
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Bioengineering
Простое, доступное и модульное структурирование клеток с использованием ДНК
Здесь мы представляем протокол к микропаттерновым клеткам с одноклеточным разрешением с использованием днк-запрограммированной адгезии. Этот протокол использует настольная фотолитографическую платформу для создания паттернов олигонуклеотидов ДНК на стеклянном слайде, а затем маркирует клеточные мембраны коммерчески доступными комплементарными олигонуклеотидами. Гибридизация олиго приводит к запрограммированной клеточной адгезии.
Относительное расположение клеток является ключевой особенностью микросреды, организующей клеточно-клеточные взаимодействия. Для изучения взаимодействий между клетками одного или другого типа оказались полезными методы микроструктурирования. ДНК-запрограммированная сборка клеток (DPAC) - это метод микроструктурирования, который нацелен на адгезию клеток к субстрату или другим клеткам с использованием гибридизации ДНК. Самые основные операции в DPAC начинаются с украшения клеточных мембран липид-модифицированными олигонуклеотидами, а затем протекают ими по субстрату, который был узорчат с комплементарными последовательностями ДНК. Клетки избирательно прилипают к субстрату только там, где они находят комплементарную последовательность ДНК. Неприлипанные клетки смываются, выявляя рисунок адгезивных клеток. Дополнительные операции включают дальнейшие раунды клеточного субстрата или клеточно-клеточной адгезии, а также перенос паттернов, образованных DPAC, во встраиваемый гидрогель для долгосрочной культуры. Ранее методы моделирования олигонуклеотидов на поверхностях и украшения клеток последовательностями ДНК требовали специализированного оборудования и пользовательского синтеза ДНК соответственно. Мы сообщаем об обновленной версии протокола, использующей недорогую настольной фотолитографическую установку и коммерчески доступные модифицированные холестерином олигонуклеотиды (ОКУ), развернутые с использованием модульного формата. Клетки, меченые CMO, с высокой эффективностью прилипают к субстрастрам с ДНК-паттернами. Этот подход может быть использован для создания нескольких типов клеток одновременно с высокой точностью и для создания массивов микротизюй, встроенных во внеклеточный матрикс. Преимущества этого метода включают его высокое разрешение, способность встраивать клетки в трехмерную микросреду без нарушения микроструктуры и гибкость в паттерне любого типа клеток.
Расположение клеток относительно друг друга в ткани является важной особенностью микросреды1,2,3,4. Методы, используемые для структурирования живых клеток в пространственно контролируемые расположения, являются ценными экспериментальными инструментами дляизучения дифференцировки4,5,6,7,8,подвижности клеток9,морфогенеза10, 11, 12,метаболизма13и клеточно-клеточных взаимодействий7,14 . Существует множество методов паттернирования клеток, каждый со своими преимуществами и недостатками3,4. Методы, которые создают адгезивные островки из белков внеклеточного матрикса (ECM), такие как микроконтактная печать и лазерная резка трафаретов, просты и масштабируемы. Тем не менее, трудно создать паттерн более одного или двух типов клеток одновременно, потому что адгезивные свойства разных типов клеток к различным молекулам ECM часто похожи15,16,17. Более сложные микроструктуры могут быть созданы с помощью световой молекулярной адсорбции (LIMAP), метода, который использует ультрафиолетовый свет для абляции областей, покрытых ПЭГ, и допускает последующую адсорбцию белка18,19. Этот процесс может быть повторен для создания микроструктур с высоким разрешением с несколькими типами клеток. Однако может произойти перекрестное связывание клеток с различными белковыми пятнами, что приводит к плохой специфичности паттерна19. Физические методы, такие как посев клеток на микромеханические реконфигурируемые устройства культивирования, могут создавать структурированные кокультуры с динамическим управлением, но без гибкости в дизайне шаблонов микроконтактной печати или LIMAP14,8. В отличие от других методов, биопечать может создавать трехмерные расположения клеток внутри гидрогелей20,21. Тем не менее, биопечатные конструкции имеют гораздо более низкое разрешение, чем другие методы микроструктурирования, со средним размером признака порядка сотен микрон22. Идеальный метод клеточного паттерна должен иметь высокое разрешение, шаблонировать несколько типов клеток, использовать оборудование и реагенты, которые легко доступны, и иметь возможность встраивать успешные паттерны в гидрогель для трехмерной (3D) клеточной культуры. В этой статье мы представляем CMO-DPAC, метод микроструктурирования клеток, который использует гибкость и скорость гибридизации ДНК для нацеливания клеточной адгезии к субстрату. Этот метод был адаптирован из наших предыдущих протоколов23,24, чтобы сделать его более доступным, модульным и доступным. Используя текущий протокол, любая лаборатория должна иметь возможность настроить полностью функциональную систему без какого-либо специализированного оборудования или опыта.
ЗАПРОГРАММИРОВАННАЯ сборка клеток ДНК (DPAC) - это мощная техника тканевой инженерии, которая моделирует клетки с одноклеточным разрешением с точным контролем расстояния между клетками и геометрией тканей. В DPAC клеточные мембраны украшены олигонуклеотидами ДНК (олиго) с использованием двух липид-модифицированных олиго, предназначенных для гибридизации на клеточной мембране. Поскольку олиго конъюгированы с гидрофобными липидами, они быстро переходят на клеточную мембрану25, где они гибридизуются, увеличивая чистую гидрофобность нековалентно связанных молекул и тем самым увеличивая их время жизни на поверхности клетки26. Олиго представлены на поверхности клетки таким образом, что они могут гибридизоваться с комплементарными олиго на других клетках или ДНК-функционализированными стеклянными слайдами для создания определенных 2D или 3D клеточных паттернов с предписанным составом, расстоянием между клетками и геометрией23,24. Узорчатые микротизры могут быть отщеплены от поверхности ферментаматически и встроены в гидрогель для длительной 3D-культуры. При использовании в сочетании с первичными клетками или стволовыми клетками полученные коллекции клеток могут подвергаться морфогенезу и формироваться в органоиды23,27,28. DPAC был применен для исследования динамики судьбы взрослых нервных стволовых клеток в ответ на конкурирующие сигналы6,29,для изучения самоорганизации эпителиальных клеток молочной железы23, 28и для генерации «тканевого оригами» посредством мезенхимальной конденсации27.
DPAC позволяет точно размещать несколько клеточных популяций и имеет значительно лучшее разрешение, чем биопринтеры на основе экструзии (порядка микрон)22,23. Кроме того, в отличие от методов моделирования на основе ECM, таких как микроконтактная печать, DPAC не требует дифференциальной адгезии различных типов клеток к поверхности с покрытием ECM15,23. Он идеально подходит для ответа на вопросы о том, как состав ткани влияет на ее поведение, как клетки интегрируют несколько клеточных и микроокружающих сигналов при принятиирешений 6,29и как пары клеток взаимодействуют друг с другом. Преимущество этого метода перед другими методами микроструктурирования заключается в том, что его можно использовать для 3D-культуры клеток в одной плоскости визуализации, облегчая покадровые исследования самоорганизации тканей и органоидного морфогенеза23,27,30.
Несмотря на эти преимущества, успешное внедрение DPAC потребовало синтеза пользовательских олигонуклеотидных реагентов и доступа к специализированному оборудованию для паттернов ДНК23,24,что ограничивает широкое распространение. Например, оптимальные липид-модифицированные олиго (ЖИО), используемые в исходном протоколе, должны быть специально синтезированы, модифицированы лигноцериновой кислотой или пальмитиновой кислотой и очищены26. Этот процесс требует использования синтезатора ДНК и высокоэффективного инструмента жидкостной хроматографии, а также покупки связанных реагентов, таких как метиламин, контролируемое вещество, которое подпадает как под институциональные, так и федеральные правила. В качестве альтернативы ЖИО могут быть приобретены на заказ оптом, но это требует значительных авансовых инвестиций в технологию.
Чтобы преодолеть эти ограничения, мы разработали пересмотренную версию DPAC, которая использует коммерчески доступные модифицированные холестерином олиго (ОКУ) вместо специально синтезированных ЖИО. Чтобы еще больше снизить затраты и повысить гибкость платформы, мы перешли на модульную трехолигосистему. Вместо того, чтобы заказывать новое модифицированное холестерином олиго для каждой уникальной клеточной популяции, пользователь этого протокола может вместо этого использовать одни и те же модифицированные холестерином олиго («Универсальный якорь» и «Универсальный ко-якорь») для каждой клеточной популяции, а затем использовать недорогое, немодифицированное олиго («Adapter Strand»), которое гибридизуется как с универсальным якорем, так и либо с амино-функционализированной ДНК на поверхности, либо с переходной нитью другого типа клеток.
Другим ограничением оригинального протокола DPAC было то, что он создавал слайды с днк-паттернами с помощью жидкого принтера с высоким разрешением (например, Nano eNabler, BioForce Nanosciences)23,24. Хотя этот прибор может похвастаться исключительным разрешением и низкими требованиями к реагентам, он недоступен для большинства учреждений и имеет относительно низкую скорость печати (примерно 1 функция с рисунком в секунду). Недавно были разработаны два фотолитографических метода для моделирования особенностей ДНК на поверхностях. Виола и его коллеги использовали полиакриламидное и бензофенонное покрытие, которое ковалентно связывает одноцепочечные олиго ДНК при воздействии ультрафиолетового света30. Используя этот метод, они смогли создать тканевые каркасы, которые претерпели крупномасштабные, запрограммированные изменения формы в результате сократимости клеток и самоорганизации. Scheideler et al. разработали метод, который использует УФ-экспозицию положительного фоторезиста для селективного воздействия амино-модифицированных олиго ДНК на альдегидно-функционализированный слайд29. После выпечки и восстановительного аминирования амино-модифицированная ДНК ковалентно связывается с поверхностью. Этот метод был использован для исследования реакции взрослых нервных стволовых клеток на пространственно представленные сигналы самообновления и дифференцировки. Эта статья адаптирует протокол Scheideler et al. для создания паттернов ДНК, которые будут захватывать клетки, меченые CMO. Этот протокол фотопаттерна может быть выполнен без использования чистой комнаты. Он использует недорогое и коммерчески доступное оборудование, которое легко развертывается на столешнице или вытяжном капоте. Использование недорогого или DIY (сделай сам) фотолитографического оборудования повышает доступность для исследователей без доступа к объектам чистых помещений и позволяет исследователям попробовать технику без больших вложений времени или ресурсов31,32. Тем не менее, лучшее разрешение и выравнивание нескольких признаков ДНК могут быть достигнуты с помощью коммерческого спин-коатера и элайнера масок, обычно встречающихся в чистых помещениях.
Здесь мы описываем метод паттернирования клеток с одноклеточным разрешением с использованием адгезии на основе ДНК. Во-первых, фотопаттернирование с положительным фоторезистом используется для создания паттернов с высоким разрешением амино-модифицированной ДНК на альдегидно-модифицированной стеклянной подложке. Затем слайд обрабатывается для уменьшения неспецифического прикрепления клеток, а ячейки потока PDMS создаются для ограничения клеток над узорчатыми областями. Затем клетки мечутся короткими олигонуклеотидами ДНК, которые функционализируются холестерином и в результате вставляются в клеточную мембрану. Затем клетки протекают по микроструктурам ДНК. Гибридизация между ДНК клеточной поверхности и ДНК на поверхности стекла приводит к специфической адгезии клеток к образцу ДНК. Неприлипание клеток смывается, выявляя адгезивный клеточный паттерн. Этот процесс может быть повторен для создания шаблонов нескольких типов клеток или для создания многослойных структур. При желании клетки могут быть полностью встроены в ECM для 3D клеточной культуры.
1. Дизайнерский эксперимент
- Спланируйте желаемый эксперимент, учитывая размер признака, расстояние между признаками, количество вовлеченных типов клеток и расположение клеток по отношению друг к другу. Обратитесь к Дополнительному файлу 1,руководству по экспериментальному проектированию, и Дополнительному файлу 2,который содержит примеры последовательностей олиго.
- Проектирование фотомаски с помощью программного обеспечения для автоматизированного проектирования. Пример фотомаски приведен в дополнительном файле 3.
- Нарисуйте прямоугольник размеров стандартного слайда микроскопа (25 мм х 75 мм).
- Нарисуйте четыре прямоугольные области шириной 10 мм и длиной 10 мм, равномерно распределенные по слайду.
- В каждой области нарисуйте объекты, которые являются нужным размером, формой и интервалом для эксперимента. Клетки будут придерживаться только этих особенностей в эксперименте.
- Чтобы создать выровненные фотомашки для нескольких типов ячеек, создайте главный чертеж со всеми наборами объектов, а затем сохраните версии, соответствующие каждому типу ячеек.
- Закажите фотомаску прозрачности с высоким разрешением (не менее 20 000 точек на дюйм) из этого чертежа САПР с функциями, нарисованными в прозрачной форме 1.2.3, и более крупными областями черного цвета.
2. Фотопаттерн ДНК на альдегид-функционализированных слайдах (протокол, адаптированный из Scheideler et al.29)
- При создании паттернов нескольких типов клеток изготовьте фидуциальные маркеры на альдегидно-функционализированном слайде перед любым паттерном ДНК, чтобы облегчить выравнивание признаков. Альтернативные методы создания фидуциальных маркеров предложены в дополнительном файле 1.
- Для создания металлических фидуциальных маркеров применяйте положительный фоторезист S1813, как описано в шагах 2.3 - 2.11. Используйте фотомаску, содержащую большие объекты, которые будет легко выровнять позже. Включите эти функции в дизайн фотомаска, которые будут использоваться для создания паттернов ДНК.
- Нанесите тонкую пленку (100 ангстрем) титана на затвор с помощью электронно-пушечных испарений29. Удалите избыток металла и фоторезиста с помощью ацетона, а затем приступайте к фотопаттернажу ДНК.
- Готовят 20 мкМ раствор 5'-амино-модифицированного олиго в буфере ДНК (50 мМ фосфата натрия в воде, рН = 8,5). Рекомендуемые последовательности олиго см. в дополнительном файле 2.
ПРИМЕЧАНИЕ: Для некоторых паттернов и применений можно использовать всего 5 мкМ амино-модифицированного олиго, поэтому может потребоваться оптимизация концентрации поверхностной ДНК. - Предварительно нагреть конфорку до 100 °C.
- Используйте двустороннюю ленту или вакуум для прикрепления альдегидного функционализированного стекла к ротору спин-коатера.
ВНИМАНИЕ: Отслоение скольжения во время отжима покрытия представляет собой риск безопасности. Всегда используйте отжимной коатер в закрытом контейнере с крышкой, например, в акриловой коробке.
ПРИМЕЧАНИЕ: Пометьте угол слайда с помощью алмазного писца или аналогичного орудия, чтобы поцарапать стекло. Это помогает с идентификацией и ориентацией слайдов после того, как фоторезист был смыт. - Используйте одноразовую пипетку, чтобы опустить положительный фоторезист на альдегидный слайд. Для ровных покрытий добавьте небольшие капли фоторезиста поперек слайда, вместо одной большой капли посередине(дополнительный рисунок 1A).
- Используя отжимной коатер, вращайте затвор со скоростью 3000 об/мин в течение 30 с.
- Поместите зажим на конфорку при 100 °C в течение 1,5 мин (мягкая выпечка), чтобы сшить фоторезист.
- Снимите затвор с конфорки. Поместите фотомаску с функциями, желаемыми для этого эксперимента, поверх слайда и взвесьте фотомаску куском стекла(дополнительный рисунок 1B, C). Покройте всю установку в непрозрачнойкоробке (Дополнительный рисунок 1D). Экспонировать с помощью УФ-лампы (длина волны 365 нм, 360 мВт, 5 дюймов от слайда, общая плотность лучистой энергии 100мДж/см2)в течение 2 мин.
ПРИМЕЧАНИЕ: ультрафиолетовый свет нарушит полимерные связи в фоторезисте под прозрачными областями фотомаски, создавая области, где ДНК позже сможет прилипать. - Разработайте слайд, погрузвшись в решение разработчика на 3-5 мин(Дополнительный рисунок 1Е).
- Смойте избыток раствора разработчика водой. Высушите под струей воздуха или азота. (Дополнительный рисунок 1F).
- Подтвердите, что фотолитография была успешной, посмотрев на слайд под микроскопом. Поскольку фоторезист чувствителен к ультрафиолетовому излучению, сделайте этот шаг быстро, а затем сохраните слайд в темноте, готовя другие слайды (если применимо).
ПРИМЕЧАНИЕ: Успешно узорчатый слайд должен иметь четко определенные края для каждого объекта, без растрескивания и искажения признаков по краям. Примеры правильной и неправильной фотолитографии приведены на дополнительном рисунке 2А. Рекомендации по устранению неполадок см. в таблице 1, если фотолитография не обеспечивает требуемое качество функций. - Добавьте каплю 20 мкМ амино-модифицированного олиго раствора (шаг 2.1) на каждую фотопятнаполненный край слайда. Используйте наконечник пипетки, чтобы аккуратно распределить каплю по всей области, стараясь не поцарапать горку. (Дополнительный рисунок 1G).
- Выпекать горку в духовке с температурой 65-70 °C до полного высыхания раствора ДНК на поверхности слайда (около 1 ч).
- Выполните восстановительное аминирование, поместив узорчатые, запеченные слайды в 15-сантиметровую чашку для клеточных культур и поместите в вытяжной вытяжной вытяжной вытяжкой поверх шейкерки. Взвесьте 100 мг боргидрида натрия. В вытяжку добавьте 40 мл фосфатно-буферного физиологического раствора (PBS), аккуратно перемешайте и добавьте в блюдо, содержащее узорчатые слайды. Дайте реакции продолжаться в течение 15 мин с легким встряхиванием.
ПРИМЕЧАНИЕ: Амин на олиго сначала образует основание Шиффа с альдегидами на поверхности скольжения. Это обратимая ковалентная связь, которая должна быть преобразована в необратимую связь перед использованием в DPAC. Добавление восстановителя (боргидрида натрия) превращает основание Шиффа во вторичный амин путем восстановительного аминирования.
ОСТОРОЖНО: Реакция боргидрида натрия с водой создает газообразный водород и будет продолжаться в течение нескольких часов или дней после начала реакции. Выполните восстановительную этап аминирования в вытяжном вытяжке и храните все отходы раствора боргидрида натрия в открытом или свободно закрытом контейнере в вытяжном вытяжке в течение не менее 24 ч. - Удалите нереактированную ДНК, промыв дважды 0,1% додецилсульфата натрия (SDS) в воде, а затем три раза дистиллированной водой. Высушите горку под струей азота или воздуха.
- Промойте слайд ацетоном, чтобы удалить оставшийся фоторезист.
ПРИМЕЧАНИЕ: На данный момент ДНК необратимо и ковалентно прикреплена к слайду, и все нереагидные функциональные группы альдегида были преобразованы в спирты. Фоторезист больше не нужен. - Если будет выровнено несколько олигов, вернитесь к шагу 2.4, выровняйте фотомаску с фидуциальными метками и повторите.
ПРИМЕЧАНИЕ: Здесь эксперимент можно приостановить. Храните слайды в вакуумном адсорбаторе. В сухих условиях горки могут храниться до 3 месяцев без потери качества.
3. Сделать слайд гидрофобным (необязательным) (протокол адаптирован из Todhunter et al.24)
ПРИМЕЧАНИЕ: Предпочтительно, но не обязательно, изменять химический состав поверхности слайда, чтобы сделать его более инертным и гидрофобным. Неспецифическое прикрепление клеток уменьшается на этих поверхностях33,тем самым облегчая неспецифическое связывание клеток с нешаблочными участками слайда. Кроме того, если узорчатые клетки в конечном итоге будут встроены в гидрогель и перенесены с слайда, обработка поверхности необходима для надежного перемещения нагруженного клетками гидрогеля по слайду без искажений или разрывов. Силанизация (тридекафтор-1,1,2,2-тетрагидрооктилом) диметилхлорсилана приводит к наличию гидрофобных фторалкильных групп на поверхности слайда.
ВНИМАНИЕ: Выполните все шаги, начиная с 3.1 и далее в химическом вытяжном вытяжке, чтобы предотвратить воздействие паров уксусной кислоты и метиленхлорида.
- Промыть горкой 10% уксусной кислотой, а затем высушить под потоком воздуха.
- В стеклянной банке Коплина готовят раствор 60 мл метиленхлорида (дихлорметана), 0,6 мл триэтиламина и 0,6 мл (тридекафтор-1,1,2,2-тетрагидрооктила) диметилхлорсилана. Перемешать металлическим шпателем.
ПРИМЕЧАНИЕ: Эти реагенты чувствительны к воде. Их следует хранить в сухих условиях и использовать как можно более свежими. - Добавьте слайд в банку Coplin, содержащую раствор силана. Поместите банку Коплина на орбитальный шейкер (установленный на 60-80 оборотов в минуту) и дайте реакции силана и слайда прогрессировать в течение 15 минут.
- Используйте металлические щипцы, чтобы удалить слайд из раствора силана. Погрузите слайд в банку Coplin, содержащую метиленхлорид, на 1 минуту, чтобы удалить избыток силана с слайда.
- Погрузите слайд в коническую трубку 50 мл, содержащую этанол. Волновать. Погрузите горку в коническую трубку 50 мл, содержащую дистиллированную воду. Волновать.
ПРИМЕЧАНИЕ: Метиленхлорид и вода не смешиваются, поэтому для удаления избытка метиленхлорида перед окончательным промывкой водой необходимо промывка этанола. - Извлеките горку из воды и осмотрите ее. Горка должна быть достаточно сухой, с любыми каплями воды, имеющими угол контакта более 90°. Дайте слайдам полностью высохнуть и храните в вакуумном осушителях до использования.
ПРИМЕЧАНИЕ: Здесь эксперимент можно приостановить. Храните горку в сухих условиях.
4. Подготовьте проточные ячейки PDMS и слайд для эксперимента
ПРИМЕЧАНИЕ: Прямоугольные ячейки потока PDMS используются для концентрации клеток над узорчатыми областями слайда. Для экспериментов, культивируемых в 3D, проточная клетка образует форму для гидрогеля.
- Изготовление СУ-8 мастер для использования в качестве пресс-формы для проточных ячеек PDMS.
- Предварительный нагрев конфорки до 95 °C.
- Добавьте 5 мл SU-8 2075 в кремниевую пластину.
- Спиновое покрытие SU-8 на пластине при 500 об/мин в течение 10 с, а затем 1000 об/мин в течение 30 с. Это должно создавать объекты высотой до 240 мкм34.
- Мягко выпекайте на конфорке не менее 45 мин.
- Снимите пластину с конфорки. Положите фотомаску (см. Дополнительный файл 4)(эмульсия стороной вниз) поверх пластины и взвесьте ее стеклянным диском, чтобы обеспечить контакт между фотомаской и слайдом.
- Экспонирование с ультрафиолетовым светом (365 нм) для лучистой плотности энергии 350 мДж/см2.
- Выпекать на конфорке 12-15 мин.
- Поместите в широкую стеклянную емкость. Крышка пластины с решением разработчика SU-8. Поместите на шейкер и развивайте во время перемешивания не менее 15 мин.
- Используйте щипцы, чтобы удалить пластину из решения разработчика. Смойте в течение 5 с, распыляя больше раствора разработчика из бутылки для брызг. Распылить изопропиловым спиртом для промывки. Если появляется белый осадок, верните пластину в решение разработчика и развивайте дольше.
- Сухая пластина под потоком воздуха или азота.
- Выпекать горку в течение 5 мин.
ПРИМЕЧАНИЕ: После создания основной пластины ее можно повторно искивать бесконечно, пока функции остаются нетронутыми.
- Подготовьте PDMS.
- В весовую лодку добавляют полидиметилсилоксановый эластомер и сшиватель в соотношении 10:1 (по массе). Энергично перемешайте, чтобы обеспечить равномерное перемешивание.
- Дегазируйте PDMS в вакуумном адсорбаторе в течение 15-30 мин, пока не будут видны пузырьки.
- Поместите мастер-вафлю в 15 см тканевую культурную чашку. Налейте PDMS на пластину. При появлении пузырьков дегазуют в вакуумном адсорбциаторе на несколько минут.
- Выпекать в духовке при 60 °C в течение 3 ч.
ПРИМЕЧАНИЕ: После выпечки проточные ячейки PDMS могут храниться на столешнице неопределенно долго.
- Подготовьте проточные ячейки PDMS к эксперименту.
- Незадолго до начала эксперимента CMO-DPAC вырежьте необходимое количество проточных ячеек PDMS из мастер-пластины. Плазму окисляют с помощью 10 куб. см / мин комнатного воздуха в течение 90 с, чтобы сделать поверхность гидрофильной.
- Вырежьте каждую отдельную проточную ячейку так, чтобы с каждой стороны осталось 1-2 мм PDMS, затем разрежьте верхнюю и нижнюю части проточной ячейки, чтобы создать вход и выход.
- Извлеките узорчатый слайд, созданный на шагах 2 и 3. Выровнять поверх фотомаски.
- Используя фотомаску в качестве эталона, поместите ячейки потока PDMS на слайд в месте расположения каждой узорчатой области.
- Добавьте 50 мкл фосфатного буферного физиологического раствора (PBS) + 1% бычьего сывороточного альбумина (BSA) на вход каждой проточной клетки, как показано на дополнительном рисунке 1H. Убедитесь, что проточная ячейка полностью заполнена PBS + 1% BSA и что нет больших пузырьков. Немедленно перейдите к шагам 5 и 6.
ПРИМЕЧАНИЕ: Блокировка с помощью BSA минимизирует неспецифическую адгезию клеток к поверхности слайда.
5. Поднимите и пометьте клетки с модифицированной холестерином ДНК
- Приготовьте растворы ДНК, модифицированные холестерином.
- Для каждого набора клеток в эксперименте смешайте вместе 3 мкл запасного раствора 100 мкМ модифицированной холестерином универсальной якорной цепи с 3 мкл 100 мкМ запасного раствора адаптерной нити. Инкубировать в течение 1 минуты. Это позволит предварительно гибридизировать олиго. Добавьте 69 мкл фосфатно-буферного физиологического раствора (PBS) для создания универсального анкерного раствора + адаптера 4 мкМ.
- Для каждого набора клеток в эксперименте добавляют 3 мкл 100 мкМ универсального раствора Co-Anchor Strand, модифицированного холестерином, к 12 мкл PBS, создавая раствор 20 мкМ.
- Подготовьте одноэлелеточную суспензию (суспензии).
- Для адгезивных клеток используйте трипсин или другой диссоциационный агент для удаления клеток из колбы культуры. Добавьте культуральные среды для нейтрализации трипсина и центрифугу для гранулирования клеток. Для неприлипательных клеток соберите клеточную суспензию и центрифугу, чтобы гранулировать клетки.
- Повторно суспендируют клеточную гранулу в 1 мл ледяного PBS или безсыворочной среды. Перенесите 1-3 миллиона клеток в микроцентрифужную трубку размером 1,5 мл. Центрифуга при 160 х г в течение 4 мин.
ПРИМЕЧАНИЕ: Если используемый тип клеток склонен к слипанию / агрегации, используйте PBS без ионов кальция и магния для всех этапов промывки, чтобы уменьшить нежелательную агрегацию клеток. Если жизнеспособность является особой проблемой для используемого типа клеток, используйте безывороточные носители вместо PBS. Среда, содержащая фетальную бычий сыворотку, не рекомендуется для маркировки клеток, поскольку она может препятствовать включению липид-модифицированных олиго. 35 35
- Пометьте клетки олиго, модифицированными холестерином.
- Повторно суспендируют клеточную гранулу в 75 мкл ледяного PBS или безсыворочной среды. Храните клетки в ведре со льдом на протяжении всего процесса маркировки и промывки, чтобы максимизировать жизнеспособность клеток и свести к минимуму потерю модифицированных холестерином олиго с поверхности клеток.
ПРИМЕЧАНИЕ: Повторное сусплирование клеток перед добавлением ДНК гарантирует, что распределение ДНК равномерно по всей клеточной популяции. - Добавьте 75 мкл раствора универсального якоря + адаптера 4 мкМ, созданного на этапе 5.1.1, в микроцентрифужную трубку, содержащую клеточную суспензию. Тщательно перемешайте путем пипетки. Инкубировать в течение 5 мин на льду.
- Добавьте 15 мкл универсального ко-анкерного раствора в трубку микроцентрифуги. Тщательно перемешайте путем пипетки. Инкубировать в течение 5 мин на льду.
- Удалите лишние олиго из клеточной суспензии. Добавьте 1 мл ледяной PBS или безымяточной среды в трубку микроцентрифуги. Смешайте с пипеткой P1000. Центрифуга при 160 х г в течение 4 мин при 4 °C. Выбросьте супернатант. Повторите еще два раза.
ПРИМЕЧАНИЕ: Если клетки склонны к слипанию, пропустите клеточную суспензию через фильтр 40 мкм перед окончательной промывкой. Если клетки склонны к адсорбции на стороне трубки микроцентрифуги, рассмотрите возможность предварительной блокировки трубки казеином.
- Повторно суспендируют клеточную гранулу в 75 мкл ледяного PBS или безсыворочной среды. Храните клетки в ведре со льдом на протяжении всего процесса маркировки и промывки, чтобы максимизировать жизнеспособность клеток и свести к минимуму потерю модифицированных холестерином олиго с поверхности клеток.
6. Паттерны клеток, меченых ДНК
- Повторное суспендировать клетки в ледяной PBS или свободной от сыворотки среде для создания клеточного плотного раствора не менее 25 миллионов клеток / мл.
ПРИМЕЧАНИЕ: Для одного слайда с использованием четырех из проточных ячеек PDMS размером 10 мм x 15 мм x 200 мкм, описанных на этапе 4, требуется около 100 мкл этой плотной клеточной суспензии. Хотя большинство из этих клеток не будут придерживаться паттерна и в конечном итоге будут отброшены, наличие чрезвычайно концентрированного раствора клеток над паттерном значительно повышает эффективность клеточного паттернинга. - Подберите горку и слегка наклоните ее. Добавьте 25 мкл клеточной суспензии на вход каждой проточной ячейки на узорчатом слайде. Удалите раствор PBS + 1% BSA из выходного отверстия, позволив суспензии ячейки заполнить проточную ячейку PDMS. Инкубировать на льду или при комнатной температуре в течение 30 с.
ПРИМЕЧАНИЕ: На этом этапе, глядя на проточную ячейку под микроскопом, следует показать плотно упакованные клетки с небольшими или вообще без зазоров, видимых между клетками. См. дополнительный рисунок 2B. - Аспирировать 5 мкл клеточной суспензии от выходного отверстия слайда и добавить ее обратно во входное отверстие. Повторите 10 раз на проточную ячейку.
ПРИМЕЧАНИЕ: Адгезия клеток, меченых CMO, к слайду с днк-паттерном происходит почти мгновенно. Протекание клеток по паттерну в несколько раз увеличивает вероятность того, что клетка будет течь по данному пятно ДНК и будет захвачена. - Аккуратно пипетку PBS или безымяточную жиму в отверстие каждой проточной ячейки, чтобы вымыть лишние клетки. Соберите клеточную суспензию с выходного отверстия. Повторить 2-4 раза или до тех пор, пока визуальный осмотр слайда под микроскопом не подтвердит, что лишних клеток не осталось.
ПРИМЕЧАНИЕ: Может быть выгодно сохранить лишние ячейки от первой промывки. Если эффективность паттернирования неудовлетворительна, избыточные ячейки могут быть центрифугированы и повторно суспендированы в меньшем объеме PBS для создания более плотного раствора, а затем процесс может быть повторен из шага 6.2. - Повторите шаги 6.1–6.4 для каждого набора ячеек в шаблоне. Для шаблонов, в которых несколько типов клеток непосредственно шаблонируются поверхностным шаблоном, начните с наименее распространенного типа клеток шаблона и закончите наиболее распространенным типом клеток.
ПРИМЕЧАНИЕ: Желательно делать каждый раунд клеточной сборки последовательно, а не объединять клетки, даже в условиях, когда все клетки помечены ортогональными последовательностями ДНК. Объединение клеток эффективно разбавляет каждую популяцию клеток и снижает эффективность паттернов. - После завершения заключительного раунда сборки ячеек следующие шаги будут варьироваться в зависимости от конкретного эксперимента. Если ячейки предназначены для того, чтобы оставаться на стекле, добавьте среду в чашку Петри, содержащую слайд, а затем осторожно используйте щипцы, чтобы подтолкнуть ячейки потока PDMS к слайду. Если клетки будут встроены в гидрогель и культивированы в 3D, перейдите к шагу 7.
7. Перенос в гидрогель для 3D культуры (опционально)
- Готовят раствор гидрогеля-предшественника, содержащий 2% ДНКазы.
ПРИМЕЧАНИЕ: Состав раствора будет варьироваться в зависимости от экспериментальной установки. Матригель и смеси матригеля и коллагена хорошо работают в этом протоколе, но возможны и другие гидрогели. - Добавьте 50 мкл раствора гидрогеля, содержащего 2% ДНКазы, на вход каждой проточной ячейки. Аспирировать лишнюю жидкость из выхода, загоняя раствор гидрогеля в проточную ячейку. Для вязких предшественников гидрогеля может потребоваться небольшое наклонение затвора, чтобы помочь гидрогелю течь в проточную ячейку.
- Инкубируют слайд при 37 °C в течение 30-45 мин (в зависимости от кинетики гелеобразования гидрогеля), чтобы гидрогель мог застыть и расщеплить адгезию на основе ДНК между клетками и поверхностью.
- Выньте каждую проточную клетку с слайда и поместите поверх раствора предшественника гидрогеля.
- Добавьте 50 мкл предшественника гидрогеля в скважину с 2-мя скважинной камерной горкой или 6-скважинной пластиной.
- Пипетка 10 мкл PBS по обе стороны от каждой проточной ячейки.
- Используйте лезвие бритвы или тонкий пинцет, чтобы распределить PBS по всей длине проточной ячейки, а затем осторожно поднимите стороны проточной ячейки, чтобы PBS устремился под гидрогель.
ПРИМЕЧАНИЕ: Это будет «плавать» гидрогель по слайду, что позволяет передавать его без искажений или разрывов. - Используйте лезвие бритвы, чтобы осторожно переместить проточную ячейку к краю стеклянного затвора.
- Перевернуть слайд. Лезвием бритвы оттолкните проточную ячейку от затвора, чтобы она приземлился на лезвие бритвы.
- Соберите проточную ячейку с лезвия бритвы с помощью изогнутых щипцов. Инвертировать проточную ячейку так, чтобы клетки были на дне, а затем поместить сверху каплю раствора предшественника гидрогеля.
- Повторите шаги 7.4.1 - 7.4.6 для каждой проточной ячейки.
- Инкубировать в течение, по меньшей мере, 30 мин, чтобы гидрогель, содержащий клеток с рисунком, мог связываться с подложкой гидрогеля, что приводит к полному встраиванием клеток с рисунком.
- Извлеките проточную ячейку PDMS.
- Добавьте достаточное количество носителей для погружения проточной ячейки PDMS.
ПРИМЕЧАНИЕ: Приток сред ослабит адгезию между гидрогелем и проточной ячейкой PDMS. - Используйте изогнутые щипцы, ориентированные вдоль длинной оси проточной ячейки, чтобы мягко подталкивать проточную ячейку, пока она не выскочет и не всплывет в среде. Соберите проточную ячейку щипцами и выбросьте.
ПРИМЕЧАНИЕ: Для достижения оптимальных результатов распределите изогнутые щипцы и приложите мягкое давление к стенкам проточной ячейки PDMS. Приложить усилие в направлении длинной оси проточной ячейки.
- Добавьте достаточное количество носителей для погружения проточной ячейки PDMS.
8. Подтвердите успешную маркировку ячеек с помощью CMO (опционально, для устранения неполадок)
- Закажите флуоресцентно модифицированный (FAM или AF647) олигонуклеотид, который комплементален поверхностной последовательности адгезии переходной нити, используемой в эксперименте.
- Пометить клетки ДНК CMO и вымыть избыток ДНК, как описано в Шаге 5. Повторное суспенд в 200 мкл ледяного PBS.
- Составляют 4 мкМ раствор флуоресцентно меченого комплементарного олигонуклеотида в PBS. Добавьте 200 мкл этого раствора в клеточную суспензию. Насиживать на льду в течение 5 мин.
- Добавьте 1 мл ледяного PBS. Смешайте. Центрифугирование клеток, чтобы гранулировать их. Удалите супернатант. Повторите этот процесс еще два раза, чтобы вымыть любую ДНК, которая не гибридизировалась.
- Выполните аналитическую проточную цитометрию для количественной оценки присутствия ДНК на поверхности клетки.
- На проточном цитометре проанализируйте контрольные клетки, которые не были помечены ДНК. Установите ворота на основе этой популяции.
- Анализ клеток, меченых CMO, которые были обработаны флуоресцентно меченым комплементарным олигонуклеотидом.
- Рассчитайте средную интенсивность флуоресценции.
Этот протокол позволяет моделировать ячейки в 2D и 3D с высокой точностью и без использования пользовательских реагентов или дорогостоящего оборудования для чистых помещений. На рисунке 1 показан обзор протокола. Во-первых, ДНК-функционализированные слайды создаются с помощью фотолитографии. Затем ячейки помечаются ОКУ. Затем клетки перетекают через слайд, где они прикрепляются только к днк-функционализированным областям слайда. После того, как лишние клетки смываются, выявляется желаемый рисунок клеток. Эти клетки могут быть культивированы на слайде или встроены в гидрогель, содержащий ДНКазу, и перенесены с слайда для 3D-культуры клеток.
Маркировка клеток ОКУ позволяет прикрепить их к слайду с узором ДНК(рисунок 2). Во-первых, модифицированная холестерином универсальная якорная нить предварительно гибридизуется с adapter Strand. Далее раствор Universal Anchor + Adapter смешивают 1:1 с клеточной суспензией. Холестерин на комплексе Universal Anchor + Adapter вставляется в клеточную мембрану. Добавление модифицированной холестерином универсальной ко-якорной цепи, которая гибридизуется с универсальной якорной нитью, улучшает стабильность комплекса CMO в клеточной мембране за счет увеличения чистой гидрофобности комплекса26. После вымывания лишней ДНК из клеточной суспензии клетки перетекают по слайду. Гибридизация между переходной нитью и поверхностной нитью ДНК приводит к прикреплению клеток к областям слайда с днк-паттерном.
Рисунок клеток создается с помощью фотолитографии для ограничения прикрепления амино-модифицированных олиго ДНК к определенным областям альдегидно-модифицированного стеклянного слайда29 (фиг.3А). Положительный фоторезист наносится спин-покрытием на альдегидно-функционализированный слайд. Затем на слайд помещается прозрачность фотомаски, и слайд подвергается воздействию ультрафиолетового излучения. После разработки области слайда, которые подвергались воздействию ультрафиолетового света, больше не покрыты фоторезистом и, таким образом, подвергаются воздействию альдегидных групп. Затем 20 мкМ раствор олигосов ДНК, модифицированных амином, опускают на слайд и распространяют, чтобы покрыть узорчатые области. Выпечка с последующим восстановительным аминированием приводит к ковалентной связи между амино-модифицированной ДНК и слайдом. Примечательно, что этот процесс может быть повторен для паттерна нескольких олиго без потери функциональности ранее узорчатых олиго(рисунок 3B). Тем не менее, следует проявлять осторожность, чтобы избежать перекрывающихся паттернов, что приводит к присутствию обоих олигов в пониженнойконцентрации (дополнительный рисунок 3). Множественные клеточные популяции могут быть построены последовательно с помощью Adapter Strands, которые отличаются своей модульной областью (20 оснований, ближайших к концу 3').
Хотя этот протокол фотошаблонирования был разработан Scheideler et al. в контексте чистой комнаты, мы продемонстрировали, что можно достичь аналогичных результатов с помощью недорогой, «домашней» фотолитографической установки, которая легко помещается в химический вытяжной вытяжной вытяжной. Установка включает в себя спин-коатер стоимостью 400 долларов США, изготовленный из двигателя постоянного тока, цифрового контроллера и коробки для тортов CD, а также ультрафиолетовую лампу, которая была собрана из отдельных компонентов и размещена в перепрофилированном контейнере для острых деталей(Дополнительный рисунок 1). Основным преимуществом установки для домашней фотолитографии является то, что она очень доступна (< 1000 долларов США за все оборудование), но при этом может создавать одноэлекторные функции. Тем не менее, использование недорогого оборудования имеет свои ограничения - например, сложнее точно выровнять фидуциальные маркеры для создания нескольких олиго ДНК без использования выравнивания маски. Мы рекомендуем эту недорогую установку фотолитографии для лабораторий, которые не имеют удобного доступа к чистой комнате или которые хотят попробовать этот метод без больших инвестиций.
Чтобы определить оптимальные условия для клеточной адгезии, запрограммированной ДНК, мы систематически варьировали концентрации нитей ДНК на клеточных поверхностях и измеряли эффективность адгезии клеток к ДНК-модифицированным стеклянным поверхностям. Концентрация Universal Anchor + Adapter Strand и Universal Co-Anchor в растворах для маркировки варьировалась на несколько порядков величины(Рисунок 4A,B),в результате чего на клетку входилось10 4 -10 6 комплексов ДНК(Дополнительный рисунок 4). Клеточная адгезия зависела от дозы, с минимальной клеточной адгезией к структуре ДНК, когда клетки были помечены ОКУ в концентрации 0,05 мкМ или менее, и высокой заполняемостью при концентрации 2,5 мкМ и выше. Поэтому в большинстве экспериментов мы использовали раствор Universal Anchor + Adapter Strand размером 2 мкМ и раствор Universal Co-Anchor 2 мкМ. Ожидается, что адгезия клеток также уменьшится, если количество ДНК, используемой на поверхности стекла, уменьшится на29 или если несоответствия между переходной нитью и поверхностной нитью увеличится. Более подробная информация о конструкции последовательности Adapter Strand приведена в дополнительном файле 2. Маркировка CMO с использованием Adapter Strands без повторов CpG не стимулировала TLR9 в HEK-клетках, экспрессирующих TLR9 мыши(дополнительный рисунок 5).
Мы предоставляем несколько демонстраций того, что пересмотренный протокол обеспечивает воспроизводимую и эффективную адгезию клеток, запрограммированную ДНК. Например, эндотелиальные клетки пупочных вен человека (HUVECs), помеченные ОКУ, с высокой эффективностью придерживались паттернов ДНК. Приклеенные HUVEC, маркированные CMO, а также HUVEC с маркировкой LMO(рисунок 5A). Клетки, выкройка с использованием CMO-DPAC, сохранили свою жизнеспособность и функциональность. Клетки, помеченные ОКУ, окрашивали кальцеином AM и гомодимером этидия для оценки жизнеспособности(дополнительный рисунок 6). Различия в жизнеспособности по сравнению с немаркированными контрольными клетками были небольшими (94% против 97%). Одиночные MDCK, смоделированные через CMO-DPAC и перенесенные в Matrigel, смогли размножаться и правильно поляризоваться после 5 дней культивации(рисунок 5B). DPAC также предоставляет средства для разработки паттернов клеток в третьем измерении(рисунок 5C). Например, многослойные, многоклеточные агрегаты могут быть созданы путем чередования слоев клеток, помеченных комплементарными ОКУ(рисунок 5C). Эти эксперименты демонстрируют, что протокол воспроизводим, не оказывает негативного влияния на жизнеспособность или функциональность клеток и дает клеточные паттерны, которые могут быть успешно культивированы в пределах одной плоскости визуализации в 3D ECM.
Предоставляя ортогональные последовательности ДНК для прямой клеточной адгезии, DPAC предоставляет средство моделирования нескольких типов клеток на одной поверхности. Чтобы реализовать эту особенность DPAC, паттерны ДНК, генерируемые фотолитографией, должны быть выровнены по отношению друг к другу. Металлические фидуциарные маркеры, нанесенные на слайд, позволяли выровнивать несколько фотомаскателей и, следовательно, структурировать несколько типов клеток одновременно. MCF10As, окрашенные различными уникальными красителями, были помечены ортогональными ОКУ и узорами для создания визуализации логотипов UC Berkeley и UCSF(рисунок 6). Этот эксперимент демонстрирует, что несколько уникальных клеточных популяций могут быть склеено вместе с высокой точностью и без перекрестного загрязнения.
Успешное моделирование клеток с использованием CMO-DPAC требует высококачественной фотолитографии, достаточной концентрации олиго на поверхности клетки, высокой плотности клеток над рисунком и достаточной промывки. Сбой любого из этих шагов влияет на конечный результат. Дополнительный рисунок 2 включает примеры изображений правильной и неправильной фотолитографии(Дополнительный рисунок 2A),желаемую плотность клеток над рисунком для создания полностью занятых паттернов(Дополнительный рисунок 2B),потерю клеток с рисунком из-за чрезмерно энергичного пипетирования на последующих этапах DPAC(Дополнительный рисунок 2C)и нежелательное слипание клеток(Дополнительный рисунок 2D). В таблице 1 приведен список распространенных точек отказа и предлагаемые способы устранения неполадок. Использование флуоресцентных комплементарных олиго рекомендуется в качестве инструмента для устранения неполадок для подтверждения наличия узорчатой ДНК на слайде и присутствия ОКУ на поверхности клетки с помощью проточной цитометрии (см. Шаг 8 протокола).
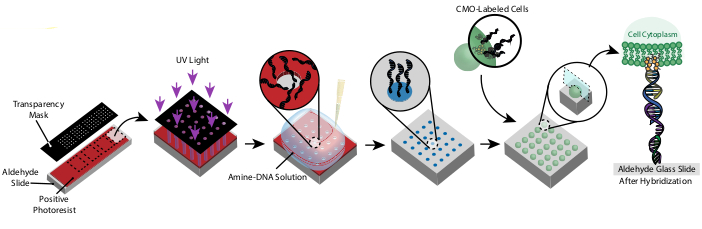
Рисунок 1:Обзор протокола CMO-DPAC. Во-первых, слайд с днк-узором создается путем покрытия альдегидного функционализированного стекла положительным фоторезистом, покрытия его прозрачной маской в желаемом рисунке и воздействия ультрафиолетового света. Фоторезист, подвергающийся воздействию ультрафиолета, смывается с разработчиком, оставляя открытые области альдегидного скольжения и позволяя связывать амино-функционализированную ДНК с поверхностью. Затем клетки маркируются ОКУ и перетекают по поверхности. ДНК на клеточной мембране гибридизуется с ДНК на поверхности, что приводит к адгезии. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
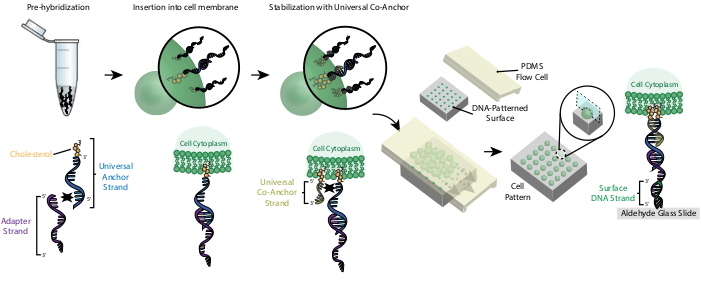
Рисунок 2:Ячейки помечаются ОКУ в поэтапном процессе. Во-первых, модифицированная холестерином универсальная якорная нить предварительно гибридизуется с adapter Strand. Далее раствор Universal Anchor + Adapter смешивают с клеточной суспензией. Холестерин на комплексе Universal Anchor + Adapter вставляется в клеточную мембрану. После инкубации модифицированная холестерином универсальная ко-якорная нить добавляется в клеточную суспензию, где она гибридизуется с универсальной якорной нитью и вставляется в клеточную мембрану. Добавление второй молекулы холестерина увеличивает чистую гидрофобность комплекса ДНК и стабилизирует его внутри мембраны26. После вымывания избытка ДНК клетки концентрируются и добавляются к проточной клетке PDMS поверх узорчатой поверхности. 3-х '-образный конец Adapter Strand гибридизируется с поверхностной нитью ДНК на стеклянном слайде, что приводит к адгезии к слайду, особенно в областях, функционализированных комплементарной ДНК. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
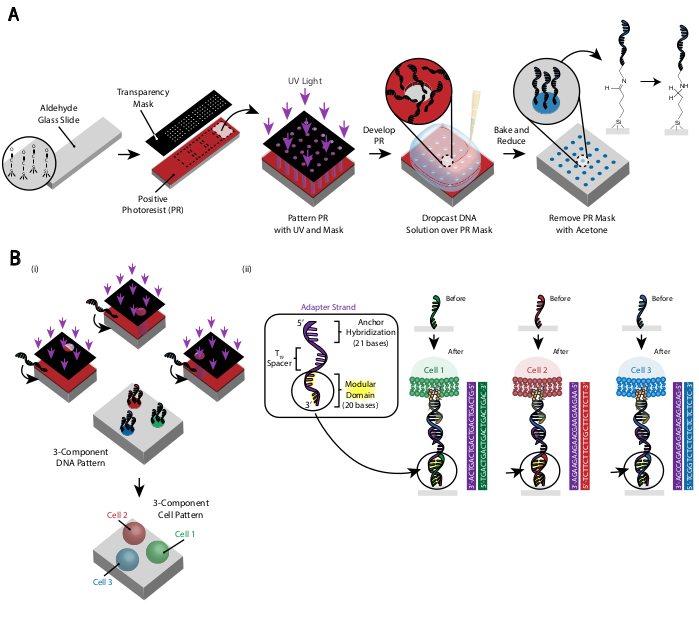
Рисунок 3:Фотолитография используется для создания слайдов с днк-паттерном, которые в конечном итоге будут диктовать размещение клеток. (A) Обзор процесса фотолитографии. Альдегидно-функционализированный слайд покрыт положительным фоторезистом. Ультрафиолетовый свет светит на слайд через прозрачную фотомаску, которая является прозрачной там, где желательна адгезия клеток. После того, как слайд разработан, области, которые ранее подвергались воздействию ультрафиолетового света, теперь подверглись воздействию альдегидных групп. 20 мкМ раствор олиго амино-функционализированной ДНК затем опускают на слайд и распределяют по узорчатым областям. Затем слайд запекают, чтобы индуцировать образование связей Шиффа (C = N) между аминовой и альдегидной группами, обратимой ковалентной связи29. Последующее восстановительное аминирование 0,25% боргидрида натрия в PBS превращает основание Шиффа во вторичный амин путем восстановительного аминирования, что приводит к необратимой связи между ДНК и слайдом. Оставшийся фоторезист затем может быть удален путем промывки ацетоном. Этотпроцесс может быть повторен для создания многокомпонентных паттернов ДНК и, следовательно, проведения экспериментов с несколькими клеточными популяциями. i) После того, как первое олиго узорчато, слайд снова покрывается фоторезистом, и протокол действует по-прежним. Выравнивание фотомашек с использованием фидуциарных маркеров необходимо для паттернов нескольких нитей ДНК. ii) Каждый тип ячейки, по образцу которого он моделируется, отличается в 20-базовой модульной области Adapter Strand. Используя ортогональные наборы комплементарных олиго, несколько типов клеток могут быть узорчаты без перекрестной адгезии. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
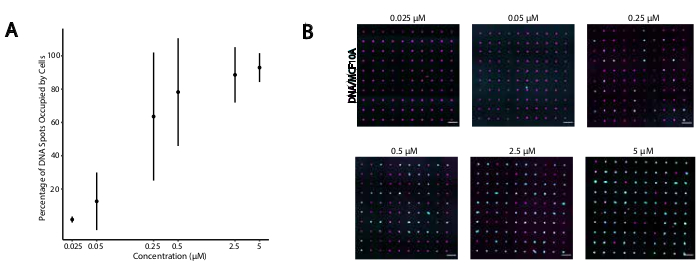
Рисунок 4:Адгезия клеток, меченых CMO, к паттернам ДНК увеличивается в зависимости от концентрации CMO во время маркировки. В этом эксперименте Universal Anchor + Adapter Strand (предварительно гибридизированный) и Universal Co-Anchor использовались в равных концентрациях. Концентрация относится к концентрации CMO в клеточной суспензии во время маркировки CMO клеток. (A)Количественная оценка процентного соотношения пятен ДНК диаметром 15 мкм, которые были заняты мечеными CMO клетками MCF10A в зависимости от концентрации CMO во время маркировки клеток. Данные, представленные в виде среднего ± стандартное отклонение от трех экспериментов. (B)Репрезентативные изображения паттернов ДНК (пурпурного) и прилипающих MCF10As (циан) при различных концентрациях ОКУ. Шкала = 100 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
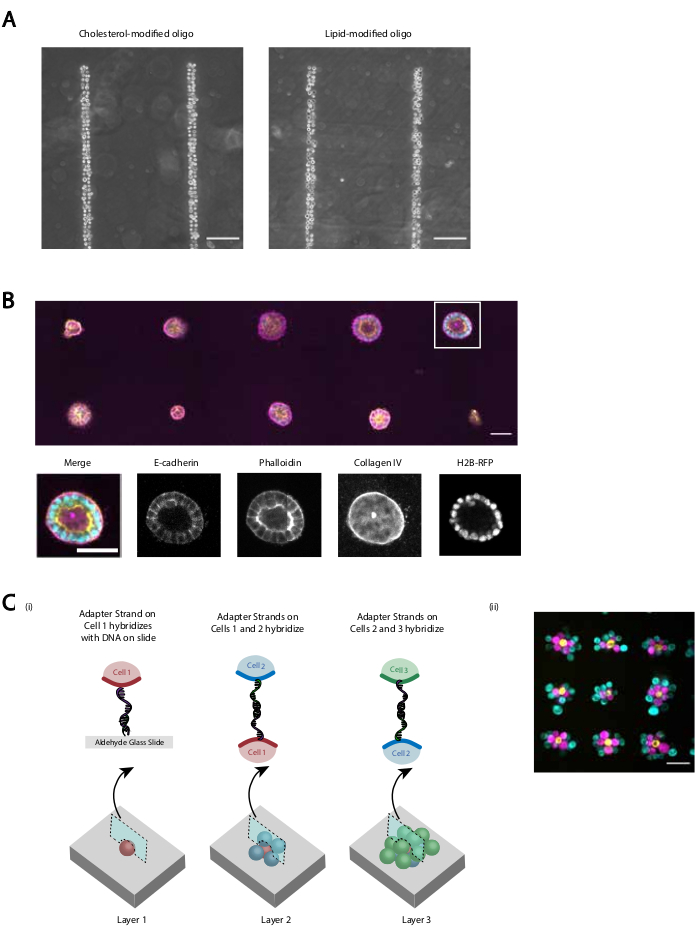
Рисунок 5:CMO-DPAC может быть использован для создания двумерных клеточных паттернов, которые впоследствии могут быть встроены в трехмерный гидрогель для культуры и/или слоистые для создания многослойных структур. (A)Прямое сравнение между мечеными CMO эндотелиальными клетками пуповинной вен человека (HUVECs) и ХУВЕКами, меченными LMO, придерживаются линейного паттерна ДНК. Оба метода маркировки клеток приводят к почти 100% заполнению паттерна ДНК. (B) Одиночные клетки почек собачки Мадина-Дарби (MDCK), экспрессирующие H2B-RFP, были построены на пятнах диаметром 15 мкм, расположенных на 200 мкм друг от друга и впоследствии встроенных в Матригель. После 120 ч культивирования полученные эпителиальные кисты фиксировали и окрашивали на Е-кадгерин, актин и коллаген IV. Сфероид в белом ящике показан подробно. Шкала = 50 мкм.(C)Многослойные клеточные структуры могут быть созданы путем маркировки отдельных клеточных популяций комплементарными переходными нитями и последовательного паттерна, чтобы каждое новое добавление клеток прилипало к клеточному слою перед ним. (i)Схема последовательного паттернирования клеточных популяций для создания многослойных структур. (ii) Трехслойные клеточные агрегаты MCF10As (визуализированные с помощью красителей) были созданы с использованием этого процесса. Шкала = 50 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.

Рисунок 6:Несколько типов клеток могут быть узорчатыми без перекрестного загрязнения или потери адгезии. Множественные олиго, модифицированные амином, были последовательно выстроены на альдегидном слайде и выровнены с помощью металлических фидуциарных маркеров. Три популяции MCF10As (циановые, пурпурные, желтые) были окрашены уникальными красителями, помеченными дополнительными ОКУ, и узором на слайде, что привело к изображению логотипов UC Berkeley и UCSF. Шкала 1 мм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Дополнительный рисунок 1: Пример изображений настольная фотолитография. (A) Скольжение на спин-коатере, покрытом положительным фоторезистом, перед спиновым покрытием. (B) Изображение прозрачной фотомаски. (C) Во время экспозиции фотомаскама зажата между слайдом с фоторезистовым покрытием и стеклянным диском. (D) Корпус для ультрафиолетовой лампы был изготовлен из перепрофилированного контейнера для резких кадров. (E) Слайд погружен в решение разработчика. (F) Разработан слайд. (G)Амино-модифицированный раствор ДНК распространяется на узорчатые области слайда. (H)Ячейки потока PDMS, размещенные поверх узорчатых областей слайда. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 2: Некоторые примеры распространенных сбоев этого протокола. (A)(i) Недопекание перед воздействием ультрафиолета или чрезмерно развивающиеся функции после экспозиции могут привести к особенностям, которые имеют зубчатые края и могут быть нерегулярными по размеру. (ii)Пример правильно фотопаттерированного слайда, который имеет чистые края вокруг объектов, одинаковый размер объекта и отсутствие явных трещин в рисунке. Шкала бара = 50 мкм. (B) Плотность ячеек имеет решающее значение для эффективности паттернов. При наблюдении за клетками поверх рисунка под микроскопом между клетками должно существовать мало промежутков, о чем свидетельствует пример изображения слева. Шкала бар = 50 мкм. (C) Узорчатые клетки могут быть чувствительны к силам жидкости, возникающим в результате чрезмерно энергичного пипетирования, которое может повредить и вытеснить узорчатые клетки. Многослойные клеточные агрегаты особенно уязвимы, так как одна клетка внизу поддерживает структуру нескольких клеток. (i)Массив клеточных агрегатов, успешно встроенных в Matrigel. (ii)Сетка клеточных агрегатов, которые смещались в результате слишком энергичного пипетирования вязкого Матригеля. (D)Может произойти слипание клеток, особенно с эпителиальными клетками. Эти сгустки обычно гомотипичны, но могут быть гетеротипическими (клетки, прилипающие к уже узорчатым клеткам другого типа), если клетки особенно липкие. Изображение показывает, что три различные популяции MCF10As были построены на массиве, состоящем из трех различных пятен ДНК размером с одну клетку (15 мкм). Большинство пятен ДНК имеют 2-4 прикрепленных клетки. Слипание может быть решено путем обработки ЭДТА или путем фильтрации сгустков перед паттерном. Шкала бара = 100 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 3: Перекрывающиеся фотошаблоны приводят к присутствию обоих олигов в пониженной концентрации. Два ортогональных амино-модифицированных олиго были фотопаттернированы последовательно, сначала вертикальная линия (Strand 1), а затем горизонтальная линия, которая перекрывала его (Strand 2). Затем олиго были визуализированы путем гибридизации с флуоресцентными комплементарными олиго. (A) Флуоресцентное изображение Strand 1. (B)Количественная оценка профиля флуоресценции Strand 1 над вертикальной линией 100 мкм, охватывающей перекрытие. (C) Флуоресцентное изображение Strand 2. (D)Количественная оценка профиля флуоресценции Strand 2 по горизонтальной линии 100 мкм, охватывающей перекрытие. Шкала бара = 50 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 4: Количественная оценка комплексов ДНК на поверхности клетки в зависимости от концентрации маркировки CMO. HUVEC маркировали различными концентрациями раствора CMO, промывали, а затем инкубировали флуоресцентной комплементарной нитью. Набор микросфер MESF (Молекулы эквивалентного растворимого фторхрома) использовался для проведения количественной проточной цитометрии и оценки количества комплексов ДНК на поверхности клетки в зависимости от концентрации CMO во время маркировки. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 5: Маркировка ОКУ не стимулирует реакцию TLR9. Был проведен эксперимент, чтобы выяснить, будет ли маркировка CMO запускать механизм обнаружения ДНК TLR9 и будет ли на это влиять CpG в последовательности Adapter Strand. HEK-клетки, экспрессирующие TLR9 мыши, инкубировали в течение ночи с 0,2 мкМ либо ODN 1826 (агонист TLR9, содержащий CpG), либо универсальный анкер CMO + универсальный ко-якорь + адаптерная нить, содержащая ту же последовательность, что и ODN 1826 (CMO-CpG), либо универсальная анкерная + универсальная ко-якорная + адаптерная нить, содержащая аналогичную последовательность, но с заменой CpG на GPC (CMO-GpC). Стимуляция TLR9 приведет к производству SEAP (секретируемой эмбриональной щелочной фосфатазы). Секреция SEAP была количественно определена колориметрическим анализом (поглощением). Условия лечения сравнивали с покоящиеся клетками, которые обрабатывались только PBS. Инкубация с CMO-GPC не стимулировала экспрессию TLR9. Инкубация с CMO-CpG была немного выше, чем в покоящихся клетках, но намного ниже, чем ODN-1826. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 6: Жизнеспособность клеток после процесса маркировки ОКУ. Чтобы оценить, как протокол влияет на жизнеспособность, HUVEC были разделены на четыре популяции: одна оставалась на льду в течение 1 ч, одна была имитирована PBS, но в остальном проходила через все этапы центрифуги и промывки, одна была помечена ОКУ, а другая была помечена ОКУ и отфильтрована через фильтр 40 мкм для удаления сгустков. Затем клетки окрашивали кальцеином AM и гомодимером этидия для оценки количества живых и мертвых клеток. Все методы лечения привели к значительному снижению жизнеспособности, чем контроль льда (односторонний ANOVA с пост-специальным анализом Tukey), но медиана жизнеспособности для CMO-маркировки (с фильтрацией или без нее) составила около 94%. Данные, собранные в результате трех независимых экспериментов. * = p < 0,05. = p < 0.0001 Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
| Результат | Возможная причина(и) | Предлагаемые исправления |
| Фотолитография – признаки трескаются | Непоследовательная или неадекватная мягкая выпечка | Увеличить время мягкой выпечки до 3 минут; проверить фактическую температуру плиты и повысить температуру по мере необходимости |
| Фотолитография – объекты не резкие или в них остается фоторезист | Недостаточное развитие | Увеличить время, которое слайд тратит на решение разработчика; включить мягкое перемешивание |
| Фотолитография – особенности, несогласованные на слайде | Ультрафиолетовый свет может быть не центрирован или не сфокусирован должным образом | Отрегулируйте настройку ультрафиолетового излучения, чтобы обеспечить коллимированный свет равномерной интенсивности |
| Клетки не прилипают к узорчатым пятнам с высокой эффективностью | Недостаточно ДНК на поверхности | Подтвердите, что ДНК присутствует на поверхности, гибридизировав слайд с флуоресцентными комплементарными олиго, а затем визуализируя под микроскопом |
| Ячейки недостаточно маркированы ОКУ | Добавление флуоресцентных комплементарных олиго в клеточную суспензию и подтверждение флуоресценции с помощью проточной цитометрии | |
| Недостаточно клеток над шаблоном | Собирайте ячейки путем вымывания из проточной ячейки PDMS, центрифуги и повторного суспендирования в меньшем объеме для концентрации клеток | |
| Слишком много оставшегося CMO в клеточной суспензии, гибридизирующейся с ДНК на слайде | Добавьте еще один шаг стирки. Обязательно удаляйте как можно больше супернатанта с каждой стиркой. | |
| Слишком большая интернализация CMO из-за времени и температуры | Работайте быстро после маркировки клеток с помощью CMO; хранить клетки и скользить по льду и использовать ледяные реагенты | |
| Клетки слипаются | Клетки не были адекватно разделены во время трипсинизации | Используйте PBS + 0,04% ЭДТА во время промывки клеток; пропустить клеточную суспензию через фильтр 35 мкм перед окончательной промывкой |
| Клетки прилипают не специкоррецит | Если в одной конкретной области – может быть из-за царапин на слайде, смещения проточных клеток PDMS или разлива ДНК за пределы области паттерна | Избегайте царапин, будьте осторожны, чтобы выровнять ячейки потока PDMS с областью шаблона |
| Если клетки прилипают везде – неадекватная блокировка или промывка | Добавьте больше смывов после структурирования клеток; пипетку более энергично во время промывок; блок с 1% BSA дольше до начала клеточного паттернинга; силанизировать слайд (дополнительный этап 3) или подтвердить, что силанизация была успешной, измерив угол контакта капли воды | |
| Пузырьки образуются внутри проточной ячейки | Погрешности пипетирования, неровная гидрофильная поверхность, создаваемая при окислении плазмы | Если пузырьки маленькие, добавьте PBS на входную часть проточной ячейки, и они могут быть вымыты. Если пузырьки больше, приложите мягкое давление к проточной ячейке PDMS, подталкивая пузырьки к входу или выходу. |
| Клетки первоначально придерживаются паттерна, но удаляются во время промывок, паттернов других типов клеток или добавления предшественника гидрогеля | Силы сдвига от слишком энергичного пипетирования могут привести к тому, что клетки отсоезываются от поверхности. | Пипетируйте более осторожно во время последующих промывок, раундов клеточного рисунка или добавления предшественников гидрогеля. Поскольку предшественники гидрогеля вязкие, они с большей вероятностью вызовут смещение рисунка, поэтому соблюдайте особую осторожность. Многослойные структуры, как правило, тяжелы сверху и более восприимчивы к смещению. |
| Ткань деформируется при 3D-переносе | Гидрогелевые палочки для скольжения | Подтверждение гидрофобности скольжения с помощью измерений угла контакта |
| Используйте лезвие бритвы, чтобы полностью поднять PDMS на обоих краях, позволяя PBS плавать под тканью | ||
| Это может произойти с чистыми гидрогелями коллагена - рассмотрите возможность корректировки концентрации белка или состава гидрогеля | ||
| Клетки не переносятся с гидрогелем и остаются на слайде | Увеличение концентрации Turbo DNAse или увеличение времени инкубации | |
| Гидрогель недостаточно твердый | Увеличить время инкубации и/или механизм гелеобразования для рассматриваемого гидрогеля (например, для коллагена, убедитесь, что рН правильный) | |
| Гидрогелевые разрывы при удалении PDMS | Сделайте pdMS проточные клетки гидрофильными, используя плазменное окисление перед началом эксперимента, чтобы они легко отделяются при добавлении среды. Используйте щипцы очень осторожно, чтобы отсоедить PDMS. |
Таблица 1: Руководство по устранению неполадок для выявления и устранения потенциальных сбоев, которые могут возникнуть в результате этого протокола. В частности, плохая адгезия клеток к рисунку может иметь много коренных причин, и это руководство должно помочь в выявлении и решении этих проблем.
Дополнительный файл 1. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный файл 2. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный файл 3. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный файл 4. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
В этой статье мы представляем подробный протокол для паттернов клеток с высоким разрешением в 2D и 3D для экспериментов с культурой клеток in vitro. В отличие от ранее опубликованных версий этого метода, представленный здесь протокол фокусируется на удобстве использования: он не требует узкоспециализированного оборудования, и все реагенты могут быть приобретены у поставщиков вместо того, чтобы требовать пользовательского синтеза. В отличие от других методов микроструктурирования клеток, этот метод является быстрым и агностичным по клеточному типу: он не требует специфической адгезии к белкам внеклеточного матрикса15. Клетки, созданные CMO-DPAC, могут быть встроены во внеклеточный матрикс, такой как матригель или коллаген, что приводит к 3D-культурам с гораздо более высоким пространственным разрешением, чем это в настоящее время возможно с помощью методов экструзионной печати22. CMO-DPAC может быть использован для создания от сотен до тысяч микроскопических объектов на слайде, что позволяет одновременно выполнять множество реплик.
Одним из наиболее важных параметров успеха этого протокола является плотность клеток, добавленных к ячейкам потока поверх узорчатого слайда. В идеале плотность должна составлять не менее 25 миллионов клеток/мл. При загрузке в проточные ячейки эта плотность ячеек приводит к почти плотно упакованному монослою клеток над рисунком(дополнительный рисунок 2B). Эти высокие плотности клеток максимизируют вероятность того, что клетка поселится непосредственно на вершине пятна ДНК и прилипнет. Уменьшение плотности ячеек снизит общую эффективность паттернов. Другим важным шагом в этом протоколе является тщательная повторная приостановка клеток в PBS или средах, свободных от сыворотки, перед добавлением раствора CMO. ОКУ очень быстро делятся на клеточные мембраны, и добавление раствора CMO непосредственно к клеточной грануле приведет к гетерогенной маркировке клеток. После добавления раствора CMO в клеточную суспензию важно тщательно перемешать путем пипетирования, чтобы ячейки были равномерно помечены СХМО. После инкубации необходимо тщательно вымыть лишние ОКУ с помощью нескольких этапов центрифугирования и промывки. Избыток свободного ОКУ, присутствующий в клеточной суспензии, будет связываться с узорчатой амино-модифицированной ДНК на стеклянном слайде, блокируя гибридизацию и адгезию CMO-модифицированных клеток в суспензии. Время также является ключевым фактором для этого протокола. Важно работать как можно быстрее при использовании ОКУ и держать клетки на льду, чтобы свести к минимуму интернализацию ОКУ и максимизировать жизнеспособность клеток. Эксперименты по проточной цитометрии показали, что ОКУ не сохраняются так долго на поверхности клеток, как ЖИО, с потерей 25% комплексов ОКУ в течение двух часов инкубации нальду 36. Кроме того, жизнеспособность клеток будет уменьшаться по мере увеличения времени обработки клеток. Жизнеспособность может быть максимизирована путем быстрой работы, удержания клеток на льду, использования ледяных реагентов и использования безсывороточных сред для обеспечения некоторых питательных веществ.
Хотя CMO-DPAC может быть мощным способом изучения клеточной биологии путем паттернирования клеток с высокой точностью, у него есть свои ограничения. Эксперименты CMO-DPAC могут быть сложными, особенно потому, что экспериментальная сложность добавляется с несколькими типами клеток, слоями или 3D-культурой клеток(Дополнительный файл 1). Экспериментальные сбои могут быть распространены при запуске этого протокола, как описано в таблице 1. Поэтому мы рекомендуем пользователям проводить проверки качества (подтверждая, что ДНК присутствует на слайде, подтверждая, что клетки достаточно помечены ДНК (шаг 8), подтверждая, что избыточные клетки были тщательно смыты и т. Д.), Чтобы убедиться, что эксперимент удался, и определить шаги, которые могут потребовать дальнейшей оптимизации. Мы надеемся, что информация, представленная в этой рукописи и ее дополнительных файлах, поможет облегчить любые необходимые способы устранения неполадок.
Холестерин представляет собой биологически активную молекулу, интернализация которой может влиять на клеточный метаболизм, экспрессию генов и текучесть мембран37,38. В предыдущем исследовании сравнивали влияние на экспрессию генов клеток, меченых CMO и LMO, с использованием секвенирования одноклеточной РНК. Клетки HEK, меченые CMO, имели измененную экспрессию генов по сравнению с немаркированными и мечеными LMO клетками36. Маркировка клеток С помощью ОКУ привела к дифференциальной экспрессии (> в 1,5 раза) восьми генов относительно немаркированных контрольных органов, включая AP2B1, который был связан с транспортом холестерина и сфинголипида (GeneCards), и MALAT1, длинную некодирующую РНК, которая регулирует накопление холестерина39. Хотя эти транскрипционные реакции незначительны, они, тем не менее, могут вызывать беспокойство, если рассматриваемый эксперимент изучает метаболизм, динамику мембран или другие пути, связанные с холестерином, в клетках.
Этот протокол является гибким и может быть скорректирован в соответствии с потребностями каждого эксперимента. Поскольку CMO вставляет себя в липидную мембрану вместо использования какого-либо конкретного рецептора, метод является агностиком клеточного типа (ЗДЕСЬ были продемонстрированы HUVECs, MCF10As, HEKs и MDCK). Хотя холестерин является другим гидрофобным якорем, чем наши ранее опубликованные ЖИО, мы до сих пор обнаружили, что они ведут себя аналогично. Таким образом, мы ожидаем, что ОКУ будут работать с любым из широкого спектра типов клеток, которые мы ранее опубликовали с ЖИО, включая, но не ограничиваясь, нервными стволовыми клетками, фибробластами, мононуклеарными клетками периферической крови, опухолевыми клетками и первичными эпителиальными клетками молочной железы6,23,27,29,36 . Маркировка CMO не стимулирует TLR9, предполагая, что протокол совместим с иммунными клетками. Мембранное включение ОКУ является функцией общего размера клетки и степени отрицательного заряда в клетке гликокаликса35. Таким образом, мы включили протокол (шаг 8) для тестирования степени включения мембраны, которая поддаться быстрой оптимизации. Специфические особенности каждого клеточного паттерна неизбежно будут варьироваться в зависимости от экспериментального дизайна (см. Дополнительный файл 1 для получения дополнительного руководства). Хотя протокол фотопаттернирования, описанный выше для паттерна ДНК, рекомендуется, любой метод пространственного ограничения капель раствора амина-ДНК должен работать, например, использование капельных принтеров высокого разрешения. Разрешение шаблона и минимальный интервал между функциями зависят от используемого метода. Также теоретически возможно комбинировать ДНК-фотопаттерные участки этого протокола с другими методами, которые использовались для маркировки клеток с ДНК, например, с ДНК, гибридизированной с мембранно-экспрессированными цинковыми пальцами40,с использованием NHS-конъюгированной ДНК41и реагируя на остатки азидо сиаловой кислоты на поверхности клетки с фосфин-конъюгированной ДНК42. . CMO-DPAC может быть применен к различным экспериментам, которые требуют жесткого контроля над расстоянием между клетками, включая исследования взаимодействий между парами клеток, эксперименты с кокультурами, изучающие передачу сигналов от клеток-отправителей к клеткам-«приемникам», и исследования влияния близлежащих внеклеточных сигналов на дифференцировку стволовых клеток6,29 . Метод также может быть использован для создания микротизов, которые могут быть использованы для изучения миграции клеток в трех измерениях, самоорганизации клеток вткани 23,27и динамического взаимодействия между клетками и ECM27. Мы надеемся, что этот протокол предоставит исследователям доступную платформу для изучения новых применений клеточного паттерна на основе ДНК с высоким разрешением в их собственных лабораториях.
Z.J.G. является консультантом и акционером Provenance Biosciences.
Авторы хотели бы поблагодарить Джереми Гарсию за тестирование этого протокола и Бхушана Харбикара за обучение работе с оборудованием в UCSF Biomedical Micro and Nanotechnology Core. Это исследование было частично поддержано грантами Программы исследований рака молочной железы Министерства обороны (W81XWH-10-1-1023 и W81XWH-13-1-0221), NIH (U01CA199315, DP2 HD080351-01, 1R01CA190843-01, 1R21EB01918181-01A и 1R21CA182375-01A1), NSF (MCB1330864) и Центра клеточного строительства UCSF (DBI-1548297), научно-технического центра NSF. O.J.S финансировался стипендией NSF Graduate Research Fellowship, стипендией Siebel и стипендией P.E.O. Z.J.G и A.R.A. являются исследователями Chan-Zuckerberg BioHub.
| Name | Company | Catalog Number | Comments |
| 2-well Chambered Coverglass w/ non-removable wells | Thermo Fisher Scientific | 155379 | |
| Acetic Acid | Sigma-Aldrich | A6283 | |
| Adapter with External SM1 Threads and Internal SM3 Thread | ThorLabs | SM3A1 | |
| Aldehyde Functionalized Slides | Schott | Nexterion Slide AL | Store under dry conditions after opening. |
| All Plastic Syringes, 1 mL | Fisher Scientific | 14-817-25 | |
| Amine-Modified DNA Oligo | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| Aspheric Condenser Lens | ThorLabs | ACL7560 | |
| Borosilicate Disc, 6in Diameter X 1/2in Thick | Chemglass | CG-1906-23 | |
| Cell Culture Dishes 60x15 mm style | Corning | 353002 | |
| Cholesterol-Modified Oligo | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| Diamond Scribe | Excelta | 475B | |
| DNA Oligonucleotide | IDT | n/a | See Supplemental File 1 for suggested sequences. |
| DPBS, no calcium, no magnesium | Thermo Fisher Scientific | 14190250 | |
| Isopropyl Alcohol | Sigma-Aldrich | 278475 | |
| Matrigel Matrix, Growth Factor Reduced | Corning | 354230 | |
| Methylene Chloride (Stabilized/Certified ACS) | Fisher Scientific | D37-4 | |
| MF-321 Developer | Kayaku Advanced Materials | n/a | |
| Microposit S1813 Positive Photoresist | Kayaku Advanced Materials | n/a | |
| Ø3" Adjustable Lens Tube, 0.81" Travel | ThorLabs | SM3V10 | |
| Oven | Thermo Scientific | 51-028-112H | |
| PE-50 Compact Benchtop Plasma Cleaning System | Plasma Etch | PE-50 | |
| Photomask (custom) | CAD/Art Services | n/a | Minimum feature size guaranteed by CAD/Art Services is 10 microns. |
| Razor Blades | Fisher Scientific | 12-640 | |
| RCT Basic Hot Plate | IKA | 3810001 | |
| Silicon Wafer (100 mm) | University Wafer | 590 | |
| Sodium Borohydride, 98%, granules | Acros Organics | 419471000 | |
| Spin Coater Kit | Instras | SCK-200 | This is a low cost option, but any spin coater that can maintain a speed of 3000 rpm will suffice. |
| SU-8 2075 | Microchem | Y111074 0500L1GL | |
| SU-8 Developer | Microchem | Y020100 4000L1PE | |
| Sylgard 184 Silicone Elastomer Kit | Dow | 2646340 | |
| Syringe Needles | Sigma-Aldrich | Z192341 | |
| T-Cube LED Driver, 1200 mA Max Drive Current | ThorLabs | LEDD1B | |
| Tridecafluoro-1,1,2,2-tetrahydrooctyl dimethylchlorosilane | Gelest | SIT8170.0 | |
| Triethylamine | Sigma-Aldrich | 90335 | |
| Turbo DNase | Thermo Fisher Scientific | AM2238 | |
| Tweezers Style N7 | VWR | 100488-324 | The curved shape of these tweezers is essential for delicately picking up the PDMS flow cells containing patterned tissues. |
| UV LED (365 nm, 190 mW (Min) Mounted LED, 700 mA) | ThorLabs | M365L2 | |
| Wafer Tweezers | Agar Scientific | T5063 | |
| WHEATON Dry-Seal vacuum desiccator | Millipore Sigma | W365885 |
- Kreeger, P. K., Strong, L. E., Masters, K. S. Engineering approaches to study cellular decision-making. Annual Review of Biomedical Engineering. , 49-72 (2018).
- Goubko, C. a., Cao, X. Patterning multiple cell types in co-cultures: A review. Materials Science and Engineering C. 29 (6), 1855 (2009).
- Sun, W., et al. The bioprinting roadmap. Biofabrication. 12 (2), 022002 (2020).
- Liu, W. F., Chen, C. S. Cellular and multicellular form and function. Advanced Drug Delivery Reviews. 59 (13), 1319-1328 (2007).
- Duffy, R. M., Sun, Y., Feinberg, A. W. Understanding the role of ECM protein composition and geometric micropatterning for engineering human skeletal muscle. Annals of Biomedical Engineering. 44 (6), 2076-2089 (2016).
- Chen, S., et al. Interrogating cellular fate decisions with high-throughput arrays of multiplexed cellular communities. Nature Communications. 7, 10309 (2016).
- Shaya, O., et al. Cell-cell contact area affects notch signaling and notch-dependent patterning. Developmental Cell. 40 (5), 505-511 (2017).
- Rao, N., et al. A co-culture device with a tunable stiffness to understand combinatorial cell-cell and cell-matrix interactions. Integrative Biology. 5 (11), 1344 (2013).
- Sriraghavan, V., Desai, R. A., Kwon, Y., Mrksich, M., Chen, C. S. Micropatterned dynamically adhesive substrates for cell migration. Langmuir. 26 (22), 17733-17738 (2010).
- Wong, L., Pegan, J. D., Gabela-Zuniga, B., Khine, M., McCloskey, K. E. Leaf-inspired microcontact printing vascular patterns. Biofabrication. 9 (2), 021001 (2017).
- Chen, T. H., et al. Directing tissue morphogenesis via self-assembly of vascular mesenchymal cells. Biomaterials. 33 (35), 9019-9026 (2012).
- Laurent, J., et al. Convergence of microengineering and cellular self-organization towards functional tissue manufacturing. Nature Biomedical Engineering. 1 (12), 939-956 (2017).
- Lin, C., Khetani, S. R. Micropatterned co-cultures of human hepatocytes and stromal cells for the assessment of drug clearance and drug-drug interactions. Current Protocols in Toxicology. 2017, 1-23 (2017).
- Hui, E. E., Bhatia, S. N. Micromechanical control of cell-cell interactions. Proceedings of the National Academy of Sciences of the United States of America. 104 (14), 5722-5726 (2007).
- D'Arcangelo, E., McGuigan, A. P. Micropatterning strategies to engineer controlled cell and tissue architecture in vitro. BioTechniques. 58 (1), 13-23 (2015).
- Martinez-Rivas, A., González-Quijano, G. K., Proa-Coronado, S., Séverac, C., Dague, E. Methods of micropatterning and manipulation of cells for biomedical applications. Micromachines. 8 (12), (2017).
- Lee, S., et al. Simple lithography-free single cell micropatterning using laser-cut stencils. Journal of Visualized Experiments. (158), e60888 (2020).
- Strale, P. O., et al. Multiprotein printing by light-induced molecular adsorption. Advanced Materials. 28 (10), 2024-2029 (2016).
- Melero, C., et al. Light-induced molecular adsorption of proteins using the primo system for micro-patterning to study cell responses to extracellular matrix proteins. Journal of Visualized Experiments. (152), e60092 (2019).
- Reid, J. A., Mollica, P. M., Bruno, R. D., Sachs, P. C. Consistent and reproducible cultures of large-scale 3D mammary epithelial structures using an accessible bioprinting platform. Breast Cancer Research. , 1-13 (2018).
- Wang, Z., Lee, S. J., Cheng, H. -. J., Yoo, J. J., Atala, A. 3D bioprinted functional and contractile cardiac tissue constructs. Acta Biomaterialia. 70, 48-56 (2018).
- Miri, A. K., et al. Effective bioprinting resolution in tissue model fabrication. Lab on a Chip. 19 (11), 2019-2037 (2019).
- Todhunter, M. E., et al. Programmed synthesis of three-dimensional tissues. Nature Methods. 12 (10), 975-981 (2015).
- Todhunter, M. E., Weber, R. J., Farlow, J., Jee, N. Y., Gartner, Z. J. Fabrication of 3D microtissue arrays by DNA programmed assembly of cells. Current Protocols in Chemical Biology. 8 (3), 147-178 (2016).
- Csizmar, C. M., Petersburg, J. R., Wagner, C. R. Programming cell-cell interactions through non-genetic membrane engineering. Cell Chemical Biology. 25 (8), 931-940 (2018).
- Weber, R. J., Liang, S. I., Selden, N. S., Desai, T. A., Gartner, Z. J. Efficient targeting of fatty-acid modified oligonucleotides to live cell membranes through stepwise assembly. Biomacromolecules. 15 (12), 4621-4626 (2014).
- Hughes, A. J., et al. Engineered tissue folding by mechanical compaction of the mesenchyme. Developmental Cell. 44 (2), 165-178 (2018).
- Weber, R. J., et al. Rapid organoid reconstitution by chemical micromolding. ACS Biomaterials Science & Engineering. 2 (11), 1851-1855 (2016).
- Scheideler, O. J., et al. Recapitulating complex biological signaling environments using a multiplexed, DNA-patterning approach. Science Advances. 6 (12), (2020).
- Viola, J. M., et al. Guiding cell network assembly using shape-morphing hydrogels. Advanced materials (Deerfield Beach, Fla.). , 2002195 (2020).
- Mohammad, A., Davis, M., Aprelev, A., Ferrone, F. A. Note: Professional grade microfluidics fabricated simply. Review of Scientific Instruments. 87 (10), 1-4 (2016).
- Lee, O. J., Chuah, H. S., Umar, R., Chen, S. K., Yusra, A. F. I. Construction of cost effective homebuilt spin coater for coating amylose-amylopectin thin films. Journal of Fundamental and Applied Sciences. 9 (2), 279 (2018).
- Webb, K., Hlady, V., Tresco, P. A. Relative importance of surface wettability and charged functional groups on NIH 3T3 fibroblast attachment, spreading, and cytoskeletal organization. Journal of Biomedical Materials Research. 41 (3), 422-430 (1998).
- Processing Guidelines for: SU-8 2025, SU-8 2035, SU-8 2050, SU-8 2075. Microchem SU-8 2000 Permanent Expoxy Negative Photoresist Available from: https://kayakuam.com/wp-content/uploads/2019/09/SU-82000DataSheet2025thru2075Ver4.pdf (2019)
- Palte, M. J., Raines, R. T. Interaction of nucleic acids with the glycocalyx. Journal of the American Chemical Society. 134 (14), 6218-6223 (2012).
- McGinnis, C. S., et al. MULTI-seq: sample multiplexing for single-cell RNA sequencing using lipid-tagged indices. Nature Methods. 16 (7), 619-626 (2019).
- Maxfield, F. R., van Meer, G. Cholesterol, the central lipid of mammalian cells. Current Opinion in Cell Biology. 22 (4), 422-429 (2010).
- Luo, J., Yang, H., Song, B. L. Mechanisms and regulation of cholesterol homeostasis. Nature Reviews Molecular Cell Biology. 21 (4), 225-245 (2020).
- Liu, L., Tan, L., Yao, J., Yang, L. Long non-coding RNA MALAT1 regulates cholesterol accumulation in ox-LDL-induced macrophages via the microRNA-17-5p/ABCA1 axis. Molecular Medicine Reports. 21 (4), 1761-1770 (2020).
- Mali, P., Aach, J., Lee, J. H., Levner, D., Nip, L., Church, G. M. Barcoding cells using cell-surface programmable DNA-binding domains. Nature Methods. 10 (5), 403-406 (2013).
- Hsiao, S. C., et al. Direct cell surface modification with DNA for the capture of primary cells and the investigation of myotube formation on defined patterns. Langmuir. 25 (12), 6985-6991 (2009).
- Gartner, Z. J., Bertozzi, C. R. Programmed assembly of 3-dimensional microtissues with defined cellular conductivity. Proceedings of the National Academy of Sciences. (17), 1-5 (2009).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved