
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Bioengineering
דפוס פשוט, זול ומודולרי של תאים המשתמשים ב- DNA
כאן אנו מציגים פרוטוקול לתאים זעירים ברזולוציה של תא בודד באמצעות הידבקות מתוכנתת DNA. פרוטוקול זה משתמש בפלטפורמת פוטוליטוגרפיה על הספסל כדי ליצור דפוסים של אוליגונוקלאוטידים DNA על שקופית זכוכית ולאחר מכן תוויות קרום התא עם אוליגונוקלאוטידים משלימים זמינים מסחרית. הכלאה של אוליגוס גורמת הידבקות תאים מתוכנתים.
המיקום היחסי של תאים הוא תכונה מרכזית של microenvironment המארגן אינטראקציות תא-תאים. כדי לחקור את האינטראקציות בין תאים מאותו סוג או סוג אחר, טכניקות micropatterning הוכיחו שימושי. הרכבה מתוכנתת של תאים (DPAC) היא טכניקת מיקרו-מפטרציה המכוונת להידבקות של תאים למצע או לתאים אחרים באמצעות הכלאת DNA. הפעולות הבסיסיות ביותר ב- DPAC מתחילות עם קישוט קרום התא עם אוליגונוקלאוטידים שעברו שינוי שומנים בדם, ואז מזרימים אותם על מצע שעוסק ברצפי DNA משלימים. תאים לדבוק באופן סלקטיבי למצע רק שם הם מוצאים רצף DNA משלים. תאים שאינם דבקים נשטפים, חושפים דפוס של תאים דבקים. פעולות נוספות כוללות סבבים נוספים של מצע תא או הידבקות בתא, כמו גם העברת התבניות שנוצרו על ידי DPAC להידרוג'ל מוטבע לתרבות ארוכת טווח. בעבר, שיטות לדפוס אוליגונוקלאוטידים על משטחים וקישוט תאים עם רצפי DNA נדרש ציוד מיוחד וסינתזת DNA מותאמת אישית, בהתאמה. אנו מדווחים על גרסה מעודכנת של הפרוטוקול, תוך שימוש בהגדרת פוטוליטוגרפיה זולה וזמין מסחרית כולסטרול שונה אוליגונוקלאוטידים (CMOs) פרוסים בפורמט מודולרי. תאים המסומנים בתווית CMO דבקים ביעילות גבוהה במצעים בדוגמת DNA. גישה זו יכולה לשמש ליצירת סוגי תאים מרובים בו-זמנית בדיוק גבוה וליצור מערכים של מיקרוטיסות המוטמעות בתוך מטריצה חוץ-תאית. היתרונות של שיטה זו כוללים את הרזולוציה הגבוהה שלה, היכולת להטמיע תאים במיקרו-סביבה תלת-ממדית מבלי לשבש את המיקרו-פטרן, וגמישות בדוגמת כל סוג תא.
המיקום של תאים ביחס זה לזה ברקמה הוא תכונה חשובה של microenvironment1,2,3,4. טכניקות המשמשות לתבנית תאים חיים לתוך סידורים מבוקרים מרחבית הם כלים ניסיוניים יקריערךלחקר בידול 4,5,6,7,8, תנועתיות תאים9, מורפוגנזה10,11,12, חילוף חומרים13, ואינטראקציות תא תא7,14 . קיימות מגוון שיטות לתבנית תאים, שלכל אחד מהם יתרונות וחסרונותמשלו 3,4. שיטות היוצרות איים דבקים של חלבוני מטריצה חוץ-תאית (ECM), כגון הדפסת מיקרו-קונטקט ושבלונות חתוכות בלייזר, הן פשוטות ומדרגיות. עם זאת, קשה דפוס יותר מסוג תא אחד או שניים בכל פעם כי המאפיינים דבק של סוגי תאים שונים למולקולות ECM שונות דומים לעתים קרובות15,16,17. מיקרופטרנים מורכבים יותר ניתן ליצור עם סופת מולקולרית הנגרמת על ידי אור (LIMAP), טכניקה המשתמשת באור UV כדי לפטור אזורים מצופים PEG ולאפשר ספיחת חלבון לאחר מכן18,19. ניתן לחזור על תהליך זה כדי ליצור מיקרו-פלטרנס ברזולוציה גבוהה עם סוגי תאים מרובים. עם זאת, קשירה צולבת של תאים טלאי חלבון שונים יכולה להתרחש, וכתוצאה מכך ספציפיות דפוס ירודה19. שיטות פיזיות כגון זריעת תאים על התקני תרבית מיקרומכניים הניתנים להגדרה מחדש יכולות ליצור תרביות משותפות מובנות עם שליטה דינמית, אך ללא הגמישות בעיצוב דפוס של הדפסת מיקרו-מגע או LIMAP14,8. שלא כמו טכניקות אחרות, ביו הדפסה יכולה ליצור סידורים תלת ממדיים של תאים בתוך הידרוג'ל20,21. עם זאת, מבנים מודפסים ביו יש רזולוציה נמוכה בהרבה מאשר טכניקות micropatterning אחרות, עם גודל תכונה ממוצע בסדר גודל של מאות מיקרון22. שיטת דפוס תאים אידיאלית תהיה ברזולוציה גבוהה, דפוס סוגי תאים מרובים, להשתמש בציוד ריאגנטים נגישים בקלות, ויש להם את היכולת להטמיע דפוסים מוצלחים לתוך הידרוג'ל עבור תרבית תאים תלת ממדית (3D). במאמר זה, אנו מציגים CMO-DPAC, טכניקת מיקרו-מפטרציה של תאים המשתמשת בגמישות ובמהירות של הכלאת DNA כדי למקד הידבקות תאים למצע. שיטה זו הותאמה מהפרוטוקולים הקודמים שלנו23,24 כדי להפוך אותה לזוהה יותר, מודולרית ונגישה יותר. באמצעות הפרוטוקול הנוכחי, כל מעבדה צריכה להיות מסוגלת להקים מערכת פונקציונלית לחלוטין ללא כל ציוד מיוחד או מומחיות.
הרכבה מתוכנתת DNA של תאים (DPAC) היא טכניקה הנדסית רקמות רבת עוצמה המתבניתת תאים ברזולוציה של תא בודד עם שליטה מדויקת על מרווח בין תאים וגיאומטריית רקמות. ב- DPAC, קרום התא מעוטר באוליגונוקלאוטידים של DNA (אוליגוס) באמצעות שני אוליגוס שעברו שינוי שומנים שנועדו להתהכלא על קרום התא. מכיוון שהאוליגוס מצומדים לליפידים הידרופוביים, הם מתחלקים במהירות לקרום התא25 שם הם מתכלאים, מגדילים את ההידרופוביה נטו של המולקולות שאינן קשורות באופן קוולנטי, ובכך משפרים את חייהם על פני התא26. האוליגו מוצגים על פני התא באופן שבו הם יכולים להתהכלא עם אוליגוס משלימים על תאים אחרים או שקופיות זכוכית פונקציונליות DNA כדי ליצור דפוסי תאים 2D או 3D מוגדרים עם הרכב שנקבע, מרווח בין תאים וגיאומטריה23,24. ניתן לבקע את המיקרוטיסות בדוגמתן מפני השטח באופן אנזימטי ולהטמיע בהידרוג'ל לתרבות תלת-ממדית ממושכת. כאשר נעשה שימוש בשילוב עם תאים ראשיים או תאי גזע, האוספים המתקבלים של תאים יכולים לעבור מורפוגנזה וליצור לתוך organoids23,27,28. DPAC הוחל לחקור את הדינמיקה של גורל תאי גזע עצביים למבוגרים בתגובה לאותות מתחרים6,29, לחקור ארגון עצמי של תאי אפיתל ממארי23,28, וליצור "אוריגמי רקמה" באמצעות עיבוי mesenchymal27.
DPAC מאפשר מיקום מדויק של אוכלוסיות תאים מרובות ויש לו רזולוציה טובה יותר באופן משמעותי מאשר ביו-הדפסה מבוססת שחול (בסדר של מיקרונים)22,23. בנוסף, שלא כמו שיטות דפוס מבוססות ECM כגון הדפסת מיקרו-חיבור, DPAC אינו דורש הידבקות דיפרנציאלית של סוגי התאים השונים למשטח מצופה ECM15,23. הוא אידיאלי לענות על שאלות על איך הרכב של רקמה משפיע על ההתנהגות שלה, איך תאים לשלב רמזים תאיים מיקרו-וירוונמנטליים מרובים בעת קבלת החלטות6,29, וכיצד זוגות של תאים אינטראקציה אחד עם השני. יתרון של שיטה זו על פני שיטות micropatterning אחרות הוא כי זה יכול לשמש עבור תרבית תאים 3D במישור הדמיה אחד, להקל על מחקרים לשגות זמן של ארגון עצמי רקמות מורפוגנזה organoid23,27,30.
למרות יתרונות אלה, יישום מוצלח של DPAC דרש סינתזה של ריאגנטים אוליגונוקלאוטיד מותאמים אישית וגישה לציוד מיוחד לדפוס DNA23,24, הגבלת אימוץ נרחב. לדוגמה, אוליגוס אופטימלית (LMOs) שינוי שומנים המשמש בפרוטוקול המקורי חייב להיות מסונתז בהתאמה אישית, שונה עם חומצה לינוקרית או חומצה פלמיטית, ומטוהר26. תהליך זה דורש שימוש בסינתיסייזר DNA ומכשיר כרומטוגרפיה נוזלי בעל ביצועים גבוהים, כמו גם רכישת ריאגנטים הקשורים כגון מתילאמין, חומר מבוקר הכפוף לתקנות מוסדיות ופדראליות כאחד. כחלופה, ניתן לרכוש LMOs בהתאמה אישית בכמויות גדולות, אך הדבר דורש השקעה משמעותית מראש בטכנולוגיה.
כדי להתגבר על מגבלות אלה, פיתחנו גרסה מתוקנת של DPAC המשתמשת באוליגו (CMOs) הזמינים מסחרית במקום LMOs מסונתזים בהתאמה אישית. כדי להפחית עוד יותר את העלויות ולהגדיל את הגמישות של הפלטפורמה, עברנו למערכת מודולרית של שלושה אוליגו. במקום להזמין אוליגו חדש שעבר שינוי כולסטרול עבור כל אוכלוסיית תאים ייחודית, משתמש בפרוטוקול זה יכול במקום זאת להשתמש באותם אוליגוס שעברו שינוי כולסטרול ("עוגן אוניברסלי" ו"עוגן קו-עוגן אוניברסלי") עבור כל אוכלוסיית תאים ולאחר מכן להעסיק אוליגו זול ולא משתנה ("סטרנד מתאם") המתכלא הן עם העוגן האוניברסלי והן עם ה- DNA המתפקד באמין על פני השטח או עם גדיל המתאם מסוג תא אחר.
מגבלה נוספת של פרוטוקול DPAC המקורי הייתה שהוא יצר את השקופיות בדוגמת DNA באמצעות מדפסת נוזלית ברזולוציה גבוהה (למשל, Nano eNabler, BioForce Nanosciences)23,24. בעוד מכשיר זה מתגאה ברזולוציה יוצאת דופן ודרישות ריאגנט נמוכות, הוא אינו זמין לרוב המוסדות ויש לו שיעור הדפסה נמוך יחסית (כ 1 תכונה בדוגמת שנייה). לאחרונה פותחו שתי שיטות פוטו-ליתוגרפיות לתבנית תכונות דנ"א על משטחים. ויולה ועמיתיו השתמשו בציפוי פוליאקרילמיד ובנזופנון שקשר בהתאמה אוליגוס DNA חד-גדילי עם חשיפה לאור UV30. בשיטה זו, הם הצליחו ליצור פיגומי רקמות עברו שינויי צורה בקנה מידה גדול ומתוכנת כתוצאה מהתכווצות התאים והארגון העצמי. Scheideler ואח ' פיתח שיטה המשתמשת בחשיפה UV של פוטוארסיסט חיובי לחשוף באופן סלקטיבי אוליגוס DNA שונה אמין לשקופית29פונקציונלית אלדהיד . לאחר אפייה ומיצוי רדוקטיבי, הדנ"א שעבר שינוי אמין קשור באופן קוולנטי לפני השטח. שיטה זו שימשה כדי לחקור את התגובה של תאי גזע עצביים בוגרים כדי להציג באופן מרחבי רמזי התחדשות עצמית ובידול. מאמר זה מתאים את הפרוטוקול של שיידלר ואח' ליצירת דפוסי ה- DNA שילכדו תאים בעלי תווית CMO. פרוטוקול צילום זה יכול להתבצע ללא שימוש בחדר נקי. הוא משתמש בציוד זול וזמין מסחרית שנפרס בקלות על ספסל או מכסה המנוע האדים. השימוש בציוד פוטו-ליתוגרפיה זול או עשה זאת בעצמך מגביר את הנגישות לחוקרים ללא גישה למתקני חדר נקי ומאפשר לחוקרים לנסות את הטכניקה ללא השקעה גדולה של זמן אומשאבים 31,32. עם זאת, רזולוציה טובה יותר ואת היישור של תכונות DNA מרובות ניתן להשיג באמצעות מעיל ספין מסחרי ו aligner מסכה בדרך כלל למצוא במתקני חדר נקי.
כאן, אנו מתארים שיטה לתבנית תאים ברזולוציה של תא בודד באמצעות הידבקות מבוססת DNA. ראשית, photopatterning עם photoresist חיובי משמש ליצירת דפוסים ברזולוציה גבוהה של DNA שונה אמין על מצע זכוכית שונה אלדהיד. לאחר מכן, השקופית מטופלת כדי להפחית את הקובץ המצורף לתאים שאינם ספציפיים ותאי זרימת PDMS נוצרים כדי להגביל תאים על אזורים בעלי תבנית. תאים מסומנים לאחר מכן עם אוליגונוקלאוטידים DNA קצרים כי הם פונקציונליים עם כולסטרול וכתוצאה מכך להכניס לתוך קרום התא. לאחר מכן, התאים זורמים מעל המיקרו-פטרנים של הדנ"א. הכלאה בין הדנ"א של פני התא לבין הדנ"א על פני הזכוכית גורמת להידבקות ספציפית של התאים לתבנית הדנ"א. תאים שאינם דבקים נשטפים, חושפים את דפוס התא החסידי. ניתן לחזור על תהליך זה כדי ליצור סוגי תאים מרובים או כדי ליצור מבנים מרובי שכבות. אם תרצה, ניתן להטביע את התאים באופן מלא ב- ECM עבור תרבית תאים תלת-ממדית.
1. ניסוי עיצוב
- תכנן את הניסוי הרצוי, בהתחשב בגודל התכונה, בריווח התכונות, מספר סוגי התאים המעורבים וסידור התאים ביחס זה לזה. עיין בקובץ משלים 1, מדריך לעיצוב ניסיוני, וקובץ משלים 2, המכיל רצפי אוליגו לדוגמה.
- עצב מסכת צילום באמצעות תוכנת עיצוב בעזרת מחשב. מסכת צילום לדוגמה מסופקת בקובץ משלים 3.
- צייר מלבן של הממדים של שקופית מיקרוסקופ סטנדרטית (25 מ"מ x 75 מ"מ).
- צייר ארבעה אזורים מלבניים ברוחב 10 מ"מ ובאורך 10 מ"מ, המופצים באופן שווה על-פני השקופית.
- בתוך כל אזור, צייר תכונות שהן הגודל, הצורה והמרווח הרצויים עבור הניסוי. תאים ידבקו רק בתכונות אלה בניסוי.
- כדי ליצור מסכות תמונה מיושרות עבור סוגי תאים מרובים, צור ציור ראשי עם כל ערכות התכונות ולאחר מכן שמור גירסאות התואמות לכל סוג תא.
- הזמינו מסכת צילום ברזולוציה גבוהה (לפחות 20,000 נקודות לאינץ') מציור CAD זה עם התכונות שצוירו ב-1.2.3 שקופות והאזורים הגדולים יותר שחורים.
2. DNA פוטופטרן על שקופיות פונקציונליות אלדהיד (פרוטוקול מותאם מ Scheideler ואח'29 )
- אם אתה יוצר דוגמאות מילוי של סוגי תאים מרובים, פברק סמנים פידמיים בשקופית בעלת התפקוד של אלדהיד לפני כל דפוס DNA כדי להקל על יישור התכונות. שיטות חלופיות ליצירת סמנים פיודואליים מוצעות בקובץ משלים 1.
- כדי ליצור סמנים פיודואליים ממתכת, החל S1813 פוטורסיסט חיובי כמתואר בשלבים 2.3 - 2.11. השתמש במסכה עם תמונה המכילה תכונות גדולות שיהיה קל ליישר מאוחר יותר. שלב תכונות אלה בעיצוב של מסכות הצילום שישמשו לדפוסי DNA.
- הפקד סרט דק (100 אנגסטרום) של טיטניום על השקופית באמצעות אידוי אקדח אלקטרונים29. הסר עודף מתכת ו photoresist באמצעות אצטון, ולאחר מכן להמשיך photopatterning DNA.
- הכן פתרון של 20 מיקרומטר של אוליגו 5'-אמין שונה במאגר DNA (50 mM של נתרן פוספט במים, pH = 8.5). ראה קובץ משלים 2 עבור רצפי אוליגו מוצעים.
הערה: ניתן להשתמש מעט כמו 5 מיקרומטר של אוליגו שונה אמין עבור כמה דפוסים ויישומים, כך ריכוז DNA פני השטח ייתכן שיהיה צורך אופטימיזציה. - מחממים מראש צלחת חמה ל 100 °C (50 °F).
- השתמש בסרט דו-צדדי או בוואקום כדי לחבר שקופית זכוכית פונקציונלית של אלדהיד לרוטור של מעיל ספין.
זהירות: ניתוק שקופיות במהלך ציפוי ספין הוא סיכון בטיחותי. השתמש תמיד במעיל ספין במיכל סגור עם מכסה, כגון קופסת אקריל.
הערה: סמן פינה של השקופית באמצעות סופר יהלומים או כלי דומה כדי לגרד את הזכוכית. פעולה זו מסייעת בזיהוי שקופיות והתמצאות לאחר שצלם הצילום נשטף. - השתמש פיפטה חד פעמית כדי להפיל את photoresist החיובי על שקופית אלדהיד. לציפויים זוגיים, הוסיפו טיפות קטנות של הפוטורסיסט על פני השקופית, במקום טיפה אחת גדולה באמצע(איור משלים 1A).
- באמצעות מעיל הספין, לסובב את השקופית ב 3000 סל"ד עבור 30 s.
- מניחים את השקופית על 100 °C כיריים למשך 1.5 דקות (אפייה רכה) כדי להצליב פוטורסיסט.
- הסר את השקופית מהחלמה החמה. מקם מסכת צילום עם התכונות הרצויות לניסוי זה על גבי השקופית ושקל את מסכת הצילום כלפי מטה עם חתיכת זכוכית(איור 1B,C). מכסים את כל ההתקנה בקופסה אטומה(איור 1D משלים). לחשוף עם מנורת UV (365 ננומטר אורך גל, 360 mW, 5 אינץ 'מן השקופית, צפיפות אנרגיה קורנת הכוללת 100 mJ / ס"מ2) במשך 2 דקות.
הערה: אור UV ישבור את הקשרים הפולימריים בפוטורסיסט מתחת לאזורים השקופים של מסכת הצילום, וייצור אזורים שבהם ה- DNA יוכל מאוחר יותר לדבוק. - פתח את השקופית על ידי טבילה בפתרון מפתחים למשך 3-5 דקות(איור משלים 1E).
- יש לשטוף את תמיסת המפתח העודף במים. יבש תחת זרם של אוויר או חנקן. (איורמשלים 1F).
- אשר כי הפוטוליתוגרפיה הצליחה על ידי התבוננות בשקופית מתחת למיקרוסקופ. מכיוון שהפוטורסיסט רגיש לאור UV, בצע שלב זה במהירות ולאחר מכן אחסן את השקופית בחושך תוך הכנת שקופיות אחרות (אם ישים).
הערה: לשקופית עם תבנית בהצלחה אמורים להיות קצוות מוגדרים בחדות עבור כל תכונה, ללא פיצוח וללא עיוות תכונות בקצוות. דוגמאות לפוטוליתוגרפיה נכונה ולא נכונה ניתנות באיור 2A המשלים. ראה טבלה 1 לקבלת הצעות לפתרון בעיות אם פוטוליתוגרפיה אינה מספקת את איכות התכונה הרצויה. - הוסף טיפה של פתרון אוליגו 20 מיקרומטר שינוי אמין (שלב 2.1) על כל אזור photopattered של השקופית. השתמש קצה pipette כדי להפיץ בעדינות את טיפה על פני האזור כולו, נזהר לא לגרד את השקופית. (איורמשלים 1G).
- אופים את המגלשה בתנור 65-70 מעלות צלזיוס עד שתמיסת הדנ"א מתייבשת לחלוטין על משטח הגלישה (כשעה).
- בצעו הפחתת צמצום על ידי הצבת השקופיות הדוגמנות והאפויות בצלחת תרבית תאים בגודל 15 ס"מ והניחו במכסה אדים על גבי שייקר. לשקול 100 מ"ג של נתרן בורוהידריד. במכסה המנוע האדים, הוסיפו 40 מ"ל של תמיסת מלח (PBS) עם אגירה בפוספט, ערבבו בעדינות והוסיפו לצלחת המכילה את השקופיות המעוצבות. תן לתגובה להמשיך במשך 15 דקות עם רועד עדין.
הערה: האמין על האוליגו יוצר תחילה בסיס שיף עם אלדהידים על משטח השקופית. זהו אג"ח קוולנטי הפיך שיש להמיר לאג"ח בלתי הפיך לפני השימוש ב- DPAC. תוספת של חומר מפחית (נתרן בורוהידריד) ממירה את בסיס שיף לאמין משני על ידי הפחתת.
זהירות: התגובה של נתרן בורוהידריד עם מים יוצרת גז מימן ותמשיך לעשות זאת במשך שעות או ימים לאחר תחילת התגובה. בצע את שלב ההפחתה במכסה המנוע של האדים ושמור על כל פסולת פתרון הבורוהידריד נתרן במיכל פתוח או עטוף באופן רופף במכסה המנוע של האדים למשך 24 שעות לפחות. - הסר DNA לא נטען על ידי שטיפת פעמיים עם 0.1% נתרן דודסיל סולפט (SDS) במים, ולאחר מכן שלוש פעמים עם מים מזוקקים. יבש את המגלשה תחת זרם של חנקן או אוויר.
- לשטוף את השקופית עם אצטון כדי להסיר את photoresist הנותר.
הערה: בשלב זה, ה- DNA היה מחובר באופן בלתי הפיך וקוולנטי לשקופית וכל הקבוצות התפקודיות של אלדהיד שלא הוקמו הוסבו לאלכוהול. הצלם כבר לא נחוץ. - אם אוליגוס מרובים יהיו בדוגמת דפוס, חזור לשלב 2.4, יישר את מסכת התמונות עם סימונים פי דו-יים וחזר על הפעולה.
הערה: ניתן להשהות את הניסוי כאן. אחסן שקופיות במתן ואקום. בתנאים יבשים, ניתן לאחסן את השקופיות עד 3 חודשים ללא אובדן איכות.
3. הפוך שקופית הידרופובית (אופציונלי) (פרוטוקול מותאם מ Todhunter ואח'24 )
הערה: זה יתרון, אבל לא נדרש, כדי לשנות את הכימיה פני השטח של השקופית כדי להפוך אותו יותר אדיש הידרופובי. קובץ מצורף לתא לא ספציפי מופחת על משטחים אלה33, ובכך מקל על איגוד תאים לא ספציפי לאזורים שאינם בעלי תבנית בשקופית. בנוסף, אם התאים בדוגמת בסופו של דבר יוטבעו בתוך הידרוג'ל ויועברו מהשקופית, הטיפול על פני השטח חיוני לתנועה אמינה של ההידרוג'ל עמוס התאים על פני השקופית ללא עיוות או קרע. סילאניזציה עם (tridecafluoro-1,1,2,2-טטרהידרוקטיל) דימתילכלורוסילאן גורמת לנוכחות של קבוצות פלואורואלקיל הידרופוביות על פני השטח של השקופית.
אזהרה: בצע את כל השלבים מ-3.1 ואילך במכסה המנוע של אדים כימיים כדי למנוע חשיפה לחומצה אצטית ולאדי מתילן כלוריד.
- יש לשטוף את הגלישה עם 10% חומצה אצטית ולאחר מכן להתייבש מתחת לזרם אוויר.
- בצנצנת קופלין מזכוכית, הכינו פתרון של 60 מ"ל מתילן כלוריד (דיכלורומתאן), 0.6 מ"ל של טריאתילאמין ו-0.6 מ"ל של דימתיל-כלורוסילאן (טריידקלאורו-1,1,2,2-טטרהידרוקטיל). מערבבים עם מרית מתכת לערבב.
הערה: ריאגנטים אלה רגישים למים. הם צריכים להיות מאוחסנים בתנאים יבשים ולהשתמש טרי ככל האפשר. - מוסיפים את השקופית לצנצנת קופלין המכילה את תמיסת הסילאן. מניחים את צנצנת קופלין על שייקר מסלולי (מוגדר ל-60-80 סל"ד) ומאפשרים לתגובת הסילאן והמגלשה להתקדם במשך 15 דקות.
- השתמש במלקחיים מתכת כדי להסיר את השקופית מפתרון סילאן. יש לטבול שקופית בצנצנת קופלין המכילה מתילן כלוריד למשך דקה אחת כדי להסיר עודף סילאן מהשקופית.
- לטבול את השקופית בצינור חרוט 50 מ"ל המכיל אתנול. להתסיס. לטבול את השקופית בצינור חרוט 50 מ"ל המכיל מים מזוקקים. להתסיס.
הערה: מתילן כלוריד ומים אינם מטעים, ולכן יש צורך בשטיפת אתנול כדי להסיר עודף מתילן כלוריד לפני שטיפה סופית במים. - הסר את המגלשה מהמים ולבדוק אותו. המגלשה צריכה להיות יבשה למדי, עם כל טיפות מים שיש זווית מגע של יותר מ 90°. אפשר לשקופיות להתייבש באופן מלא ולאחסן במתייבש ואקום עד לשימוש.
הערה: ניתן להשהות את הניסוי כאן. אחסן את השקופית בתנאים יבשים.
4. הכן תאי זרימת PDMS ושקופית לניסוי
הערה: תאי זרימת PDMS מלבניים משמשים לרכז את התאים מעל האזורים בעלי הדוגמת של השקופית. לניסויים בתרבית בתלת-ממד, תאי הזרימה יוצרים תבנית עבור ההידרוג'ל.
- הפוך SU-8 מאסטר לשימוש כעובש עבור תאי זרימת PDMS.
- לפני החום חם ל 95 °C (5 °F).
- הוסף 5 מ"ל של SU-8 2075 לופל סיליקון.
- ספין מעיל SU-8 על הוופל ב 500 סל"ד עבור 10s, ואחריו 1,000 סל"ד עבור 30s. פעולה זו אמורה ליצור תכונות בגובה של עד 240 מיקרומטר34.
- רכים אופים את הוופל על הכיריים לפחות 45 דקות.
- מוציאים את הוופל מהחם. שים את מסכת הצילום (ראה קובץ משלים 4) (צד אמולסיה כלפי מטה) על גבי הוופל והכביד עליו עם דיסק זכוכית כדי להבטיח מגע בין מסכת הצילום לשקופית.
- לחשוף עם אור UV (365 ננומטר) עבור צפיפות אנרגיה קורנת של 350 mJ / ס"מ2.
- אופים רקיק על הכיריים במשך 12-15 דקות.
- מניחים רקיק במיכל זכוכית רחב. לכסות רקיק עם פתרון מפתח SU-8. מניחים על שייקר ומתפתחים תוך כדי עצבנות במשך 15 דקות לפחות.
- השתמש במלקחיים כדי להסיר את הוופל מפתרון המפתח. יש לשטוף במשך 5 s על ידי ריסוס תמיסת מפתחים נוספת מבקבוק התזה. יש לרסס עם אלכוהול איזופרופיל לשטיפה. אם מופיע משקעים לבנים, החזר את הוופל לפתרון המפתח והתפתח למשך זמן רב יותר.
- רקיק יבש מתחת לזרם של אוויר או חנקן.
- אופים שקופית במשך 5 דקות.
הערה: לאחר יצירת הוופל הראשי, ניתן לעשות בו שימוש חוזר ללא הגבלת זמן כל עוד התכונות נשארות ללא פגע.
- הכן PDMS.
- בסירת שקילה, הוסף אלסטומר רב-ידילסילוקסן וצליבה ביחס של 10:1 (במסה). מערבבים במרץ כדי להבטיח ערבוב אחיד.
- פירוק גז PDMS במתן ואקום במשך 15-30 דקות עד שלא ניתן לראות בועות נוספות.
- מניחים את הוופל הראשי בצלחת תרבית רקמות 15 ס"מ. יוצקים PDMS על הוופל. אם מופיעות בועות, יש פירוק גז במתן ואקום למשך מספר דקות.
- אופים בתנור 60 מעלות צלזיוס במשך 3 שעות.
הערה: לאחר האפייה, ניתן לאחסן תאי זרימת PDMS על הספסל ללא הגבלת זמן.
- הכן תאי זרימת PDMS לניסוי.
- זמן קצר לפני תחילת ניסוי CMO-DPAC, חתוך את המספר הנדרש של תאי זרימת PDMS מן הוופל הראשי. פלזמה להתחמצן עם אוויר חדר 10 סמ"ק / דקה עבור 90 s כדי להפוך את פני השטח הידרופילי.
- חותכים כל תא זרימה בודד כך שיש 1-2 מ"מ של PDMS שנותרו בכל צד, ולאחר מכן חתכו את החלק העליון והתחתון של תא הזרימה כדי ליצור פתח ושקע.
- אחזר שקופית בדוגמת דפוס שנוצרה בשלבים 2 ו- 3. ישר על גבי מסכת תמונה.
- באמצעות מסכת התמונות כהפניה, מקם את תאי הזרימה PDMS בשקופית במיקום של כל אזור עם תבנית.
- הוסף 50 μL של תמיסת מלח חוצץ פוספט (PBS) + 1% אלבומין סרום בקר (BSA) ל- inlet של כל תא זרימה, כפי שמוצג איור משלים 1H. ודא כי תא הזרימה מלא לחלוטין על ידי PBS + 1% BSA וכי אין בועות גדולות. המשך מיד לשלבים 5 ו-6.
הערה: חסימה באמצעות BSA ממזערת הידבקות תאים לא ספציפית למשטח השקופית.
5. להרים ולתייג תאים עם DNA שונה כולסטרול
- הכן את פתרונות ה- DNA שעברו שינוי כולסטרול.
- עבור כל קבוצה של תאים בניסוי, לערבב יחד 3 μL של פתרון מלאי 100 מיקרומטר של גדיל עוגן אוניברסלי שונה כולסטרול עם 3 μL של פתרון מלאי 100 מיקרומטר של מתאם סטרנד. דגירה לדקה אחת. זה יקדם-הכלאה את האוליגו. הוסף 69 μL של תמיסת מלח חוצץ פוספט (PBS) כדי ליצור פתרון עוגן אוניברסלי + מתאם 4 מיקרומטר.
- עבור כל קבוצה של תאים בניסוי, להוסיף 3 μL של 100 μM אוניברסלי כולסטרול שונה כולסטרול מניות פתרון 12 μL של PBS, יצירת פתרון 20 מיקרומטר.
- הכן את ההשעיות של תא בודד.
- עבור תאים חסידים, השתמש טריפסין או סוכן דיסוציאציה אחר כדי להסיר את התאים מבקבוק התרבות. הוסף מדיה תרבית כדי לנטרל את טריפסין וצנטריפוגה כדי גלולה התאים. עבור תאים שאינם דבקים, לאסוף את השעיית התא צנטריפוגה כדי כדורי התאים.
- resuspend גלולה התא ב 1 מ"ל של PBS קר כקרח או מדיה ללא סרום. העבר 1-3 מיליון תאים לצינור מיקרוצנטריפוגה 1.5 מ"ל. צנטריפוגה ב 160 x g במשך 4 דקות.
הערה: אם סוג התא הנמצא בשימוש נוטה לגוש/צבירה, השתמש ב- PBS ללא יוני סידן ומגנזיום לכל שלבי הכביסה כדי להפחית צבירה לא רצויה של תאים. אם הכדאיות היא דאגה מיוחדת לסוג התא הנמצא בשימוש, השתמש במדיה נטולת סרום במקום PBS. מדיה המכילה סרום בקר עוברי אינה מומלצת לתיוג תאים מכיוון שהיא עלולה לעכב שילוב של אוליגוס שעברו שינוי שומנים בדם. 35
- לתייג את התאים עם אוליגוס שינוי כולסטרול.
- resuspend גלולה התא ב 75 μL של PBS קר כקרח או מדיה ללא סרום. שמור את התאים בדלי קרח לאורך כל תהליך התיוג והשטיפה כדי למקסם את הכדאיות של התא ולמזער את אובדן האוליגו שעבר שינוי כולסטרול מפני השטח של התא.
הערה: שימוש חוזר בתאים לפני הוספת ה- DNA מבטיח כי התפלגות ה- DNA אחידה על פני אוכלוסיית התאים. - הוסף את 75 μL של 4 מיקרומטר אוניברסלי עוגן + פתרון מתאם שנוצר בשלב 5.1.1 לצינור microcentrifuge המכיל את מתלה התא. מערבבים היטב על ידי צנרת. דגירה במשך 5 דקות על קרח.
- הוסף 15 μL של פתרון העוגן המשותף האוניברסלי לצינור microcentrifuge. מערבבים היטב על ידי צנרת. דגירה במשך 5 דקות על קרח.
- הסר אוליגוס עודף מן השעיית התא. הוסיפו מ"ל אחד של PBS קר כקרח או מדיה נטולת סרום לצינור המיקרוצנטריפוגה. מערבבים עם פיפטה P1000. צנטריפוגה ב 160 x g במשך 4 דקות ב 4 °C (50 °F). זרוק את סופר-טבעי. חזור על הפעולה פעמיים נוספות.
הערה: אם תאים נוטים להתגבש, להעביר את ההשעיה התא דרך מסנן 40 מיקרומטר לפני הכביסה הסופית. אם תאים נוטים ספיחה על הצד של צינור microcentrifuge, לשקול מראש חסימת הצינור עם קזאין.
- resuspend גלולה התא ב 75 μL של PBS קר כקרח או מדיה ללא סרום. שמור את התאים בדלי קרח לאורך כל תהליך התיוג והשטיפה כדי למקסם את הכדאיות של התא ולמזער את אובדן האוליגו שעבר שינוי כולסטרול מפני השטח של התא.
6. תבנית התאים המסומנים בדנ"א
- Resuspend התאים PBS קר כקרח או מדיה ללא סרום כדי ליצור פתרון תא צפוף של לפחות 25 מיליון תאים / מ"ל.
הערה: עבור שקופית אחת באמצעות ארבעה של 10 מ"מ x 15 מ"מ x 200 מיקרומטר תאי זרימה PDMS המתוארים בשלב 4, נדרש כ 100 μL של השעיית תא צפוף זה. למרות שרוב התאים האלה לא ידבקו בדפוס ובסופו של דבר ייזרקו, פתרון מרוכז מאוד של תאים על פני התבנית משפר באופן דרמטי את היעילות של דפוס התא. - הרם שקופית והטה אותה מעט. הוסף 25 μL של השעיית תא ל- inlet של כל תא זרימה בשקופית עם התבנית. הסר את פתרון PBS + 1% BSA מהשקע, ומאפשר להשעיית התא למלא את תא הזרימה PDMS. דגירה על קרח או בטמפרטורת החדר במשך 30 מעלות.
הערה: בשלב זה, התבוננות בתא הזרימה מתחת למיקרוסקופ צריכה להראות תאים צפופים עם מעט מאוד רווחים גלויים בין תאים. ראו איור משלים 2B. - שאף 5 μL של השעיית תא מהשקע של השקופית ולהוסיף אותו בחזרה לתוך הכניסה. חזור על הפעולה 10 פעמים בכל תא זרימה.
הערה: הידבקותם של תאים המסומנים בתווית CMO לשקופית בדוגמת DNA היא כמעט מיידית. זרימת התאים מעל התבנית מספר פעמים מגדילה את ההסתברות שתא יזרום מעל כתם דנ"א נתון ויתפס. - צנרת PBS בעדינות או מדיה ללא סרום לתוך הכניסה של כל תא זרימה לשטוף תאים עודפים. לאסוף את השעיית התא מהשקע. חזור על הפעולה 2-4 פעמים או עד בדיקה חזותית של השקופית מתחת למיקרוסקופ מאשר כי לא נותרו תאים עודפים.
הערה: זה יכול להיות יתרון כדי להציל את התאים עודפים מן לשטוף הראשון. אם יעילות התבניות אינה משביעת רצון, ניתן לצנטריפוג את התאים העודפים ולתעבד מחדש בנפח נמוך יותר של PBS כדי ליצור פתרון צפוף יותר בתאים, ואז ניתן לחזור על התהליך בשלב 6.2. - חזור על שלבים 6.1-6.4 עבור כל ערכת תאים בתבנית. עבור תבניות שבהן סוגי תאים מרובים מעוצבים ישירות על-ידי תבנית פני השטח, התחל בסוג התא הפחות שופע של התבנית וסיים בסוג התא הנפוץ ביותר.
הערה: מומלץ לעשות כל סיבוב של הרכבה סלולרית ברצף במקום לאגד את התאים, אפילו בתנאים שבהם התאים מסומנים כולם עם רצפי DNA אורתוגונליים. איגום התאים מדלל ביעילות כל אוכלוסיית תאים ומפחית את יעילות התבנית. - לאחר השלמת הסיבוב האחרון של הרכבת התא, השלבים הבאים ישתנו בהתאם לניסוי הספציפי. אם התאים נועדו להישאר על הזכוכית, הוסיפו מדיה לצלחת פטרי המכילה את השקופית ולאחר מכן השתמשו בעדינות במלקחיים כדי לדחוף את תאי זרימת ה-PDMS מהשקופית. אם התאים יוטבעו בהידרוג'ל ויתרבדו בתלת-ממד, המשיכו לשלב 7.
7. העבר להידרוג'ל לתרבות תלת-ממדית (אופציונלי)
- הכן פתרון מבשר הידרוג'ל המכיל 2% DNase.
הערה: הרכב הפתרון ישתנה בהתאם להתקנה ניסיונית. מטריגל ותערובות של מטריגל וקולגן אני עובד היטב בפרוטוקול זה, אבל הידרוג'לים אחרים אפשריים גם כן. - הוסף 50 μL של פתרון הידרוג'ל המכיל 2% DNase ל- inlet של כל תא זרימה. שאפו את הנוזל העודף מהשקע, נהיגה פתרון הידרוג'ל לתוך תא הזרימה. עבור מבשרי הידרוג'ל צמיגים, הטיית השקופית מעט עשויה להידרש כדי לעזור לזרימת ההידרוג'ל לתא הזרימה.
- לדגור על השקופית ב 37 °C (30-45 דקות(בהתאם קינטיקה ג'ל הידרוג'ל) כדי לאפשר הידרוג'ל להגדיר ולבקע את ההידבקות מבוססת ה- DNA בין התאים לבין פני השטח.
- הסר כל תא זרימה מהשקופית והצב מעל תמיסת מבשרת הידרוג'ל.
- הוסף 50 μL של הקדמה הידרוג'ל לבאר של שקופית תא 2-באר או צלחת 6-באר.
- Pipette 10 μL של PBS משני צדי כל תא זרימה.
- השתמש בסכין גילוח או פינצטה עדינה כדי להפיץ את ה- PBS לאורך מלא של תא הזרימה, ולאחר מכן הרם בעדינות את צידי תא הזרימה כך שה- PBS ימהר מתחת להידרוג'ל.
הערה: זה יהיה "לצוף" הידרוג'ל על פני השקופית, המאפשר העברה ללא עיוות או קרע. - השתמש בסכין גילוח כדי להזיז בעדינות את תא הזרימה לקצה מגלשת הזכוכית.
- הפוך את השקופית. עם סכין הגילוח, לדחוף את תא הזרימה מהמגלשה כך שהוא נוחת על גבי סכין הגילוח.
- בחר את תא הזרימה את סכין הגילוח באמצעות מלקחיים מעוקלים. הפוך את תא הזרימה כך שהתאים נמצאים בתחתית, ולאחר מכן מניחים על גבי טיפת תמיסת הקדמת הידרוג'ל.
- חזור על שלבים 7.4.1 - 7.4.6 עבור כל תא זרימה.
- דגירה במשך 30 דקות לפחות, כך הידרוג'ל המכיל את התאים בדוגמת יכול להיקשר underlay הידרוג'ל, וכתוצאה מכך הטמעה מלאה של התאים בדוגמת.
- הסר את תא הזרימה PDMS.
- הוסיפו מספיק מדיה כדי לטבול את תא הזרימה של PDMS.
הערה: זרם המדיה ישחרר את ההידבקות בין ההידרוג'ל לתא הזרימה PDMS. - השתמש במלקחיים מעוקלים, המכוונים לאורך הציר הארוך של תא הזרימה, כדי לדחוף בעדינות את תא הזרימה עד שהוא קופץ וצף לתוך המדיה. לאסוף את תא הזרימה עם מלקחיים ולהשליך.
הערה: לקבלת תוצאות מיטביות, פזורים את המלקחיים המעוקלים והפעילו לחץ עדין על קירות תא הזרימה PDMS. החל כוח בכיוון הציר הארוך של תא הזרימה.
- הוסיפו מספיק מדיה כדי לטבול את תא הזרימה של PDMS.
8. אשר תיוג מוצלח של תאים עם CMO (אופציונלי, לפתרון בעיות)
- הזמינו אוליגונוקלאוטיד שעבר שינוי פלואורסצנטרי (FAM או AF647) המשלים את רצף הידבקות פני השטח של סטרנד המתאם הנמצא בשימוש בניסוי.
- סמן תאים עם DNA CMO ושטוף את ה- DNA העודף כמתואר בשלב 5. Resuspend ב 200 μL של PBS קר כקרח.
- הפוך פתרון 4 מיקרומטר של oligonucleotide משלימה שכותרתו פלואורסצנטית ב- PBS. הוסף 200 μL של פתרון זה השעיית התא. דגירה על קרח במשך 5 דקות.
- הוסף 1 מ"ל של PBS קר כקרח. מערבבים, צנטריפוגות את התאים כדי ללגלג אותם. הסר סופר-טבעי. חזור על תהליך זה פעמיים נוספות כדי לשטוף את כל ה- DNA שלא היברידי.
- בצע ציטומטריית זרימה אנליטית כדי לכמת את נוכחות ה- DNA על פני התא.
- על ציטומטר זרימה, לנתח תאי בקרה שלא תויגו עם DNA. להקים שערים המבוססים על אוכלוסייה זו.
- נתח תאים בעלי תווית CMO שטופלו באוליגונוקלאוטיד משלים עם תווית פלואורסצנטית.
- חשב עוצמת פלואורסצנטיות ממוצעת.
פרוטוקול זה מאפשר דפוס תאים 2D ותלת-ממד עם דיוק גבוה וללא שימוש ריאגנטים מותאמים אישית או ציוד ניקוי יקר. איור 1 מציג מבט כולל על הפרוטוקול. ראשית, שקופיות פונקציונליות DNA נוצרות באמצעות פוטוליתוגרפיה. לאחר מכן, תאים מסומנים ב- CMOs. לאחר מכן, התאים זורמים מעל השקופית, שם הם מתחברים רק לאזורים הפונקציונליים של ה- DNA של השקופית. לאחר תאים עודפים נשטפים, התבנית הרצויה של תאים מתגלה. תאים אלה ניתנים לתרבית בשקופית או מוטבעים בהידרוג'ל המכיל DNase ומועברים מהשקופית עבור תרבית תאים תלת-ממדית.
תיוג של תאים עם CMOs מאפשר את ההתקשרות שלהם לשקופית בדוגמת DNA (איור 2). ראשית, סטרנד העוגן האוניברסלי שעבר שינוי כולסטרול הוא היברידי מראש עם סטרנד המתאם. לאחר מכן, פתרון העוגן האוניברסלי + מתאם מעורבב 1:1 עם ההשעיה של התא. הכולסטרול על קומפלקס העוגן האוניברסלי + מתאם מכניס לתוך קרום התא. תוספת של סטרנד עוגן אוניברסלי שונה כולסטרול, אשר הכלאה עם גדיל עוגן אוניברסלי, משפר את היציבות של קומפלקס CMO בקרום התא על ידי הגדלת ההידרופוביה נטו של מורכב26. לאחר שטיפת ה- DNA העודף מהשעיית התא, התאים זורמים מעל השקופית. הכלאה בין סטרנד המתאם לבין גדיל ה- DNA של פני השטח גורמת לחיבור של תאים לאזורים בדוגמת DNA של השקופית.
התבנית של התאים נוצרת באמצעות פוטוליטוגרפיה כדי להגביל את ההתקשרות של אוליגוס DNA שעברו שינוי אמין לאזורים ספציפיים של שקופית זכוכית מותאמת אלדהיד29 (איור 3A). פוטורסיסט חיובי מצופה ספין על שקופית פונקציונלית אלדהיד. לאחר מכן, מסכת צילום של שקיפות ממוקמת על גבי השקופית והשקופית נחשפת לאור UV. לאחר התפתחות, אזורי השקופית שנחשפו לאור UV אינם מצופים עוד בפוטורסיסט ולכן חשפו קבוצות אלדהיד. פתרון 20 מיקרומטר של אוליגוס DNA שונה אמין הוא ירד לאחר מכן על השקופית להתפשט כדי לכסות את האזורים בדוגמת. אפייה ואחריה הפחתת תוצאות קשר קוולנטי בין ה- DNA שעבר שינוי אמין לבין השקופית. למרבה הפלא, ניתן לחזור על תהליך זה כדי ליצור אוליגוס מרובים ללא כל אובדן פונקציונליות של אוליגוס בדוגמת בעבר (איור 3B). עם זאת, יש להקפיד להימנע מדפוסים חופפים, מה שגורם לנוכחות של שני האוליגו בריכוז מופחת (איור 3 משלים). ניתן ליצור דפוסים של אוכלוסיות תאים מרובות ברצף באמצעות גדילי מתאם השונים בתחום המודולרי שלהם (20 הבסיסים הקרובים ביותר לקצה 3').
למרות פרוטוקול photopatterning זה פותח על ידי Scheideler ואח 'בהקשר של חדר נקי, הוכחנו כי ניתן להשיג תוצאות דומות עם זול, "לחלוט הביתה" הגדרת פוטוליתוגרפיה שמתאימה בקלות בתוך מכסה המנוע אדים כימי. ההתקנה כוללת מעיל ספין בשווי 400 דולר העשוי ממנוע DC, בקר דיגיטלי וקופסת עוגת תקליטורים, וכן מנורת UV שהורכבה מרכיבים בודדים ושוכנת במיכל חדות המיועד מחדש(איור 1 משלים). היתרון העיקרי של הגדרת פוטוליתוגרפיה לחלוט הביתה הוא שזה זול מאוד (< $1000 עבור כל הציוד) תוך עדיין להיות מסוגל ליצור תכונות בגודל תא יחיד. עם זאת, השימוש בציוד זול יש מגבלות שלה - למשל, זה מאתגר יותר ליישר במדויק סמנים fiducial דפוס אוליגוס DNA מרובים ללא שימוש של קשת מסכה. אנו ממליצים על הגדרת פוטוליתוגרפיה זולה זו למעבדות שאין להן גישה נוחה לחדר נקי או שרוצות לנסות שיטה זו ללא השקעה גדולה.
כדי לזהות תנאים אופטימליים להידבקות תאים מתוכנתי DNA, שינינו באופן שיטתי את הריכוזים של גדילי DNA על משטחי התא ומדדנו את היעילות של הידבקות תאים למשטחי זכוכית שעברו שינוי דנ"א. הריכוז של עוגן אוניברסלי + מתאם סטרנד ועוגן קו-עוגן אוניברסלי בפתרונות תיוג היו מגוונים על פני מספר סדרי גודל (איור 4A,B),וכתוצאה מכך 104 - 106 מתחמי DNA לתא(איור 4 משלים). הידבקות התא הייתה תלויה במינון, עם הידבקות תאים מינימלית לתבנית ה- DNA כאשר תאים סומנו עם CMOs בריכוז של 0.05 מיקרומטר או פחות, ותפוסה גבוהה בריכוז של 2.5 מיקרומטר ומעלה. אנו, אם כן, השתמשנו בפתרון 2 מיקרומטר של עוגן אוניברסלי + מתאם סטרנד ופתרון 2 מיקרומטר של עוגן קו-עוגן אוניברסלי ברוב הניסויים. הידבקות התא צפויה גם להקטין אם כמות ה- DNA המשמש על משטח הזכוכיתירדה 29 או אם אי התאמות בין גדיל מתאם גדיל משטח גדל. מידע נוסף אודות עיצוב רצף מתאם Strand מסופק בקובץ משלים 2. תיוג CMO באמצעות גדילי מתאם ללא חזרות CpG לא לעורר TLR9 בתאי HEK מבטא עכבר TLR9 (איור משלים 5).
אנו מספקים מספר הדגמות שהפרוטוקול המתוקן מספק הידבקות תאית מתוכנתת DNA ניתנת לשחזור ויעילה. לדוגמה, תאי אנדותל וריד טבור אנושי (HUVECs) המסומנים עם CMOs דבק דפוסי DNA עם יעילות גבוהה. HUVECs עם תווית CMO דבקה כמו גם HUVECs עם תווית LMO(איור 5A). תאים בעלי תבנית באמצעות CMO-DPAC שמרו על הכדאיות והפונקציונליות שלהם. תאים המסומנים ב-CMOs הוכתמו על ידי קלצ'ין AM ואתידיום הומודימר כדי להעריך את הכדאיות(איור משלים 6). ההבדלים הכדאיות לעומת תאי בקרה ללא תווית היו קטנים (94% לעומת 97%). חברי פרל"ק בודדים שהועברו באמצעות CMO-DPAC והועברו למטריג'ל הצליחו להתרבות ולקטב כראוי לאחר 5 ימים של תרבות(איור 5B). DPAC מספק גם אמצעי להרחבת דפוסי תאים לתוך המימד השלישי(איור 5C). לדוגמה, ניתן ליצור אגרגטים רב-שכבתיים ורב-תאיים על-ידי שכבות לסירוגין של תאים המסומנים ב- CMOs משלימים (איור 5C). ניסויים אלה מוכיחים כי הפרוטוקול ניתן לשחזור, אינו משפיע לרעה על הכדאיות או הפונקציונליות של התאים, ומניב דפוסים תאיים שניתן לתרבות בהצלחה בתוך מישור הדמיה יחיד ב- ECM תלת-ממדי.
על-ידי מתן רצפי דנ"א אורתוגונליים להדבקה ישירה של תאים, DPAC מספק אמצעי ליצירת דפוס של סוגי תאים מרובים על משטח יחיד. כדי ליישם תכונה זו של DPAC, דפוסי DNA שנוצרו על ידי פוטוליתוגרפיה חייבים להיות מיושרים ביחס זה לזה. סמני נאמנות מתכת שהופקדו על השקופית מותר יישור של מסכות צילום מרובות ולכן דפוס של סוגי תאים מרובים בבת אחת. MCF10As מוכתמים בצבעים ייחודיים שונים סומנו עם CMOs אורתוגונל ודוגמת כדי ליצור הדמיה של הלוגואים UC ברקלי ו- UCSF(איור 6). ניסוי זה מדגים כי אוכלוסיות תאים ייחודיות מרובות יכולות להיות מעוצבות יחד עם דיוק גבוה וללא זיהום צולב.
דפוס מוצלח של תאים באמצעות CMO-DPAC דורש פוטוליטוגרפיה באיכות גבוהה, ריכוז מספיק של אוליגו על פני התא, צפיפות גבוהה של תאים על פני התבנית, ושטיפה מספקת. כישלון של אחד מהצעדים הללו משפיע על התוצאה הסופית. איור 2 משלים כולל תמונות לדוגמה של פוטוליתוגרפיה נכונה ולא נכונה (איור משלים 2A), צפיפות התאים הרצויה מעל התבנית ליצירת דפוסים תפוסים לחלוטין (איור משלים 2B),אובדן תאים בדוגמת עקב צנרת נמרצת מדי במהלך השלבים הבאים של DPAC (איור משלים 2C),וגושים לא רצויים של תאים (איור 2D משלים). טבלה 1 כוללת רשימה של נקודות כשל נפוצות ופתרון הבעיות המוצע. השימוש באוליגו משלימים פלואורסצנטיים מומלץ ככלי לפתרון בעיות כדי לאשר את נוכחותו של DNA בדוגמת בשקופית ואת נוכחותם של CMOs על פני התא על ידי ציטומטריית זרימה (ראה שלב 8 של פרוטוקול).
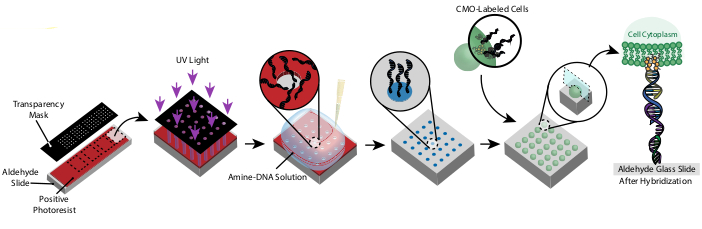
איור 1: מבט כולל על פרוטוקול CMO-DPAC. ראשית, שקופית בדוגמת DNA נוצרת על ידי ציפוי שקופית זכוכית פונקציונלית אלדהיד עם פוטורזיסט חיובי, מכסה אותו במסכת שקיפות בתבנית הרצויה, וחשיפה לאור UV. הפוטורסיסט חשוף UV נשטף עם מפתח, משאיר אזורים חשופים של שקופית אלדהיד ומאפשר כריכה של DNA פונקציונלי אמין על פני השטח. לאחר מכן, התאים מסומנים ב- CMOs וזורמים על פני השטח. הדנ"א על קרום התא מתהפך לדנ"א על פני השטח, וכתוצאה מכך הידבקות. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
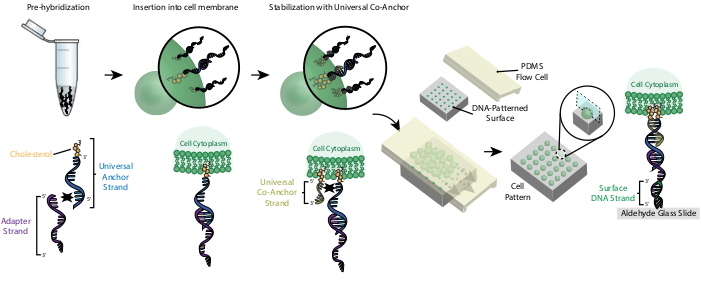
איור 2: תאים מסומנים ב- CMOs בתהליך צעד. ראשית, סטרנד העוגן האוניברסלי שעבר שינוי כולסטרול הוא היברידי מראש עם סטרנד המתאם. לאחר מכן, פתרון העוגן האוניברסלי + מתאם מעורבב עם מתלה התא. הכולסטרול על קומפלקס העוגן האוניברסלי + מתאם מכניס לתוך קרום התא. לאחר הדגירה, סטרנד העוגן האוניברסלי שעבר שינוי כולסטרול נוסף להשעיית התא, שם הוא מתכלא עם גדיל העוגן האוניברסלי ומכניס לקרום התא. התוספת של מולקולת הכולסטרול השנייה מגבירה את ההידרופוביה נטו של קומפלקס ה- DNA ומייצבת אותו בתוך הממברנה26. לאחר שטיפת ה- DNA העודף, התאים מרוכזים ומתווספים לתא זרימת PDMS על גבי פני השטח בדוגמת. הקצה של המתאם סטרנד מתכלה עם סטרנד ה- DNA של פני השטח על שקופית הזכוכית, וכתוצאה מכך הידבקות בשקופית במיוחד באזורים פונקציונליים עם DNA משלים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
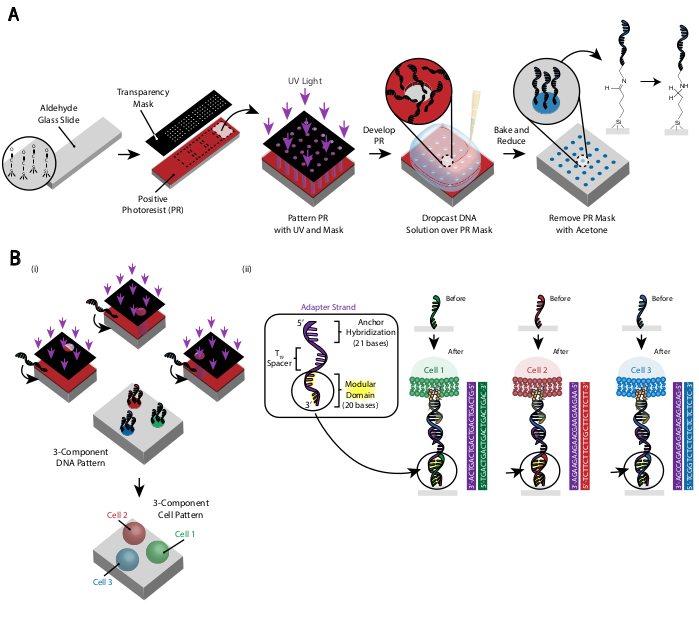
איור 3: פוטוליתוגרפיה משמשת ליצירת שקופיות בדוגמת DNA שבסופו של דבר יכתיבו את מיקום התאים. (A)מבט כולל על תהליך פוטוליתוגרפיה. שקופית פונקציונלית של אלדהיד מצופה בפוטורסיסט חיובי. אור UV זורח על השקופית דרך מסכת צילום שקיפות שקופה במקום שבו נדרשת הידבקות תאים. לאחר פיתוח השקופית, האזורים שנחשפו בעבר לאור UV חשפו כעת קבוצות אלדהיד. פתרון של 20 מיקרומטר של אוליגו DNA פונקציונלי אמין הוא ירד לאחר מכן על השקופית להתפשט על פני האזורים בדוגמת. לאחר מכן, השקופית אפויה כדי לגרום להיווצרות קשרים של שיף (C= N) בין קבוצות האמין והאלדהיד, קשר קוולנטי הפיך29. לאחר מכן, הפחתת ספיגה עם 0.25% נתרן בורוהידיד ב-PBS ממירה את בסיס שיף לאמין משני על ידי הפחתת הכנסה, וכתוצאה מכך קשר בלתי הפיך בין ה- DNA לבין השקופית. לאחר מכן ניתן להסיר את הפוטורסיסט הנותר על ידי שטיפה עם אצטון. (B)ניתן לחזור על תהליך זה כדי ליצור דפוסי DNA מרובי רכיבים ולכן לבצע ניסויים עם אוכלוסיות תאים מרובות. (i) לאחר האוליגו הראשון הוא בדוגמת, השקופית מצופה שוב photoresist ואת הפרוטוקול ממשיך כמו קודם. יישור של מסכות הצילום באמצעות סמני נאמנות נחוץ לדפוס גדילי DNA מרובים. (ii) כל סוג תא בדוגמת דפוס שונה בקבוצת המחשבים המודולרית בעלת 20 הבסיסים של המתאם Strand. באמצעות קבוצות אורתוגונליות של אוליגוס משלימים, סוגי תאים מרובים יכולים להיות בדוגמת ללא הידבקות צולבת. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
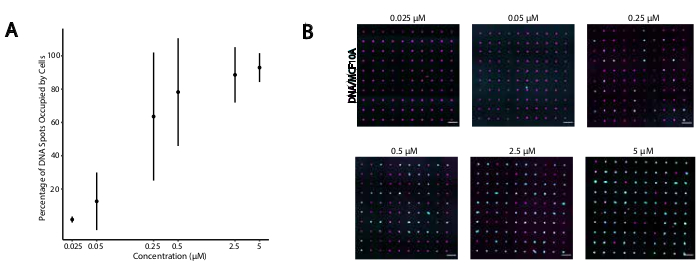
איור 4: הידבקות של תאים המסומנים על-ידי CMO לדפוסי דנ"א עולה כפונקציה של ריכוז CMO במהלך התיוג. בניסוי זה, העוגן האוניברסלי + סטרנד מתאם (טרום היברידי) ואת עוגן קו אוניברסלי שימש בריכוזים שווים. ריכוז מתייחס לריכוז של CMO בהשעיית התא במהלך תיוג CMO של תאים. (A)כימות האחוז של כתמי DNA בקוטר 15 מיקרומטר שנכבשו על ידי תאי MCF10A המסומנים CMO כפונקציה של ריכוז CMO במהלך תיוג תאים. הנתונים המיוצגים כממוצע ± סטיית התקן משלושה ניסויים. (B)תמונות מייצגות של דפוסי ה- DNA (מגנטה) ו- MCF10As (ציאן) דבק בריכוזים שונים של CMO. סרגל קנה מידה = 100 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
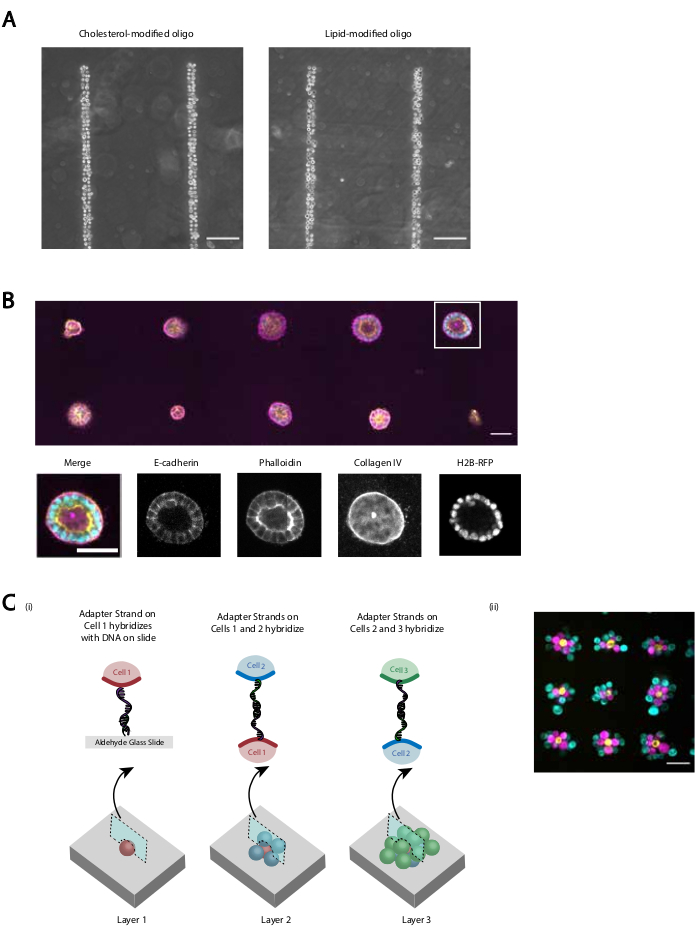
איור 5: ניתן להשתמש ב-CMO-DPAC ליצירת דפוסי תאים דו-ממדיים שניתן להטמיע לאחר מכן בהידרוג'ל תלת-ממדי לתרבות ו/או לשכבה ליצירת מבנים רב-שכבתיים. (A)השוואה ישירה בין תאי אנדותל ורידים אנושיים המסומנים על ידי CMO (HUVECs) לבין HUVECs המסומנים בתווית LMO דבקו בתבנית DNA ליניארית. שתי השיטות לתיוג תאים גורמות לתפוסה של כמעט 100% בדפוס הדנ"א. (B)תאי כליה יחידים של מאדין-דארבי (MDCKs) המביעים H2B-RFP היו בדוגמת כתמים בקוטר 15 מיקרומטר במרווח זה מזה של 200 מיקרומטר ולאחר מכן מוטמעים מטריגל. לאחר 120 שעות של תרבות, ציסטות אפיתל וכתוצאה מכך תוקנו ומוכתמים עבור E-cadherin, actin, וקולגן IV. ספרואיד בקופסה לבנה מוצג בפירוט. סרגל קנה מידה = 50 מיקרומטר. (C)ניתן ליצור מבנים סלולריים רב שכבתיים על-ידי סימון אוכלוסיות תאים נפרדות עם גדילי מתאם משלימים ותבניות ברצף, כך שכל תוספת חדשה של תאים דבקה בשכבת התא לפניה. (i)שרטוט של דפוס רציף של אוכלוסיות תאים כדי ליצור מבנים רב שכבתיים. (ii) אגרגטים של תאים תלת-שכבתיים של MCF10As (דמיינו באמצעות צבעים) נוצרו באמצעות תהליך זה. סרגל קנה מידה = 50 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

איור 6: ניתן דפוס של סוגי תאים מרובים ללא זיהום צולב או אובדן הידבקות. אוליגוס דנ"א מרובים שעברו שינוי אמין נדגמו ברצף על שקופית אלדהיד ויישרו באמצעות סמני נאמנות ממתכת. שלוש אוכלוסיות של MCF10As (ציאן, מגנטה, צהוב) הוכתמו בצבעים ייחודיים המסומנים ב- CMOs משלימים, ודוגמנו על השקופית, וכתוצאה מכך תמונה של הלוגו של UC ברקלי ו- UCSF. סרגל קנה מידה 1 מ"מ. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
איור משלים 1: תמונות לדוגמה של הגדרת הפוטוליתוגרפיה של הספסל. (A)החלק על מעיל ספין, מכוסה פוטורסיסט חיובי, לפני ציפוי ספין. (B)תמונה של מסכת תמונות של שקיפות. (ג)במהלך החשיפה, מסכת הצילום נדחקת בין המגלשה מצופה הפוטורסיסט לדיסק זכוכית. (D)דיור למנורת UV היה עשוי ממיכל חדים ייעודי. (E)שקופית שקועה בפתרון מפתחים. (ו)שקופית מפותחת. (G)תמיסת DNA במין שונה התפשטה על אזורים בדוגמת השקופית. (H)תאי זרימת PDMS הממוקמים על גבי אזורים בדוגמת מילוי של השקופית. נא לחץ כאן כדי להוריד קובץ זה.
איור 2 משלים: כמה דוגמאות לכשלים נפוצים בפרוטוקול זה. (A)(i)תת אפייה לפני חשיפה לקרינת UV או תכונות פיתוח יתר לאחר החשיפה עלולות לגרום לתכונות שיש להן קצוות משוננים ועשויות להיות לא סדירות בגודלן. (ii)דוגמה לשקופית עם צילומים נכונים עם קצוות נקיים סביב תכונות, גודל תכונה אחיד וללא סדקים ברורים בתבנית. סרגל קנה מידה = 50 מיקרומטר. (B)צפיפות התאים חיונית ליעילות התבנית. בעת התבוננות בתאים מעל התבנית מתחת למיקרוסקופ, יש קיימים פערים מעטים בין תאים, כפי שמעידה התמונה לדוגמה משמאל. סרגל קנה מידה = 50 מיקרומטר. (C)תאים בדוגמת דפוס יכולים להיות רגישים לכוחות נוזלים הנובעים מצינורות נמרצים מדי, אשר יכולים לפגוע ולחלץ את התאים בדוגמת. צבירי תאים מרובי שכבות פגיעים במיוחד, מכיוון שתא אחד בתחתית תומך במבנה של תאים מרובים. (i)מערך של צבירות תאים מוטבע בהצלחה לתוך מטריגל. (ii)רשת של אגרגטים תאים שהתנתקו כתוצאה ממטריג צמיג צנרת במרץ רב מדי. (ד)גוש תאים יכול להתרחש, במיוחד עם תאי אפיתל. גושים אלה הם בדרך כלל הומוטיפיקים אך יכולים להיות הטרוטיפיקים (תאים הנצמדים לתאים שכבר בדוגמתם מסוג אחר) אם התאים דביקים במיוחד. התמונה מראה שלוש אוכלוסיות שונות של MCF10As היו בדוגמת על מערך המורכב משלושה כתמי DNA שונים בגודל תא יחיד (15 מיקרומטר). ברוב כתמי הדנ"א מחוברים 2-4 תאים. ניתן לפתור את הגושים על ידי טיפול EDTA או על ידי סינון הגושים לפני הדפוס. סרגל קנה מידה = 100 מיקרומטר. נא לחץ כאן כדי להוריד קובץ זה.
איור משלים 3: פוטופטרנים חופפים גורמים לנוכחות של שני האוליגו בריכוז מופחת. שני אוליגוס שעברו שינוי אמין אורתוגונלית היו פוטו-מרופטים ברצף, תחילה קו אנכי (Strand 1), ואחריו קו אופקי שחפף אותו (סטרנד 2). האוליגו היו אז לדמיין על ידי הכלאה עם אוליגוס משלימים פלואורסצנטי. (א)תמונת פלואורסצנטיות של סטרנד 1. (B)כימות פרופיל הפלואורסצנטיות של סטרנד 1 על פני קו אנכי של 100 מיקרומטר המשתרע על פני החפיפה. (C)תמונת פלואורסצנטיות של סטרנד 2. (D)כימות של פרופיל הפלואורסצנטיות של סטרנד 2 על פני קו אופקי של 100 מיקרומטר המשתרע על פני החפיפה. סרגל קנה מידה = 50 מיקרומטר. נא לחץ כאן כדי להוריד קובץ זה.
איור משלים 4: כימות מתחמי DNA על פני התא כפונקציה של ריכוז תיוג CMO. HUVECs סומנו בריכוזים שונים של פתרון CMO, נשטף, ולאחר מכן דגירה עם גדיל משלימה פלואורסצנטית. ערכת מיקרוספירה של MESF (מולקולות של פלואורוכרום מסיס שווה ערך) שימשה לביצוע ציטומטריית זרימה כמותית ולהעריך את מספר מתחמי ה- DNA על פני התא כפונקציה של ריכוז CMO במהלך התיוג. נא לחץ כאן כדי להוריד קובץ זה.
איור משלים 5: תיוג CMO אינו מעורר את תגובת TLR9. בוצע ניסוי כדי לראות אם תיוג CMO יפעיל את מנגנון זיהוי ה- DNA של TLR9 והאם זה יושפע על ידי CPGs ברצף סטרנד מתאם. תאי HEK המבטאים עכבר TLR9 דוגרו בן לילה עם 0.2 מיקרומטר של ODN 1826 (אגוניסט TLR9 המכיל CpG), עוגן אוניברסלי CMO + עוגן אוניברסלי + גדיל מתאם המכיל את אותו רצף כמו ODN 1826 (CMO-CpG), או עוגן אוניברסלי CMO + עוגן משותף אוניברסלי + גדיל מתאם המכיל רצף דומה אך עם החלפת ה- CPGs עם GPCs (CMO-GpC). גירוי TLR9 יגרום לייצור SEAP (פוספטאז אלקליין עוברי מופרש). הפרשת SEAP כותמה על ידי מבחנת צבע (ספיגה). תנאי הטיפול הושוו לתאי מנוחה שטופלו רק עם PBS. דגירה עם CMO-GPC לא לעורר ביטוי TLR9. דגירה עם CMO-CpG היה מעט גבוה יותר מאשר תאים נחים אבל הרבה יותר נמוך מאשר ODN-1826. נא לחץ כאן כדי להוריד קובץ זה.
איור 6 משלים: הכדאיות של תאים לאחר תהליך תיוג CMO. כדי להעריך כיצד הפרוטוקול משפיע על הכדאיות, HUVECs חולקו לארבע אוכלוסיות: אחת נשארה על קרח במשך שעה אחת, אחת סומנה בדומה עם PBS אך אחרת נלקחה דרך כל שלבי הצנטריפוגות והשטיפה, אחת סומנה ב- CMOs, ואחת סומנה ב- CMOs ומסוננת באמצעות מסנן של 40 מיקרומטר כדי להסיר גושים. התאים היו מוכתמים אז עם calcein AM ואתידיום homodimer כדי להעריך את מספר התאים החיים והמתים. כל הטיפולים הביאו לירידה משמעותית בכדאיות מאשר בקרת הקרח (ANOVA בכיוון אחד עם ניתוח טוקי לאחר הוק), אך הכדאיות החציונית לתיוג CMO (עם או בלי סינון) הייתה כ -94%. נתונים שנאספו משלושה ניסויים עצמאיים. * = p < 0.05. = p < 0.0001 אנא לחץ כאן כדי להוריד קובץ זה.
| תוצאה | סיבות אפשריות | תיקונים מוצעים |
| פוטוליתוגרפיה – התכונות סדוקות | אפייה רכה לא עקבית או לא מספקת | להגדיל את הזמן של אפייה רכה עד 3 דקות; לאמת את הטמפרטורה בפועל של כיריים ולהעלות את הטמפרטורה לפי הצורך |
| פוטוליתוגרפיה – התכונות אינן חדות או שנשארו בתוכן פוטורזיסט | תת-פיתוח | הגדל את הזמן שהשקופית מבלה בפתרון מפתחים; שילוב של עצבנות עדינה |
| פוטוליתוגרפיה – תכונות שאינן עקביות בשקופית | ייתכן שאור UV אינו מרוכז או לא ממוקד כראוי | התאם את הגדרת תאורת UV כדי להבטיח אור קולימי בעוצמה אחידה |
| תאים אינם נצמדים לכתמים בדוגמת דפוס עם יעילות גבוהה | אין מספיק דנ"א על פני השטח | אשר כי DNA קיים על פני השטח על ידי הכלאת השקופית עם אוליגוס משלימים פלואורסצנטי ולאחר מכן הדמיה תחת מיקרוסקופ |
| תאים מסומנים כראוי עם CMO | הוסף אוליגוס משלימים פלואורסצנטיים להשעיית תאים ולאשר פלואורסצנטיות באמצעות ציטומטריית זרימה | |
| אין מספיק תאים על-פני תבנית | איסוף תאים על-ידי שטיפה מתא זרימת PDMS, צנטריפוגה והשהייתם מחדש בנפח נמוך יותר כדי לרכז את התאים | |
| יותר מדי CMO שנותר בהשעיית תאים, הכלאה עם DNA בשקופית | הוסף שלב כביסה נוסף. הקפד להסיר כמה שיותר supernatant ככל האפשר עם כל לשטוף. | |
| יותר מדי הפנמה של CMO עקב זמן וטמפרטורה | לעבוד במהירות לאחר תיוג התאים עם CMO; לשמור על תאים ולהחליק על קרח ולהשתמש ריאגנטים קרים כקרח | |
| תאים מתגושמות | התאים לא הופרדו כראוי במהלך טריפסיניזציה | השתמש PBS + 0.04% EDTA במהלך שטיפת תאים; לעבור השעיית תא דרך מסנן 35 מיקרומטר לפני הכביסה הסופית |
| תאים דבקים באופן לא ספציפי | אם באזור מסוים אחד – יכול להיות בגלל שריטות בשקופית, אי התאמה של תאי זרימת PDMS, או שפיכה של DNA מחוץ לאזור התבנית | הימנע משריטות, הקפד ליישר את תאי זרימת PDMS לאזור דוגמת המילוי |
| אם התאים דבקים בכל מקום – חסימה או כביסה לקויים | להוסיף שטיפות נוספות לאחר דפוס התאים; צינור במרץ רב יותר במהלך שטיפות; לחסום עם 1% BSA למשך זמן רב יותר לפני תחילת תבנית התא; שקופית סילאניז (שלב 3 אופציונלי) או אישור שסילאניזציה הצליחה על ידי מדידת זווית המגע של טיפת מים | |
| נוצרות בועות בתוך תא זרימה | שגיאות צנרת, משטח הידרופילי לא אחיד שנוצר במהלך חמצון פלזמה | אם הבועות קטנות, הוסף PBS ל- Inlet של תא הזרימה וייתכן שהן נשטפו החוצה. אם הבועות גדולות יותר, יש להפעיל לחץ עדין על תא הזרימה PDMS, תוך הסטת הבועות לכיוון המפרצון או השקע. |
| תאים בתחילה לדבוק דפוס אבל מוסרים במהלך שטיפות, דפוס של סוגי תאים אחרים, או הוספת מבשר הידרוג'ל | כוחות הגימור מצינורות במרץ רב מדי יכולים לגרום לתאים להתנתק מפני השטח | יש להזרים בעדינות רבה יותר במהלך השטיפות הבאות, סבבים של דפוס תאים או הוספת מבשרי הידרוג'ל. מכיוון שמבשרי ההידרוג'ל הם צמיגים, הם נוטים יותר לגרום לתבנית להתנתק, אז היזהרו במיוחד. מבנים רב שכבתיים נוטים להיות כבדים ביותר והם רגישים יותר להיות מופרים. |
| עיוותי רקמות במהלך העברה תלת-ממדית | הידרוג'ל מקלות להחליק | אישור הידרופוביה של שקופית באמצעות מדידות זווית מגע |
| השתמש בסכין גילוח כדי להרים PDMS במלואו בשני הקצוות, ומאפשר PBS לצוף מתחת לרקמה | ||
| זה יכול לקרות עם הידרוג'ל קולגן טהור – שקול להתאים את ריכוז החלבון או הרכב ההידרוג'ל | ||
| תאים לא עוברים עם ההידרוג'ל ונשארים בשקופית | להגביר את ריכוז ה-DNAse של טורבו או להגדיל את זמן הדגירה | |
| הידרוג'ל אינו מוצק מספיק | להגדיל את זמן הדגירה ו/או את מנגנון הג'ל עבור הידרוג'ל המדובר (למשל קולגן, לוודא pH נכון) | |
| דמעות הידרוג'ל עם הסרת PDMS | הפוך PDMS לזרום תאים הידרופילי באמצעות חמצון פלזמה לפני תחילת הניסוי, כך שהם להתנתק בקלות עם הוספת מדיה. השתמש במלקחיים בעדינות רבה כדי לנתק את ה- PDMS. |
טבלה 1: מדריך לפתרון בעיות כדי לזהות ולפתור כשלים פוטנציאליים שעלולים לנבוע מפרוטוקול זה. בפרט, הידבקות לקויה של תאים לתבנית יכולה להיות סיבות שורש רבות ומדריך זה צריך לעזור עם זיהוי ופתרון של בעיות אלה.
קובץ משלים 1. נא לחץ כאן כדי להוריד קובץ זה.
קובץ משלים 2. נא לחץ כאן כדי להוריד קובץ זה.
קובץ משלים 3. נא לחץ כאן כדי להוריד קובץ זה.
קובץ משלים 4. נא לחץ כאן כדי להוריד קובץ זה.
במאמר זה, אנו מציגים פרוטוקול מפורט לתבנית ברזולוציה גבוהה של תאים ב- 2D ותלת-ממד לניסויים בתרבית תאי במבחנה. שלא כמו גרסאות שפורסמו בעבר של שיטה זו, הפרוטוקול המוצג כאן מתמקד שימושיות: זה לא דורש ציוד מיוחד מאוד ואת כל ריאגנטים ניתן לרכוש מספקים במקום לדרוש סינתזה מותאמת אישית. שלא כמו שיטות אחרות של מיקרו-מפטרציה של תאים, שיטה זו מהירה ואגנוסטית מסוג תא: היא אינה דורשת הידבקות ספציפית לחלבוני מטריצה חוץ-תאיים15. תאים בדוגמת CMO-DPAC יכולים להיות מוטבעים בתוך מטריצה חוץ תאית כגון מטריגל או קולגן, וכתוצאה מכך תרביות תלת-ממד עם רזולוציה מרחבית גבוהה בהרבה ממה שניתן כיום בשיטות מבוססות הדפסה שחול22. ניתן להשתמש ב- CMO-DPAC כדי ליצור מאות עד אלפי תכונות מיקרוסקופיות לכל שקופית, ומאפשר לבצע שכפולים רבים בו-זמנית.
אחד הפרמטרים החשובים ביותר בהצלחת פרוטוקול זה הוא צפיפות התאים שנוספו לתאי הזרימה מעל השקופית בדוגמת דפוס. באופן אידיאלי, הצפיפות צריכה להיות לפחות 25 מיליון תאים / מ"ל. כאשר נטען לתאי הזרימה, צפיפות זו של תאים גורמת לשכבת תאים כמעט צפופה מעל התבנית (איור 2B משלים). צפיפות תאים גבוהה אלה ממקסמות את ההסתברות שתא יתיישב ישירות על גבי כתם DNA וידבק. הפחתת צפיפות התאים תקטין את יעילות התבנית הכוללת. צעד קריטי נוסף בפרוטוקול זה הוא השעיה מחדש של התאים ב-PBS או במדיה נטולת סרום לפני הוספת פתרון CMO. מחיצת CMOs במהירות רבה לתוך קרום התא והוספת פתרון CMO ישירות לכדור תא תגרום תיוג הטרוגניים של תאים. לאחר הוספת פתרון CMO להשעיית התא, חשוב לערבב ביסודיות על ידי pipetting, כך התאים מסומנים באופן אחיד עם CMOs. לאחר הדגירה, יש צורך לשטוף ביסודיות את CMOs עודף באמצעות צנטריפוגה מרובים ולשטוף צעדים. עודף CMO חינם נוכח ההשעיה התא יהיה לאגד את ה- DNA בדוגמת אמין שונה על שקופית הזכוכית, חסימת הכלאה והידבקות של תאים מהונדסים CMO בהשעיה. זמן הוא גם שיקול מרכזי עבור פרוטוקול זה. חשוב לעבוד מהר ככל האפשר בעת שימוש ב- CMOs ולשמור על התאים על הקרח על מנת למזער את ההפנמה של CMOs ולמקסם את הכדאיות התא. ניסויי ציטומטריית זרימה הראו כי CMOs אינם נמשכים זמן רב על פני התא כמו LMOs, עם אובדן של 25% של מתחמי CMO במשך שעתיים של דגירה על קרח36. יתר על כן, הכדאיות של תאים תקטן ככל שזמן הטיפול בתאים יגדל. ניתן למקסם את הכדאיות על ידי עבודה מהירה, שמירה על תאים על קרח, שימוש בריאגנטים קרים כקרח ושימוש במדיה ללא סרום כדי לספק חומרים מזינים מסוימים.
למרות CMO-DPAC יכול להיות דרך רבת עוצמה של לימוד ביולוגיה של התא על ידי דפוס תאים עם דיוק גבוה, יש לו את המגבלות שלה. ניסויי CMO-DPAC יכולים להיות מאתגרים, במיוחד כאשר המורכבות הניסיונית מתווספת עם סוגי תאים מרובים, שכבות או תרבית תאיםתלת-ממדית ( קובץ משלים 1). כשלים ניסיוניים יכולים להיות נפוצים בעת הפעלת פרוטוקול זה, כמתואר בטבלה 1. לכן, אנו ממליצים למשתמשים להנהיג בדיקות בקרת איכות (המאשרות כי ה- DNA קיים בשקופית, המאשרות כי תאים מסומנים מספיק עם DNA (שלב 8), המאשר כי תאים עודפים נשטפו ביסודיות וכו ') כדי לוודא שהניסוי מצליח ולזהות שלבים שעשויים לדרוש אופטימיזציה נוספת. אנו מקווים כי המידע המסופק בכתב יד זה ובקבצים המשלימים שלו יסייע להקל על כל פתרון בעיות נדרש.
כולסטרול הוא מולקולה ביואקטיבית שהפנמה שלה עשויה להשפיע על חילוף החומרים של התא, ביטוי גנים ונזילות ממברנה37,38. מחקר קודם השווה את ההשפעות על ביטוי גנים של תאים בעלי תווית CMO ו- LMO באמצעות רצף RNA של תאים בודדים. תאי HEK המסומנים בתווית CMO שינו את ביטוי הגנים בהשוואה לתאים לא מתויגים ומסומנים ב-LMO36. תיוג תאים עם CMOs הביא ביטוי דיפרנציאלי (> פי 1.5) של שמונה גנים ביחס לבקרות ללא תווית, כולל AP2B1, אשר נקשר כולסטרול ותחבורה sphingolipid (GeneCards), ו MALAT1, RNA ארוך שאינו קידוד המווסת הצטברותכולסטרול 39. בעוד מינורי, תגובות שעתוק אלה עשויות בכל זאת להיות דאגה אם הניסוי המדובר הוא לימוד חילוף החומרים, דינמיקה ממברנה, או מסלולים אחרים הקשורים כולסטרול בתאים.
פרוטוקול זה גמיש וניתן לכוונן אותו כך לענות על הצרכים של כל ניסוי. מכיוון שה- CMO מכניס את עצמו לתוך קרום השומנים במקום להשתמש בכל קולטן ספציפי, השיטה היא אגנוסטית מסוג התא (HUVECs, MCF10As, HEKs ו- MDCKs הוכחו כאן). למרות כולסטרול הוא עוגן הידרופובי שונה מאשר LMOs שלנו שפורסם בעבר, עד כה מצאנו אותם להתנהג באופן דומה. לכן, היינו מצפים CMOs לעבוד עם כל מגוון רחב של סוגי תאים שפרסמנו בעבר עם LMOs, כולל אך לא מוגבל לתאי גזע עצביים, פיברובלסטים, תאים מונונוקלאריים דם היקפי, תאים סרטניים, ותאי אפיתל הממכרהעיקריים 6,23,27,29,36 . תיוג CMO אינו מגרה את TLR9, דבר המצביע על כך שהפרוטוקול תואם לתאי מערכת החיסון. שילוב ממברנה של CMO הוא פונקציה של גודל התא הכולל ואת מידת המטען השלילי בגליקוקליקס התא35. לכן, כללנו פרוטוקול (שלב 8) לבדיקת היקף שילוב הממברנה כי הוא מקובל אופטימיזציה מהירה. התכונות הספציפיות של כל תבנית תא ישתנו באופן בלתי נמנע בהתאם לעיצוב הניסיוני (ראה קובץ משלים 1 לקבלת הדרכה נוספת). למרות פרוטוקול photopatterning המתואר לעיל עבור דפוס ה- DNA מומלץ, כל שיטה של confining מרחבי טיפות של פתרון אמין-DNA צריך לעבוד, כגון השימוש במדפסות טיפות ברזולוציה גבוהה. רזולוציית התבניות ומרווח התכונות המינימלי ישתנו בהתאם לפעולת השירות שבה נעשה שימוש. תיאורטית ניתן גם לשלב את קטעי ה-DNA-photopatterning של פרוטוקול זה עם שיטות אחרות ששימשו לתיוג תאים עם DNA, כגון עם DNA היברידי לאצבעות אבץ מבוטאותממברנה 40, באמצעות DNA מצומד NHS41, ותגובה שאריות חומצה סיאלית אזידו על פני התא עם DNA פוספין מצומד42 . ניתן להחיל CMO-DPAC על מגוון ניסויים הדורשים שליטה הדוקה על המרווח בין תאים, כולל מחקרים על האינטראקציות בין זוגות תאים, ניסויים בתרבית משותפת הבוחנים העברת אותות מתאי "השולח" לתאי "מקלט", וחקירות של ההשפעה של רמזים חוץ-תאיים סמוכים על בידול תאי גזע6,29 . השיטה יכולה לשמש גם ליצירת microtissues שניתן להשתמש בהם כדי לחקור את העברת התאים בשלושה ממדים, את הארגון העצמי של תאים לתוךרקמות 23,27, ואת יחסי הגומלין הדינמיים בין תאים ו- ECM27. אנו מקווים כי פרוטוקול זה יספק לחוקרים פלטפורמה נגישה לחקור יישומים חדשים של דפוס תאים מבוססי DNA ברזולוציה גבוהה במעבדות שלהם.
ז.ג.ג. הוא יועץ ובעל מניות ב-Provenance Biosciences.
המחברים רוצים להודות לג'רמי גרסיה על שבחן פרוטוקול זה ובושאן חרקיקאר על מתן הכשרה על הציוד בליבת המיקרו והננוטכנולוגיה הביו-רפואית של UCSF. מחקר זה נתמך בחלקו על ידי מענקים מהתוכנית לחקר סרטן השד של משרד ההגנה (W81XWH-10-1-1023 ו- W81XWH-13-1-0221), NIH (U01CA199315, DP2 HD080351-01, 1R01CA190843-01, 1R21EB019181-01A, ו- 1R21CA182375-01A1), NSF (MCB1330864) ומרכז UCSF לבנייה סלולרית (DBI-1548297), מרכז מדע וטכנולוגיה NSF. O.J.S מומן על ידי מלגת מחקר בוגר NSF, מלגת סייבל, ומלגת P.E.O. זי.ג'יי.ג'י וא.ר.א הם חוקרי ביו-הוב של צ'אן-צוקרברג.
| Name | Company | Catalog Number | Comments |
| 2-well Chambered Coverglass w/ non-removable wells | Thermo Fisher Scientific | 155379 | |
| Acetic Acid | Sigma-Aldrich | A6283 | |
| Adapter with External SM1 Threads and Internal SM3 Thread | ThorLabs | SM3A1 | |
| Aldehyde Functionalized Slides | Schott | Nexterion Slide AL | Store under dry conditions after opening. |
| All Plastic Syringes, 1 mL | Fisher Scientific | 14-817-25 | |
| Amine-Modified DNA Oligo | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| Aspheric Condenser Lens | ThorLabs | ACL7560 | |
| Borosilicate Disc, 6in Diameter X 1/2in Thick | Chemglass | CG-1906-23 | |
| Cell Culture Dishes 60x15 mm style | Corning | 353002 | |
| Cholesterol-Modified Oligo | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| Diamond Scribe | Excelta | 475B | |
| DNA Oligonucleotide | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| DPBS, no calcium, no magnesium | Thermo Fisher Scientific | 14190250 | |
| Isopropyl Alcohol | Sigma-Aldrich | 278475 | |
| Matrigel Matrix, Growth Factor Reduced | Corning | 354230 | |
| Methylene Chloride (Stabilized/Certified ACS) | Fisher Scientific | D37-4 | |
| MF-321 Developer | Kayaku Advanced Materials | n/a | |
| Microposit S1813 Positive Photoresist | Kayaku Advanced Materials | n/a | |
| Ø3" Adjustable Lens Tube, 0.81" Travel | ThorLabs | SM3V10 | |
| Oven | Thermo Scientific | 51-028-112H | |
| PE-50 Compact Benchtop Plasma Cleaning System | Plasma Etch | PE-50 | |
| Photomask (custom) | CAD/Art Services | n/a | Minimum feature size guaranteed by CAD/Art Services is 10 microns. |
| Razor Blades | Fisher Scientific | 12-640 | |
| RCT Basic Hot Plate | IKA | 3810001 | |
| Silicon Wafer (100 mm) | University Wafer | 590 | |
| Sodium Borohydride, 98%, granules | Acros Organics | 419471000 | |
| Spin Coater Kit | Instras | SCK-200 | This is a low cost option, but any spin coater that can maintain a speed of 3000 rpm will suffice. |
| SU-8 2075 | Microchem | Y111074 0500L1GL | |
| SU-8 Developer | Microchem | Y020100 4000L1PE | |
| Sylgard 184 Silicone Elastomer Kit | Dow | 2646340 | |
| Syringe Needles | Sigma-Aldrich | Z192341 | |
| T-Cube LED Driver, 1200 mA Max Drive Current | ThorLabs | LEDD1B | |
| Tridecafluoro-1,1,2,2-tetrahydrooctyl dimethylchlorosilane | Gelest | SIT8170.0 | |
| Triethylamine | Sigma-Aldrich | 90335 | |
| Turbo DNase | Thermo Fisher Scientific | AM2238 | |
| Tweezers Style N7 | VWR | 100488-324 | The curved shape of these tweezers is essential for delicately picking up the PDMS flow cells containing patterned tissues. |
| UV LED (365 nm, 190 mW (Min) Mounted LED, 700 mA) | ThorLabs | M365L2 | |
| Wafer Tweezers | Agar Scientific | T5063 | |
| WHEATON Dry-Seal vacuum desiccator | Millipore Sigma | W365885 |
- Kreeger, P. K., Strong, L. E., Masters, K. S. Engineering approaches to study cellular decision-making. Annual Review of Biomedical Engineering. , 49-72 (2018).
- Goubko, C. a., Cao, X. Patterning multiple cell types in co-cultures: A review. Materials Science and Engineering C. 29 (6), 1855 (2009).
- Sun, W., et al. The bioprinting roadmap. Biofabrication. 12 (2), 022002 (2020).
- Liu, W. F., Chen, C. S. Cellular and multicellular form and function. Advanced Drug Delivery Reviews. 59 (13), 1319-1328 (2007).
- Duffy, R. M., Sun, Y., Feinberg, A. W. Understanding the role of ECM protein composition and geometric micropatterning for engineering human skeletal muscle. Annals of Biomedical Engineering. 44 (6), 2076-2089 (2016).
- Chen, S., et al. Interrogating cellular fate decisions with high-throughput arrays of multiplexed cellular communities. Nature Communications. 7, 10309 (2016).
- Shaya, O., et al. Cell-cell contact area affects notch signaling and notch-dependent patterning. Developmental Cell. 40 (5), 505-511 (2017).
- Rao, N., et al. A co-culture device with a tunable stiffness to understand combinatorial cell-cell and cell-matrix interactions. Integrative Biology. 5 (11), 1344 (2013).
- Sriraghavan, V., Desai, R. A., Kwon, Y., Mrksich, M., Chen, C. S. Micropatterned dynamically adhesive substrates for cell migration. Langmuir. 26 (22), 17733-17738 (2010).
- Wong, L., Pegan, J. D., Gabela-Zuniga, B., Khine, M., McCloskey, K. E. Leaf-inspired microcontact printing vascular patterns. Biofabrication. 9 (2), 021001 (2017).
- Chen, T. H., et al. Directing tissue morphogenesis via self-assembly of vascular mesenchymal cells. Biomaterials. 33 (35), 9019-9026 (2012).
- Laurent, J., et al. Convergence of microengineering and cellular self-organization towards functional tissue manufacturing. Nature Biomedical Engineering. 1 (12), 939-956 (2017).
- Lin, C., Khetani, S. R. Micropatterned co-cultures of human hepatocytes and stromal cells for the assessment of drug clearance and drug-drug interactions. Current Protocols in Toxicology. 2017, 1-23 (2017).
- Hui, E. E., Bhatia, S. N. Micromechanical control of cell-cell interactions. Proceedings of the National Academy of Sciences of the United States of America. 104 (14), 5722-5726 (2007).
- D'Arcangelo, E., McGuigan, A. P. Micropatterning strategies to engineer controlled cell and tissue architecture in vitro. BioTechniques. 58 (1), 13-23 (2015).
- Martinez-Rivas, A., González-Quijano, G. K., Proa-Coronado, S., Séverac, C., Dague, E. Methods of micropatterning and manipulation of cells for biomedical applications. Micromachines. 8 (12), (2017).
- Lee, S., et al. Simple lithography-free single cell micropatterning using laser-cut stencils. Journal of Visualized Experiments. (158), e60888 (2020).
- Strale, P. O., et al. Multiprotein printing by light-induced molecular adsorption. Advanced Materials. 28 (10), 2024-2029 (2016).
- Melero, C., et al. Light-induced molecular adsorption of proteins using the primo system for micro-patterning to study cell responses to extracellular matrix proteins. Journal of Visualized Experiments. (152), e60092 (2019).
- Reid, J. A., Mollica, P. M., Bruno, R. D., Sachs, P. C. Consistent and reproducible cultures of large-scale 3D mammary epithelial structures using an accessible bioprinting platform. Breast Cancer Research. , 1-13 (2018).
- Wang, Z., Lee, S. J., Cheng, H. -. J., Yoo, J. J., Atala, A. 3D bioprinted functional and contractile cardiac tissue constructs. Acta Biomaterialia. 70, 48-56 (2018).
- Miri, A. K., et al. Effective bioprinting resolution in tissue model fabrication. Lab on a Chip. 19 (11), 2019-2037 (2019).
- Todhunter, M. E., et al. Programmed synthesis of three-dimensional tissues. Nature Methods. 12 (10), 975-981 (2015).
- Todhunter, M. E., Weber, R. J., Farlow, J., Jee, N. Y., Gartner, Z. J. Fabrication of 3D microtissue arrays by DNA programmed assembly of cells. Current Protocols in Chemical Biology. 8 (3), 147-178 (2016).
- Csizmar, C. M., Petersburg, J. R., Wagner, C. R. Programming cell-cell interactions through non-genetic membrane engineering. Cell Chemical Biology. 25 (8), 931-940 (2018).
- Weber, R. J., Liang, S. I., Selden, N. S., Desai, T. A., Gartner, Z. J. Efficient targeting of fatty-acid modified oligonucleotides to live cell membranes through stepwise assembly. Biomacromolecules. 15 (12), 4621-4626 (2014).
- Hughes, A. J., et al. Engineered tissue folding by mechanical compaction of the mesenchyme. Developmental Cell. 44 (2), 165-178 (2018).
- Weber, R. J., et al. Rapid organoid reconstitution by chemical micromolding. ACS Biomaterials Science & Engineering. 2 (11), 1851-1855 (2016).
- Scheideler, O. J., et al. Recapitulating complex biological signaling environments using a multiplexed, DNA-patterning approach. Science Advances. 6 (12), (2020).
- Viola, J. M., et al. Guiding cell network assembly using shape-morphing hydrogels. Advanced materials (Deerfield Beach, Fla.). , 2002195 (2020).
- Mohammad, A., Davis, M., Aprelev, A., Ferrone, F. A. Note: Professional grade microfluidics fabricated simply. Review of Scientific Instruments. 87 (10), 1-4 (2016).
- Lee, O. J., Chuah, H. S., Umar, R., Chen, S. K., Yusra, A. F. I. Construction of cost effective homebuilt spin coater for coating amylose-amylopectin thin films. Journal of Fundamental and Applied Sciences. 9 (2), 279 (2018).
- Webb, K., Hlady, V., Tresco, P. A. Relative importance of surface wettability and charged functional groups on NIH 3T3 fibroblast attachment, spreading, and cytoskeletal organization. Journal of Biomedical Materials Research. 41 (3), 422-430 (1998).
- Processing Guidelines for: SU-8 2025, SU-8 2035, SU-8 2050, SU-8 2075. Microchem SU-8 2000 Permanent Expoxy Negative Photoresist Available from: https://kayakuam.com/wp-content/uploads/2019/09/SU-82000DataSheet2025thru2075Ver4.pdf (2019)
- Palte, M. J., Raines, R. T. Interaction of nucleic acids with the glycocalyx. Journal of the American Chemical Society. 134 (14), 6218-6223 (2012).
- McGinnis, C. S., et al. MULTI-seq: sample multiplexing for single-cell RNA sequencing using lipid-tagged indices. Nature Methods. 16 (7), 619-626 (2019).
- Maxfield, F. R., van Meer, G. Cholesterol, the central lipid of mammalian cells. Current Opinion in Cell Biology. 22 (4), 422-429 (2010).
- Luo, J., Yang, H., Song, B. L. Mechanisms and regulation of cholesterol homeostasis. Nature Reviews Molecular Cell Biology. 21 (4), 225-245 (2020).
- Liu, L., Tan, L., Yao, J., Yang, L. Long non-coding RNA MALAT1 regulates cholesterol accumulation in ox-LDL-induced macrophages via the microRNA-17-5p/ABCA1 axis. Molecular Medicine Reports. 21 (4), 1761-1770 (2020).
- Mali, P., Aach, J., Lee, J. H., Levner, D., Nip, L., Church, G. M. Barcoding cells using cell-surface programmable DNA-binding domains. Nature Methods. 10 (5), 403-406 (2013).
- Hsiao, S. C., et al. Direct cell surface modification with DNA for the capture of primary cells and the investigation of myotube formation on defined patterns. Langmuir. 25 (12), 6985-6991 (2009).
- Gartner, Z. J., Bertozzi, C. R. Programmed assembly of 3-dimensional microtissues with defined cellular conductivity. Proceedings of the National Academy of Sciences. (17), 1-5 (2009).
Tags
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved