
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Bioengineering
Manipolazione di singole cellule staminali neurali e neuroni nelle fette cerebrali utilizzando la microiniezione robotica
Questo protocollo dimostra l'uso di una piattaforma robotica per la microiniezione in singole cellule staminali neurali e neuroni nelle fette cerebrali. Questa tecnica è versatile e offre un metodo per tracciare le cellule nei tessuti ad alta risoluzione spaziale.
Una domanda centrale nella neurobiologia dello sviluppo è come le cellule staminali neurali e progenitrici formano il cervello. Per rispondere a questa domanda, è necessario etichettare, manipolare e seguire singole cellule nel tessuto cerebrale con alta risoluzione nel tempo. Questo compito è estremamente impegnativo a causa della complessità dei tessuti nel cervello. Abbiamo recentemente sviluppato un robot, che guida un ago di microiniezione nel tessuto cerebrale utilizzando immagini acquisite da un microscopio per fornire volumi di soluzione femtolitro in singole cellule. L'operazione robotica aumenta risultando in una resa complessiva che è un ordine di grandezza maggiore della microiniezione manuale e consente un'etichettatura precisa e una manipolazione flessibile di singole cellule nel tessuto vivente. Con questo, si possono microiniettare centinaia di cellule all'interno di una singola fetta organotipico. Questo articolo dimostra l'uso del robot di microiniezione per la microiniezione automatizzata di cellule progenitrici neurali e neuroni nelle fette del tessuto cerebrale. Più in generale, può essere utilizzato su qualsiasi tessuto epiteliale con una superficie che può essere raggiunta dalla pipetta. Una volta impostato, il robot di microiniezione può eseguire 15 o più microiniezioni al minuto. Il robot di microiniezione a causa della sua produttività e versalità renderà la microiniezione una tecnica di manipolazione cellulare ad alte prestazioni ampiamente semplice da utilizzare in bioingegneria, biotecnologia e biofisica per eseguire analisi a cellule singole in fette cerebrali organotipiche.
Questo protocollo descrive l'uso di un robot per colpire e manipolare singole cellule nelle fette del tessuto cerebrale, concentrandosi in particolare su singole cellule staminali neurali e neuroni. Il robot è stato sviluppato per affrontare una questione centrale nella neurobiologia dello sviluppo, cioè come le cellule staminali neurali e progenitrici contribuiscono alla morfogenesi cerebrale1,2,3,4,5. Per rispondere a questa domanda, è necessario etichettare e tracciare singole cellule staminali neurali e seguire la loro progressione del lignaggio nel tempo per correlare il comportamento delle singole cellule con la morfogenesi tissutale. Questo può essere ottenuto in diversi modi, ad esempio elettroporando il tessuto cerebrale in utero o etichettando una singola cellula usando matrici lipofile. Sebbene potenti, questi metodi mancano di una precisa risoluzione a singola cella (elettroporazione) e /o della possibilità di manipolare lo spazio intracellulare (colorante lipofilo). La microiniezione in singole cellule è stata sviluppata per superare questasfida 6,7,8. Durante la microiniezione, una pipetta viene brevemente inserita in una singola cellula all'interno del tessuto intatto sotto pressione per microiniettare volumi femtoliter di reagenti9. In precedenza abbiamo descritto una procedura manuale per la microiniezione di singole cellule staminali neurali nel tessuto organotipico(Figura 1A)10,11. La microiniezione nelle cellule staminali neurali si basa sull'uso di una micropipetta che viene inserita in singole cellule staminali neurali per iniettare una soluzione contenente un colorante fluorescente, insieme ad altre molecole di interesse. Il targeting selettivo delle cellule staminali neurali si ottiene avvicinando al telencefalo in via di sviluppo attraverso la superficie ventricolare (o ventricolare, vedi cartone animato nella figura 1A), che è formato dalla membrana plasmatica apicale dei progenitori apicali (cartone animato nella figura 1A). Questo processo deve essere ripetuto per ogni cella che lo sperimentatore desidera iniettare. Inoltre, il successo della microiniezione dipende dal controllo preciso della profondità e della durata dell'iniezione di micropipette nel tessuto. Pertanto, nonostante i vantaggi unici, la microiniezione manuale è estremamente noiosa e richiede una notevole pratica per funzionare a velocità e resa ragionevoli, rendendo questa tecnica difficile da usare in modo scalabile. Per superare questa limitazione, abbiamo recentemente sviluppato un robot guidato dall'immagine, l'Autoiniettore12 (o robot di microiniezione) in grado di eseguire automaticamente microiniezioni in singole cellule.
Il robot di microiniezione utilizza algoritmi microscopici di imaging e visione artificiale per indirizzare con precisione posizioni specifiche nello spazio 3D all'interno del tessuto per la microiniezione(Figura 1B). Il robot di microiniezione può essere costruito apportando modifiche relativamente semplici a una configurazione di microiniezione esistente. Lo schema generale del robot di microiniezione è mostrato nella figura 1C. Una pipetta è montata in un supporto per pipette attaccato a un manipolatore a tre assi. Una fotocamera al microscopio viene utilizzata per acquisire immagini del tessuto e dell'ago di microiniezione. Un sistema di regolazione della pressione personalizzato viene utilizzato per controllare la pressione all'interno della pipetta e un micromanipolatore programmabile viene utilizzato per controllare la posizione della pipetta del microiniettore. Le immagini della telecamera della pipetta di tessuto e microiniezione vengono utilizzate per determinare la posizione spaziale della punta della pipetta di microiniezione e le posizioni in cui devono essere eseguite le microiniezioni. Il software calcola quindi le traiettorie necessarie per spostare la pipetta all'interno del tessuto. Tutto l'hardware è controllato dal software che abbiamo sviluppato in precedenza. Tutti i software sono scritti in linguaggio di codifica (ad esempio Python e Arduino) e possono essere scaricati da https://github.com/bsbrl/Autoinjector con le istruzioni. L'interfaccia grafica utente (GUI) consente all'utente di visualizzare il tessuto e la micropipetta e di personalizzare la traiettoria della microiniezione. Il nostro sistema può essere stabilito utilizzando modifiche relativamente semplici a un microscopio invertito dotato di filtri a campo luminoso ed epi-fluorescenza.
In primo luogo, forniamo istruzioni sulla preparazione di fette di tessuto organotipico cerebrale per la microiniezione. Quindi il protocollo illustra l'avvio del robot di microiniezione seguito da fasi preparatorie, come la calibrazione del movimento della pipetta, che devono essere fatte prima della microiniezione. Questo è seguito dalla definizione dei parametri di iniezione. Successivamente, l'utente può definire la traiettoria utilizzata dal robot di microiniezione e avviare la procedura di iniezione. Il tessuto microiniettato (in questo caso fette di tessuto organotipico cerebrale) può essere tenuto in coltura per diversi periodi di tempo a seconda del progettosperimentale 10,11. Il tessuto può essere elaborato per seguire e studiare l'identità e il destino delle cellule iniettate e la loro progenie. In alternativa, le cellule microiniettate possono essere seguite utilizzando l'imaging dal vivo. Nell'ambito di questo protocollo, dimostriamo l'uso del robot per microiniettare automaticamente le cellule progenitrici neurali in fette organotipiche di telencefalo dorsale E14.5 del mouse. Il robot è inoltre in grado di microiniettare nei neuroni dei neonati nel telencefalo del topo, così come nel telencefalo fetale umano12.
In sintesi, descriviamo una piattaforma robotica che può essere utilizzata per seguire e manipolare singole cellule nei tessuti. La piattaforma fa uso di pressione ed è, quindi, estremamente versatile per quanto riguarda la natura chimica del composto da iniettare. Inoltre, può essere adattato a cellule bersaglio diverse dalle cellule staminali. Ci aspettiamo che il nostro sistema sia facilmente adattabile anche ad altri sistemi modello.
Tutti gli studi sugli animali sono stati condotti in conformità con la legislazione tedesca sul benessere degli animali e le licenze necessarie sono state ottenute dalla Commissione etica regionale per la sperimentazione animale di Dresda, Germania (Tierversuchskommission, Landesdirektion Dresden). Le fette organotipiche sono state preparate a partire da E14.5 o E16.5 C57BL/6 telencefalo embrionale del topo (Janvier Labs).
1. Installazione di software
- Seguire le istruzioni per installare il software da https://github.com/bsbrl/Autoinjector.
2. Preparazione di reagenti e pipette
- Agarosio: Preparare il 3% di agarosio sciogliendo separatamente 3 g di agarosio ad ampia gamma e 3 g di agarosio a punto a bassa fusione in 100 ml di PBS di coltura cellulare in due bottiglie di vetro separate da 200 ml, rispettivamente. Conservare a temperatura ambiente per un massimo di 3 mesi.
- Soluzione di tirodo: Sciogliere 1 g di bicarbonato di sodio e sale di Tirodo (utilizzare il contenuto dell'intera bottiglia) e 13 ml di 1 M HEPES in 1 L di acqua distillata. Regolare il pH a 7,4. Filtrare la soluzione attraverso un filtro bottle-top da 0,2 μm.
- Mezzo di coltura a fette (SCM): Aggiungere 10 mL di siero di ratto, 1 mL di glutammina 2 mM, 1 mL penicillina-streptomicina (100x), 1 mL di integratore N-2 (100x), 2 mL di integratore B27 (50x) e 1 mL di HEPES (pH 7,3) tampone in 84 mL di mezzo neurobasale. Aliquota 5 mL di SCM in tubi da 15 mL. Conservare a -20 °C.
- CO2- Mezzo di microiniezione indipendente (CIMM): Preparare 5x soluzione di glucosio basso modificata DMEM (senza rosso fenolo) sciogliendo la polvere in 200 mL di acqua distillata. Filtrare la soluzione attraverso un filtro bottle-top da 0,2 μm (per la polvere DMEM, utilizzare il contenuto dell'intera bottiglia). Per preparare 100 mL di CIMM, mescolare 20 mL di soluzione modificata DMEM 5x, 1 mL di tampone HEPES, 1 mL di integratore N2 (100x), 2 mL di integratore B27 (50x), 1 mL di penicillina-streptomicina (100x), 1 mL di glutammina da 2 mM e 74 mL di acqua distillata. Conservare la soluzione a 4 °C.
- Tampone di ricostituzione: Preparare il tampone di ricostituzione sciogliendo 262 mM NaHCO3, 0,05 N NaOH, 200 mM HEPES in acqua distillata. Sterilizzare la soluzione filtrando attraverso un sistema di filtrazione a bottiglia da 0,22 μm in una bottiglia di vetro sterile. Aliquota 500 μL di tampone di ricostituzione in tubi di microcentrifugo ermetici. Conservare a 4 °C.
- Materiale colorante a microiniezione: Sciogliere il Dextran etichettato fluorescentmente in acqua distillata libera da RNasi (concentrazione finale 10 μg/μL). Preparare 5 aliquote μL e conservare a -20 °C fino all'uso.
- Estrarre le pipette di microiniezione dai capillari di vetro borosilicato (diametro esterno di 1,2 mm, diametro interno di 0,94 mm) utilizzando l'estrattore di micropipette. Proteggere le pipette dalla polvere. Non conservare pipette per più di 2 - 3 giorni. Per questo esperimento, i parametri di trazione erano HEAT: temperatura della rampa +1 – 5; PULL: 100; VEL: 110; DEL: 100. HEAT e VEL sono i parametri che influenzano la maggior parte della forma e delle dimensioni della pipetta.
NOTA: La pipetta di microiniezione ottimale ha una punta lunga e flessibile, per evitare danni alle cellule durante la microiniezione.
3. Preparazione delle fette di tessuto
- Sciogliere l'agarosio di vasta gamma del 3% utilizzando un forno a microonde prima della dissezione del tessuto cerebrale. Non lasciare che l'agarosio si solidifichiamo mantenendo in un bagno d'acqua a 37 °C prima dell'incorporamento. Assicurarsi che le pipette siano protette dalla polvere. Non conservare pipette per più di 2 - 3 giorni.
- Scongelare un'aliquota di SCM e riscaldare cimm da 10 a 12 mL e 20 mL della soluzione di Tyrode a 37 °C utilizzando un bagno d'acqua.
- Mescolare il tracciante fluorescente (Dextran-3000 o Dextran-10000-Alexa coniugato; concentrazione finale 5 - 10 μg/μL) con le altre sostanze chimiche da iniettare. Centrifugare la soluzione di microiniezione a 16.000 x g per 30 min a 4 °C. Raccogli il supernatante e trasferisci in un nuovo tubo. Mantenere la soluzione di microiniezione sul ghiaccio fino all'uso.
- Usa le teste degli embrioni di topo E13.5 - E16.5 per preparare fette di tessuto organotipico del telencefalo. Rimuovere la pelle e aprire il cranio usando le forcep, muovendosi lungo la linea mediana. Sezionare il cervello embrionale dal cranio aperto e rimuovere le meningi che coprono il tessuto cerebrale a partire dal lato ventrale del cervello. Lasciare l'intero cervello sezionato nella soluzione di Tyrode su un blocco riscaldante a 37 °C.
NOTA: Tutte le fasi di dissezione descritte al 3.4 devono essere eseguite nella soluzione di Tyrode prebellita. - Versare l'ampia gamma di agarosio fuso in uno stampo di incorporamento usa e getta. Quando l'agarosio viene raffreddato a 38 - 39 °C, trasferire accuratamente il cervello (un massimo di 4) in esso usando una pipetta Pasteur. Usa sempre le punte tagliate per questo passaggio.
- Mescolare l'agarosio intorno al tessuto usando una spatola o un paio di #1 Dumont senza toccare il tessuto. Lasciare che l'agarosio si solidifichiamo a temperatura ambiente. Una volta che l'agarosio si è solidificato, tagliare l'agarosio in eccesso che circonda il tessuto.
- Riempire il vassoio buffer con PBS. Orientare il cervello con l'asse rostro-caudale del tessuto perpendicolare al vassoio (utilizzare come punto di riferimento i bulbi olfattivi, che rappresentano la parte più rostrale del cervello). Tagliare fette da 250 μm usando un vibratomo.
- Riempire una piastra di Petri di 3,5 cm con 2 mL di supporti prebellici. Utilizzando una pipetta Pasteur di plastica, trasferire fette (10 - 15) in questo piatto. Una volta fatto, spostare la piastra di Petri con le fette nell'incubatore di coltura delle fette. Mantenere le fette a 37 °C in un'atmosfera umidificata contenente il 40% O2 / 5% CO2 / 55% N2 fino all'uso.
4. Microiniezione
- Accendere il computer, il microscopio, la fotocamera del microscopio, i manipolatori, il carro di pressione e il sensore di pressione. Caricare l'applicazione facendo clic sul file "launchapp.py" nella cartella principale scaricata da GitHub e specificare le impostazioni del dispositivo nella schermata popup (vedere il passaggio 1.1 per le istruzioni di installazione).
- Creare una pressione esterna per evitare intasamenti indesiderati prima di immergere la pipetta nella soluzione. Per applicare pressione alla pipetta, far scorrere la barra di pressione di compensazione al 24-45% e fare clic su Imposta valori. Successivamente, sintonizzare la pressione su una pressione sufficiente ruotando la manopola della valvola a pressione meccanica a 1 - 2 PSI (69 - 138 mbar) come indicato dal sensore di pressione.
- Trasferire le fette in una piastra di Petri di 3,5 cm contenente 2 mL di CIMM prerifatti. Posizionare le fette da microiniettare al centro della piastra di Petri. Trasferire la piastra di Petri allo stadio di microiniezione preriscaldato (37 °C).
- Caricare la pipetta di microiniezione con 1,4 –1,6 μL di soluzione microiniettata (dal passaggio 3.3) utilizzando una pipetta di plastica a punta lunga. Inserire la pipetta di microiniezione sul supporto della pipetta.
- Utilizzando l'ingrandimento più basso al microscopio, mettere a fuoco la fetta e guidare la micropipetta in questo campo visivo (FOV) in modo che sia focalizzata sullo stesso piano della destinazione della fetta. Passare l'uscita del microscopio alla fotocamera per visualizzare l'FOV nell'applicazione.
- Fare clic sul pulsante di ingrandimento in alto a sinistra dell'interfaccia per avviare la calibrazione del dispositivo. Verrà visualizzata una finestra per selezionare l'ingrandimento. Selezionare l'ingrandimento 10x o qualsiasi ingrandimento impostato sull'obiettivo (ad esempio, 4x, 10x, 20x, 40x) e premere OK. Il software presuppone che l'obiettivo interno sia 10x (l'ingrandimento obiettivo più comune).
- Rifocalizzare la punta della pipetta utilizzando la ruota micrometrica del microscopio e fare clic sulla punta della pipetta con il cursore. Premere quindi il pulsante 1.1 e premere OK nella finestra popup. La pipetta si muoverà nella direzione Y. Fate clic sulla punta della pipetta e premete il pulsante passo 1.2. Infine, immettete 45 nella casella Angolo pipetta (Pipette angle) e premete l'angolo Imposta (Set angle).
- Immettere i parametri desiderati nel pannello Controlli di microiniezione automatizzati. Per la microiniezione in progenitori apicali impostare la distanza di iniezione a 20 - 40 μm e la profondità a 10 - 15 μm. Per la microiniezione nei neuroni impostare la distanza di iniezione 30 - 40 μm dal lato basale e la profondità a 10 - 30 μm a seconda di ciò che viene preso di mira. Imposta sempre la velocità al 100%. Fare clic su Imposta valori.
NOTA: La distanza di avvicinamento è la distanza che la pipetta estrae dal tessuto prima di passare alla distanza di iniezione successiva, la profondità è la profondità nel tessuto che va la microiniezione, la spaziatura è la distanza lungo la linea tra le iniezioni sequenziali, la velocità è la velocità della pipetta in μm / s. - Fate clic sul pulsante Disegna spigolo (Draw edge) e trascinate il cursore lungo la traiettoria desiderata nella finestra popup per definire la traiettoria di iniezione. Per le cellule staminali progenitrici di microiniezione, il lato ventrale della superficie del telencefalo è mirato, come mostrato nella figura 2A. Portare la pipetta all'inizio della linea e fare clic sulla punta della pipetta. Fate clic su Esegui traiettoria (Run trajectory) per avviare la microiniezione. Ripetere questo passaggio per ogni piano di iniezione mirato (di solito fatto per 3 - 4 piani con 40 - 75 iniezioni per piano).
5. Coltura tissutale e lavorazione delle fette tissutali per l'immunofluorescenza
- Preparare la miscela di collagene (1,5 mg/mL): A un tubo aggiungere 1,25 mL della soluzione matriciale, 0,5 ml di acqua distillata, 0,5 ml di soluzione DMEM-F12 5x e 0,25 ml di tampone di ricostituzione. Tienilo sul ghiaccio fino all'uso.
- Ottenere la piastra di Petri contenente le fette microiniettate dalla camera di incubazione della coltura della fetta e immergere le fette nella miscela di collagene.
- Trasferire le fette insieme a 200 - 300 μL di miscela di collagene in un pozzo di 14 mm di un piatto con fondo di vetro da 35 mm. Assicurarsi che le fette siano coperte con molto meno collagene. Questo set up consente le condizioni ottimali per i nutrienti e l'assorbimento di ossigeno.
- Orientare le sezioni assicurando al contempo che vi sia spazio sufficiente tra le fette utilizzando due coppie di forcep. Incubare la piastra di Petri per 5 minuti a 37 °C utilizzando un blocco riscaldante per consentire al collagene di solidificarsi. Considera questa volta come t = 0 delle impostazioni cultura delle fette.
- Spostare la piastra di Petri nell'incubatore di coltura delle fette per altri 40 minuti. Quindi aggiungere 2 mL dell'SCM prebellico. Le sezioni vengono mantenute in coltura fino al punto di tempo desiderato.
- Togliere le fette dall'incubatore di coltura slice e aspirare l'SCM. Lavare le fette incorporate nel collagene con 1x PBS. Aggiungere il 4% (wt/vol) di paraformaldeide (in tampone fosfato da 120 mM, pH 7,4) e lasciare il tessuto a RT per 30 minuti. Quindi spostarlo a 4 °C per consentire la fissazione notturna.
- Aspirare la soluzione di paraformaldeide il giorno successivo ed eseguire 1 lavaggio PBS. Per rimuovere le fette dal collagene, utilizzare due paia di forcep per estrarre delicatamente le fette sotto uno stereomicroscopio.
- Utilizzare un forno a microonde per sciogliere l'agarosio a basso punto di fusione del 3% (wt/vol) per la lavorazione delle fette microiniettate. Versare l'agarosio fuso in uno stampo di incorporamento usa e getta e lasciarlo raffreddare a circa 38 - 39 °C.
- Trasferire le fette di tessuto dal passo 5.7 in questo stampo contenente agarosio a bassa fusione utilizzando una pipetta Pasteur di plastica. Assicurarsi che il lato pio della fetta sia verso l'alto e che la superficie ventricolare sia verso il basso. Se necessario orientarsi di conseguenza. Lasciare raffreddare l'agarosio a RT per solidificarsi.
- Tagliare l'agarosio extra che circonda le fette. Orientare il blocco di agarosio per assicurarsi che la superficie di taglio sia parallela alla lama di taglio del vibratomo. Usando il vibratoma, tagliare sezioni spesse 50 μm.
- Riempire un piatto da 24 po 'con 1x PBS. Trasferire le sezioni in questo piatto utilizzando un pennello a punta fine. Eseguire l'immunofluorescenza secondo i protocolli standard.
La microiniezione ha lo scopo di tracciare e manipolare singole cellule staminali neurali e la loro progenie nei tessuti viventi e di seguire la loro progressione del lignaggio in un ambiente fisiologico. In questo articolo, abbiamo dimostrato l'uso del robot di microiniezione per il targeting e l'iniezione automatica di fette organotipiche del telecephalon del topo. La figura 2 illustra le immagini rappresentative delle cellule progenitrici iniettate con successo e la figura 3 illustra i neuroni appena nati iniettati. Quando vengono iniettate con colorante Dextran Alexa-488 (o Alexa-A555), le cellule appaiono completamente riempite con il colorante. Per quanto riguarda i progenitori apicali (Figura 2) l'imaging confocale consente di ricostruire ad alta risoluzione spaziale la morfologia cellulare, la presenza - o assenza - dell'attacco apicale e basale, e di combinare l'indagine morfologica con l'espressione marcatore. Combinando questi criteri, l'utente può assegnare un destino cellulare specifico alle cellule microiniettate e alla loro progenie. Per quanto riguarda l'iniezione neuronale, l'utente può ricostruire la morfologia neuronale, compresa la struttura e le caratteristiche della dendrite apicale e dell'assone. La microiniezione automatizzata può fornire una produttività significativamente più elevata rispetto alla microiniezione manuale (Figura 2B). Inoltre, l'etichettatura EdU conferma che la vitalità delle celle non è influenzata dall'automazione (Figura 2C). Mantenere la fetta organotipica in coltura consente di seguire la progressione del lignaggio delle cellule microiniettate (abbiamo mostrato 4 - 24h nella figura 2D). Se la soluzione di microiniezione contiene materiale genetico (guide DNA, mRNA, CRISPR-Cas9) o proteine ricombinanti, ciò consente di studiare se e come la progressione del lignaggio è influenzata dalla manipolazione.
La microiniezione in singole cellule staminali neurali nei tessuti fornisce un'eccellente risoluzione a singola cellula e per questo motivo è stata utilizzata per sezionare la biologia cellulare della progressione delle cellule staminali neurali e della transizione del destino (Figura 3A). La microiniezione consente la fornitura di miscele complesse di sostanze chimiche. In precedenza abbiamo utilizzato questa funzione per studiare l'accoppiamento giunzionale nelle cellule progenitrici neurali mescolando permeabile alla giunzione gap con coloranti fluorescenti impermeabili slanciati12. Abbiamo esteso il lavoro precedente studiando l'accoppiamento giunzionale nei neuroni appena nati, iniettando Lucifer Yellow insieme a Dextran-A555 (Figura 3B). Come mostrato nella figura 3B, una proporzione di neuroni piramidali appena nati sono accoppiati attraverso giunzioni gap ai neuroni vicini. Questa osservazione è coerente con l'idea che i neuroni immaturi comunichino attraverso lagiunzione gap 13,14. Inoltre, il targeting dei neuroni mostra che l'uso del robot di microiniezione può essere generalizzato a diversi tipi di cellule nel cervello dei mammiferi in via di sviluppo. Questa configurazione sperimentale sarà utile per sezionare la biologia cellulare dei neuroni nei tessuti, ad esempio fornendo oligopeptidi specifici per interferire con le interazioni proteina-proteina.
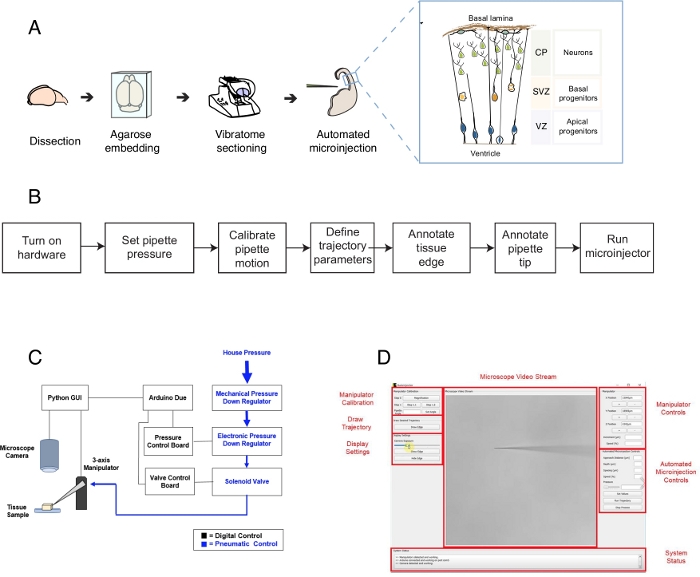
Figura 1: Configurazione e protocollo automatizzati della microiniezione. (A) Protocollo globale per la preparazione dei tessuti e le microiniezioni automatizzate mediante il robot di microiniezione. Inserto destro: Schema cartoon del telencefalo del mouse destinato alla microiniezione in questo protocollo. (B) Diagramma di flusso delle fasi automatizzate di microiniezione. (C) Schema dell'hardware del robot a microiniezione. (D) Interfaccia grafica utente (GUI) del software utilizzato per controllare e far funzionare il robot di microiniezione. Questa cifra è adattata dalrif. Clicca qui per visualizzare una versione più grande di questa figura.
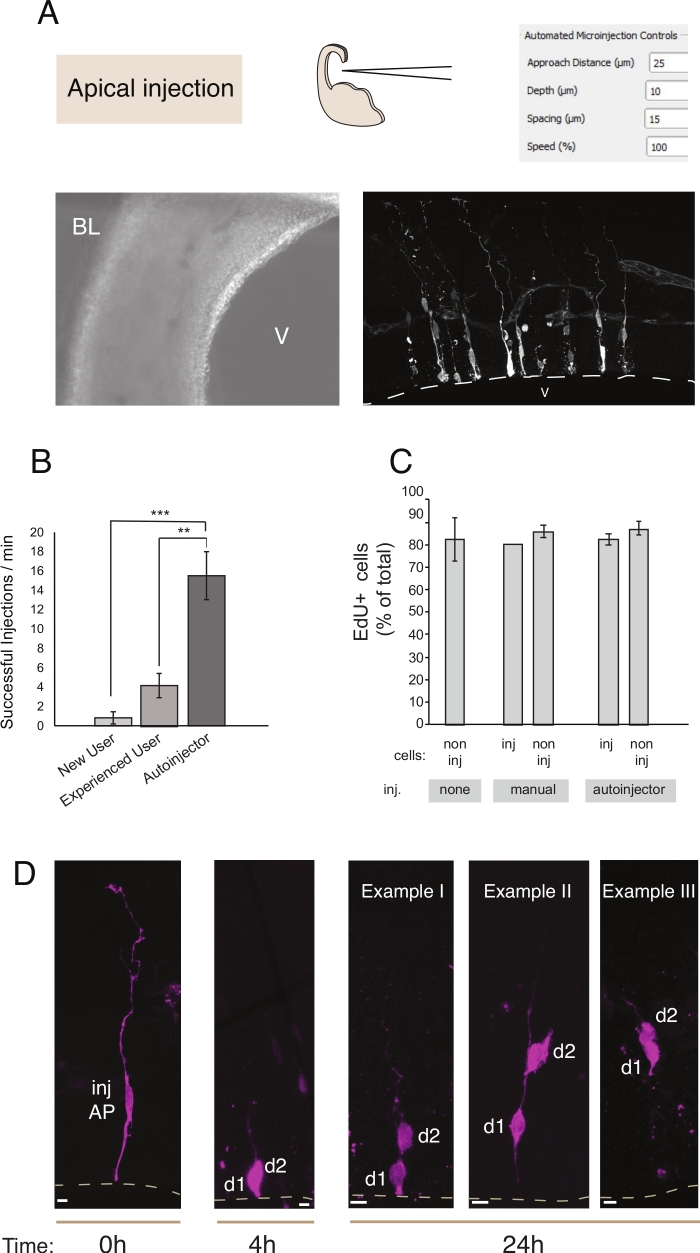
Figura 2: Microiniezione robotica in progenitori apicali. Risultati schematici e attesi quando si utilizza il robot di microiniezione per indirizzare i progenitori apicali (AP) attraverso la superficie apicale (iniezione apicale). (A) Riga superiore. A sinistra: schema del processo. A destra: GUI con parametri rilevanti per l'iniezione apicale. Riga inferiore. A sinistra: immagine di contrasto di fase scattata durante la procedura di iniezione (V: ventricolo; BL: lamina basale). A destra: risultati rappresentativi che mostrano I POP microiniettati. La linea tratteggiata rappresenta il ventricolo (V). Barra di scala: 10 μm. (B) Iniezioni di successo al minuto per un utente alle prime armi sul sistema di microiniezione manuale, un utente esperto sul sistema di microiniezione manuale e il robot di microiniezione. (C) Incorporazione dell'EDU nelle cellule microiniettate e nelle cellule non iniettate nell'area iniettata. Le fette organotipiche di telencefalo dorsale E14.5 del mouse sono state (i) non iniettate o (ii) sottoposte a microiniezione manuale o automatizzata (fetta iniettata) utilizzando Dextran-A488 (per manuale e autoiniettore). Le fette sono state tenute in coltura in presenza di EdU per 24 ore, poi sono state fissate e macchiate per DAPI ed EdU. Le cellule iniettate e non iniettate nell'area iniettata sono state valutate per positività edU. (D) Uso del robot di microiniezione Tracciatura del lignaggio. Un colorante fluorescente (Dx3-A555, magenta) viene iniettato in una singola cellula staminali neurale (t = 0 h). Il colorante fluorescente è diviso alle cellule figlie (d1, d2) durante la mitosi. Ciò consente di seguire la progenie della cellula iniettata (t = 4 h e 24 h) e rivelare la progressione del lignaggio nel tempo. Per t = 24 h, mostriamo diversi esempi della progenie che ci si aspetta di trovare. Barre di scala: 10 μm. I grafici in B e Csono presi dalrif.
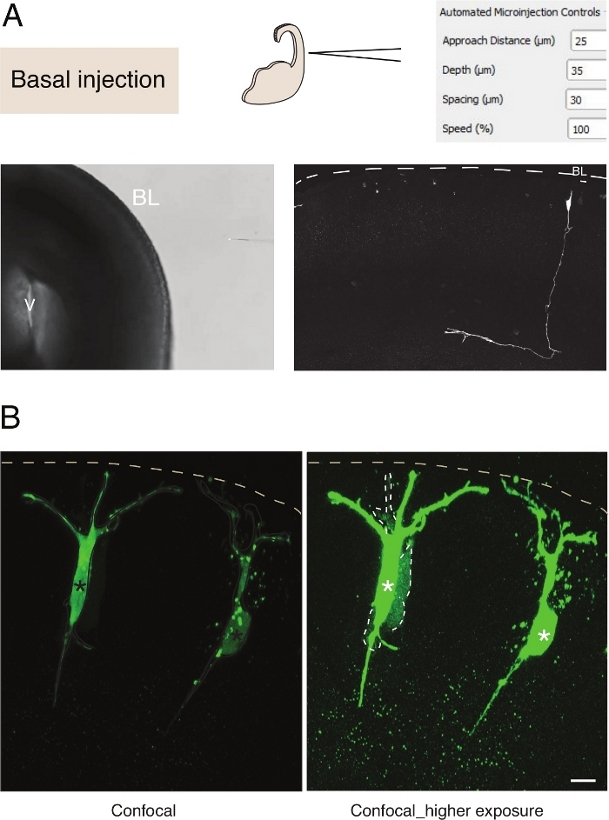
Figura 3: Microiniezione robotica nei neuroni. Risultati schematici e attesi quando si utilizza il robot di microiniezione per colpire i neuroni piramidali (N) attraverso la superficie basale (iniezione basale). (A) Riga superiore. A sinistra: schema del processo. A destra: GUI con parametri rilevanti per l'iniezione basale. Riga inferiore. A sinistra: immagine di contrasto di fase scattata durante la procedura di iniezione (V: ventricolo; BL: lamina basale). A destra: risultati rappresentativi che mostrano una linea N. Tratteggiata microiniettata rappresenta la lamina basale (BL). Barra di scala: 10 μm. (B) Uso dell'autoiniettore per studiare la comunicazione giunzione gap nei tessuti. Ai neuroni piramidali è stata iniettata una soluzione contenente due coloranti: il Dx-A555 a snodo junctional-impermeabile (non mostrato) e il lucifero giallo permeabile a giunzione gap (verde). Dx-A555 è confinato alla cella di destinazione (asterischi), mentre il LY si diffonde alle celle collegate tramite giunzione gap alla cella di destinazione (linee tratteggiate). Pannello sinistro: Bassa esposizione, sono visibili solo le cellule microiniettate. Pannello destro: l'esposizione elevata consente la visualizzazione delle celle iniettate e delle celle accoppiate (linee tratteggiate). Barra di scala: 10 μm. Fare clic qui per visualizzare una versione più grande di questa figura.
File aggiuntivo: risoluzione di diversi errori comuni che si verificano durante la microiniezione. Clicca qui per scaricare questo file.
La microiniezione in singole cellule staminali neurali nei tessuti fornisce un'eccellente risoluzione a singola cellula e per questo motivo è stata utilizzata per sezionare la biologia cellulare della progressione delle cellule staminali neurali e della transizione del destino (Figura 2;vedi anche 10,11,12). La procedura automatizzata di microiniezione può essere eseguita su altri tipi di cellule sia nei topi embrionali che nel tessuto cerebrale umano. I risultati rappresentativi della microiniezione dei neuroni appena nati prendendo di mira la superficie basale del telencefalo sono mostrati nella figura 3.
Il principio qui stabilito può essere applicato per colpire diversi tipi di cellule nel cervello embrionale del topo e nel cervello umano. Abbiamo precedentemente dimostrato che il robot di microiniezione può anche essere utilizzato per colpire singole cellule progenitrici nel cervello posteriore del topo e nel telencefalo e nei neuroni appena nati nel topo e nell'uomo sviluppando la neocorteccia12. Per ottenere i migliori risultati della procedura di iniezione, è necessario ottimizzare tutti i passaggi prima di iniziare l'iniezione. È importante considerare attentamente e ottimizzare la preparazione di fette di tessuto organotipico vitali e ben conservate dal tessuto cerebrale (Figura 1). È fondamentale essere rapidi nella procedura di dissezione e affezione illustrata nella figura 1. Per l'iniezione apicale destinata agli AP, è necessario scegliere le fette che mostrano l'orientamento ideale della superficie apicale. Per l'iniezione di AP, l'orientamento ideale è la superficie apicale perpendicolare al fondo della piastra di Petri. Qualsiasi altro orientamento sarà anche permissivo, tuttavia, la superficie apicale perpendicolare alla piastra di Petri fornisce una superficie più ampia per l'iniezione, aumentando così il successo dell'iniezione. Per l'iniezione nei neuroni, l'orientamento della fetta gioca poco o nessun effetto.
Una volta selezionate le fette da iniettare, la procedura di iniezione per fetta richiede circa 5 minuti. Considerando che si lavora con il tessuto vivente, si consiglia vivamente di accelerare la procedura di iniezione. A tal fine si consiglia di impostare tutti i parametri per l'iniezione tramite la GUI (Figura 1D) prima che il tessuto sia pronto, per ridurre eventuali tempi di attesa non necessari. Per la risoluzione dei problemi si rimanda al file supplementare.
In caso di coltura di fette a lungo termine, i passaggi successivi alla procedura di microiniezione automatizzata possono influire sullo stato delle cellule e quindi sull'esperimento. Pertanto, si consiglia vivamente di eseguire un test di controllo qualità e ottimizzare le condizioni delle impostazioni cultura delle sezioni. Per valutare la vitalità cellulare dopo la procedura di affettamento e iniezione, abbiamo eseguito l'etichettatura EdU durante la coltura e abbiamo quantificato il numero di nuclei pyknotic (un proxy per le cellule apoptotiche) nelle colture e nel tessuto iniettato12. Queste quantificazioni non hanno rivelato alcun impatto significativo della microiniezione sulla vitalità dei tessuti (figura 2C). Si consiglia di eseguire controlli di qualità simili stabilendo al contempo la pipeline di affettare e microiniettare i tessuti organotipici in laboratorio.
Rispetto alla microiniezione manuale, il robot di microiniezione offre diversi vantaggi. In primo luogo, la curva di apprendimento per l'utente è meno ripida rispetto all'iniezione manuale: un nuovo utente raggiungerà un'elevata competenza dopo un numero limitato di sessioni, in genere 1 o 2. In secondo luogo, nel caso della microiniezione manuale, una competenza comparabile richiede mesi di formazione. La procedura di iniezione è più rapida ed efficiente (Figura 2B). Abbiamo quantificato questi parametri e abbiamo scoperto che il robot di microiniezione ha sovraperformato un utente manuale qualificato per quanto riguarda il successo dell'iniezione (% di iniezione riuscita / numero totale di iniezioni) e nel numero totale di iniezioni per unità di tempo12. Ciò si traduce in un aumento complessivo del 300% dell'efficienza di iniezione (% di iniezione/min di successo) per il robot di microiniezione rispetto a un utente qualificato. L'aumento dell'efficienza è stato ancora più pronunciato quando si confronta il robot di microiniezione con un utente principiante e ha raggiunto il 700%. Ultimo ma non meno importante, il robot di microiniezione può essere facilmente programmato per esplorare sistematicamente tutti i parametri spaziali. Ciò è particolarmente vantaggioso quando si adatta il robot di microiniezione per colpire nuove cellule o tessuti o quando si utilizza il robot di microiniezione per scopi che richiedono una diversa risoluzione spaziale.
La costruzione del robot di microiniezione richiede modifiche minime a un microscopio a epi-fluorescenza esistente12. In precedenza abbiamo fornito istruzioni per questo adattamento a https://github.com/bsbrl/Autoinjector. Una volta configurato l'hardware, questo protocollo fornisce dettagli metodologici chiave per intraprendere con successo microiniezioni automatizzate. Nel complesso, il robot di microiniezione ha un tasso di iniezione di successo di 15,52 + 2,48 iniezioni / min, che è 15 volte superiore a un utente inesperto (1,09 ± 0,67 iniezioni / min) e 3 volte superiore a un utente esperto (4,95 ± 1,05 iniezioni / min)12. Questo miglioramento del tasso di iniezione riuscito consente sia agli utenti alle prime armi che agli utenti esperti di iniettare più cellule in un lasso di tempo più breve, essenziale per preservare la vitalità dei tessuti. Inoltre, il robot di microiniezione è personalizzabile e la traiettoria, la profondità dell'iniezione, il numero di iniezioni, la spaziatura tra le iniezioni possono essere tutti sintonizzati utilizzando la GUI. Queste caratteristiche consentono al robot di microiniezione di essere utilizzato come strumento per ottimizzare esperimenti precedentemente laboriosi ed esplorare esperimenti fondamentalmente nuovi che richiedono una resa più elevata di quanto precedentemente possibile.
I principali limiti della procedura di microiniezione che abbiamo descritto qui sono legati alla preparazione di fette di tessuto, un passaggio cruciale che richiede un'ampia ottimizzazione. Inoltre, la microiniezione si basa sulla presenza di una superficie che può essere avvicinata dalla pipetta di vetro. Questa funzione limita il tipo di tessuti e posizioni tissutali che possono essere presi di mira tramite microiniezione utilizzando la configurazione presente.
Il robot di microiniezione utilizza attualmente l'imaging a campo luminoso ed è stato utilizzato in preparati in vitro per fette cerebrali. In futuro, il robot di microiniezione potrebbe essere combinato con l'imaging a 2 fotoni per aumentare la specificità del targeting a singola cellula in vivo per l'etichettatura molecolare o colorante. Tali sforzi sono già stati compiuti per l'elettrofisiologia acella singola 15,16. Il dispositivo corrente richiede l'osservazione manuale della procedura di microiniezione. Le versioni future potrebbero includere strategie per la pulizia delle pipette di microiniezione ostrute17 o l'integrazione di robot di movimentazione dei fluidi18 per microiniezioni multiplexate e completamente autonome. Questi dispositivi potrebbero aumentare la scala di microiniezione per ordini di grandezza. L'adattamento di algoritmi per il controllo parallelo di più pipette di microiniezione19 potrebbe consentire la consegna multiplexata di dozzine di coloranti e reagenti molecolari nelle stesse cellule all'interno degli stessi esperimenti. Questo ha il potenziale per aprire nuove strade per lo screening molecolare nei tessuti.
Il robot di microiniezione potrebbe essere usato per taggare cellule identificate funzionalmente utilizzando codici a barre DNA o RNA. Questo potrebbe a sua volta essere combinato con altre tecniche di analisi a singola cellula, come il sequenziamento dell'RNA a singola cella (scRNAseq) e la microscopia elettronica. I nostri risultati preliminari mostrano che le cellule microiniettate e la loro progenie possono essere recuperate e isolate utilizzando la dissociazione tissutale seguita dallo smistamento FACS (Taverna, risultati inediti). Le celle ordinate FACS possono quindi essere utilizzate per scRNAseq. Inoltre, i risultati preliminari mostrano che le capacità di risoluzione a singola cellula del robot di microiniezione possono essere utilizzate in combinazione con l'analisi microscopica elettronica per esplorare la biologia cellulare sulle cellule staminali neurali nei tessuti ad alta risoluzione spaziale (Taverna e Wilsch-Bräuninger, risultati inediti). Questi dati suggeriscono che il robot di microiniezione può essere utilizzato come strumento per la microscopia della luce correlativa e dell'elettrone nei tessuti e, in senso più ampio, per l'analisi multimodale dell'identità cellulare e del comportamento nei tessuti.
La microiniezione si basa sull'uso della pressione e ci si può permettere soluzioni di iniezione ad alta complessità molecolare (ad esempio, un intero trascritoma). Questa caratteristica della microiniezione è stata sfruttata in passato per isolare e clonare i recettori ligandi-gated20. Lungo questa linea, il robot di microiniezione potrebbe essere utilizzato per modellare e studiare tratti multigenici a livello cellulare. Combinato con una strategia di sub-pooling, il robot di microiniezione potrebbe anche essere utilizzato come piattaforma per identificare l'insieme minimo di geni che guidano un certo comportamento tratto / cellulare. Finora, il robot di microiniezione è stato utilizzato per manipolare la biochimica della cellula attraverso la consegna di mRNA, DNA o proteine ricombinanti10,21,22. Prevediamo un'applicazione del robot di microiniezione nel sondare la biofisica dello spazio intracellulare, ad esempio fornendo nanomateriali o nanomacchine che consentono il rilevamento e/o la manipolazione delle proprietà biofisiche dello spazio intracellulare.
Gli autori non hanno nulla da rivelare.
Gli autori vorrebbero riconoscere la Fondazione Nomis (ET). SBK riconosce i fondi del dipartimento di Ingegneria Meccanica, College of Science and Engineering, iniziativa MnDRIVE RSAM del Dipartimento di istruzione superiore dell'Università del Minnesota, Minnesota, National Institutes of Health (NIH) 1R21NS103098-01, 1R01NS111028, 1R34NS111654, 1R21NS112886 e 1R21 NS111196. GS è stato supportato dalla National Science Foundation Graduate Research Fellowship e dalla borsa di formazione NSF IGERT.
| Name | Company | Catalog Number | Comments |
| Chemicals | |||
| Agarose, Low Melt | Carl Roth | Cat# 6351.2 | |
| Agarose, Wild Range | Sigma-Aldrich | Cat# A2790 | |
| Best-CA 221 Glue | Best Klebstoffe GmbH & Co.KG | Cat# CA221-10ml | |
| B-27 Supplement | Thermo Fisher Scientific | Cat# 17504044 | |
| Cellmatrix Type-IA (Collagen, Type !) | FUJIFILM Wako Chemicals | Cat# 637-00653 | |
| Distilled Water | |||
| DMEM-F12, CO2 independent (w/o Phenol red) | Sigma-Aldrich | Cat# D2906 | |
| DMEM-F12, CO2 independent (with Phenol red) | Sigma-Aldrich | Cat# D8900 | |
| HEPES-NAOH, pH 7.2, 1M (HEPES buffer) | Carl Roth | Cat# 9105.3 | |
| L-Glutamine, 200 mM | Thermo Fisher Sientific | Cat# 25030024 | |
| Mowiol 4-88 | Sigma-Aldrich | Cat# 81381 | |
| N-2 Supplement | Thermo Fisher Scientific | Cat# 17502048 | |
| Neurobasal Medium | Thermo Fisher Scientific | Cat# 21103049 | |
| Nuclease-free water | Thermo Fisher Scientific | Cat# AM9937 | |
| O2 (40%), CO2 (5%), N2 (55%) Mix, 50 liters | |||
| Paraformaldehyde | Merck | Cat# 818715 | |
| PBS | |||
| Penicillin-Streptomycin (10,000 U/mL) | Thermo Fisher Scientific | Cat# 15140122 | |
| Rat serum | Charles River Laboratories | ||
| Japan | |||
| Sodium bicarbonate (NaHCO3) | Merck | Cat# 106323 | |
| Sodium hydroxide (NaOH) | Merck | Cat# 106482 | |
| Tyrode’s salt | Sigma | Cat# T2145-10x1L) | |
| Equipment | |||
| Borosilicate glass capillaries, 1.2 mm outer diameter x 0.94 mm inner diameter | Sutter Instruments | Cat# BF-120-94-10 | |
| Bottle-top filter system, 500 mL | Corning | Cat# 430769 | |
| Computer PC | |||
| Custom pressure rig | Custom pressure rig | ||
| Electronic pressure regulator | Parker Hannifin | Cat# 990-005101-002 | |
| Falcon tubes, 15 mL | Corning | Cat# 430791 | |
| Falcon tubes, 50 mL | Corning | Cat# 430829 | |
| Fine-tip paintbrush | |||
| Flaming/ Brown micropipette puller | Sutter Instruments | Cat# P-97 | |
| Forceps, Dumont no. 3 | Fine Science Tools | Cat# 11231-30 | |
| Forceps, Dumont no. 5 | Fine Science Tools | Cat# 11255-20 | |
| Forceps, Dumont no. 55 | Fine Science Tools | Cat# 11252-20 | |
| Heating block | Labtech International | Cat # Dri block Digi2 | |
| Inverted fluorescence microscope | Zeiss | Cat# Axiovert 200 | |
| Light source | Olympus | Cat# Highlight 3100 | |
| Manual pressure regulator | McMaster Carr | Cat# 0-60 PSI 41795K3 | |
| Microloader Tips | Eppendorf | Cat# 5242956.003 | |
| Microcontroller | Arduino | Cat# Arduino Due | |
| Microscope camera Hamamatsu Orca Flash 4.0 V3 | |||
| Motorized stage XY for microscope | |||
| Multiwell plate, 24 wells | Nunc | Cat# 142475 | |
| Pasteur pipettes, plastic | |||
| Petri dish, 60 x 15 mm | Greiner | Cat# 628102 | |
| Petri dish, 35 x 10 mm | Nunc | Cat# 153066 | |
| Petri dish, 34 x 14 mm, including Microwell no. 1.5 cover glass | MatTek | Cat# P35G-1.5-14-C | |
| Pipette holder | Warner Instruments | Cat# 64-2354 MP-s12u | |
| Pipette and tips | |||
| Puller filament, 3.0-mm square box filament | Sutter Instrument | Cat# FB330B | |
| Slice culture incubation box | MPI-CBG | Cat# custom made | |
| Solenoid valve | Cat# LHDA053321H-A | ||
| Stereomicroscope | Olympus | Cat# SZX12 | |
| Tabletop centrifuge | Heraeus | Cat# 5431622 | |
| Thermometer | |||
| Three-axis Manipulator | Sensapex Inc | Cat# tree-axis uMP | |
| Vibratome | Leica | Cat# VT1000s | |
| Whole-embryo-culture-system incubator | Ikemoto Company | Cat# RKI-10-0310 | |
| Waterbath | |||
| Software and Algorithms | |||
| Arduino | Arduino | ||
| Fiji | RRID: SCR_002285 | ||
| Python | Python Software foundation | Python 2.7.12 | |
| ZEN | RRID: SCR_013672 |
- Taverna, E., Götz, M., Huttner, W. B. The Cell Biology of Neurogenesis: Toward an Understanding of the Development and Evolution of the Neocortex. Annual Review of Cell and Developmental Biology. 30 (1), 465-502 (2014).
- Götz, M., Huttner, W. B. The cell biology of neurogenesis. Nature Reviews Molecular Cell Biology. 6 (10), 777-788 (2005).
- Di Lullo, E., Kriegstein, A. R. The use of brain organoids to investigate neural development and disease. Nature Reviews Neuroscience. 18 (10), 573-584 (2017).
- Lancaster, M. A., Knoblich, J. A. Organogenesisin a dish: Modeling development and disease using organoid technologies. Science. 345 (6194), 1247125 (2014).
- Kretzschmar, K., Clevers, H. Organoids: Modeling Development and the Stem Cell Niche in a Dish. Developmental Cell. 38 (6), 590-600 (2016).
- Pepperkok, R. et al. Automatic microinjection system facilitates detection of growth inhibitory mRNA. Proceedings of the National Academy of Sciences of the United States of America. 85 (18), 6748-6752 (1988).
- Pepperkok, R., Lowe, M., Burke, B., Kreis, T. E. Three distinct steps in transport of vesicular stomatitis virus glycoprotein from the ER to the cell surface in vivo with differential sensitivities to GTPγS. Journal of Cell Science. 111 (13), 1877-1888 (1998).
- Pepperkok, R. et al. β-COP is essential for biosynthetic membrane transport from the endoplasmic reticulum to the Golgi complex in vivo. Cell. 74 (1), 71-82 (1993).
- Ansorge, W., Pepperkok, R. Performance of an automated system for capillary microinjection into living cells. Journal of Biochemical and Biophysical Methods. 16 (4), 283-292 (1988).
- Taverna, E., Haffner, C., Pepperkok, R., Huttner, W. B. A new approach to manipulate the fate of single neural stem cells in tissue. Nature Neuroscience. 15 (2), 329-337 (2012).
- Wong, F. K., Haffner, C., Huttner, W. B., Taverna, E. Microinjection of membrane-impermeable molecules into single neural stem cells in brain tissue. Nature Protocols. 9 (5), 1170-1182 (2014).
- Shull, G., Haffner, C., Huttner, W. B., Kodandaramaiah, S. B., Taverna, E. Robotic platform for microinjection into single cells in brain tissue. EMBO Reports. 20 (10), e47880 (2019).
- Jabeen, S., Thirumalai, V. The interplay between electrical and chemical synaptogenesis. Journal of Neurophysiology. 120 (4), 1914-1922 (2018).
- Nagy, J. I., Pereda, A. E., Rash, J. E. Electrical synapses in mammalian CNS: Past eras, present focus and future directions. Biochimica et Biophysica Acta - Biomembranes. 1860 (1), 102-123 (2018).
- Suk, H.J. et al. Closed-loop real-time imaging enables fully automated cell-targeted patch-clamp neural recording in vivo. Neuron. 95 (5), 1037-1047 (2017).
- Annecchino, L. A. et al. Robotic automation of in vivo two-photon targeted whole-cell patch-clamp electrophysiology. Neuron. 95 (5), 1048-1055 (2017).
- Kolb, I. et al. Cleaning patch-clamp pipettes for immediate reuse. Scientific Reports. 6, 35001 (2016).
- Holst, G. L. et al. Autonomous patch-clamp robot for functional characterization of neurons in vivo: development and application to mouse visual cortex. Journal of Neurophysiology. 121 (6), 2341-2357 (2019).
- Kodandaramaiah, S. B. et al. Multi-neuron intracellular recording 1 in vivo via interacting autopatching 2 robots. ELife. 7, 24656 (2018).
- Lubbert, H. et al. cDNA cloning of a serotonin 5-HT1c receptor by electrophysiological assays of mRNA-injected Xenopus oocytes (RNA fractionation/hybrid depletion/hybrid selection/choroid plexus/voltage clamp). Neurobiology. 84 (2) 4332-4336 (1987).
- Florio, M. et al. Human-specific gene ARHGAP11B promotes basal progenitor amplification and neocortex expansion. Science. 347 (6229), 1465-1470 (2015).
- Kalebic, N. et al. CRISPR/Cas9-induced disruption of gene expression in mouse embryonic brain and single neural stem cells in vivo. EMBO Reports. 17 (3), 338-348 (2016).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved