
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Bioengineering
Eenvoudige, betaalbare en modulaire patronen van cellen met behulp van DNA
Hier presenteren we een protocol voor micropatrintercellen met eencellige resolutie met behulp van DNA-geprogrammeerde adhesie. Dit protocol maakt gebruik van een benchtop fotolithografieplatform om patronen van DNA-oligonucleotiden op een glasplaat te creëren en vervolgens celmembranen te labelen met in de handel verkrijgbare complementaire oligonucleotiden. Hybridisatie van de oligo's resulteert in geprogrammeerde celadhesie.
De relatieve positionering van cellen is een belangrijk kenmerk van de micro-omgeving die cel-cel interacties organiseert. Om de interacties tussen cellen van hetzelfde of een ander type te bestuderen, zijn micropatterningtechnieken nuttig gebleken. DNA Programmed Assembly of Cells (DPAC) is een micropatterning-techniek die zich richt op de hechting van cellen aan een substraat of andere cellen met behulp van DNA-hybridisatie. De meest elementaire operaties in DPAC beginnen met het versieren van celmembranen met lipide-gemodificeerde oligonucleotiden en stromen ze vervolgens over een substraat dat is gemodelleerd met complementaire DNA-sequenties. Cellen hechten zich selectief aan het substraat alleen waar ze een complementaire DNA-sequentie vinden. Niet-hechtende cellen worden weggespoeld, waardoor een patroon van aanhangende cellen wordt onthuld. Aanvullende bewerkingen omvatten verdere rondes van celsubstraat of cel-celadhesie, evenals het overbrengen van de patronen gevormd door DPAC naar een inbeddingshydrogel voor langdurige kweek. Voorheen vereisten methoden voor het patroon van oligonucleotiden op oppervlakken en het versieren van cellen met DNA-sequenties respectievelijk gespecialiseerde apparatuur en aangepaste DNA-synthese. We rapporteren een bijgewerkte versie van het protocol, met behulp van een goedkope benchtop fotolithografie-opstelling en commercieel beschikbare cholesterol gemodificeerde oligonucleotiden (CMO's) geïmplementeerd met behulp van een modulair formaat. CMO-gelabelde cellen hechten zich met hoge efficiëntie aan substraten met DNA-patronen. Deze aanpak kan worden gebruikt om meerdere celtypen tegelijk met hoge precisie te modelleren en om arrays van microtissues te creëren die zijn ingebed in een extracellulaire matrix. Voordelen van deze methode zijn de hoge resolutie, het vermogen om cellen in een driedimensionale micro-omgeving in te bedden zonder het micropatroon te verstoren en flexibiliteit bij het patroon van elk celtype.
De positionering van cellen ten opzichte van elkaar in een weefsel is een belangrijk kenmerk van de micro-omgeving1,2,3,4. Technieken die worden gebruikt om levende cellen in ruimtelijk gecontroleerde arrangementen te modelleren, zijn waardevolle experimentele hulpmiddelen voor het bestuderen van differentiatie4,5,6,7,8,celmotiliteit9,morfogenese10,11,12,metabolisme 13en cel-celinteracties7,14 . Er bestaan verschillende methoden voor het patroon van cellen, elk met hun eigen voor- ennadelen 3,4. Methoden die kleefeilanden van extracellulaire matrix (ECM) eiwitten creëren, zoals microcontactprinten en lasergesneden stencils, zijn eenvoudig en schaalbaar. Het is echter moeilijk om meer dan één of twee celtypen tegelijk te modelleren, omdat de kleefeigenschappen van verschillende celtypen met verschillende ECM-moleculen vaak vergelijkbaar zijn15,16,17. Complexere micropatrines kunnen worden gemaakt met lichtgeïnduceerde moleculaire adsorptie (LIMAP), een techniek die UV-licht gebruikt om PEG-gecoate gebieden te aborteren en daaropvolgende eiwitasorptie mogelijk te maken18,19. Dit proces kan worden herhaald om micropatrinterns met hoge resolutie te maken met meerdere celtypen. Kruisbinding van cellen aan de verschillende eiwitpleisters kan echter optreden, wat resulteert in een slechte patroonspecifi specificiteit19. Fysische methoden zoals het zaaien van cellen op micromechanische herconfigureerbare kweekapparaten kunnen gestructureerde coculturen creëren met dynamische controle, maar zonder de flexibiliteit in patroonontwerp van microcontactprinten of LIMAP14,8. In tegenstelling tot de andere technieken kan bioprinting driedimensionale arrangementen van cellen in hydrogelscreëren 20,21. Biogeprinte constructies hebben echter een veel lagere resolutie dan andere micropatterningtechnieken, met een gemiddelde functiegrootte in de orde van honderden micron22. Een ideale celpatroonmethode zou een hoge resolutie hebben, meerdere celtypen patroonen, apparatuur en reagentia gebruiken die gemakkelijk toegankelijk zijn en de mogelijkheid hebben om succesvolle patronen in een hydrogel in te bedden voor driedimensionale (3D) celcultuur. In dit artikel presenteren we CMO-DPAC, een celmicropatterningtechniek die de flexibiliteit en snelheid van DNA-hybridisatie gebruikt om celadhesie aan een substraat te targeten. Deze methode is aangepast van onze vorige protocollen23,24 om het betaalbaarder, modulair en toegankelijker te maken. Met behulp van het huidige protocol zou elk lab in staat moeten zijn om een volledig functioneel systeem op te zetten zonder gespecialiseerde apparatuur of expertise.
DNA Programmed Assembly of Cells (DPAC) is een krachtige tissue engineering-techniek die cellen met eencellige resolutie patronen met nauwkeurige controle over cel-celafstand en weefselgeometrie. In DPAC worden celmembranen versierd met DNA-oligonucleotiden (oligo's) met behulp van twee lipide-gemodificeerde oligo's die zijn ontworpen om te hybridiseren op het celmembraan. Omdat de oligo's worden geconjugeerd aan hydrofobe lipiden, verdelen ze zich snel naar het celmembraan25 waar ze hybridiseren, waardoor de netto hydrofobiciteit van de niet-covalent gebonden moleculen toeneemt en daardoor hun levensduur aan het celoppervlak verbetert26. De oligo's worden op het celoppervlak gepresenteerd op een manier waarop ze kunnen hybridiseren met complementaire oligo's op andere cellen of DNA-gefunctionaliseerde glazen dia's om gedefinieerde 2D- of 3D-celpatronen te creëren met voorgeschreven samenstelling, cel-celafstand en geometrie23,24. De microtissues met patroon kunnen enzymatisch van het oppervlak worden afgesplitst en ingebed in een hydrogel voor langdurige 3D-cultuur. Bij gebruik in combinatie met primaire cellen of stamcellen kunnen de resulterende verzamelingen cellen morfogenese ondergaan en zich vormen tot organoïden23,27,28. DPAC is toegepast om de dynamiek van het lot van volwassen neurale stamcellen te onderzoeken in reactie op concurrerendesignalen 6,29,om zelforganisatie van borstepitheelcellen23,28te bestuderen en om "weefselorigami" te genereren door mesenchymale condensatie27.
DPAC maakt de precieze plaatsing van meerdere celpopulaties mogelijk en heeft een aanzienlijk betere resolutie dan op extrusie gebaseerde bioprinters (in de orde van micron)22,23. Bovendien vereist DPAC, in tegenstelling tot ECM-gebaseerde patroonmethoden zoals microcontactprinten, geen differentiële hechting van de verschillende celtypen aan een ECM-gecoat oppervlak15,23. Het is ideaal voor het beantwoorden van vragen over hoe de samenstelling van een weefsel het gedrag beïnvloedt, hoe cellen meerdere cellulaire en micro-omgevingssignalen integreren bij het nemen van beslissingen6,29en hoe paren cellen met elkaar interageren. Een voordeel van deze methode ten opzichte van andere micropatterningmethoden is dat het kan worden gebruikt voor 3D-celkweek in een enkel beeldvormingsvlak, waardoor time-lapse studies van weefselzelforganisatie en organoïde morfogenese23,27,30worden vergemakkelijkt.
Ondanks deze voordelen heeft een succesvolle implementatie van DPAC de synthese van aangepaste oligonucleotide-reagentia en toegang tot gespecialiseerde apparatuur voor DNA-patronen23,24, waardoor de wijdverspreide acceptatie wordt beperkt. De optimale lipide-gemodificeerde oligo's (LMO's) die in het oorspronkelijke protocol worden gebruikt, moeten bijvoorbeeld op maat worden gesynthetiseerd, gemodificeerd met lignocerinezuur of palmitinezuur en gezuiverd26. Dit proces vereist het gebruik van een DNA-synthesizer en een hoogwaardig vloeistofchromatografie-instrument, evenals de aankoop van de bijbehorende reagentia zoals methylamine, een gereguleerde stof die onderworpen is aan zowel institutionele als federale regelgeving. Als alternatief kunnen LMO's op maat worden gekocht in bulk, maar dit vereist een aanzienlijke investering vooraf in de technologie.
Om deze beperkingen te overwinnen, hebben we een herziene versie van DPAC ontwikkeld die commercieel verkrijgbare cholesterol-gemodificeerde oligo's (CMO's) gebruikt in plaats van de op maat gesynthetiseerde LMO's. Om de kosten verder te verlagen en de flexibiliteit van het platform te vergroten, zijn we overgestapt op een modulair, drie-oligo systeem. In plaats van een nieuw cholesterolgemodificeerd oligo te bestellen voor elke unieke celpopulatie, kan een gebruiker van dit protocol in plaats daarvan dezelfde cholesterol-gemodificeerde oligo's ("Universal Anchor" en "Universal Co-Anchor") gebruiken voor elke celpopulatie en vervolgens een goedkoop, ongewijzigd oligo ("Adapter Strand") gebruiken dat hybridiseert met zowel het Universal Anchor als het amine-gefunctionaliseerde DNA op het oppervlak of de Adapter Strand van een ander celtype.
Een andere beperking van het oorspronkelijke DPAC-protocol was dat het de DIA's met DNA-patroon creëerde met behulp van een vloeistofprinter met hoge resolutie (bijv. Nano eNabler, BioForce Nanosciences)23,24. Hoewel dit instrument buitengewone resolutie- en lage reagensvereisten heeft, is het niet beschikbaar voor de meeste instellingen en heeft het een relatief lage afdruksnelheid (ongeveer 1 functiepatroon per seconde). Onlangs zijn twee fotolithografische methoden ontwikkeld om DNA-kenmerken op oppervlakken te modelleren. Viola en collega's gebruikten een polyacrylamide- en benzofenoncoating die covalent gebonden enkelstrengs DNA-oligo's bij blootstelling aan UV-licht30. Met behulp van deze methode waren ze in staat om weefselsteigers te maken die grootschalige, geprogrammeerde vormveranderingen ondergingen als gevolg van celcontractiliteit en zelforganisatie. Scheideler et al. ontwikkelden een methode die UV-blootstelling van een positieve fotoresist gebruikt om selectief amine-gemodificeerde DNA-oligo's bloot te stellen aan een aldehyde-gefunctionaliseerde dia29. Na het bakken en reductieve aminatie wordt het amine-gemodificeerde DNA covalent aan het oppervlak gebonden. Deze methode werd gebruikt om de reactie van volwassen neurale stamcellen op ruimtelijk gepresenteerde zelfvernieuwings- en differentiatiesignalen te onderzoeken. Dit artikel past het protocol van Scheideler et al. aan om de DNA-patronen te creëren die CMO-gelabelde cellen zullen vastleggen. Dit fotopatterning protocol kan worden uitgevoerd zonder gebruik te maken van een cleanroom. Het maakt gebruik van goedkope en commercieel verkrijgbare apparatuur die gemakkelijk kan worden ingezet op een tafel of zuurkast. Het gebruik van goedkope of doe-het-zelf (doe-het-zelf) fotolithografie-apparatuur verhoogt de toegankelijkheid voor onderzoekers zonder toegang tot cleanroomfaciliteiten en stelt onderzoekers in staat om de techniek uit te proberen zonder een grote investering van tijd of middelen31,32. Een betere resolutie en de uitlijning van meerdere DNA-kenmerken kan echter worden bereikt door gebruik te maken van de commerciële spincoater en maskeruitlijner die vaak wordt aangetroffen in cleanroomfaciliteiten.
Hier beschrijven we een methode om cellen te modelleren met eencellige resolutie met behulp van dna-gebaseerde adhesie. Ten eerste wordt fotopatterning met een positieve fotoresist gebruikt om patronen met hoge resolutie van amine-gemodificeerd DNA op een aldehyde-gemodificeerd glassubstraat te creëren. Vervolgens wordt de dia behandeld om niet-specifieke celaanhechting te verminderen en worden PDMS-stroomcellen gemaakt om cellen over patroongebieden te beperken. Cellen worden vervolgens gelabeld met korte DNA-oligonucleotiden die worden gefunctionaliseerd met cholesterol en als gevolg daarvan in het celmembraan worden ingebracht. De cellen worden vervolgens over de DNA-micropatrines gevloeid. Hybridisatie tussen het celoppervlak-DNA en het DNA op het glazen oppervlak resulteert in een specifieke hechting van de cellen aan het DNA-patroon. Niet-hechtende cellen worden weggespoeld, waardoor het aanhangende celpatroon wordt onthuld. Dit proces kan worden herhaald om meerdere celtypen te modelleren of om meerlaagse structuren te maken. Indien gewenst kunnen de cellen volledig worden ingebed in een ECM voor 3D-celkweek.
1. Ontwerpexperiment
- Plan het gewenste experiment, rekening houdend met de grootte van het kenmerk, de afstand tussen de functies, het aantal betrokken celtypen en de rangschikking van cellen ten opzichte van elkaar. Raadpleeg Supplemental File 1, een gids voor experimenteel ontwerp, en Supplemental File 2, dat voorbeeld oligosequenties bevat.
- Ontwerp fotomasker met behulp van computerondersteunde ontwerpsoftware. Een voorbeeldfotomasker is te zien in Aanvullend bestand 3.
- Teken een rechthoek met de afmetingen van een standaard microscoopglaasje (25 mm x 75 mm).
- Teken vier rechthoekige gebieden van 10 mm breed en 10 mm lang, gelijkmatig verdeeld over de dia.
- Teken binnen elk gebied kenmerken die de gewenste grootte, vorm en afstand voor het experiment hebben. Cellen zullen zich alleen aan deze kenmerken in het experiment hechten.
- Als u uitgelijnde fotomaskers voor meerdere celtypen wilt maken, maakt u een hoofdtekening met alle sets functies en slaat u versies op die overeenkomen met elk celtype.
- Bestel een hoge resolutie (minstens 20.000 dots per inch) transparantie fotomasker uit deze CAD tekening met de functies getekend in 1.2.3 transparant en de grotere gebieden zwart.
2. Fotopattern DNA op aldehyde-gefunctionaliseerde dia's (protocol aangepast van Scheideler et al.29 )
- Als u meerdere celtypen patroont, fabriceert u fiduciële markers op de aldehyde-gefunctionaliseerde dia vóór DNA-patronen om de uitlijning van kenmerken te vergemakkelijken. Alternatieve methoden voor het maken van fiduciale markers worden voorgesteld in Supplemental File 1.
- Als u metalen fiduciale markeringen wilt maken, past u S1813 positieve fotoresist toe zoals beschreven in stappen 2.3 - 2.11. Gebruik een fotomasker dat grote functies bevat die later gemakkelijk uit te lijnen zijn. Neem deze functies op in het ontwerp van de fotomaskers die zullen worden gebruikt voor DNA-patronen.
- Deponeer een dunne film (100 Angstroms) van titanium op de dia met behulp van elektronen-pistool verdamping29. Verwijder overtollig metaal en fotoresist met aceton en ga vervolgens verder met de DNA-fotopatterning.
- Bereid een 20 μM-oplossing van een 5'-amine-gemodificeerd oligo in DNA-buffer (50 mM natriumfosfaat in water, pH = 8,5). Zie Aanvullend bestand 2 voor voorgestelde oligosequenties.
OPMERKING: Het is mogelijk om slechts 5 μM amine-gemodificeerd oligo te gebruiken voor sommige patronen en toepassingen, dus de DNA-concentratie van het oppervlak moet mogelijk worden geoptimaliseerd. - Verwarm een kookplaat voor op 100 °C.
- Gebruik dubbelzijdige tape of een vacuüm om een aldehyde-gefunctionaliseerde glasplaat aan de rotor van een spincoater te bevestigen.
LET OP: Het losmaken van de schuif tijdens het spincoaten is een veiligheidsrisico. Gebruik de spincoater altijd in een bijgesloten container met deksel, zoals een acryldoos.
OPMERKING: Label een hoek van de dia met behulp van een diamantscribent of een soortgelijk werktuig om het glas te krassen. Dit helpt bij de identificatie en oriëntatie van dia's nadat de fotoresist is weggespoeld. - Gebruik een wegwerppipet om de positieve fotoresist op de aldehydeplaat te laten vallen. Voeg voor gelijkmatige coatings kleine druppels van de fotoresist toe over de dia, in plaats van één grote druppel in het midden(aanvullende figuur 1A).
- Gebruik de spincoater om de glijbaan 30 s op 3000 tpm te laten draaien.
- Plaats de dia op een kookplaat van 100 °C gedurende 1,5 min (zacht bakken) om de fotoresistent te kruisen.
- Verwijder de dia van de kookplaat. Plaats een fotomasker met de voor dit experiment gewenste kenmerken bovenop de dia en weeg het fotomasker met een stuk glas(aanvullende figuur 1B, C). Bedek de hele opstelling in een ondoorzichtige doos(aanvullende figuur 1D). Belicht met een UV-lamp (365 nm golflengte, 360 mW, 5 inch van dia, totale stralingsenergiedichtheid 100 mJ / cm2) gedurende 2 minuten.
OPMERKING: UV-licht zal de polymeerbindingen in de fotoresist onder transparante gebieden van het fotomasker verbreken, waardoor gebieden ontstaan waar DNA zich later kan hechten. - Ontwikkel de dia door 3-5 minuten onder te dompelen in de ontwikkelaarsoplossing(aanvullende figuur 1E).
- Spoel overtollige ontwikkelaarsoplossing weg met water. Droog onder een stroom lucht of stikstof. (Aanvullende figuur 1F).
- Bevestig dat de fotolithografie succesvol was door naar de dia onder de microscoop te kijken. Omdat de fotoresist UV-lichtgevoelig is, voert u deze stap snel uit en slaat u de dia vervolgens op in het donker terwijl u andere dia's voorbereidt (indien van toepassing).
OPMERKING: Een dia met een succesvol patroon moet voor elk kenmerk scherp gedefinieerde randen hebben, geen scheuren en geen functievervorming aan de randen. Voorbeelden van correcte en onjuiste fotolithografie zijn te vinden in aanvullende figuur 2A. Zie tabel 1 voor suggesties voor het oplossen van problemen als fotolithografie niet de gewenste functiekwaliteit biedt. - Voeg een druppel van de 20 μM amine-gemodificeerde oligo-oplossing (stap 2.1) toe aan elk fotogepatterd gebied van de dia. Gebruik een pipetpunt om de druppel voorzichtig over het hele gebied te verspreiden, waarbij u voorzichtig moet oppassen dat u niet aan de dia krabt. (Aanvullende figuur 1G).
- Bak de dia in een oven van 65-70 °C totdat de DNA-oplossing volledig is opgedroogd op het diaoppervlak (ongeveer 1 uur).
- Voer reductieve aminatie uit door de patroonvormige, gebakken dia's in een celcultuurschaal van 15 cm te plaatsen en plaats deze in een zuurkast bovenop een shaker. Weeg 100 mg natriumboorhydride af. Voeg in een zuurkast 40 ml fosfaat-gebufferde zoutoplossing (PBS) toe, meng voorzichtig en voeg toe aan de schaal met de patroonglaasjes. Laat de reactie 15 minuten doorgaan met zacht schudden.
OPMERKING: Het amine op het oligo vormt eerst een Schiff-basis met de aldehyden op het diaoppervlak. Dit is een omkeerbare covalente binding die moet worden omgezet in een onomkeerbare binding voor gebruik in DPAC. Toevoeging van een reductiemiddel (natriumboorhydride) zet de Schiff-base om in een secundair amine door reductieve aminatie.
LET OP: De reactie van natriumboorhydride met water creëert waterstofgas en zal dit blijven doen gedurende uren of dagen nadat de reactie begint. Voer de reductieve aminatiestap uit in een zuurkast en bewaar al het afval van natriumboorhydride-oplossing gedurende ten minste 24 uur in een open of los afgedekte container in de zuurkast. - Verwijder niet-gereageerd DNA door tweemaal te wassen met 0,1% natriumdodecylodeylsulfaat (SDS) in water en vervolgens drie keer met gedestilleerd water. Droog de glijbaan onder een stroom stikstof of lucht.
- Spoel de dia af met aceton om de resterende fotoresist te verwijderen.
OPMERKING: Op dit punt is het DNA onomkeerbaar en covalent aan de dia bevestigd en zijn alle niet-gereageerde aldehyde-functionele groepen omgezet in alcoholen. De fotoresist is niet meer nodig. - Als er meerdere oligo's een patroon hebben, keert u terug naar stap 2.4, lijnt u het fotomasker uit met fiduciële tekens en herhaalt u dit.
OPMERKING: Het experiment kan hier worden gepauzeerd. Bewaar dia's in een vacuümsiccator. Onder droge omstandigheden kunnen de platen tot 3 maanden worden bewaard zonder kwaliteitsverlies.
3. Maak slide hydrofoob (optioneel) (protocol aangepast van Todhunter et al.24 )
OPMERKING: Het is voordelig, maar niet vereist, om de oppervlaktechemie van de dia te wijzigen om deze meer inert en hydrofoob te maken. Niet-specifieke celaanhechting wordt op deze oppervlakken verminderd33, waardoor de niet-specifieke binding van cellen aan niet-patroonachtige delen van de dia wordt verlicht. Bovendien, als de cellen met een patroon uiteindelijk in een hydrogel worden ingebed en van de dia worden overgebracht, is de oppervlaktebehandeling essentieel voor een betrouwbare beweging van de met cellen beladen hydrogel over de dia zonder vervorming of scheuren. Silaniseren met (tridecafluor-1,1,2,2-tetrahydrooctyl) dimethylchloorsilaan resulteert in de aanwezigheid van hydrofobe fluoroalkylgroepen op het diaoppervlak.
LET OP: Voer alle stappen vanaf 3.1 uit in een chemische zuurkast om blootstelling aan azijnzuur en methyleenchloridedampen te voorkomen.
- Spoel de glijd af met 10% azijnzuur en droog vervolgens onder een luchtstroom.
- Bereid in een glazen Coplin-pot een oplossing van 60 ml methyleenchloride (dichloormethaan), 0,6 ml triethylamine en 0,6 ml (tridecafluor-1,1,2,2-tetrahydrooctyl) dimethylchloorsilaan. Roer met een metalen spatel om te mengen.
OPMERKING: Deze reagentia zijn gevoelig voor water. Ze moeten onder droge omstandigheden worden opgeslagen en zo vers mogelijk worden gebruikt. - Voeg de dia toe aan de Coplin-pot met de silaanoplossing. Plaats coplin pot op een orbitale shaker (ingesteld op 60-80 rpm) en laat de reactie van het silaan en de dia gedurende 15 minuten verlopen.
- Gebruik een metalen tang om de glijbaan uit de silaanoplossing te verwijderen. Dompel de glijpartij gedurende 1 minuut onder in een Coplin-pot met methyleenchloride om overtollig silaan van de dia te verwijderen.
- Dompel de dia onder in een conische buis van 50 ml met ethanol. Agiteren. Dompel de glijbaan onder in een conische buis van 50 ml met gedestilleerd water. Agiteren.
OPMERKING: Methyleenchloride en water zijn niet mengbaar, dus een ethanolspoeling is nodig om overtollig methyleenchloride te verwijderen voor de laatste waterspoeling. - Haal de glijbaan uit het water en inspecteer deze. De glijbaan moet vrij droog zijn, waarbij eventuele waterdruppels een contacthoek van meer dan 90° hebben. Laat de glijden volledig drogen en bewaar ze in een vacuümsiccator tot gebruik.
OPMERKING: Het experiment kan hier worden gepauzeerd. Bewaar de dia onder droge omstandigheden.
4. Bereid PDMS-stroomcellen en dia voor op experimenten
OPMERKING: Rechthoekige PDMS-stromingscellen worden gebruikt om de cellen te concentreren over de patroongebieden van de dia. Voor experimenten gekweekt in 3D vormen de flowcellen een mal voor de hydrogel.
- Maak SU-8 master om te gebruiken als mal voor PDMS-flowcellen.
- Verwarm de kookplaat voor tot 95 °C.
- Voeg 5 ml SU-8 2075 toe aan een siliciumwafer.
- Spin coat de SU-8 op de wafer bij 500 rpm voor 10s, gevolgd door 1.000 rpm voor 30s. Dit moet functies creëren tot 240 μm in hoogte34.
- Bak de wafel zachtjes op de kookplaat gedurende minstens 45 min.
- Verwijder de wafer van de kookplaat. Plaats het fotomasker (zie Aanvullend bestand 4) (emulsie met de zijkant naar beneden) bovenop de wafer en verzwaar het met een glazen schijf om contact tussen het fotomasker en de dia te garanderen.
- Belicht met UV-licht (365 nm) voor een stralingsenergiedichtheid van 350 mJ/cm2.
- Bak wafel op de kookplaat gedurende 12-15 min.
- Plaats de wafer in een brede glazen container. Dekwafer af met su-8 developer oplossing. Plaats op een shaker en ontwikkel tijdens het roeren gedurende ten minste 15 minuten.
- Gebruik een tang om de wafer uit de ontwikkelaarsoplossing te verwijderen. Spoel gedurende 5 s door meer ontwikkelaarsoplossing uit een spuitfles te spuiten. Spray met isopropylalcohol om af te spoelen. Als er een wit neerslag verschijnt, brengt u de wafer terug naar de ontwikkelaarsoplossing en ontwikkelt u deze langer.
- Droge wafer onder een stroom lucht of stikstof.
- Bakschuif gedurende 5 min.
OPMERKING: Zodra de masterwafer is gemaakt, kan deze voor onbepaalde tijd worden hergebruikt zolang de functies intact blijven.
- Pdms voorbereiden.
- Voeg in een weegboot polydimethylsiloxaanelastomeer en crosslinker toe in een verhouding van 10:1 (bij massa). Roer krachtig om een gelijkmatige menging te garanderen.
- Ontgas de PDMS in een vacuümsiccator gedurende 15-30 minuten totdat er geen bellen meer zichtbaar zijn.
- Plaats de masterwafer in een 15 cm weefselkweekschaal. Giet PDMS over de wafer. Als er bellen verschijnen, ontgass dan een paar minuten in een vacuümsiccator.
- Bak 3 uur in de oven van 60 °C.
OPMERKING: Na het bakken kunnen PDMS-flowcellen voor onbepaalde tijd op de tafel worden bewaard.
- Bereid PDMS-stroomcellen voor op het experiment.
- Kort voordat u een CMO-DPAC-experiment start, knipt u het vereiste aantal PDMS-stroomcellen uit de masterwafer. Plasma oxideert met 10 cc/min kamerlucht gedurende 90 s om het oppervlak hydrofiel te maken.
- Knip elke afzonderlijke stroomcel uit zodat er aan elke kant 1-2 mm PDMS overblijft en snijd vervolgens de boven- en onderkant van de stroomcel open om een inlaat en uitlaat te creëren.
- Haal dia met patronen op die is gemaakt in stap 2 en 3. Lijn uit op het fotomasker.
- Gebruik het fotomasker als referentie en plaats de PDMS-stroomcellen op de dia op de locatie van elk patroongebied.
- Voeg 50 μL fosfaat gebufferde zoutoplossing (PBS) + 1% runderserumalbumine (BSA) toe aan de inlaat van elke stroomcel, zoals weergegeven in aanvullende figuur 1H. Controleer of de stroomcel volledig gevuld is door de PBS + 1% BSA en dat er geen grote bubbels zijn. Ga onmiddellijk verder met stap 5 en 6.
OPMERKING: Blokkeren met BSA minimaliseert de niet-specifieke celhechting aan het diaoppervlak.
5. Lift en label cellen met cholesterol-gemodificeerd DNA
- Bereid de cholesterol-gemodificeerde DNA-oplossingen voor.
- Meng voor elke set cellen in het experiment 3 μL van een 100 μM-stamoplossing van de cholesterolge gemodificeerde Universal Anchor Strand met 3 μL van een 100 μM-stamoplossing van een adapterstreng. Incubeer gedurende 1 minuut. Dit zal de oligo's pre-hybridiseren. Voeg 69 μL fosfaat-gebufferde zoutoplossing (PBS) toe om een 4 μM Universal Anchor + Adapter-oplossing te creëren.
- Voeg voor elke set cellen in het experiment 3 μL van een 100 μM Universele cholesterol-gemodificeerde Co-Anchor Strand stockoplossing toe aan 12 μL PBS, waardoor een 20 μM-oplossing ontstaat.
- Bereid de eencellige suspensie(suspensie) voor.
- Gebruik voor aanhankelijke cellen trypsine of een ander dissociatiemiddel om de cellen uit de kweekkolf te verwijderen. Voeg kweekmedia toe om de trypsine te neutraliseren en centrifugeer om de cellen te pelleteren. Voor niet-hechtende cellen, verzamel de celsuspensie en centrifugeer om de cellen te pelleteren.
- Resuspend de celkorrel in 1 ml ijskoude PBS of serumvrije media. Breng 1-3 miljoen cellen over in een microcentrifugebuis van 1,5 ml. Centrifugeer bij 160 x g gedurende 4 min.
OPMERKING: Als het gebruikte celtype gevoelig is voor klonteren/aggregeren, gebruik dan PBS zonder calcium- en magnesiumionen voor alle wasstappen om ongewenste celaggregatie te verminderen. Als levensvatbaarheid een bijzondere zorg is voor het celtype dat wordt gebruikt, gebruik dan serumvrije media in plaats van PBS. Media die foetaal runderserum bevatten, worden niet aanbevolen voor celetikettering omdat dit de opname van lipide-gemodificeerde oligo's kan belemmeren. 35
- Label de cellen met cholesterol-gemodificeerde oligo's.
- Resuspend de celkorrel in 75 μL ijskoude PBS of serumvrije media. Bewaar de cellen in een ijsemmer tijdens het etiketterings- en wasproces om de levensvatbaarheid van de cel te maximaliseren en het verlies van de cholesterol-gemodificeerde oligo's van het celoppervlak te minimaliseren.
OPMERKING: Het opnieuw opsuspenden van de cellen voordat het DNA wordt toegevoegd, zorgt ervoor dat de verdeling van DNA uniform is over de celpopulatie. - Voeg de 75 μL van de 4 μM Universal Anchor + Adapter-oplossing die in stap 5.1.1 is gemaakt, toe aan de microcentrifugebuis met de celsuspensie. Meng grondig door te pipetteren. Incubeer gedurende 5 minuten op ijs.
- Voeg 15 μL van de Universele Co-Anchor Oplossing toe aan de microcentrifugebuis. Meng grondig door te pipetteren. Incubeer gedurende 5 minuten op ijs.
- Verwijder overtollige oligo's uit de celsuspensie. Voeg 1 ml ijskoude PBS of serumvrije media toe aan de microcentrifugebuis. Meng met een P1000 pipet. Centrifugeer bij 160 x g gedurende 4 minuten bij 4 °C. Gooi het supernatant weg. Herhaal dit nog twee keer.
OPMERKING: Als cellen gevoelig zijn voor klonteren, passeer de celsuspensie door een filter van 40 μm voor de laatste wasbeurt. Als cellen gevoelig zijn voor adsorptie aan de zijkant van de microcentrifugebuis, overweeg dan om de buis vooraf te blokkeren met caseïne.
- Resuspend de celkorrel in 75 μL ijskoude PBS of serumvrije media. Bewaar de cellen in een ijsemmer tijdens het etiketterings- en wasproces om de levensvatbaarheid van de cel te maximaliseren en het verlies van de cholesterol-gemodificeerde oligo's van het celoppervlak te minimaliseren.
6. Patroon de DNA-gelabelde cellen
- Resuspend de cellen in ijskoude PBS of serumvrije media om een celdichte oplossing van ten minste 25 miljoen cellen / ml te creëren.
OPMERKING: Voor één dia met vier van de PDMS-flowcellen van 10 mm x 15 mm x 200 μm die in stap 4 worden beschreven, is ongeveer 100 μL van deze dichte celsuspensie vereist. Hoewel de meeste van deze cellen zich niet aan het patroon zullen houden en uiteindelijk zullen worden weggegooid, verbetert het hebben van een extreem geconcentreerde oplossing van cellen over het patroon de efficiëntie van celpatronen dramatisch. - Pak de glijbaan op en kantel hem iets. Voeg 25 μL celsuspensie toe aan de inlaat van elke stroomcel op de patroondia. Verwijder de PBS + 1% BSA-oplossing uit de uitlaat, zodat de celsuspensie de PDMS-stroomcel kan vullen. Incubeer op ijs of bij kamertemperatuur gedurende 30 s.
OPMERKING: Op dit punt zou het kijken naar de stroomcel onder een microscoop dicht opeengepakte cellen moeten laten zien met weinig tot geen openingen zichtbaar tussen cellen. Zie aanvullende figuur 2B. - Aspirateer 5 μL celsuspensie uit de uitlaat van de dia en voeg deze terug toe aan de inlaat. Herhaal dit 10 keer per stroomcel.
OPMERKING: De hechting van CMO-gelabelde cellen aan de DIA met DNA-patroon is bijna onmiddellijk. Door de cellen meerdere keren over het patroon te laten stromen, neemt de kans toe dat een cel over een bepaalde DNA-plek stroomt en wordt gevangen. - Pipetteer PBS of serumvrije media voorzichtig in de inlaat van elke stroomcel om overtollige cellen uit te spoelen. Verzamel de celsuspensie uit de uitlaat. Herhaal dit 2-4 keer of totdat een visuele inspectie van het dia onder de microscoop bevestigt dat er geen overtollige cellen overblijven.
OPMERKING: Het kan voordelig zijn om de overtollige cellen van de eerste wasbeurt te redden. Als de patroonefficiëntie onbevredigend is, kunnen de overtollige cellen worden gecentrifugeerd en opnieuw worden uitgegeven in een lager volume PBS om een meer celdichte oplossing te creëren, en vervolgens kan het proces worden herhaald vanaf stap 6.2. - Herhaal stap 6.1-6.4 voor elke set cellen in het patroon. Voor patronen waarin meerdere celtypen rechtstreeks worden gemodelleerd door de oppervlaktesjabloon, begint u met het minst voorkomende celtype van het patroon en eindigt u met het meest voorkomende celtype.
OPMERKING: Het is raadzaam om elke ronde van cellulaire assemblage sequentieel te doen in plaats van de cellen te bundelen, zelfs in omstandigheden waarin de cellen allemaal zijn gelabeld met orthogonale DNA-sequenties. Het poolen van de cellen verdunt effectief elke celpopulatie en vermindert de efficiëntie van patronen. - Nadat de laatste ronde van celassemblage is voltooid, zullen de volgende stappen variëren op basis van het specifieke experiment. Als de cellen bedoeld zijn om op het glas te blijven, voegt u media toe aan een petrischaal met de dia en gebruikt u vervolgens voorzichtig een tang om de PDMS-stroomcellen van de dia te duwen. Als de cellen worden ingebed in een hydrogel en in 3D worden gekweekt, gaat u verder met stap 7.
7. Overdracht in hydrogel voor 3D-cultuur (optioneel)
- Bereid een hydrogelprecursoroplossing met 2% DNase.
OPMERKING: De samenstelling van de oplossing varieert op basis van de experimentele opstelling. Matrigel en mengsels van Matrigel en collageen ik werk goed in dit protocol, maar ook andere hydrogels zijn mogelijk. - Voeg 50 μL hydrogeloplossing met 2% DNase toe aan de inlaat van elke stroomcel. Adem de overtollige vloeistof uit de uitlaat op en drijf de hydrogeloplossing in de stroomcel. Voor viskeuze hydrogelprecursoren kan het nodig zijn om de schuif iets te kantelen om de hydrogel in de stroomcel te laten stromen.
- Incubeer de dia bij 37 °C gedurende 30-45 minuten (afhankelijk van de hydrogelgelationkinetiek) om de hydrogel te laten uitzetten en de op DNA gebaseerde hechting tussen de cellen en het oppervlak te splitsen.
- Verwijder elke stroomcel van de dia en plaats deze bovenop de hydrogelvoorloperoplossing.
- Voeg 50 μL hydrogelvoorloper toe aan een put van een 2-well kamer slide of een 6-well plaat.
- Pipetteer 10 μL PBS aan weerszijden van elke stroomcel.
- Gebruik een scheermesje of een fine-point pincet om de PBS over de volledige lengte van de stroomcel te verdelen en til vervolgens voorzichtig de zijkanten van de stroomcel op zodat de PBS onder de hydrogel door stroomt.
OPMERKING: Dit zal de hydrogel over de dia "laten zweven", waardoor overdracht mogelijk is zonder vervorming of scheuren. - Gebruik een scheermesje om de stroomcel voorzichtig naar de rand van de glasplaat te verplaatsen.
- Keer de dia om. Duw met het scheermesje de stroomcel van de glijbaan zodat deze bovenop het scheermesje landt.
- Pluk de flowcel van het scheermesje met een gebogen tang. Keer de stroomcel om zodat de cellen zich aan de onderkant bevinden en plaats deze vervolgens bovenop de druppel hydrogelvoorloperoplossing.
- Herhaal stap 7.4.1 - 7.4.6 voor elke stroomcel.
- Incubeer gedurende ten minste 30 minuten zodat de hydrogel die de patrooncellen bevat, zich kan binden aan de hydrogel-onderlaag, wat resulteert in de volledige inbedding van de patrooncellen.
- Verwijder de PDMS-stroomcel.
- Voeg voldoende media toe om de PDMS-stroomcel onder te dompelen.
OPMERKING: De instroom van media zal de hechting tussen de hydrogel en de PDMS-stroomcel losser maken. - Gebruik een gebogen tang, georiënteerd langs de lange as van de stroomcel, om de stroomcel zachtjes te duwen totdat deze eraf springt en in de media zweeft. Verzamel de stroomcel met een tang en gooi weg.
OPMERKING: Voor optimale resultaten spreidt u de gebogen tang en oefent u zachte druk uit op de wanden van de PDMS-stroomcel. Oefen kracht uit in de richting van de lange as van de stroomcel.
- Voeg voldoende media toe om de PDMS-stroomcel onder te dompelen.
8. Bevestig succesvolle labeling van cellen met CMO (optioneel, voor probleemoplossing)
- Bestel een fluorescerend gemodificeerd (FAM of AF647) oligonucleotide dat complementair is aan de oppervlakteadhesiesequentie van de adapterstreng die in het experiment wordt gebruikt.
- Label cellen met CMO-DNA en spoel overtollig DNA uit zoals beschreven in stap 5. Resuspend in 200 μL ijskoude PBS.
- Maak een 4 μM-oplossing van het fluorescerend gelabelde complementaire oligonucleotide in PBS. Voeg 200 μL van deze oplossing toe aan de celsuspensie. Incubeer op ijs gedurende 5 min.
- Voeg 1 ml ijskoude PBS toe. Meng. Centrifugeer de cellen om ze te pelleteren. Verwijder supernatant. Herhaal dit proces nog twee keer om DNA dat niet is gehybridiseerd uit te spoelen.
- Voer analytische flowcytometrie uit om de aanwezigheid van DNA op het celoppervlak te kwantificeren.
- Analyseer op een flowcytometer controlecellen die niet zijn gelabeld met DNA. Stel poorten op basis van deze populatie.
- Analyseer CMO-gelabelde cellen die zijn behandeld met een fluorescerend gelabeld complementair oligonucleotide.
- Bereken de gemiddelde fluorescentie-intensiteit.
Dit protocol maakt het mogelijk om cellen in 2D en 3D te modelleren met hoge precisie en zonder het gebruik van aangepaste reagentia of dure cleanroomapparatuur. Figuur 1 toont een overzicht van het protocol. Ten eerste worden DNA-gefunctionaliseerde dia's gemaakt door middel van fotolithografie. Vervolgens worden cellen gelabeld met CMO's. De cellen worden vervolgens over de dia gestroomd, waar ze zich alleen hechten aan de DNA-gefunctionaliseerde gebieden van de dia. Nadat overtollige cellen zijn weggespoeld, wordt het gewenste patroon van cellen onthuld. Deze cellen kunnen op de dia worden gekweekt of ingebed in een hydrogel met DNase en van de dia worden overgebracht voor 3D-celkweek.
Labeling van cellen met CMO's maakt hun hechting aan de DNA-patroondia mogelijk(figuur 2). Ten eerste is de cholesterol-gemodificeerde Universal Anchor Strand voorgehybridiseerd met de Adapter Strand. Vervolgens wordt de Universal Anchor + Adapter-oplossing 1:1 gemengd met de celophanging. Het cholesterol op het Universal Anchor + Adapter complex wordt in het celmembraan geplaatst. Toevoeging van de cholesterol-gemodificeerde Universal Co-Anchor Strand, die hybridiseert met de Universal Anchor Strand, verbetert de stabiliteit van het CMO-complex in het celmembraan door de netto hydrofobiciteit van het complex te verhogen26. Na het uitwassen van het overtollige DNA uit de celsuspensie, worden de cellen over de dia gestroomd. Hybridisatie tussen de adapterstreng en de oppervlakte-DNA-streng resulteert in hechting van cellen aan de DNA-patroongebieden van de dia.
Het patroon van de cellen wordt gecreëerd door fotolithografie te gebruiken om de aanhechting van amine-gemodificeerde DNA-oligo's aan specifieke gebieden van een aldehyde-gemodificeerde glasplaat29 te beperken (Figuur 3A). Positive photoresist wordt gespind op een aldehyde-gefunctionaliseerde dia. Vervolgens wordt een transparant fotomasker bovenop de dia geplaatst en wordt de dia blootgesteld aan UV-licht. Na ontwikkeling zijn de gebieden van de dia die werden blootgesteld aan UV-licht niet langer bedekt met fotoresist en hebben dus aldehydegroepen blootgelegd. Een 20 μM-oplossing van amine-gemodificeerde DNA-oligo's wordt vervolgens op de dia gedropt en verspreid om de patroongebieden te bedekken. Bakken gevolgd door reductieve aminatie resulteert in een covalente binding tussen het amine-gemodificeerde DNA en de dia. Opmerkelijk is dat dit proces kan worden herhaald om meerdere oligo's te modelleren zonder enig verlies van functionaliteit van de eerder gemodeleerdeoligo's ( Figuur 3B). Er moet echter voor worden gezorgd dat overlappende patronen worden vermeden, wat resulteert in de aanwezigheid van beide oligo's bij een verminderde concentratie(aanvullende figuur 3). Meerdere celpopulaties kunnen sequentieel worden gemodelleerd door adapterstrengen te gebruiken die verschillen in hun modulaire domein (de 20 basen die het dichtst bij het 3'-uiteinde liggen).
Hoewel dit fotopatterningprotocol is ontwikkeld door Scheideler et al. in het kader van een cleanroom, hebben we aangetoond dat het mogelijk is om vergelijkbare resultaten te bereiken met een goedkope, "home-brew" fotolithografie-opstelling die gemakkelijk in een chemische zuurkast past. De opstelling omvat een spincoater van $ 400 gemaakt van een DC-motor, digitale controller en cd-cakedoos, evenals een UV-lamp die is samengesteld uit individuele componenten en is ondergebracht in een hergebruikte naaldencontainer(aanvullende figuur 1). Het belangrijkste voordeel van de home-brew fotolithografie-opstelling is dat het zeer betaalbaar is (< $ 1000 voor alle apparatuur) terwijl het nog steeds functies van een enkele cel kan maken. Het gebruik van goedkope apparatuur heeft echter zijn beperkingen - het is bijvoorbeeld een grotere uitdaging om fiduciale markers nauwkeurig uit te lijnen om meerdere DNA-oligo's te patroon zonder gebruik te maken van een maskeruitlijner. We raden deze goedkope fotolithografie-opstelling aan voor laboratoria die geen gemakkelijke toegang hebben tot een schone kamer of die deze methode willen proberen zonder een grote investering.
Om optimale omstandigheden voor DNA-geprogrammeerde celadhesie te identificeren, varieerden we systematisch de concentraties van DNA-strengen op celoppervlakken en maten we de efficiëntie van celadhesie aan DNA-gemodificeerde glasoppervlakken. De concentratie van Universal Anchor + Adapter Strand en Universal Co-Anchor in etiketteeroplossingen varieerde over verschillende ordes van grootte(figuur 4A,B),wat resulteerde in 104 - 106 DNA-complexen per cel(aanvullende figuur 4). Celadhesie was dosisafhankelijk, met minimale celadhesie aan het DNA-patroon wanneer cellen werden gelabeld met CMO's in een concentratie van 0,05 μM of minder, en hoge bezetting bij een concentratie van 2,5 μM en hoger. Daarom gebruikten we in de meeste experimenten een 2 μM-oplossing van Universal Anchor + Adapter Strand en 2 μM-oplossing van Universal Co-Anchor. De celadhesie zou naar verwachting ook afnemen als de hoeveelheid DNA die op het glasoppervlak wordt gebruikt29 afneemt of als de mismatches tussen de adapterstreng en de oppervlaktestreng toenemen. Meer informatie over het ontwerp van adapterstrengsequenties vindt u in Aanvullend bestand 2. CMO-etikettering met adapterstrengen zonder CpG-herhalingen stimuleerde TLR9 niet in HEK-cellen die muis-TLR9 tot expressie brengen(aanvullende figuur 5).
We geven verschillende demonstraties dat het herziene protocol reproduceerbare en efficiënte DNA-geprogrammeerde celadhesie biedt. Menselijke navelstreng endotheelcellen (HUVECs) gelabeld met CMO's kleefden bijvoorbeeld met hoge efficiëntie aan DNA-patronen. HUVEC's met een CMO-label en huvec's met LMO-label(figuur 5A). Cellen met een patroon met CMO-DPAC behielden hun levensvatbaarheid en functionaliteit. Cellen gelabeld met CMO's werden gekleurd door calceïne AM en ethidium homodimeer om de levensvatbaarheid te beoordelen (aanvullende figuur 6). Verschillen in levensvatbaarheid in vergelijking met niet-gelabelde controlecellen waren klein (94% versus 97%). Enkele MDCK's met een patroon via CMO-DPAC en overgebracht naar Matrigel konden zich na 5 dagen cultuur correct vermenigvuldigen en polariseren(figuur 5B). DPAC biedt ook een middel om patronen van cellen in de derde dimensie uit te werken(figuur 5C). Meerlagige, meercellige aggregaten kunnen bijvoorbeeld worden gemaakt door afwisselende lagen cellen te markeren met complementaire CMO's(figuur 5C). Deze experimenten tonen aan dat het protocol reproduceerbaar is, de levensvatbaarheid of functionaliteit van cellen niet negatief beïnvloedt en cellulaire patronen oplevert die met succes kunnen worden gekweekt binnen een enkel beeldvormingsvlak in een 3D-ECM.
Door orthogonale DNA-sequenties te leveren om celadhesie te sturen, biedt DPAC een middel om meerdere celtypen op één oppervlak te modelleren. Om deze functie van DPAC te implementeren, moeten DNA-patronen die door fotolithografie worden gegenereerd, ten opzichte van elkaar worden uitgelijnd. Metalen fiduciaire markers die op de dia werden afgezet, maakten de uitlijning van meerdere fotomaskers mogelijk en dus de patroonvorming van meerdere celtypen tegelijk. MCF10A's gekleurd met verschillende unieke kleurstoffen werden gelabeld met orthogonale CMO's en een patroon om een visualisatie van de UC Berkeley- en UCSF-logo's te maken(Figuur 6). Dit experiment toont aan dat meerdere unieke celpopulaties met hoge precisie en zonder kruisbesmetting aan elkaar kunnen worden gemodelleerd.
Succesvolle patrooning van cellen met behulp van CMO-DPAC vereist hoogwaardige fotolithografie, voldoende concentratie van oligo op het celoppervlak, een hoge dichtheid van cellen over het patroon en voldoende wassen. Het mislukken van een van deze stappen heeft invloed op het eindresultaat. Aanvullende figuur 2 bevat voorbeeldafbeeldingen van correcte en onjuiste fotolithografie (aanvullende figuur 2A), de gewenste celdichtheid over het patroon om volledig bezette patronen te creëren (aanvullende figuur 2B), het verlies van patrooncellen als gevolg van te krachtig pipetteren tijdens volgende stappen van DPAC (aanvullende figuur 2C) en ongewenste samenklontering van cellen (aanvullende figuur 2D). Tabel 1 bevat een lijst met veelvoorkomende storingspunten en de voorgestelde probleemoplossing. Het gebruik van fluorescerende complementaire oligo's wordt aanbevolen als een hulpmiddel voor het oplossen van problemen om de aanwezigheid van DNA met patronen op de dia en de aanwezigheid van CMO's op het celoppervlak te bevestigen door middel van flowcytometrie (zie stap 8 van het protocol).
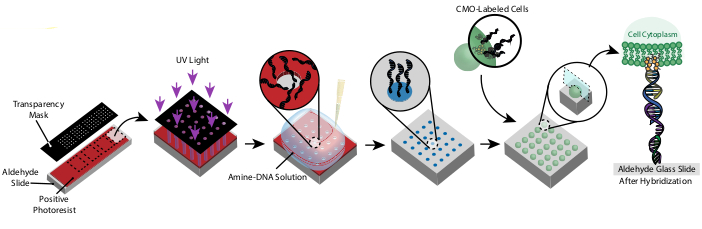
Figuur 1: Overzicht van het CMO-DPAC protocol. Eerst wordt een dia met DNA-patroon gemaakt door een aldehyde-gefunctionaliseerde glasplaat te coaten met een positieve fotoresist, deze te bedekken met een transparantiemasker in het gewenste patroon en bloot te stellen aan UV-licht. De UV-blootgestelde fotoresist wordt weggespoeld met de ontwikkelaar, waardoor blootgestelde delen van de aldehyde-dia achterblijven en de binding van amine-gefunctionaliseerd DNA aan het oppervlak mogelijk wordt. Cellen worden vervolgens gelabeld met CMO's en over het oppervlak gestroomd. Het DNA op het celmembraan hybridiseert naar het DNA aan het oppervlak, wat resulteert in hechting. Klik hier om een grotere versie van deze figuur te bekijken.
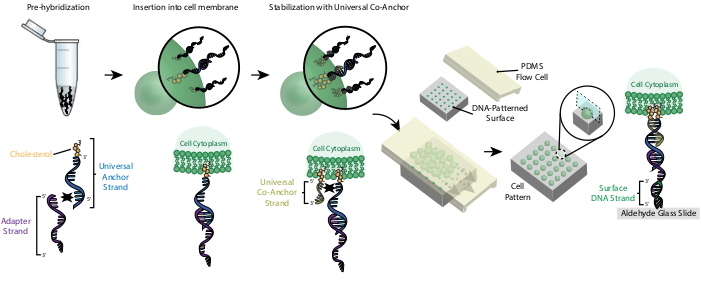
Figuur 2: Cellen worden gelabeld met CMO's in een stapsgewijs proces. Ten eerste is de cholesterol-gemodificeerde Universal Anchor Strand voorgehybridiseerd met de Adapter Strand. Vervolgens wordt de Universal Anchor + Adapter-oplossing gemengd met de celophanging. Het cholesterol op het Universal Anchor + Adapter complex wordt in het celmembraan geplaatst. Na incubatie wordt de cholesterol-gemodificeerde Universal Co-Anchor Strand toegevoegd aan de celsuspensie, waar het hybridiseert met de Universal Anchor Strand en in het celmembraan wordt ingebracht. De toevoeging van het tweede cholesterolmolecuul verhoogt de netto hydrofobiciteit van het DNA-complex en stabiliseert het in hetmembraan 26. Na het uitwassen van het overtollige DNA worden de cellen geconcentreerd en toegevoegd aan een PDMS-stroomcel bovenop het patroonoppervlak. Het 3'-uiteinde van de adapterstreng hybridiseert met de Oppervlakte-DNA-streng op de glasplaat, wat resulteert in hechting aan de dia, specifiek in regio's die zijn gefunctionaliseerd met complementair DNA. Klik hier om een grotere versie van deze figuur te bekijken.
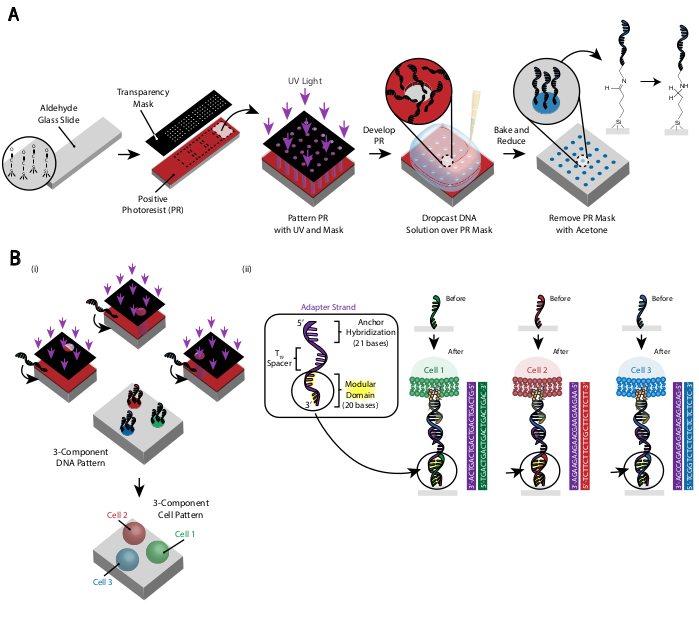
Figuur 3: Fotolithografie wordt gebruikt om de DNA-patroondia's te maken die uiteindelijk de plaatsing van cellen zullen dicteren. (A) Overzicht van het fotolithografieproces. Een aldehyde-gefunctionaliseerde dia is gespind met een positieve fotoresist. UV-licht schijnt op de dia door een transparant fotomasker dat transparant is waar celadhesie gewenst is. Nadat de dia is ontwikkeld, hebben de regio's die eerder aan UV-licht werden blootgesteld nu aldehydegroepen blootgesteld. Een 20 μM-oplossing van een amine-gefunctionaliseerd DNA-oligo wordt vervolgens op de dia gedropt en verspreid over de patroongebieden. De dia wordt vervolgens gebakken om de vorming van Schiff-bindingen (C = N) tussen de amine- en aldehydegroepen te induceren, een omkeerbare covalente binding29. Daaropvolgende reductieve aminatie met 0,25% natriumboorhydride in PBS zet de Schiff-base om in een secundair amine door reductieve aminatie, wat resulteert in een onomkeerbare binding tussen het DNA en de dia. De resterende fotoresist kan vervolgens worden verwijderd door te spoelen met aceton. (B) Dit proces kan worden herhaald om meercomponenten DNA-patronen te creëren en daarom experimenten uit te voeren met meerdere celpopulaties. (i) Nadat het eerste oligo is gemodelleerd, wordt de dia opnieuw bedekt met fotoresist en gaat het protocol verder zoals voorheen. Uitlijning van de fotomaskers met behulp van fiduciaire markers is noodzakelijk voor het patroon van meerdere DNA-strengen. (ii) Elk celtype met een patroon verschilt in het modulaire domein met 20 basissen van de adapterstreng. Door orthogonale sets van complementaire oligo's te gebruiken, kunnen meerdere celtypen worden gemodelleerd zonder kruisadhesie. Klik hier om een grotere versie van deze figuur te bekijken.
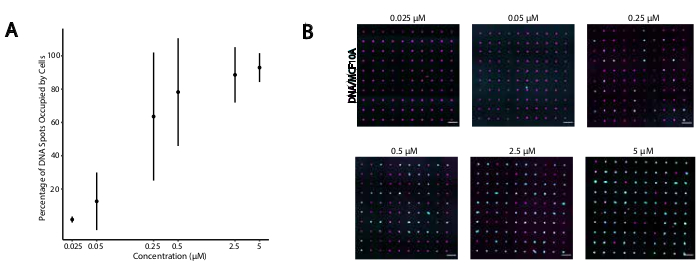
Figuur 4: De hechting van CMO-gelabelde cellen aan DNA-patronen neemt toe als functie van de CMO-concentratie tijdens het labelen. In dit experiment werden de Universal Anchor + Adapter Strand (pre-hybridized) en de Universal Co-Anchor in gelijke concentraties gebruikt. Concentratie verwijst naar de concentratie van CMO in de celsuspensie tijdens CMO-labeling van cellen. A)Kwantificering van het percentage DNA-vlekken met een diameter van 15 μm dat werd ingenomen door CMO-gelabelde MCF10A-cellen als functie van de CMO-concentratie tijdens celetikettering. Gegevens weergegeven als de gemiddelde ± standaarddeviatie van drie experimenten. (B) Representatieve beelden van de DNA-patronen (magenta) en aan gehechte MCF10A's (cyaan) bij verschillende concentraties CMO. Schaalbalk = 100 μm. Klik hier om een grotere versie van deze figuur te bekijken.
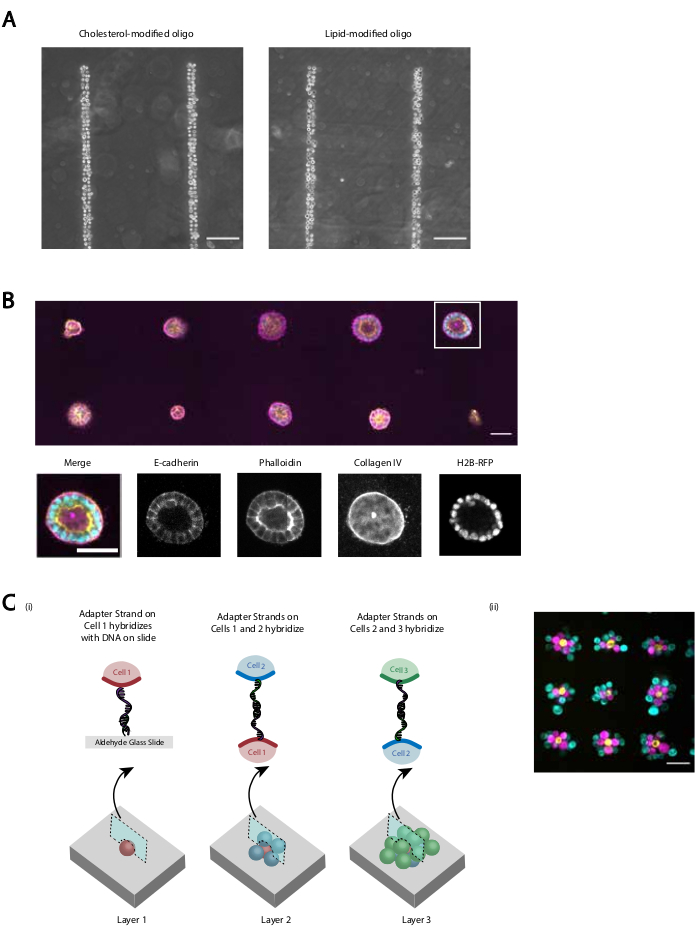
Figuur 5: CMO-DPAC kan worden gebruikt om tweedimensionale celpatronen te creëren die vervolgens kunnen worden ingebed in een driedimensionale hydrogel voor cultuur en / of gelaagd om meerlagige structuren te creëren. (A) Directe vergelijking tussen CMO-gelabelde menselijke navelstreng endotheliale cellen (HUVECs) en LMO-gelabelde HUVECs hielden vast aan een lineair DNA-patroon. Beide methoden van celetikettering resulteren in bijna 100% bezetting van het DNA-patroon. (B)Single Madin-Darby Canine Kidney cells (MDCKs) die H2B-RFP tot expressie brengen, werden gemodelleerd naar plekken met een diameter van 15 μm op een afstand van 200 μm en vervolgens ingebed in Matrigel. Na 120 uur cultuur werden de resulterende epitheliale cysten gefixeerd en gekleurd voor E-cadherine, actine en collageen IV. Sferoïde in witte doos wordt in detail weergegeven. Schaalbalk = 50 μm. (C) Meerlagige cellulaire structuren kunnen worden gemaakt door afzonderlijke celpopulaties te labelen met complementaire adapterstrengen en opeenvolgende patronen, zodat elke nieuwe toevoeging van cellen zich hecht aan de cellaag ervoor. (i) Een schema van de sequentiële patroons van celpopulaties om meerlagige structuren te creëren. ii) Drielaagse celaggregaten van MCF10A's (gevisualiseerd met behulp van kleurstoffen) werden gemaakt met behulp van dit proces. Schaalbalk = 50 μm. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 6: Meerdere celtypen kunnen worden gemodelleerd zonder kruisbesmetting of verlies van hechting. Meerdere amine-gemodificeerde DNA-oligo's werden sequentieel op een aldehyde-dia gemodelleerd en uitgelijnd door gebruik te maken van metalen fiduciaire markers. Drie populaties mcf10a's (cyaan, magenta, geel) werden gekleurd met unieke kleurstoffen gelabeld met complementaire CMO's en op de dia gemodelleerd, wat resulteerde in een afbeelding van de UC Berkeley- en UCSF-logo's. Schaalbalk 1 mm. Klik hier om een grotere versie van deze figuur te bekijken.
Aanvullende figuur 1: Voorbeeldafbeeldingen van de fotolithografie-opstelling op de tafel. (A) Schuif op spin coater, bedekt met positieve fotoresist, vóór spin coating. (B) Afbeelding van transparantie fotomasker. (C)Tijdens de belichting wordt het fotomasker ingeklemd tussen de fotoresistente dia en een glazen schijf. (D) Behuizing voor UV-lamp is gemaakt van een hergebruikte naaldencontainer. (E) Dia ondergedompeld in ontwikkelaar oplossing. (F) Ontwikkelde dia. (G) Amine-gemodificeerde DNA-oplossing verspreid over patroongebieden van de dia. (H) PDMS-stroomcellen die bovenop patroongebieden van de dia worden geplaatst. Klik hier om dit bestand te downloaden.
Aanvullende figuur 2: Enkele voorbeelden van veelvoorkomende fouten van dit protocol. (A) (i) Onderbakken vóór UV-blootstelling of overontwikkeling van kenmerken na blootstelling kan resulteren in kenmerken met gekartelde randen en kunnen onregelmatig van grootte zijn. (ii) Een voorbeeld van een correct gefotopatterde dia met schone randen rond functies, uniforme functiegrootte en geen duidelijke scheuren in het patroon. Schaalbalk = 50 μm. (B) Celdichtheid is van cruciaal belang voor de efficiëntie van patronen. Bij het observeren van de cellen bovenop het patroon onder een microscoop, zouden er weinig openingen tussen cellen moeten bestaan, zoals blijkt uit de voorbeeldafbeelding aan de linkerkant. Schaalbalk = 50 μm. (C) Patrooncellen kunnen gevoelig zijn voor vloeistofkrachten die voortvloeien uit te krachtig pipetteren, wat de patrooncellen kan beschadigen en losmaken. Meerlaagse celaggregaten zijn bijzonder kwetsbaar, omdat één cel aan de onderkant een structuur van meerdere cellen ondersteunt. (i) Een array van celaggregaten die met succes in Matrigel zijn ingebed. (ii) Een raster van celaggregaten dat losraakt als gevolg van het te krachtig pipetteren van viskeuze Matrigel. (D) Klonteren van cellen kan optreden, vooral bij epitheelcellen. Deze klonten zijn meestal homotypisch, maar kunnen heterotypisch zijn (cellen die zich hechten aan reeds patrooncellen van een ander type) als de cellen bijzonder kleverig zijn. Afbeelding toont drie verschillende populaties van MCF10A's werden gemodelleerd naar een array bestaande uit drie verschillende DNA-vlekken van eencellige grootte (15 μm). De meeste DNA-vlekken hebben 2-4 cellen bevestigd. Klonteren kan worden opgelost door EDTA-behandeling of door de klonten eruit te filteren voordat ze worden gemodelleerd. Schaalbalk = 100 μm. Klik hier om dit bestand te downloaden.
Aanvullende figuur 3: Overlappende fotopatrine resulteert in aanwezigheid van beide oligo's bij verminderde concentratie. Twee orthogonale amine-gemodificeerde oligo's werden achtereenvolgens gefotopatterd, eerst een verticale lijn (Streng 1), gevolgd door een horizontale lijn die het overlapte (Streng 2). De oligo's werden vervolgens gevisualiseerd door hybridisatie met fluorescerende complementaire oligo's. (A) Fluorescentiebeeld van streng 1. B)Kwantificering van het fluorescentieprofiel van streng 1 over een verticale lijn van 100 μm die de overlapping overspant. (C) Fluorescentiebeeld van streng 2. D)Kwantificering van het fluorescentieprofiel van streng 2 over een horizontale lijn van 100 μm die de overlapping overspant. Schaalbalk = 50 μm. Klik hier om dit bestand te downloaden.
Aanvullende figuur 4: Kwantificering van DNA-complexen op het celoppervlak als functie van cmo-etiketteringsconcentratie. HUVECs werden gelabeld met verschillende concentraties CMO-oplossing, gewassen en vervolgens geïncubeerd met een fluorescerende complementaire streng. Een MESF (Molecules of Equivalent Soluble Fluorochrome) microsfeerkit werd gebruikt om kwantitatieve flowcytometrie uit te brengen en het aantal DNA-complexen op het celoppervlak te schatten als functie van de CMO-concentratie tijdens het labelen. Klik hier om dit bestand te downloaden.
Aanvullende figuur 5: CMO-etikettering stimuleert de TLR9-respons niet. Er werd een experiment uitgevoerd om te zien of CMO-etikettering het DNA-detectiemechanisme van TLR9 zou activeren en of dit zou worden beïnvloed door CpG's in de Adapter Strand-sequentie. HEK-cellen die muis-TLR9 tot expressie brengen, werden 's nachts geïncubeerd met 0,2 μM van ODN 1826 (een CpG-bevattende TLR9-agonist), CMO Universal Anchor + Universal Co-Anchor + Adapter Strand met dezelfde sequentie als ODN 1826 (CMO-CpG), of CMO Universal Anchor + Universal Co-Anchor + Adapter Strand met een vergelijkbare sequentie, maar met vervanging van de CLB's door GpCs (CMO-GpC). TLR9-stimulatie zou resulteren in de productie van SEAP (uitgescheiden embryonale alkalische fosfatase). SEAP-secretie werd gekwantificeerd door een colorimetrische test (absorptie). Behandelingscondities werden vergeleken met rustende cellen die alleen met PBS werden behandeld. Incubatie met CMO-GPC stimuleerde de TLR9-expressie niet. Incubatie met CMO-CpG was iets hoger dan rustcellen, maar veel lager dan ODN-1826. Klik hier om dit bestand te downloaden.
Aanvullende figuur 6: Levensvatbaarheid van cellen na cmo-etiketteringsproces. Om te beoordelen hoe het protocol de levensvatbaarheid beïnvloedt, werden HUVECs opgesplitst in vier populaties: één bleef 1 uur op ijs, één werd mock-labeled met PBS maar verder door alle centrifuge- en wasstappen genomen, één werd gelabeld met CMO's en één werd gelabeld met CMO's en gefilterd door een 40 μm-filter om klonten te verwijderen. De cellen werden vervolgens gekleurd met calceïne AM en ethidium homodimeer om het aantal levende en dode cellen te beoordelen. Alle behandelingen resulteerden in een significant verminderde levensvatbaarheid dan de ijscontrole (eenrichtings-ANOVA met Tukey post-hoc analyse), maar de mediane levensvatbaarheid voor CMO-labeling (met of zonder filtering) was ongeveer 94%. Gegevens verzameld uit drie onafhankelijke experimenten. * = p < 0,05. = p < 0.0001 Klik hier om dit bestand te downloaden.
| Resultaat | Mogelijke oorzaak(en) | Voorgestelde oplossingen |
| Fotolithografie – functies zijn gekraakt | Inconsistent of ontoereikend zachtbakken | Verhoog de tijd van zachtbakken tot 3 minuten; controleer de werkelijke temperatuur van de kookplaat en verhoog de temperatuur indien nodig |
| Fotolithografie – functies zijn niet scherp of hebben fotoresist in zich | Onderontwikkeling | Verhoog de tijd die dia besteedt aan de ontwikkelaarsoplossing; zachte agitatie opnemen |
| Fotolithografie - functies inconsistent over dia | UV-licht is mogelijk niet gecentreerd of niet goed gericht | Pas de instelling van het UV-licht aan om gecollimeerd licht van uniforme intensiteit te garanderen |
| Cellen hechten zich niet aan patroonvlekken met een hoog rendement | Te weinig DNA aan de oppervlakte | Bevestig dat DNA op het oppervlak aanwezig is door de dia te hybridiseren met fluorescerende complementaire oligo's en vervolgens onder microscoop af te beelden |
| Cellen zijn onvoldoende gelabeld met CMO | Voeg fluorescerende complementaire oligo's toe aan celsuspensie en bevestig fluorescentie via flowcytometrie | |
| Niet genoeg cellen over patroon | Verzamel cellen door uit te spoelen uit PDMS-stroomcel, centrifugeer en suspensie opnieuw in een lager volume om de cellen te concentreren | |
| Te veel resterende CMO in celsuspensie, hybridiseren met DNA op dia | Voeg nog een wasstap toe. Zorg ervoor dat u bij elke wasbeurt zoveel mogelijk bovennatuurlijk verwijdert. | |
| Te veel internalisering van CMO door tijd en temperatuur | Werk snel na het labelen van de cellen met CMO; cellen bewaren en op ijs glijden en ijskoude reagentia gebruiken | |
| Cellen klonteren | Cellen werden niet voldoende gescheiden tijdens trypsinisatie | Gebruik PBS + 0,04% EDTA tijdens het waspen van cellen; laat de celsuspensie door een filter van 35 μm gaan voor de laatste wasbeurt |
| Cellen hechten zich niet-specifiek | Als in een specifiek gebied - kan te wijten zijn aan krassen op de dia, verkeerde uitlijning van PDMS-stromingscellen of morsen van DNA buiten het patroongebied | Vermijd krassen, zorg ervoor dat u de PDMS-stroomcellen uitlijnt met het patroongebied |
| Als cellen zich overal aan vastknechten – onvoldoende blokkeren of wassen | Voeg meer wasbeurten toe na het patroon van de cellen; pipetteer krachtiger tijdens het waspen; blok met 1% BSA langer voordat met celpatronen wordt begonnen; silaniseren (optionele stap 3) of bevestigen dat silanisatie succesvol was door de contacthoek van waterdruppel te meten | |
| Bubbels vormen zich in de stroomcel | Pipetteerfouten, ongelijk hydrofiel oppervlak gecreëerd tijdens plasma-oxidatie | Als de bubbels klein zijn, voeg dan PBS toe aan de inlaat van de stroomcel en ze kunnen worden weggespoeld. Als de bellen groter zijn, oefent u zachte druk uit op de PDMS-stroomcel en duwt u de bellen naar de inlaat of uitlaat. |
| Cellen hechten zich in eerste instantie aan het patroon, maar worden verwijderd tijdens het wassen, het patroon van andere celtypen of het toevoegen van de hydrogelvoorloper | De schuifkrachten van te krachtig pipetteren kunnen ervoor zorgen dat de cellen loskomen van het oppervlak | Pipetteer zachter tijdens volgende wasbeurten, rondes van celpatronen of het toevoegen van hydrogelvoorlopers. Omdat de hydrogelvoorlopers stroperig zijn, is de kans groter dat ze ervoor zorgen dat het patroon losraakt, dus wees extra voorzichtig. Meerlaagse structuren hebben de neiging om topzwaar te zijn en zijn gevoeliger voor losraken. |
| Weefsel vervormt tijdens 3D-overdracht | Hydrogel plakt om te schuiven | Bevestig de hydrofobiciteit van de dia met behulp van contacthoekmetingen |
| Gebruik scheermesje om PDMS volledig aan beide randen op te tillen, waardoor PBS onder het weefsel kan zweven | ||
| Dit kan gebeuren met pure collageen hydrogels – overweeg om de eiwitconcentratie of samenstelling van hydrogel aan te passen | ||
| Cellen gaan niet over met de hydrogel en blijven op de dia | Verhoog de Turbo DNAse-concentratie of verhoog de incubatietijd | |
| Hydrogel is niet stevig genoeg | Verhoog de incubatietijd en/of het gelationmechanisme voor de hydrogel in kwestie (bijv. voor collageen, zorg ervoor dat de pH correct is) | |
| Hydrogel scheurt bij het verwijderen van PDMS | Maak PDMS-flowcellen hydrofiel met behulp van plasma-oxidatie voordat ze met het experiment beginnen, zodat ze gemakkelijk loskomen bij het toevoegen van media. Gebruik een tang heel voorzichtig om het PDMS los te maken. |
Tabel 1: Een handleiding voor het oplossen van problemen om mogelijke fouten die kunnen voortvloeien uit dit protocol te identificeren en op te lossen. In het bijzonder kan een slechte hechting van cellen aan het patroon veel hoofdoorzaken hebben en deze gids zou moeten helpen bij het identificeren en oplossen van die problemen.
Aanvullend dossier 1. Klik hier om dit bestand te downloaden.
Aanvullend dossier 2. Klik hier om dit bestand te downloaden.
Aanvullend dossier 3. Klik hier om dit bestand te downloaden.
Aanvullend dossier 4. Klik hier om dit bestand te downloaden.
In dit artikel presenteren we een gedetailleerd protocol voor hoge resolutie patroons van cellen in 2D en 3D voor in vitro celkweekexperimenten. In tegenstelling tot eerder gepubliceerde versies van deze methode, richt het hier gepresenteerde protocol zich op bruikbaarheid: het vereist geen zeer gespecialiseerde apparatuur en alle reagentia kunnen bij leveranciers worden gekocht in plaats van aangepaste synthese te vereisen. In tegenstelling tot andere celmicropatterningmethoden is deze methode snel en celtype agnostisch: het vereist geen specifieke hechting aan extracellulaire matrixeiwitten15. Cellen met een patroon van CMO-DPAC kunnen worden ingebed in een extracellulaire matrix zoals Matrigel of collageen, wat resulteert in 3D-culturen met een veel hogere ruimtelijke resolutie dan momenteel mogelijk is met op extrusieprinten gebaseerde methoden22. CMO-DPAC kan worden gebruikt om honderden tot duizenden microscopische functies per dia te maken, waardoor veel replicaties tegelijkertijd kunnen worden uitgevoerd.
Een van de belangrijkste parameters in het succes van dit protocol is de dichtheid van cellen die worden toegevoegd aan de stroomcellen bovenop de dia met patroon. Idealiter zou de dichtheid minstens 25 miljoen cellen / ml moeten zijn. Wanneer deze dichtheid van cellen in de stroomcellen wordt geladen, resulteert deze in een bijna dicht opeengepakte monolaag van cellen boven het patroon(aanvullende figuur 2B). Deze hoge celdichtheden maximaliseren de kans dat een cel zich direct op een DNA-plek nestelt en zich hecht. Het verminderen van de celdichtheid zal de algehele patroonefficiëntie verminderen. Een andere cruciale stap in dit protocol is het grondig opnieuw opschorten van de cellen in PBS of serumvrije media voordat de CMO-oplossing wordt toegevoegd. De CMO's verdelen zeer snel in celmembranen en het toevoegen van de CMO-oplossing rechtstreeks aan een celkorrel zal resulteren in heterogene etikettering van cellen. Na het toevoegen van de CMO-oplossing aan de celsuspensie, is het belangrijk om grondig te mengen door te pipetteren, zodat de cellen uniform worden gelabeld met de CMO's. Na de incubaties is het noodzakelijk om de overtollige CMO's grondig uit te spoelen door middel van meerdere centrifugeer- en wasstappen. Overtollige vrije CMO aanwezig in de celsuspensie zal binden aan het patroon amine-gemodificeerd DNA op de glasplaat, waardoor hybridisatie en adhesie van de CMO-gemodificeerde cellen in suspensie wordt geblokkeerd. Tijd is ook een belangrijke overweging voor dit protocol. Het is belangrijk om zo snel mogelijk te werken bij het gebruik van CMO's en om de cellen op ijs te houden om de internalisatie van de CMO's te minimaliseren en de levensvatbaarheid van cellen te maximaliseren. Flowcytometrie-experimenten hebben aangetoond dat CMO's niet zo lang op het celoppervlak blijven bestaan als LMO's, met 25% verlies van CMO-complexen gedurende twee uur incubatie op ijs36. Bovendien zal de levensvatbaarheid van cellen afnemen naarmate de verwerkingstijd van de cel toeneemt. De levensvatbaarheid kan worden gemaximaliseerd door snel te werken, cellen op ijs te houden, ijskoude reagentia te gebruiken en serumvrije media te gebruiken om sommige voedingsstoffen te leveren.
Hoewel CMO-DPAC een krachtige manier kan zijn om celbiologie te bestuderen door cellen met hoge precisie te patroonen, heeft het zijn beperkingen. CMO-DPAC-experimenten kunnen een uitdaging zijn, vooral omdat de experimentele complexiteit wordt toegevoegd met meerdere celtypen, lagen of 3D-celcultuur(Supplemental File 1). Experimentele storingen kunnen vaak voorkomen bij het starten van dit protocol, zoals beschreven in tabel 1. Daarom raden we gebruikers aan om kwaliteitscontroles in te stellen (bevestigen dat DNA aanwezig is op de dia, bevestigen dat cellen voldoende zijn gelabeld met DNA (stap 8), bevestigen dat overtollige cellen grondig zijn weggespoeld, enz.) om ervoor te zorgen dat het experiment slaagt en om stappen te identificeren die mogelijk verdere optimalisatie vereisen. We hopen dat de informatie in dit manuscript en de aanvullende bestanden zal helpen bij het oplossen van de vereiste problemen.
Cholesterol is een bioactief molecuul waarvan de internalisatie het celmetabolisme, genexpressie en membraanfluïditeit kan beïnvloeden37,38. Een eerdere studie vergeleek de effecten op genexpressie van CMO- en LMO-gelabelde cellen met behulp van single cell RNA sequencing. CMO-gelabelde HEK-cellen hadden de genexpressie veranderd in vergelijking met niet-gelabelde en LMO-gelabelde cellen36. Het labelen van cellen met CMO's resulteerde in de differentiële expressie (> 1,5-voudig) van acht genen ten opzichte van niet-gelabelde controles, waaronder AP2B1, dat is gekoppeld aan cholesterol en sfingolipidetransport (GeneCards), en MALAT1, een lang niet-coderend RNA dat cholesterolaccumulatie reguleert39. Hoewel klein, kunnen deze transcriptionele reacties niettemin van belang zijn als het experiment in kwestie het metabolisme, de membraandynamica of andere cholesterolgerelateerde routes in cellen bestudeert.
Dit protocol is flexibel en kan worden aangepast aan de behoeften van elk experiment. Omdat de CMO zichzelf in het lipidemembraan inbrengt in plaats van een specifieke receptor te gebruiken, is de methode celtype-agnostisch (HUVECs, MCF10As, HEKs en MDCKs zijn hier aangetoond). Hoewel cholesterol een ander hydrofoob anker is dan onze eerder gepubliceerde LMO's, hebben we tot nu toe ontdekt dat ze zich op dezelfde manier gedragen. We zouden dus verwachten dat de CMO's werken met een van de grote verscheidenheid aan celtypen die we eerder met LMO's hebben gepubliceerd, inclusief maar niet beperkt tot neurale stamcellen, fibroblasten, perifere mononucleaire bloedcellen, tumorcellen en primaire borstepitheelcellen6,23,27,29,36 . CMO-etikettering stimuleert TLR9 niet, wat suggereert dat het protocol compatibel is met immuuncellen. Membraanintegratie van de CMO is een functie van de totale celgrootte en de mate van negatieve lading in de cel glycocalyx35. Daarom hebben we een protocol (stap 8) opgenomen voor het testen van de mate van membraanintegratie die vatbaar is voor snelle optimalisatie. De specifieke kenmerken van elk celpatroon zullen onvermijdelijk variëren op basis van het experimentele ontwerp (zie Aanvullend bestand 1 voor meer richtlijnen). Hoewel het hierboven beschreven fotopatterningprotocol voor het patroon van het DNA wordt aanbevolen, zou elke methode voor het ruimtelijk beperken van druppels amine-DNA-oplossing moeten werken, zoals het gebruik van druppelprinters met hoge resolutie. De patroonresolutie en de minimale functieafstand variëren afhankelijk van de gebruikte methode. Het is ook theoretisch mogelijk om de DNA-fotopatterende secties van dit protocol te combineren met andere methoden die zijn gebruikt om cellen met DNA te labelen, zoals met DNA gehybridiseerd tot membraan-uitgedrukt zinkvingers40, met behulp van NHS-geconjugeerd DNA41, en reagerende azido siaalzuurresiduen op het celoppervlak met fosfine-geconjugeerd DNA42 . CMO-DPAC kan worden toegepast op een verscheidenheid aan experimenten die een strikte controle over cel-celafstand vereisen, waaronder studies van de interacties tussen paren cellen, co-kweekexperimenten die kijken naar de overdracht van signalen van "zender" -cellen naar "ontvanger" -cellen, en onderzoeken naar het effect van nabijgelegen extracellulaire signalen op stamceldifferentiatie6,29 . De methode kan ook worden gebruikt om microtissues te maken die kunnen worden gebruikt om celmigratie in drie dimensies te bestuderen, de zelforganisatie van cellen in weefsels23,27en het dynamische samenspel tussen cellen en de ECM27. We hopen dat dit protocol onderzoekers een toegankelijk platform biedt om nieuwe toepassingen van op DNA gebaseerde celpatronen met hoge resolutie in hun eigen laboratoria te verkennen.
Z.J.G. is adviseur en aandeelhouder in Provenance Biosciences.
De auteurs willen Jeremy Garcia bedanken voor het testen van dit protocol en Bhushan Kharbikar voor het geven van training over de apparatuur in de UCSF Biomedical Micro and Nanotechnology Core. Dit onderzoek werd gedeeltelijk ondersteund door subsidies van het Department of Defense Breast Cancer Research Program (W81XWH-10-1-1023 en W81XWH-13-1-0221), NIH (U01CA199315, DP2 HD080351-01, 1R01CA190843-01, 1R21EB019181-01A en 1R21CA182375-01A1), de NSF (MCB1330864) en het UCSF Center for Cellular Construction (DBI-1548297), een NSF Science and Technology Center. O.J.S werd gefinancierd door een NSF Graduate Research Fellowship, een Siebel-beurs en een P.E.O. Scholarship. Z.J.G en A.R.A. zijn Chan-Zuckerberg BioHub Investigators.
| Name | Company | Catalog Number | Comments |
| 2-well Chambered Coverglass w/ non-removable wells | Thermo Fisher Scientific | 155379 | |
| Acetic Acid | Sigma-Aldrich | A6283 | |
| Adapter with External SM1 Threads and Internal SM3 Thread | ThorLabs | SM3A1 | |
| Aldehyde Functionalized Slides | Schott | Nexterion Slide AL | Store under dry conditions after opening. |
| All Plastic Syringes, 1 mL | Fisher Scientific | 14-817-25 | |
| Amine-Modified DNA Oligo | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| Aspheric Condenser Lens | ThorLabs | ACL7560 | |
| Borosilicate Disc, 6in Diameter X 1/2in Thick | Chemglass | CG-1906-23 | |
| Cell Culture Dishes 60x15 mm style | Corning | 353002 | |
| Cholesterol-Modified Oligo | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| Diamond Scribe | Excelta | 475B | |
| DNA Oligonucleotide | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| DPBS, no calcium, no magnesium | Thermo Fisher Scientific | 14190250 | |
| Isopropyl Alcohol | Sigma-Aldrich | 278475 | |
| Matrigel Matrix, Growth Factor Reduced | Corning | 354230 | |
| Methylene Chloride (Stabilized/Certified ACS) | Fisher Scientific | D37-4 | |
| MF-321 Developer | Kayaku Advanced Materials | n/a | |
| Microposit S1813 Positive Photoresist | Kayaku Advanced Materials | n/a | |
| Ø3" Adjustable Lens Tube, 0.81" Travel | ThorLabs | SM3V10 | |
| Oven | Thermo Scientific | 51-028-112H | |
| PE-50 Compact Benchtop Plasma Cleaning System | Plasma Etch | PE-50 | |
| Photomask (custom) | CAD/Art Services | n/a | Minimum feature size guaranteed by CAD/Art Services is 10 microns. |
| Razor Blades | Fisher Scientific | 12-640 | |
| RCT Basic Hot Plate | IKA | 3810001 | |
| Silicon Wafer (100 mm) | University Wafer | 590 | |
| Sodium Borohydride, 98%, granules | Acros Organics | 419471000 | |
| Spin Coater Kit | Instras | SCK-200 | This is a low cost option, but any spin coater that can maintain a speed of 3000 rpm will suffice. |
| SU-8 2075 | Microchem | Y111074 0500L1GL | |
| SU-8 Developer | Microchem | Y020100 4000L1PE | |
| Sylgard 184 Silicone Elastomer Kit | Dow | 2646340 | |
| Syringe Needles | Sigma-Aldrich | Z192341 | |
| T-Cube LED Driver, 1200 mA Max Drive Current | ThorLabs | LEDD1B | |
| Tridecafluoro-1,1,2,2-tetrahydrooctyl dimethylchlorosilane | Gelest | SIT8170.0 | |
| Triethylamine | Sigma-Aldrich | 90335 | |
| Turbo DNase | Thermo Fisher Scientific | AM2238 | |
| Tweezers Style N7 | VWR | 100488-324 | The curved shape of these tweezers is essential for delicately picking up the PDMS flow cells containing patterned tissues. |
| UV LED (365 nm, 190 mW (Min) Mounted LED, 700 mA) | ThorLabs | M365L2 | |
| Wafer Tweezers | Agar Scientific | T5063 | |
| WHEATON Dry-Seal vacuum desiccator | Millipore Sigma | W365885 |
- Kreeger, P. K., Strong, L. E., Masters, K. S. Engineering approaches to study cellular decision-making. Annual Review of Biomedical Engineering. , 49-72 (2018).
- Goubko, C. a., Cao, X. Patterning multiple cell types in co-cultures: A review. Materials Science and Engineering C. 29 (6), 1855 (2009).
- Sun, W., et al. The bioprinting roadmap. Biofabrication. 12 (2), 022002 (2020).
- Liu, W. F., Chen, C. S. Cellular and multicellular form and function. Advanced Drug Delivery Reviews. 59 (13), 1319-1328 (2007).
- Duffy, R. M., Sun, Y., Feinberg, A. W. Understanding the role of ECM protein composition and geometric micropatterning for engineering human skeletal muscle. Annals of Biomedical Engineering. 44 (6), 2076-2089 (2016).
- Chen, S., et al. Interrogating cellular fate decisions with high-throughput arrays of multiplexed cellular communities. Nature Communications. 7, 10309 (2016).
- Shaya, O., et al. Cell-cell contact area affects notch signaling and notch-dependent patterning. Developmental Cell. 40 (5), 505-511 (2017).
- Rao, N., et al. A co-culture device with a tunable stiffness to understand combinatorial cell-cell and cell-matrix interactions. Integrative Biology. 5 (11), 1344 (2013).
- Sriraghavan, V., Desai, R. A., Kwon, Y., Mrksich, M., Chen, C. S. Micropatterned dynamically adhesive substrates for cell migration. Langmuir. 26 (22), 17733-17738 (2010).
- Wong, L., Pegan, J. D., Gabela-Zuniga, B., Khine, M., McCloskey, K. E. Leaf-inspired microcontact printing vascular patterns. Biofabrication. 9 (2), 021001 (2017).
- Chen, T. H., et al. Directing tissue morphogenesis via self-assembly of vascular mesenchymal cells. Biomaterials. 33 (35), 9019-9026 (2012).
- Laurent, J., et al. Convergence of microengineering and cellular self-organization towards functional tissue manufacturing. Nature Biomedical Engineering. 1 (12), 939-956 (2017).
- Lin, C., Khetani, S. R. Micropatterned co-cultures of human hepatocytes and stromal cells for the assessment of drug clearance and drug-drug interactions. Current Protocols in Toxicology. 2017, 1-23 (2017).
- Hui, E. E., Bhatia, S. N. Micromechanical control of cell-cell interactions. Proceedings of the National Academy of Sciences of the United States of America. 104 (14), 5722-5726 (2007).
- D'Arcangelo, E., McGuigan, A. P. Micropatterning strategies to engineer controlled cell and tissue architecture in vitro. BioTechniques. 58 (1), 13-23 (2015).
- Martinez-Rivas, A., González-Quijano, G. K., Proa-Coronado, S., Séverac, C., Dague, E. Methods of micropatterning and manipulation of cells for biomedical applications. Micromachines. 8 (12), (2017).
- Lee, S., et al. Simple lithography-free single cell micropatterning using laser-cut stencils. Journal of Visualized Experiments. (158), e60888 (2020).
- Strale, P. O., et al. Multiprotein printing by light-induced molecular adsorption. Advanced Materials. 28 (10), 2024-2029 (2016).
- Melero, C., et al. Light-induced molecular adsorption of proteins using the primo system for micro-patterning to study cell responses to extracellular matrix proteins. Journal of Visualized Experiments. (152), e60092 (2019).
- Reid, J. A., Mollica, P. M., Bruno, R. D., Sachs, P. C. Consistent and reproducible cultures of large-scale 3D mammary epithelial structures using an accessible bioprinting platform. Breast Cancer Research. , 1-13 (2018).
- Wang, Z., Lee, S. J., Cheng, H. -. J., Yoo, J. J., Atala, A. 3D bioprinted functional and contractile cardiac tissue constructs. Acta Biomaterialia. 70, 48-56 (2018).
- Miri, A. K., et al. Effective bioprinting resolution in tissue model fabrication. Lab on a Chip. 19 (11), 2019-2037 (2019).
- Todhunter, M. E., et al. Programmed synthesis of three-dimensional tissues. Nature Methods. 12 (10), 975-981 (2015).
- Todhunter, M. E., Weber, R. J., Farlow, J., Jee, N. Y., Gartner, Z. J. Fabrication of 3D microtissue arrays by DNA programmed assembly of cells. Current Protocols in Chemical Biology. 8 (3), 147-178 (2016).
- Csizmar, C. M., Petersburg, J. R., Wagner, C. R. Programming cell-cell interactions through non-genetic membrane engineering. Cell Chemical Biology. 25 (8), 931-940 (2018).
- Weber, R. J., Liang, S. I., Selden, N. S., Desai, T. A., Gartner, Z. J. Efficient targeting of fatty-acid modified oligonucleotides to live cell membranes through stepwise assembly. Biomacromolecules. 15 (12), 4621-4626 (2014).
- Hughes, A. J., et al. Engineered tissue folding by mechanical compaction of the mesenchyme. Developmental Cell. 44 (2), 165-178 (2018).
- Weber, R. J., et al. Rapid organoid reconstitution by chemical micromolding. ACS Biomaterials Science & Engineering. 2 (11), 1851-1855 (2016).
- Scheideler, O. J., et al. Recapitulating complex biological signaling environments using a multiplexed, DNA-patterning approach. Science Advances. 6 (12), (2020).
- Viola, J. M., et al. Guiding cell network assembly using shape-morphing hydrogels. Advanced materials (Deerfield Beach, Fla.). , 2002195 (2020).
- Mohammad, A., Davis, M., Aprelev, A., Ferrone, F. A. Note: Professional grade microfluidics fabricated simply. Review of Scientific Instruments. 87 (10), 1-4 (2016).
- Lee, O. J., Chuah, H. S., Umar, R., Chen, S. K., Yusra, A. F. I. Construction of cost effective homebuilt spin coater for coating amylose-amylopectin thin films. Journal of Fundamental and Applied Sciences. 9 (2), 279 (2018).
- Webb, K., Hlady, V., Tresco, P. A. Relative importance of surface wettability and charged functional groups on NIH 3T3 fibroblast attachment, spreading, and cytoskeletal organization. Journal of Biomedical Materials Research. 41 (3), 422-430 (1998).
- Processing Guidelines for: SU-8 2025, SU-8 2035, SU-8 2050, SU-8 2075. Microchem SU-8 2000 Permanent Expoxy Negative Photoresist Available from: https://kayakuam.com/wp-content/uploads/2019/09/SU-82000DataSheet2025thru2075Ver4.pdf (2019)
- Palte, M. J., Raines, R. T. Interaction of nucleic acids with the glycocalyx. Journal of the American Chemical Society. 134 (14), 6218-6223 (2012).
- McGinnis, C. S., et al. MULTI-seq: sample multiplexing for single-cell RNA sequencing using lipid-tagged indices. Nature Methods. 16 (7), 619-626 (2019).
- Maxfield, F. R., van Meer, G. Cholesterol, the central lipid of mammalian cells. Current Opinion in Cell Biology. 22 (4), 422-429 (2010).
- Luo, J., Yang, H., Song, B. L. Mechanisms and regulation of cholesterol homeostasis. Nature Reviews Molecular Cell Biology. 21 (4), 225-245 (2020).
- Liu, L., Tan, L., Yao, J., Yang, L. Long non-coding RNA MALAT1 regulates cholesterol accumulation in ox-LDL-induced macrophages via the microRNA-17-5p/ABCA1 axis. Molecular Medicine Reports. 21 (4), 1761-1770 (2020).
- Mali, P., Aach, J., Lee, J. H., Levner, D., Nip, L., Church, G. M. Barcoding cells using cell-surface programmable DNA-binding domains. Nature Methods. 10 (5), 403-406 (2013).
- Hsiao, S. C., et al. Direct cell surface modification with DNA for the capture of primary cells and the investigation of myotube formation on defined patterns. Langmuir. 25 (12), 6985-6991 (2009).
- Gartner, Z. J., Bertozzi, C. R. Programmed assembly of 3-dimensional microtissues with defined cellular conductivity. Proceedings of the National Academy of Sciences. (17), 1-5 (2009).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved