описывает новую настройку, названную парадигмой нейтрального содействия (NCP). Этот модифицированный протокол RT-PP использует небольшой размер и прозрачность соединительного коридора, которого мышь естественным образом избегает из-за ее узкого и прозрачного состава. Путем сопряжения обоих основных отсеков со светостимуляцией и только оставляя коридор свободным от светостимуляции, установка NCP может быть использована для проверки того, заставит ли оптогеническая стимуляция мыши проводить больше времени в коридоре, чтобы избежать получения оптогенетической стимуляции. Сравнивая время, проведенное в двух светлых отсеках, с временем, проведенным в коридоре, можно провести проверку неприятия, вызванное оптогенетически. Эксперимент NCP состоит из двух последовательных ежедневных сессий, где оптогенетика мышей получают стимуляцию (30 мин каждый) для измерения предпочтений в режиме реального времени, и один сеанс без лазера (15 мин) для оценки условных ответов аналогично тем, которые в RT-PP Протокол.
Протоколы RT-PP и NCP, представленные ниже, были недавно проверены в нашей лаборатории в исследовании того, как различные типы нейронов, расположенных в брюшной тегментальной области (VTA), участвуют в различных аспектах поведения, связанного с вознаграждением12. Здесь, чтобы иллюстрировать реализацию протоколов РТ-ПП и NCP, допамина транспортер (DAT)-Cre19 и везикулярный глутамат транспортер 2 (VGLUT2)-Cre20 трансгенных мышей были стеротактически введены с AAV проведения флоксированный каналrhodopsin2 (ChR2) ДНК, построенный в VTA Поведенческие реакции, полученные при анализе этих мышей с использованием предоставленных протоколов RT-PP и NCP, показывают, как активация дофаминергических и глутаматергических нейронов в ВТА приводит к различным поведенческим реакциям (Рисунок 1).
Пошаговые протоколы для парадигм RT-PP и NCP предоставляются с информацией, начиная от генотипирования трансгенных мышей, стереотаксических вирусных инъекций и размещения волоконно-оптических, до программирования отслеживания программного обеспечения для лазерного контроля и поведенческих Оценки. Кроме того, обсуждаются предложения по изменению протокола с точки зрения параметров стимуляции и экспериментальных аспектов, которые могут повлиять на научный результат. Хотя протоколы описаны в контексте VTA, они могут быть применены к любой области мозга или нейронной популяции, при условии, соответствующие инструменты оптогенетики, такие как соответствующие Cre-драйвер и флоксированные опсины, доступны.
протокол
Это исследование было проведено с использованием гетерозиготных DAT-Cre19 и VGLUT2-Cre20 мышей обоих полов, в возрасте от 8 недель и весом 20 г. Все эксперименты проводились в соответствии со шведским (Закон о защите животных SFS 1998:56) и законодательством Европейского союза (Конвенция ETS 123 и Директива 2010/63/EU) с разрешения местных комитетов по этике животных.
1. Генотипирование мышей
- Возьмите ухо биопсии с помощью уха перфоратор для использования для генотипирования трансгенных мышей.
- Подготовьте удары уха для выполнения реакции полимеразы цепной реакции (ПЦР) с использованием специально горитов.
ПРИМЕЧАНИЕ: В этом протоколе были использованы праймеры Cre-directed.- Добавьте 75 кл люсис буфера (буфер 1: 250 мМ NaOH, 2 мМ EDTA) в каждой трубке 1,5 мл, содержащей ушной пунш.
- Инкубировать в нагревательном блоке при 96 градусах по Цельсию в течение 30 мин.
- Дайте образцам остыть в течение 5 мин, а затем добавьте 75 qL буфера нейтрализации (буфер 2: 400 мМ Tris-HCl pH 8.0).
- Выполните ПЦР в соответствии со стандартными процедурами12,21 с использованием соответствующих грунтовок (здесь: Cre FW 5'-ACGAGGATGATGTTCGCAA-3', Cre REV 5'-ACCGACGATGAAGCATGTTTAG-3').
ВНИМАНИЕ: Работайте на льду под капотом ПЦР и обращайте внимание на то, чтобы не загрязнять реагенты и образцы.- Подготовьте мастер-микс ПЦР. Умножьте следующие тома в зависимости от количества образцов, которые будут проанализированы, включая соответствующие контрольные образцы. Смешайте реагенты для одной реакции окончательного объема 25 л в следующем порядке: дистиллированная вода (18,9 л), 10-x буфер с MgCl2 (2,5 л), смесь dNTP 10 мМ (смесь dNTP (10 мМ dNTP) 0,5 л), 10 мкм вперед грунтовка (1 зЛ), 10 ММ обратной грунтовки (1 Зл), 5 U / L ДНК полимераза (0,1 л), и шаблон ДНК (1 Л; будут добавлены в следующих шагах).
ПРИМЕЧАНИЕ: Всегда добавляйте негатив, положительный и пустой (без шаблона ДНК) контроль, чтобы обеспечить действительные результаты. - Добавьте 24 зЛ мастер-миксвв в трубках ПЦР.
- Добавьте 1 зл шаблона ДНК (исходя из ушной пунш от каждой мыши) в каждой трубке ПЦР.
- Centrifuge ПЦР трубки кратко обеспечить шаблон ДНК находится внутри мастер смеси.
- Выполните ПЦР с тепловым велосипедистом, используя программу велоспорта в таблице 1.
- Подготовьте мастер-микс ПЦР. Умножьте следующие тома в зависимости от количества образцов, которые будут проанализированы, включая соответствующие контрольные образцы. Смешайте реагенты для одной реакции окончательного объема 25 л в следующем порядке: дистиллированная вода (18,9 л), 10-x буфер с MgCl2 (2,5 л), смесь dNTP 10 мМ (смесь dNTP (10 мМ dNTP) 0,5 л), 10 мкм вперед грунтовка (1 зЛ), 10 ММ обратной грунтовки (1 Зл), 5 U / L ДНК полимераза (0,1 л), и шаблон ДНК (1 Л; будут добавлены в следующих шагах).
- Подготовьте гель агарозы для запуска образцов с помощью электрофореза.
ПРИМЕЧАНИЕ: Размер будет зависеть от количества образцов, которые должны быть проанализированы.- Добавьте 1% w/v агарозного порошка в буфер е 1x Tris-ацетат-EDTA (TAE) в стеклянной бутылке. Тепло в микроволновой печи, пока агароза полностью растворяется, регулярно проверяя, что она не кипит.
ВНИМАНИЕ: Примите меры предосторожности, чтобы избежать ожогов. - Дайте гелю остыть примерно до 50 градусов по Цельсию и добавьте пятно геля нуклеиновой кислоты (0,5 л/50 мл геля).
- Налейте гель в литье лоток, содержащий хорошо гребни и оставить его в комнатной температуре, пока он не станет полностью затвердевших. Снимите расчески осторожно.
- Заполните электрофорекс бак с 1x TAE буфера и поместите гель в бак.
- Добавьте 2 Зл из 1x КРАНа загрузки ДНК в каждом из образцов ДНК.
- Загрузите 4 зл и рясти ДНК в первую скважину геля, затем приступаем к загрузке полного объема образцов в оставшиеся скважины.
- Установите источник питания электрофореза до 140 В и проработав 25–30 мин.
- Поместите гель под ультрафиолетовым источником и сфотографировать результаты.
- Добавьте 1% w/v агарозного порошка в буфер е 1x Tris-ацетат-EDTA (TAE) в стеклянной бутылке. Тепло в микроволновой печи, пока агароза полностью растворяется, регулярно проверяя, что она не кипит.
2. Стереотаксическая хирургия
- После генотипирования, отдельные мыши поддержанию те положительные для Cre. Подождите, пока они, по крайней мере 8 недель и весят 20 г, чтобы выполнить операцию.
- Обезвить окружающую среду и стерилизовать хирургические инструменты для выполнения операции в асептических условиях.
- Вводят мышей подкожно с анальгетиком 30 мин до операции.
- Анестезия мышей с изофлюран (2-3% в нормальном воздухе для индукции и 1,5-2,0% для поддержания анестезии). Обеспечить адекватный уровень анестезии достигается путем тестирования отсутствия болевые рефлексы, мягко щипать нос мыши. Отрегулируйте доставку изофруран соответственно.
- Поместите мышь на стереотаксический аппарат. Добавьте смазку для глаз, чтобы предотвратить повреждение глаз из-за сухости и побрить волосы верхней части черепа. Используйте грелку для поддержания температуры мыши стабильной.
- Введите под кожу черепа 100 л местной анестезии и дайте 5 мин на то, чтобы ввести в действие.
- Подготовьте место разреза тремя круговыми применениями алкоголя или стерильного солья, чередующегося с йодом. Используйте стерильный кончик хлопка и инициируйте применение от линии разреза, наружу.
- Аккуратно поднимите кожу щипками и сделайте разрез в 1,5 см вдоль ротрокадальной оси хирургическими ножницами, чтобы выявить поверхность черепа.
- Используя хлопчатобумажную палочку, нанесите h2O2 раствор, чтобы удалить периостеум.
- Промыть череп стерильным сольниковиком и высушить с помощью стерильных аппликаторов кончиков хлопка.
- Найдите брегму и лямбу.
- Выясните выравнивание плоского черепа, позиционируя кончик иглы для инъекций, нарежденный на стереотаксической раме, на брегму и лямбду. Измерьте вентральные координаты для каждой позиции и сравните. Когда череп плоский, вентральные координаты как для брегмы, так и для лямбды идентичны. Если нет, отрегулируйте положение головы и снова произвносите измерения.
- Найти и отметить положение (AP: -3.45 мм, ML: -0,2 мм от bregma в соответствии с Франклином и Paxinos22), где инъекции Cre-зависимого вируса и имплантации оптического волокна будет проходить и сделать небольшое отверстие с помощью микро-дрель.
- Загрузите 400 нл вируса в шприц 10 Л, установленный на стереотаксическом аппарате с помощью точного насоса.
- Нижняя игла (34 G, слежена) тщательно и вводить 300 nL Cre-зависимых оптогенетического вируса(AAV5-EF1a-DIO-ChR2 (H134)-eYFP 5,6 х 1012 vg/mL) в VTA (AP: -3.45 мм, ML: -0,2 мм от брегмы и -4,4 мм от поверхности черепа, в соответствии с Франклином и Paxinos22) на 100 nL/min скорость инъекций с помощью точного насоса.
- После инъекции оставьте иглу на месте еще 10 мин, чтобы обеспечить диффузию вируса(рисунок 2А).
- Медленно вытяните иглу из места инъекции.
- Сделайте небольшие отверстия с помощью микродрейля, чтобы соответствовать якорные винты, которые стабилизируют оптическое волокно и стоматологический цементный комплекс.
- Возьмите брегма координаты снова и имплантировать оптическое волокно (200 мкм диаметр, 0,37 NA) по адресу: AP: -3,45 мм, ML: -0,2 мм от брегмы и -4,0 мм от поверхности черепа (Рисунок 2B) в соответствии с Франклин и Paxinos22.
- Защищайте волокно на черепе с помощью зубного цемента. Применить достаточно цемента вокруг оптического волокна ferule, чтобы закрепить его на череп, но обратите внимание, чтобы оставить 3'4 мм верхней части ferule свободной от цемента, чтобы подключение патч шнура (Рисунок 2C).
ПРИМЕЧАНИЕ: Обратите внимание, чтобы не заполнить отверстие с цементом, как это может привести к повреждению тканей мозга. Гемостатические материалы могут быть добавлены в отверстие, чтобы предотвратить это. - Используйте клей ткани или усваиваемые швы, чтобы закрыть любую открытую рану и оставить животное, чтобы восстановить, по крайней мере две недели. Дайте дополнительную дозу анальгетика 12-24 ч после операции.
3. Настройка управления лазерным источником
- Используйте микроконтроллеры одной платы для управления лазерным источником. Напишите сценарий с помощью соответствующего программного обеспечения. Загрузите скрипт на доску микроконтроллера, используя соответствующий кабель подключения к компьютеру.
ПРИМЕЧАНИЕ: Скрипт должен включать внешнюю модуляцию (ввод) из программного обеспечения отслеживания через коробку TTL, и выход на лазер для управления параметрами стимуляции. Для 10 мс шириной импульса на частоте 20 Гц, используйте сценарий, найденный в дополнительном файле кодирования. - Подключите доску к лазеру и tTL-коробке слежения.
- Используйте сетевой кабель для подключения окна TTL к доске (контакт 5 для предоставленного сценария)(рисунок 3A,C).
- Убедитесь, что лазер настроен на управление внешней модуляцией и подключить лазер к доске с помощью кабельного кабеля FC/PC (контакт 13 для данного сценария) (Рисунок 3B, C).
- Подключите соответствующие штыри к наземным частям доски.
- Подключите лазерный источник к оптическому волокну.
- Подключите лазерный источник к роторном суставу(рисунок 3D).
- Подключите патч шнур(Рисунок 3E) к роторный сустав.
- Стабилизировать роторный сустав над аппаратом, но за пределами зоны записи. Убедитесь, что длина волоконно-оптического патч шнура является целесообразным, чтобы позволить мыши двигаться без трудностей на арене(Рисунок 3F).
4. Настройка эксперимента для подхода RT-PP в рамках программного обеспечения для отслеживания
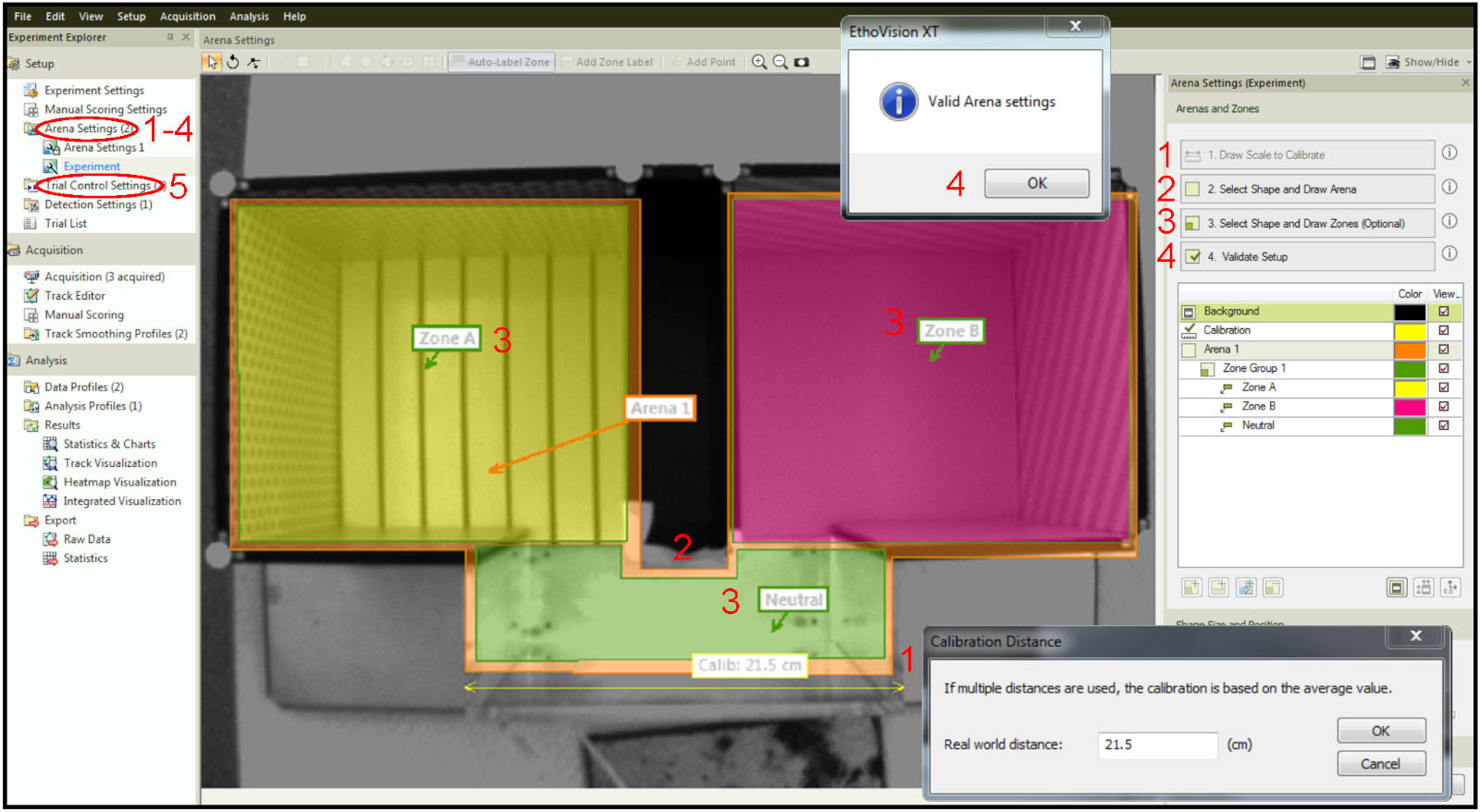
- Откалибровать установку арены. Используйте линейку для измерения определенной части физического аппарата, нарисуйте линию, соответствующую части, измеренной на изображении в программном обеспечении под шкалой рисования, для калибровки и введите уже известное значение (шаг 1 на рисунке 4).
- Дизайн арены. Нарисуйте область, где движение мышей будет записано (шаг 2 на рисунке 4).
- Создайте зоны. Нарисуйте зоны, которые в конечном итоге будут назначены в качестве лазерных парных, лазерных и "нейтральных" (шаг 3 на рисунке 4).
- Проверка настройки, чтобы подтвердить, что нет никаких противоречивых параметров, например зон за пределами арены (шаг 4 на рисунке 4)
- Установите экспериментальные параметры под настройками управления пробным производным (шаг 5 на рисунке 4).
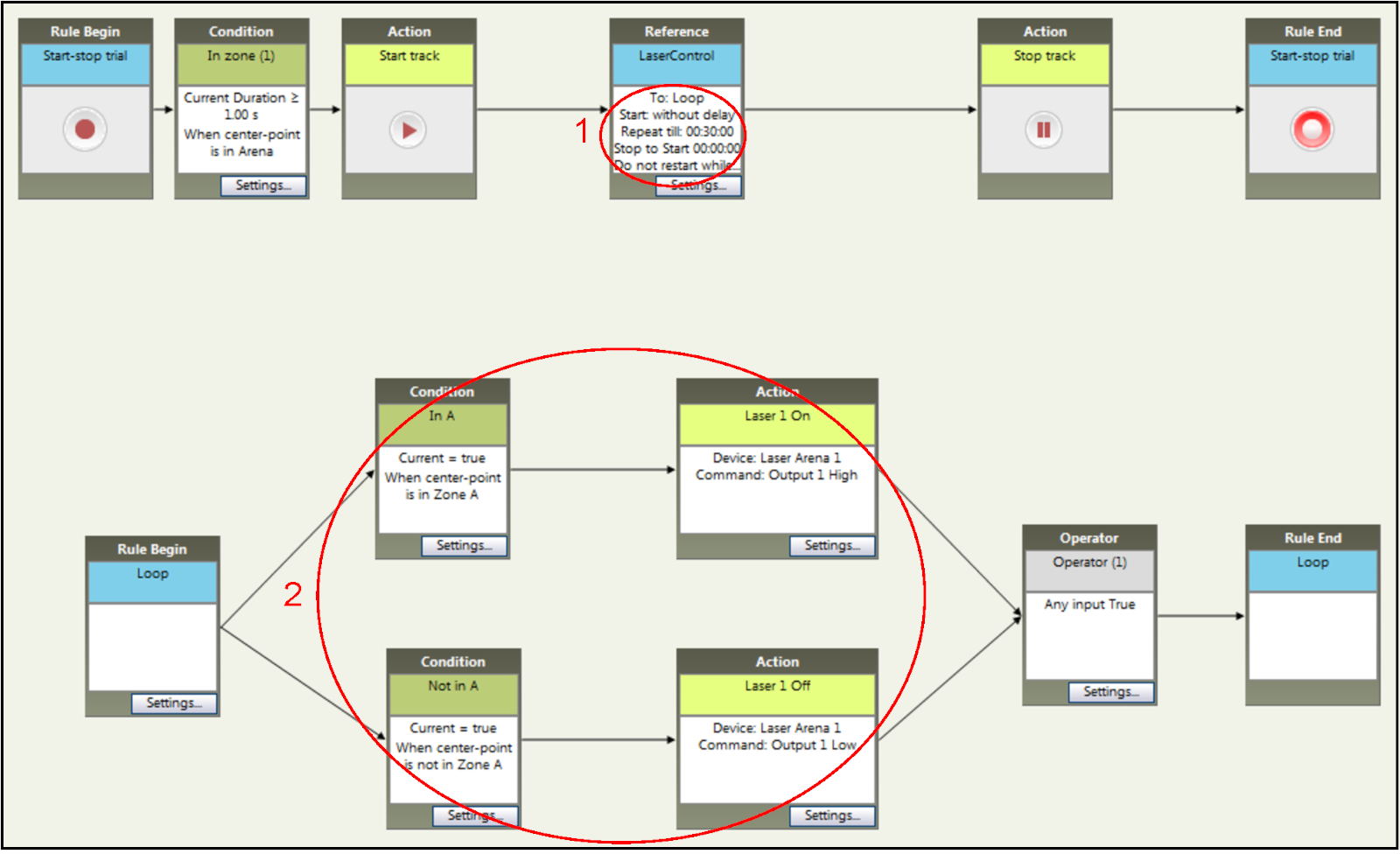
- Установите время испытания, как показано на шаг 1 в рисунке 5 для 30 мин RT-PP сессии.
- Убедившись, что "аппаратный контроль" включен, назначить отсек, как лазер-парной, в которой вход мыши вызовет сигнал TTL через программное обеспечение отслеживания на доске микроконтроллера. На рисунке 5 (шаг 2) отсек с лазерным сопряжением представляет собой отсек А. Для фазы разворота, переключатель отсеков так отсек B будет лазером в паре и отсек А будет неспаренным. Сделайте это, заменив A с B и B с A в программном обеспечении.
5. Модификация установки для проверки аверсивных свойств стимуляции с использованием подхода NCP
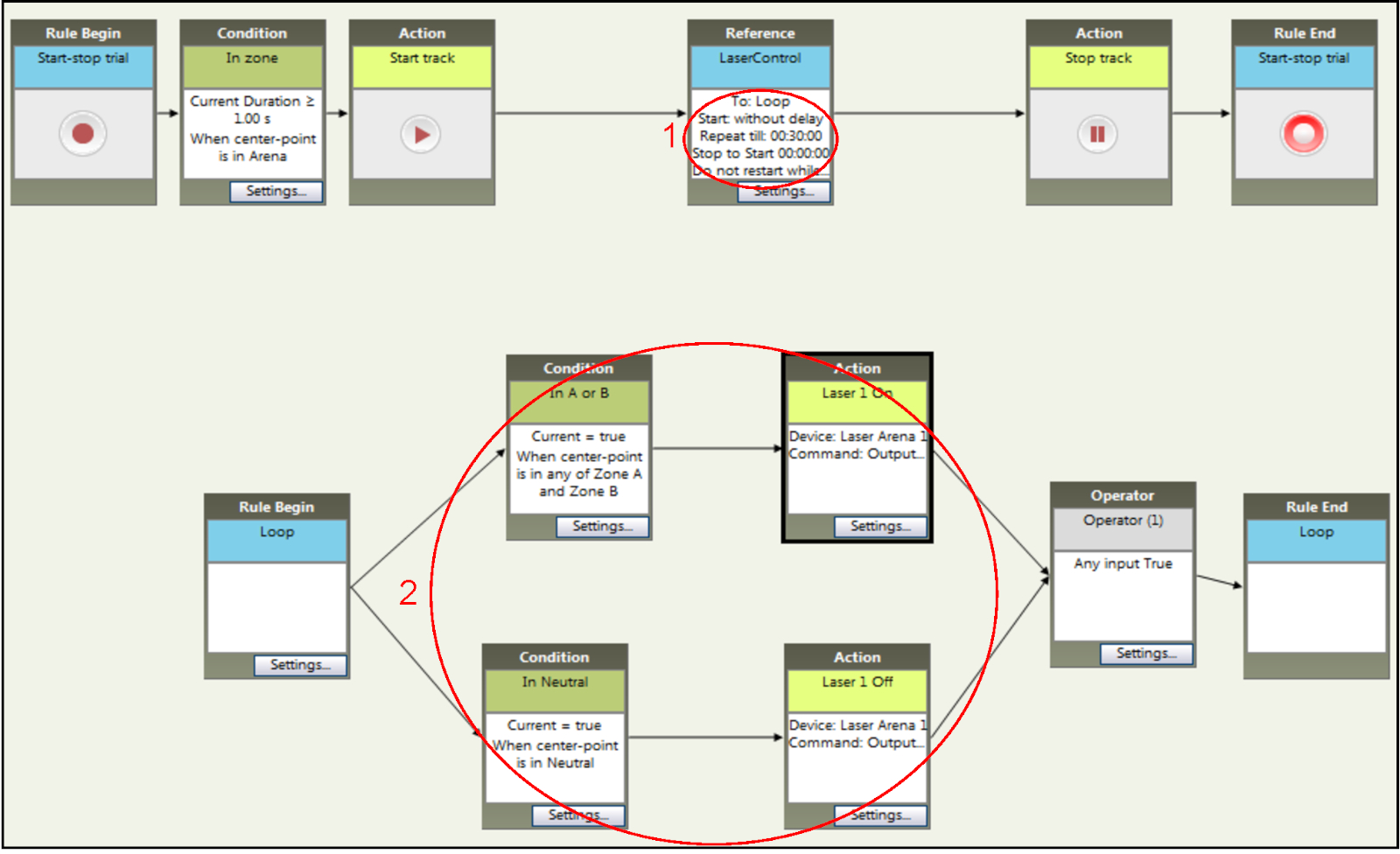
- Следуйте шагам 4.1 к 4.4,4, как описано ранее.
- Установите время эксперимента до 30 мин через опцию "Повторить до" в настройках коробки "Reference" (шаг 1 на рисунке 6).
- Назначьте как зоны A и B в паре (шаг 2 на рисунке 6), добавив "когда центральная точка находится в любой из зоны А и зоны B" для окна состояния, связанного с настройками для отсеков A и B. Обратите внимание, что лазер выключится, когда животное находится в нейтральном отсеке.
6. Проведение эксперимента с использованием лазерной стимуляции
- Настройка настроек обнаружения.
- Используйте манекен, чтобы напоминать мышь, чтобы обеспечить соответствующие настройки обнаружения.
- Поместите манекен в один отсек аппарата и используйте автоматизированную установку с динамическим вычитанием.
- Снимите манекен и поместите его в противоположном отсеке. Убедитесь, что манекен полностью обнаружен, и если нет, настроить настройки с помощью программного обеспечения для достижения надлежащего обнаружения.
- Во время этого шага также проверить, если стимуляция работает, как это должно. Начните приобретение с использованием ранее настроенных параметров пробного управления и поместите манекен в отсек для парных лазеров и посмотреть, если стимуляция срабатывает как следует. Затем поместите манекен в неспаренный и / или нейтральный отсек и посмотреть, если стимуляция остановлена.
- Используйте счетчик мощности с датчиком, чтобы установить мощность лазера до 10 мВт с помощью ручки на лазере(рисунок 3B). Выполняйте этот шаг каждый раз, когда используется лазерная стимуляция.
ВНИМАНИЕ: Используйте защитное оборудование для глаз, так как прямое воздействие лазерного света может привести к необратимому повреждению глаз. - Поместите мышь в аппарат.
- Аккуратно выняйте мышь из клетки и подключите волоконно-оптический имплантат к волоконно-оптическому патч-пуповину с помощью керамического рукава.
- Поместите мышь осторожно в нейтральном отсеке трехотсекного аппарата.
- Подождите, пока мышь не будет обнаружена программным обеспечением.
- Удалите вертикальные раздвижные двери, ограничивающие попадание животного в основные отсеки.
- Разрешить животному свободно исследовать без каких-либо нарушений.
ПРИМЕЧАНИЕ: Та же процедура соблюдается, когда животное не получает стимуляции, за исключением того, что шаг 6.2 не нужен, и что лазер выключен все время.
Результаты
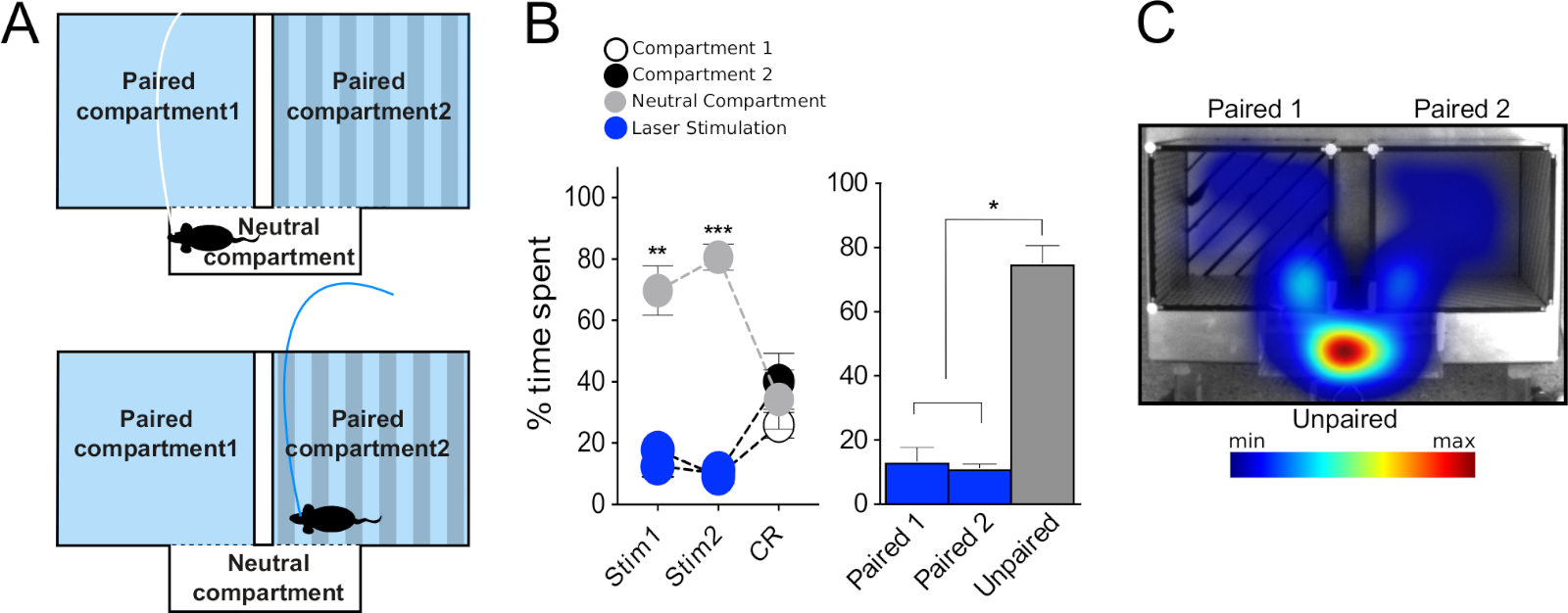
Трехкомпонентный аппарат(рисунок 3F) подходит для устранения эффектов, принося имеющее сярприг, и для оценки в режиме реального времени полезных или аверсивных свойств прямой стимуляции нейронов с помощью оптогенетики. Он состоит из двух основных отсеков (20 см (W) х 18 см (L) х 25 см (H) и одного меньшего соединительного отсека (20 см (20 см (W) х 7 см (L) x 25 см (H). Основные отсеки имеют различные текстуры стены и пола и узоры для того, чтобы облегчить ассоциативное обучение, в то время как соединительное/нейтральное отделение является узким и прозрачным, поэтому мыши проводят в нем естественно меньше времени. Как описано выше, программное обеспечение для отслеживания может быть использовано для записи нескольких поведенческих параметров мышей, включая движение и время, проведенное в каждом отсеке, и для управления лазерной стимуляции. Весь эксперимент RT-PP проходит в течение 8 сессий(рисунок 1А)и позволяет как оценивать полезные или отрицательные свойства прямой стимуляции (дни 3, 4, 6 и 7), так и формирование ассоциаций, положительных или отрицательных, в ответ на предыдущий опыт (дни 5 и 8, "CR").
Во-первых, мы протестировали мышей DAT-Cre, вводили вирус AAV-ChR2-eYFP в VTA для таргетинга дофаминергических нейронов. В соответствии с литературой, мы заметили, что мыши предпочитали проводить время в отсеке в паре со стимуляцией(Рисунок 1B, фаза 1, дни 3 и 4, синие круги, двусторонний повторный анализ «RM» анализа дисперсии «ANOVA», эффект отсека F (2,18) 141, p slt; 0.001; эффект сеанса x отсека F(12,108) Tukey после hoc испытания в паре против unpaired р злт; 0,001). В фазе разворота, где сопряжение отсеков с лазерной стимуляцией было переключено, подтвердило эти наблюдения(рисунок 1B, дни 6 и 7, синие круги, пост-специальный тест Туки в паре против непарного отсека р злт; 0.001), таким образом исключая возможность того, что результаты, полученные от фазы 1, были связаны с боковыми смещениями или случайными параметрами. Усреднение времени, проведенного в каждом отсеке в течение четырех дней RT-PP подтвердило, что мыши провели в среднем около 70% своего времени в лазерном парном отсеке, в отличие от непарного (20%) и нейтральные (10%) отсеки(Рисунок 1B бар график, односторонний RM ANOVA, эффект стимуляции F(2,6) 139, р lt; 0,001, Tukey должность, как в паре против непарных и нейтральных отсеков р/л; 0,001). Кроме того, в отсутствие стимуляции, в дни 5 и 8, мыши провели значительно больше времени в отсеке ранее в паре с лазерной стимуляции (Tukey после hoc p qlt; 0.001), указывая, что предыдущий опыт был достаточен, чтобы вызвать ассоциативное поведение обучения отражается как "поиск" стимуляции. Эти данные соответствуют литературе и показывают, что нынешний метод может быть надежно использован для изучения полезных эффектов оптогенетической стимуляции конкретных нейронных популяций в ВТА.
Затем мы протестировали мышей VGLUT2-Cre, впрыснутых с помощью AAV-ChR2-eYFP в VTA, как указано выше, для целевой глутаматергических нейронов VTA. В этом эксперименте мы наблюдали противоположный поведенческий фенотип от того, который продемонстрировали мыши DAT-Cre. Таким образом, мыши избегали отсека в паре со стимуляцией и проводили больше времени в неспаренных в течение всех дней RT-PP(рисунок 1C слева, двусторонний RM ANOVA, эффект отсека F(2,12) 40,9, р-л; 0.001; эффект сеанса x отсекаF (12,72) Tukey после hoc испытания в паре против unpaired р Злт; 0,001; Рисунок 1C право, односторонний RM ANOVA эффект стимуляции F(2,6) 162, р-л; 0,001, Tukey после hoc в паре против непарных и нейтральных отсеков р злт; 0,001). Интересно, что во время«C»дней 5 и 8, мыши не показали явного избегания ранее парного отсека (никаких различий между парными и непарными отсеками). Вполне возможно, что отсутствие условной реакции связано с недостаточным временем, проведенным в лазерном парном отсеке, что помешало образованию связей между лазерной активацией и конкретной средой, где это произошло. Для дальнейшего изучения этого фенотипа избежания мы использовали измененный протокол, который мы назвали «нейтральным предпочтением отсека», сокращенно NCP. В этом эксперименте оба основных отсека были сопряжены со стимуляцией, а нейтральный отсек оставался свободным от стимуляции(рисунок 7А). Мы предположили, что, если стимуляция обладает аверсивными свойствами, то мышь будет вынуждена проводить время в меньшем, нейтральном отсеке, чтобы избежать этого. Действительно, в оба дня стимуляции (Stim1 и Stim2) мыши провели большую часть времени в нейтральном отсеке (около 80%) по сравнению с парными отсеками(рисунок 7B, C;слева: двусторонний эффект RM ANOVA отсека F(2,8) 70,9, стр. 0.001; эффект сеанса x-купе F(4,16) 6,9, стр. 0,002, пост-стимуляция Туки 1 "Стимулирование 1" нейтрального отсека против отсека 1 и 2 р/л; 0,01, "Стимулирование 2" нейтральный отсек против купе 1 и 2 р.т; 0.018, Tukey's post hoc test paired 1 and 2 vs neutral p zlt; 0.05). Как и в случае с«CR»во время теста RT-PP, мыши, казалось, не образовывались в негативной связи между отсеками и стимуляцией; то есть, при отсутствии стимуляции (CR), они исследовали все отсеки в той же степени(рисунок 7B, нет различий между временем, проведенным в парных отсеках и нейтральном отсеке). Результаты этих экспериментов подтвердили поведенческий фенотип, наблюдаемый во время установки RT-PP, и тем самым поддерживают комбинаторную реализацию парадигм RT-PP и NCP.

Рисунок 1: Поведенческое тестирование с использованием оптогенетики в парадигме RT-PP. (A) Схематическое представление временной шкалы экспериментов. (B, C) Вверху слева: график, представляющий процент времени, затраченного в каждом отсеке на протяжении всего эксперимента RT-PP для DAT-Cre (N No 10) и VGLUT2-Cre (N no 7) мышей, введенных с помощью AAV-ChR2-eYFP. Синие круги: отсек с лазерной парой; белые, черные круги: основные отсеки; серые круги: нейтральный отсек. Вверху справа: средний процент времени, проведенного в каждом отсеке в течение дней 3, 4, 6 и 7 (RT-PP). Внизу: репрезентативные тепловые карты времени, проведенного в каждом отсеке для DAT-Cre и мыши VGLUT2-Cre. Все данные, как правило, распределены (тест Шапиро-Вилк). Результаты представлены в виде среднего значения - SEM. -стр. 0,05, п.л.; 0,01,п/д; 0,001 парный и нейтральный отсек; Р-л; 0,01, п.л.; 0,001 непарного и нейтрального отсека. Эта цифра была изменена с Bimpisidis и др.12. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: Хирургическая процедура для оптогенетических экспериментов. (A) Инъекция cre-зависимого вирусного вектора в VTA. (B) Имплантация оптического волокна над местом инъекции. Обратите внимание на якорные винты, используемые для стабилизации. (C) Постоянное закрепление волокна на черепе с использованием зубного цемента. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 3: Оборудование, используемое в экспериментах по оптогенетике. (A) Коробка TTL, используемая в исследованиях. Он получает входные данные от программного обеспечения отслеживания и посылает сигналы TTL на доску микроконтроллера. (B) Передний (вверху) и задний вид (внизу) лазерного источника, используемого для экспериментов. (C) Доска микроконтроллера, используемая для управления лазерной стимуляцией. Обратите внимание на соединения из коробки TTL и к источнику лазера. (D) Ротари сустава. (E) Волоконно-оптический патч аккорд, используемый в экспериментах. (F) Трехкомпонентный аппарат, используемый для экспериментов RT-PP и NCP. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 4: Проектирование арены и зон в рамках программного обеспечения для отслеживания. Шаг 1: Калибровка установки. Шаг 2: Рисунок всей арены. Шаг 3: Зоны рисования на арене. Шаг 4: Настройка проверки. Шаг 5: вкладка настройки trial Control для настройки параметров времени и стимуляции. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 5: Настройка параметров времени и стимуляции эксперимента RT-PP в рамках программного обеспечения для отслеживания. Добавление конкретных правил на продолжительность (шаг 1) и условия для стимуляции света (шаг 2). Условия могут быть легко изменены в соответствии с требованиями для фазы разворота. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 6: Настройка параметров времени и стимуляции экспериментов NCP в рамках программного обеспечения для отслеживания. Продолжительность сеансов стимуляции (шаг 1) аналогична продолжительности RT-PP, но условия для активации световой стимуляции (шаг 2) различны. Вход в основной отсек (здесь названная зона А и зона B) приводит к оптогенетической стимуляции, которая прекращается только тогда, когда мышь входит в нейтральный отсек. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 7: Поведенческое тестирование с использованием оптогенетики в парадигме NCP. (A) Схематическое представление экспериментальной установки. (B) Слева: график, представляющий процент времени, проведенного в каждом отсеке в течение двух дней стимуляции (Stim1 и Stim 2) и во время условного ответа (CR) сессии для VGLUT2-Cre мышей вводили с AAV-ChR2 в VTA (N No 5). Справа: средний процент времени, проведенного в каждом отсеке в течение двух дней стимуляции NCP. (C) Представитель тепловой карты времени, проведенного в каждом отсеке для мыши VGLUT2-Cre в течение одного из дней стимуляции. Данные обычно распространялись (тест Шапиро-Вилк). Результаты представлены в виде среднего значения : SEM. Эта цифра была изменена с Bimpisidis и др.12. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
| Шаг | Температура | Длительность | Циклов |
| 1. Первоначальная денатурация | 95 кв. c | 4 мин. | 1 |
| 2. Денатурация | 95 кв. c | 30 с | 30 |
| 3. Аннеалинг | 55 кк с | 30 с | |
| 4. Продление | 72 кк с | 40 с | |
| 5. Окончательное продление | 72 кк с | 6 мин. | 1 |
| 6. Удерживайте | 4 кк с | До тех пор, пока остановился экспериментатор | 1 |
Таблица 1: Программа велосипедного движения PCR.
Дополнительный файл кодирования. Пожалуйста, нажмите здесь, чтобы просмотреть этот файл (Право нажмите, чтобы скачать).
Обсуждение
В текущем исследовании мы представляем два пошаговых протокола о том, как выполнять различные типы анализа предпочтений местом с использованием оптогенетики у мышей. Протоколы были использованы для оценки награждения или аверсивных поведенческих фенотипов нейронов VTA(Рисунок 1 и Рисунок 6)12, но могут быть использованы для изучения поведенческой роли нейронов в других областях мозга, а также.
Несколько недавних исследований описали RT-PP парадигмы в двух-купе23,24 и трех-отсеков аппаратов13,14,15,16,17,18. В текущих протоколах описываются подробные настройки протоколов RT-PP и NCP в трехкомпонентном аппарате, напоминающем те, которые традиционно используются в экспериментах CPP для оценки поведенческих эффектов при администрировании наркотиков, связанных со злоупотреблением. В то время как результаты представлены только здесь как процент времени, проведенного мышью в каждом отсеке, программное обеспечение отслеживания позволяет анализировать несколько других поведенческих параметров, таких как переходы в зоны, скорость, время, проведенное неподвижно и многое другое. Анализ различных параметров может быть важен для интерпретации данных.
Текущие протоколы RT-PP являются гибкими и могут быть изменены для проверки, имеют ли различные типы моделей стимуляции полезные эффекты. Параметры лазерного управления можно легко изменить либо через скрипт доски микроконтроллера, либо внутри программного обеспечения для отслеживания, демонстрируя универсальность установки. Мы предлагаем 20 Гц частоты стимуляции, которая находится в пределах диапазона, а иногда и ниже, частот, применяемых в предыдущих исследованиях с использованием того же варианта опсина (ChR2/H134R) для изучения дофаминергических и глутамергических нейронов и их терминалов13,14,16,17,18,23,24,25,26. Недавние исследования показали, что более высокие частоты стимуляции могут иметь противоположное влияние на поведение, чем более низкие, и что эти эффекты опосредованы через блок деполяризации, вызванные более высокими частотами28. Аналогичным образом, различия в поведенческих выход были показаны при стимулировании глутамерных и ГАМК нейронов в боковой преоптической области15. Эти исследования изучили нейроны различных областей, чем VTA и наибольшие эффекты наблюдались на высоких частотах не-глутаматергических нейронов15,28. Наш выбор на 20 Гц основан на предыдущих исследованиях глутамергических и дофаминергических нейронов VTA, демонстрирующих, что при различных частотах стимуляции, связанных с вознаграждением поведенческих выход не значительно изменен24,26.
Дополнительным параметром, который может быть скорректирован и который может повлиять на экспериментальный результат, является сила источника света. Более высокая мощность лазера может увеличить размер светостимулируемой области, что может быть полезным в некоторых видах экспериментов, но с недостатком повышения температуры5. Действительно, недавнее исследование показало, что лазерно-индуцированное повышение температуры может изменить физиологию мозга и повлиять на поведенческие измерения29. Эти наблюдения подчеркивают важность включения opsin-негативного контроля в экспериментальном дизайне. В текущем протоколе, мы использовали 10 мВт лазерной мощности, которая аналогична и ранее было показано, чтобы быть эффективным в стимулировании дофаминергических и глутаматергических нейронов в VTA16,24,26. При организации экспериментов важно обращать внимание на размер области, в которой расположены интересные клетки, а также на свойства волоконно-оптического и патч-провода (числовая диафрагма, диаметр ядра). Эти параметры необходимы для учета при выполнении расчетов, связанных с лазерной мощностью. Для получения подробной информации можно использовать калькулятор, разработанный лабораторией Карла Дейссерота(http://web.stanford.edu/group/dlab/cgi-bin/graph/chart.php).
Гистологическая проверка рекомбинации Cre-Lox является еще одним критическим аспектом при применении оптогенетических экспериментов. Проверка эффективности рекомбинации всегда должна проходить в пилотной когорте до начала любых поведенческих экспериментов в большой группе животных. Это важно не только по этическим соображениям, но и по оптимизированной экспериментальной продукции. Каждая вирусная конструкция может показать переменную специфичность для различных типов нейронов и в различных регионах5, параметр, который может повлиять на эксперименты в непредсказуемых и даже вводящих в заблуждение способов. Например, мы ранее подтвердили рекомбинацию вирусов AAV5 в VTA мышей DAT-Cre и обнаружили, что односторонних инъекций было достаточно для того, чтобы нацелить большую часть интересуемых областей. Когда мы затем изучали пространственно ограниченных субпопуляций в рамках VTA, таких, как один характеризуется NeuroD6 выражение, мы заметили, что двусторонние вирусные инъекции были более эффективными для целевой большее число нейронов дает более выраженные поведенческие эффекты на оптогенетической свет-стимуляции12. Кроме того, время от операции до начала поведенческих экспериментов должно быть рассмотрено тщательно. Две недели достаточно времени для конструкции ДНК ChR2, чтобы быть выражены в клеточных телах, как мы показываем здесь, но больше времени ожидания (8 недель) может потребоваться, если следователь тестирует эффект стимуляции в проекционных областях13,14,15,17.
Стоит отметить, что объем вводимых вирусов (в нашем случае 300 нл) может быть подходящим при изучении нейронов в ВТА, но объем и титр должны быть скорректированы в зависимости от эффективности трансдукции и размера исследуемой структуры. Кроме того, для двусторонних структур, расположенных на расстоянии от посредствероальной оси, может потребоваться проведение двусторонних инъекций, а также для двусторонней имплантации волоконно-оптических веществ для обеспечения активации/ингибирования в обоих полушариях.
Наконец, всегда необходимо провести посмертный гистологический анализ, чтобы подтвердить и подтвердить эффективность рекомбинации Cre-Lox и проверить правильное место имплантации оптического волокна в нужном месте. Неожиданные, чрезмерно ограниченные или чрезмерные Cre-Lox рекомбинации может произойти из-за неизвестного распределения нейронов, выражающих Cre за пределами предполагаемой области, или из-за различий в вирус серотип, плохое обращение с вирусом, засорение шприц для доставки вируса или других проблем, связанных с хирургическим вмешательством. Проверка удовлетворительной рекомбинации Cre-Lox и правильной волоконно-оптической имплантации должна быть проведена для подтверждения любых статистических результатов поведенческих оценок для того, чтобы сделать безопасные выводы.
С точки зрения данных, представленных здесь в качестве примеров того, как две поведенческие парадигмы могут быть использованы, значительное предпочтение свето-парной стороне, полученной оптогенетической стимуляции дофаминергических нейронов в VTA путем анализа DAT-Cre мышей в RT-PP парадигмы, как ожидается, на основе предыдущих выводов23,25,27,27 в то время как избегание этой стороны показали ВТ-2. VGLUT2 нейронов VTA и их прогнозы были показаны, чтобы быть вовлечены в награду и отвращение16,17,24,30,31, и поэтому мы провели анализ NCP для оценки явного поведения избегания наблюдается в текущей установки RT-PP более подробно. Используя узкий, прозрачный коридор в качестве единственного нелегкого парного отсека для подтверждения аверсивных свойств стимуляции глутаметатергических нейронов VTA, очевидно, что в данном конкретном трехкомпонентном установке оптогенетическая активация этих нейронов вызывает аверсивную реакцию. Эти эксперименты, которые были показаны здесь, чтобы иллюстрировать ситуации, которые могут извлечь выгоду из использования как RT-PP и NCP протоколов, были частью недавно опубликованного исследования, и полный набор данных, а также обсуждения в отношении этих выводов можно найти в этой публикации12.
В дополнение к NCP, альтернативные способы подтверждения отвращения включают сильное освещение области в открытой области поля при сопряжении остальной части арены для лазерной активации, или выполнять активную задачу избежания, в которой мышь должна выполнить определенную модель поведения, чтобы прекратить лазерную стимуляцию15.
Подводя итог, описанные протоколы предоставляют важную информацию о том, как успешно выполнять анализ RT-PP и NCP наиболее эффективным способом, чтобы разгадать роль активации нейронов в вознаграждении и отвращение. В зависимости от научной гипотезы, с помощью этих протоколов можно проанализировать ряд параметров, и каждый протокол также может быть объединен с другими проверенными парадигмами для оптимизированного поведенческого анализа, реализующего оптогенетику для решения конкретного мозга областей и нейронов, представляющих интерес.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Наши источники финансирования с благодарностью признаются: Университет Уппсалы, Ветенскапсрет (Шведский исследовательский совет), Хьёрнфонден, Паркинсонфонд, Исследовательские фонды Бертила Хёллстена, ОЭ и Эдла Иоханссон, Зоологиск Форскнинг и Элен. В Уппсалском университете содержались животные, а в поведенческом центре Университета Уппсалы проводились эксперименты.
Материалы
| Name | Company | Catalog Number | Comments |
| AAV-Cre dependent virus | UNC Vector Core | - | a great variety of viruses to suit any project's needs |
| Agarose | VWR Life Science | 443666A | |
| Buffer for PCR | KAPA BIOSYSTEMS | KB1003 | 10x, contains 1.5mM MgCl2 at 1x |
| Bupivacaine (Marcain) | Aspen | N01BB01 | local anesthetic, 5 mg/ml solution, requires prescription |
| Carprofen (Norocarp) | N-Vet | 27636 | anti-inflammatory, analgesic; 50 mg/ml solution, requires prescription |
| dNTP set | Thermo Fisher Scientific | R0181 | 100mM, have to be dilluted to 10mM and mixed |
| DNA ladder | Thermo Fisher Scientific | SM0243 | 100 bp, 50 μg Gene Ruler |
| DNA loading dye | Thermo Fisher Scientific | R0611 | 6x, dilute to 1x before using |
| Ear puncher | AgnThos | AT7000 | ear puncher to take tissue samples for PCR or to mark animals |
| Fiberoptic patchcords | Doric Lenses | MFP_200/240/900-0.22_1m_FC-ZF1.25 | |
| Implantable fiberoptics | Doric Lenses | MFC_200/245-0.37_5mm_ZF1.25_FLT | the properties of the fibers might change depending on the experiment |
| Infusion pump for virus injections | AgnThos | Legato 130 | contains remote pump to secure the syringe directly on the stereotexi frame |
| Isoflurane (Forane) | Baxter | N01AB06 | Volatile anesthetic, requires prescription |
| Jewelry screws | AgnThos | MCS1x2 | |
| Laser source | Marwell Laser Systems | CNI MBL-III-473-100mW | |
| Microcontroller board | Arduino | "Uno" board | can be ordered from several companies |
| Microdrill | AgnThos | 1474 | could be ordered with or without stereotaxic holder |
| Needle (34G) | World Precision Instruments | NF36BV | |
| Nucleic Acid gel stain - GelRed | Biotium | 41003-T | |
| PCR tubes | Axygen | PCR-0208-CP-C | |
| Power meter | Thorlabs | PM100D | |
| Place Preference Apparatus | Panlab | 76-0278 | |
| Rotary joint | Doric Lenses | FRJ_1x1_FC-FC | |
| Sleeves | Doric Lenses | SLEEVE_ZR_1-25 | mating sleeve to connect the patchcord with the implanted optic fiber |
| Stabilization cement | Ivoclar Vivadent | Tetric EvoFlow | |
| Syringe (10ul) | World Precision Instruments | NanoFil | |
| Taq polymerase | KAPA BIOSYSTEMS | KE1000 | 500U |
| TAE buffer | Omega BIO-TEK | SKU:AC10089 | 50x concentration, has to be dilluted before use |
| Thermal cycler | BIO-RAD S1000 | 1852148 | necessary to perfrom PCR reactions |
| Tissue glue | AgnThos | Vetbond | |
| Tracking software | Noldus | Ethovision XT | |
| TTL box | Noldus | Noldus USB-IO box | |
| UV transilluminator | Azure Biosystems | c200 model |
Ссылки
- Fenno, L., Yizhar, O., Deisseroth, K. The Development and Application of Optogenetics. Annual Review of Neuroscience. 34 (1), 389-412 (2011).
- Kim, C. K., Adhikari, A., Deisseroth, K. Integration of optogenetics with complementary methodologies in systems neuroscience. Nature Reviews Neuroscience. 18 (4), 222-235 (2017).
- Deisseroth, K. Optogenetics: 10 years of microbial opsins in neuroscience. Nature Neuroscience. 18 (9), 1213-1225 (2015).
- Nagy, A. Cre recombinase: The universal reagent for genome tailoring. Genesis. 26 (2), 99-109 (2000).
- Yizhar, O., Fenno, L. E., Davidson, T. J., Mogri, M., Deisseroth, K. Optogenetics in Neural Systems. Neuron. 71 (1), 9-34 (2011).
- Pupe, S., Wallén-Mackenzie, &. #. 1. 9. 7. ;. Cre-driven optogenetics in the heterogeneous genetic panorama of the VTA. Trends in Neurosciences. 38 (6), 375-386 (2015).
- Spanagel, R. Animal models of addiction. Dialogues in Clinical Neuroscience. 19 (3), 247-258 (2017).
- Tzschentke, T. M. Measuring reward with the conditioned place preference (CPP) paradigm: Update of the last decade. Addiction Biology. 12 (3-4), 227 (2007).
- Hoffman, D. C. The use of place conditioning in studying the neuropharmacology of drug reinforcement. Brain Research Bulletin. 23 (4-5), 373-387 (1989).
- Huston, J. P., Silva, M. A. D. S., Topic, B., Müller, C. P. What's conditioned in conditioned place preference?. Trends in Pharmacological Sciences. 34 (3), 162-166 (2013).
- Bardo, M. T., Bevins, R. A. Conditioned place preference: What does it add to our preclinical understanding of drug reward?. Psychopharmacology. 153 (1), 31-43 (2000).
- Bimpisidis, Z., et al. The NeuroD6 subtype of VTA neurons contributes to psychostimulant sensitization and behavioral reinforcement. eNeuro. 6 (3), e0066 (2019).
- Root, D. H., Mejias-Aponte, C. A., Qi, J., Morales, M. Role of Glutamatergic Projections from Ventral Tegmental Area to Lateral Habenula in Aversive Conditioning. Journal of Neuroscience. 34 (42), 13906-13910 (2014).
- Steidl, S., Wang, H., Ordonez, M., Zhang, S., Morales, M. Optogenetic excitation in the ventral tegmental area of glutamatergic or cholinergic inputs from the laterodorsal tegmental area drives reward. European Journal of Neuroscience. 45 (4), 559-571 (2017).
- Barker, D. J., et al. Lateral Preoptic Control of the Lateral Habenula through Convergent Glutamate and GABA Transmission. Cell Reports. 21 (7), 1757-1769 (2017).
- Wang, H. L., Qi, J., Zhang, S., Wang, H., Morales, M. Rewarding Effects of Optical Stimulation of Ventral Tegmental Area Glutamatergic Neurons. The Journal of Neuroscience. 35 (48), 15948-15954 (2015).
- Qi, J., et al. VTA glutamatergic inputs to nucleus accumbens drive aversion by acting on GABAergic interneurons. Nature Neuroscience. 19, 725-733 (2016).
- Qi, J., et al. A glutamatergic reward input from the dorsal raphe to ventral tegmental area dopamine neurons. Nature Communications. 5, 5390 (2014).
- Ekstrand, M. I., et al. Progressive parkinsonism in mice with respiratory-chain-deficient dopamine neurons. Proceedings of the National Academy of Sciences of the United States of America. 104 (4), 1325-1330 (2007).
- Borgius, L., Restrepo, C. E., Leao, R. N., Saleh, N., Kiehn, O. A transgenic mouse line for molecular genetic analysis of excitatory glutamatergic neurons. Molecular and Cellular Neuroscience. 45 (3), 245-257 (2010).
- Papathanou, M., et al. Targeting VGLUT2 in Mature Dopamine Neurons Decreases Mesoaccumbal Glutamatergic Transmission and Identifies a Role for Glutamate Co-release in Synaptic Plasticity by Increasing Baseline AMPA/NMDA Ratio. Frontiers in Neural Circuits. 12, 64 (2018).
- Franklin, K. B. J., Paxinos, G. . The mouse brain in stereotaxic coordinates. , (2008).
- Tsai, H. C., et al. Phasic firing in dopaminergic neurons is sufficient for behavioral conditioning. Science. 324 (5930), 1080-1084 (2009).
- Yoo, J. H., et al. Ventral tegmental area glutamate neurons co-release GABA and promote positive reinforcement. Nature Communications. 7, 1-13 (2016).
- Pascoli, V., Terrier, J., Hiver, A., Lüscher, C. Sufficiency of Mesolimbic Dopamine Neuron Stimulation for the Progression to Addiction. Neuron. 88 (5), 1054-1066 (2015).
- Ilango, A., Kesner, A. J., Broker, C. J., Wang, D. V., Ikemoto, S. Phasic excitation of ventral tegmental dopamine neurons potentiates the initiation of conditioned approach behavior: parametric and reinforcement-schedule analyses. Frontiers in Behavioral Neuroscience. 8, 155 (2014).
- Kim, K. M., et al. Optogenetic mimicry of the transient activation of dopamine neurons by natural reward is sufficient for operant reinforcement. PLoS ONE. 7 (4), 1-8 (2012).
- Kroeger, D., et al. Galanin neurons in the ventrolateral preoptic area promote sleep and heat loss in mice. Nature Communications. 9, 4129 (2018).
- Owen, S. F., Liu, M. H., Kreitzer, A. C. Thermal constraints on in vivo optogenetic manipulations. Nature Neuroscience. 22 (7), 1061-1065 (2019).
- Root, D. H., Estrin, D. J., Morales, M. Aversion or Salience Signaling by Ventral Tegmental Area Glutamate Neurons. iScience. 2, 51-62 (2018).
- Lammel, S., et al. Diversity of transgenic mouse models for selective targeting of midbrain dopamine neurons. Neuron. 85 (2), 429-438 (2015).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены