Crioconservazione delle cellule germinali primordiali e rinascita dei ceppi di Drosophila
* Questi autori hanno contribuito in egual misura
In questo articolo
Riepilogo
È altamente auspicabile un metodo di conservazione a lungo termine per i ceppi di Drosophila come alternativa al frequente trasferimento di mosche adulte in fiale di cibo fresco. Questo protocollo descrive la crioconservazione delle cellule germinali primordiali di Drosophila e la rinascita del ceppo attraverso il loro trapianto in embrioni ospiti agametici.
Abstract
I ceppi di Drosophila devono essere mantenuti mediante il frequente trasferimento di mosche adulte in fiale nuove. Ciò comporta il pericolo di deterioramento mutazionale e cambiamenti fenotipici. Lo sviluppo di un metodo alternativo per la conservazione a lungo termine senza tali modifiche è quindi imperativo. Nonostante i precedenti tentativi di successo, la crioconservazione degli embrioni di Drosophila non è ancora di uso pratico a causa della bassa riproducibilità. Qui, descriviamo un protocollo per la crioconservazione delle cellule germinali primordiali (PGC) e la rinascita del ceppo tramite trapianto di PGC crioconservate in embrioni ospiti agametici di Drosophila melanogaster (D. melanogaster). Le PGC sono altamente permeabili agli agenti crioprotettivi (CPA) e la variazione evolutiva e morfologica tra i ceppi è meno problematica rispetto alla crioconservazione degli embrioni. In questo metodo, le PGC vengono raccolte da circa 30 embrioni donatori, caricate in un ago dopo il trattamento CPA e quindi crioconservate in azoto liquido. Per produrre gameti derivati da donatori, le PGC crioconservate in un ago vengono scongelate e quindi depositate in circa 15 embrioni ospiti agagametici. Con questo protocollo è stata raggiunta una frequenza di almeno il 15% di moscerini fertili e il numero di progenie per coppia fertile è sempre stato più che sufficiente per far rivivere il ceppo originale (il numero medio di progenie è stato di 77,2 ± 7,1), indicando la capacità delle PGC crioconservate di diventare cellule staminali germinali. Il numero medio di mosche fertili per ago era di 1,1 ± 0,2 e 9 aghi su 26 hanno prodotto due o più progenie fertile. Si è scoperto che 11 aghi sono sufficienti per produrre 6 o più progenie, in cui è probabile che siano inclusi almeno una femmina e un maschio. L'ospite agagametico permette di ravvivare rapidamente il ceppo semplicemente incrociando mosche femmine e maschi appena emerse. Inoltre, le PGC hanno il potenziale per essere utilizzate in applicazioni di ingegneria genetica, come l'editing del genoma.
Introduzione
Il mantenimento dei ceppi di Drosophila mediante il trasferimento di mosche adulte in nuove fiale di cibo comporta inevitabilmente l'accumulo di mutazioni e cambiamenti epigenetici nel tempo. Lo sviluppo di un metodo alternativo per il mantenimento a lungo termine dei ceppi di Drosophila senza tali cambiamenti è imperativo, soprattutto per i ceppi di riferimento in cui l'intero genoma deve essere mantenuto. Sono stati descritti diversi tentativi riusciti di crioconservare embrioni o ovaie di Drosophila 1,2,3. Sfortunatamente, non sono ancora di uso pratico a causa della bassa riproducibilità. Infatti, gli embrioni in fase iniziale hanno un basso tasso di sopravvivenza dopo la crioconservazione a causa del loro alto contenuto di tuorlo, che impedisce la permeazione e la diffusione dell'agente crioprotettivo (CPA) 2,3. La permeabilità al CPA è anche fortemente limitata dagli strati cerosi degli embrioni in fase avanzata. È difficile e dispendioso in termini di tempo trovare un periodo di tempo specifico per ceppo in cui gli embrioni abbiano un alto tasso di sopravvivenza e uno strato di cera più sottile. Recentemente, Zhan et al.4 hanno migliorato i metodi per la permeabilizzazione degli embrioni, il carico di CPA e la vitrificazione e hanno crioconservato con successo embrioni di ceppi multipli. Tuttavia, i metodi non sono facili da applicare perché la vitalità degli embrioni dopo la permeabilizzazione tende ad essere scarsa. Pertanto, sono ancora necessari ulteriori miglioramenti e lo sviluppo di approcci alternativi. I metodi che prevedono la crioconservazione delle cellule germinali primordiali (PGC) sono un approccio alternativo per il mantenimento a lungo termine dei ceppi di Drosophila.
Il trapianto di PGC (chiamato anche cellula polare) è stato utilizzato per generare chimere germinali, in particolare femmine, per studiare processi come gli effetti materni delle mutazioni letali zigotiche e la determinazione del sesso delle cellule germinali 5,6,7,8,9,10,11,12 . Le PGC sono molto più piccole degli embrioni e sono probabilmente altamente permeabili alla maggior parte dei crioprotettori. Inoltre, la variazione evolutiva e morfologica tra i ceppi è meno problematica e un ospite agametico consente un rapido ripristino di interi genomi. Recentemente abbiamo sviluppato un nuovo metodo di crioconservazione PGC13, che previene i cambiamenti genetici ed epigenetici altrimenti inevitabili nei ceppi di Drosophila. Qui presentiamo il protocollo dettagliato.
Questo metodo di crioconservazione richiede competenze specifiche nella manipolazione e nella strumentazione delle PGC. Mentre un approccio graduale può essere una soluzione efficiente per coloro che non hanno familiarità con esso, potrebbe non essere adatto per i piccoli laboratori a causa dei requisiti della strumentazione. Questo protocollo di crioconservazione PGC può essere adattato più facilmente per l'uso con diverse specie di Drosophila e diverse specie di insetti rispetto ai protocolli di crioconservazione degli embrioni a causa delle minori differenze di sviluppo e morfologiche. Le PGC possono anche essere potenzialmente utilizzate in applicazioni di ingegneria genetica, come l'editing del genoma 14,15,16. In sintesi, questo metodo può essere utilizzato nei centri di stoccaggio e in altri laboratori per mantenere mosche e altri ceppi di insetti per periodi di tempo prolungati senza cambiamenti.
Protocollo
1. Preparazione dell'attrezzatura
- Sistema di micromanipolatore: assemblare un sistema di micromanipolatore per raccogliere e trapiantare le cellule (Figura 1A).
- Vetrini per la raccolta di PGC (Figura 2A)
- Per preparare la colla all'eptano, tagliare un nastro biadesivo lungo circa 30 cm e immergerlo per una notte in 7 ml di soluzione di eptano tecnico (normale).
- Disegna due linee di riferimento parallele per l'allineamento dell'embrione sul retro di un vetrino.
- Stendere gocce della colla eptanica di cui sopra sul vetrino (sul lato senza le linee) utilizzando una pipetta Pasteur. Asciugare all'aria la superficie del vetrino fino a quando non diventa bianca.
- Ripetere l'aggiunta e la stesura di gocce di colla eptano e asciugare nuovamente il vetrino.
NOTA: La colla impedisce alle soluzioni liquide di diffondersi sulla superficie piana e facilita il caricamento delle soluzioni acquose in un ago. - Per realizzare cornici per piscine embrionali, incolla tre strati di nastro vinilico standard di 0,2 mm di spessore, come il nastro isolante, su un tagliere. Taglia il nastro in rettangoli larghi 1,5 cm. Quindi, taglia via tutti e tre gli strati di nastro, lasciando una cornice di 2 o 3 mm.
NOTA: Dopo aver allineato gli embrioni, viene apposta una cornice per il pool di embrioni, per formare un pool per gli embrioni.
- Aghi per trapianti
NOTA: Tutti gli aghi disponibili in commercio al momento di questo studio erano troppo stretti o troppo larghi per la crioconservazione PGC.- Crea un ago usando un capillare di vetro e un estrattore. Utilizziamo un estrattore NARISHIGE PN-31 con il livello del riscaldatore a 85.0-98.4, il livello principale del magnete a 57.8 e il sottolivello del magnete a 45.0.
- Per realizzare un ago con uno spessore della parete approssimativo di 1 μm e una punta di circa 200 μm di lunghezza con un diametro interno di 10-12 μm, lucidare la punta dell'ago nel seguente processo in tre fasi (Figura 3). Per prima cosa, macinare la punta dell'ago con un angolo di 30° a una velocità di 780 giri/min fino a quando la punta ha un diametro interno di 10-12 μm. Questa prima fase di macinazione dura circa 1 ora.
NOTA: Per evitare di rompere la punta dell'ago, ruotare prima la mola e poi spostare delicatamente l'ago verso il basso sulla mola. - Disegna una linea sulla parte superiore dell'ago per tracciare l'angolo desiderato. Ruotare l'ago in senso antiorario di 90° e lucidarlo nuovamente a una velocità di 180 giri/min. Questa operazione richiede circa 5 minuti.
- Ruotare l'ago in senso orario di 45° e lucidarlo a una velocità di 180 giri/min per un secondo.
- Posizionare un vetrino di raccolta con una goccia di miscela di acido cromico (ATTENZIONE: tossico) sul tavolino del microscopio. Fissare l'ago al supporto del capillare (Figura 1D) con un angolo di 10°-13° rispetto alla superficie del vetrino, spostare con cautela l'ago verso il basso e immergere la punta nella miscela di acido cromico.
- Tirando e spingendo lo stantuffo (Figura 1B), caricare e scaricare meccanicamente la soluzione dall'ago più volte per rimuovere i detriti di vetro nell'ago. Assicurati di pulire anche la parete esterna.
- Lavare l'interno e l'esterno dell'ago due volte con acqua distillata per rimuovere completamente l'acido cromico.
2. Raccolta e crioconservazione delle PGC
- Raccolta di embrioni
- Trasferire un numero adeguato di mosche del ceppo donatore di interesse (circa 450 per ciascun sesso per la coppetta di raccolta degli embrioni) in una coppetta per la raccolta degli embrioni con una piastra per la raccolta degli embrioni (Figura 1E) e incubarle a 25 °C. Di solito utilizziamo mosche parentali di 3-5 giorni che vengono allevate in condizioni meno affollate a temperatura ambiente (23-25 °C).
- Eseguire due pre-pre-pre-prelievi di 30 minuti e scartare le uova deposte. Poiché le femmine possono trattenere le uova fecondate che si sviluppano nell'ovidotto, questo passaggio è necessario per sincronizzare la deposizione delle uova nella fase 2.1.3 (Figura 4).
- Dopo i due pre-pre-pre-prelievi, raccogliere gli embrioni per 50 minuti e poi incubare gli embrioni raccolti in una camera umidificata a 25 °C per consentire agli embrioni di svilupparsi fino allo stadio di blastoderma (stadio iniziale 517). Il tempo di incubazione è solitamente di 100 minuti, ma può essere esteso fino a 120 minuti, a seconda del ceppo (Figura 4).
NOTA: Una camera umidificata viene realizzata posizionando un tovagliolo di carta umido sul fondo di una scatola di plastica e spruzzandolo con una nebbia d'acqua prima dell'uso. Negli embrioni in stadio 5 iniziale, la formazione di PGC è completa, ma la cellularizzazione somatica non lo è. Lo stadio esatto di un embrione viene determinato al microscopio composto nella fase 2.4.
- Embrioni decorionanti
- Depositare una goccia di acqua distillata su un filtro a maglie di acciaio inossidabile (150 mesh, apertura 109 μm, diametro filo 60 μm; Figura 1F). Usando una pinza, raccogli gli embrioni dalla piastra di raccolta degli embrioni e mettili nella goccia d'acqua.
- Premere la carta velina contro il colino da sotto per assorbire l'acqua. Aggiungere goccioline di soluzione fresca di ipoclorito di sodio al 5% (come Cl) agli embrioni e picchiettare continuamente il colino per 10 s.
- Lavare gli embrioni spruzzandoli direttamente con acqua distillata e premere la carta velina contro il colino da sotto per assorbire l'acqua. Ripetere questo passaggio 3 volte.
- Allineamento di embrioni decoroniati
- Sotto uno stereomicroscopio, usa le pinze per trasferire gli embrioni. Allineare gli embrioni decoroniati in due file su un vetrino per la raccolta di PGC lungo le due linee di riferimento (Figura 2A). Gli embrioni sono orientati con la parte anteriore a destra (il lato da manipolare) e con il lato ventrale rivolto verso l'alto.
NOTA: Questa fase dovrebbe essere completata in 20 minuti, durante i quali di solito allineiamo circa 40 embrioni. - Applicare un telaio per la piscina embrionale attorno agli embrioni sul vetrino per la raccolta di PGC. Far cadere 1 μL di soluzione CPA (1x soluzione Ephrussi-Beadle Ringer, EBR, contenente il 20% di glicole etilenico e 1 M saccarosio; 1x EBR: 130 mM NaCl, 5 mM KCl, 2 mM CaCl2 e 10 mM Hepes a pH 6,9) in due punti separati nell'area racchiusa dal telaio e riempire la piscina con olio di silicone per evitare che gli embrioni si secchino (Figura 2A).
NOTA: Per preparare la soluzione di CPA, sciogliere completamente 10,26 g di saccarosio in circa 20 mL di H2O distillato contenente 3 mL di soluzione 10 x EBR. Aggiungere 6 mL di glicole etilenico e poi aggiungere H2O distillato fino a 30 mL. Dopo un'accurata miscelazione, filtrare la soluzione attraverso una membrana monouso da 0,22 mm.
- Sotto uno stereomicroscopio, usa le pinze per trasferire gli embrioni. Allineare gli embrioni decoroniati in due file su un vetrino per la raccolta di PGC lungo le due linee di riferimento (Figura 2A). Gli embrioni sono orientati con la parte anteriore a destra (il lato da manipolare) e con il lato ventrale rivolto verso l'alto.
- Raccolta di PGC
- Posizionare il vetrino per la raccolta PGC del passaggio 2.3.2 sul tavolino di un microscopio dotato di un sistema di micromanipolazione. Collegare l'ago al supporto del capillare e portare il primo embrione nella fila sinistra e la punta dell'ago sullo stesso piano focale. Caricare l'olio di silicone nell'ago per 2-3 s.
- Iniziare la raccolta di PGC dagli embrioni nella riga di sinistra. Utilizzando un obiettivo 20x, spostare delicatamente la punta dell'ago sulla superficie dell'estremità anteriore dell'embrione e penetrare l'embrione verso l'estremità posteriore, non spostando l'ago ma spostando il tavolino del microscopio.
- Quando la punta dell'ago raggiunge l'estremità posteriore, ritrarre leggermente l'ago e scaricare completamente il tuorlo nell'ago appena all'interno dello strato di cellule somatiche.
- Mantenendo costante la pressione nell'ago, spostare la punta dell'ago sui PGC appena all'interno del polo posteriore e delicatamente, ma senza impiegare molto tempo, caricare i PGC.
- Estrarre rapidamente l'ago dall'embrione e scaricare il tuorlo e altri contaminanti dall'ago nel pool di olio di silicone, mantenendo i PGC nell'ago. Quindi, caricare l'olio di silicone pulito dalla piscina.
- Ripetere i passaggi da 2.4.2 a 2.4.5 per gli altri embrioni nella fila di sinistra. Prima di prelevare le PGC da un nuovo embrione, depositare la maggior quantità possibile di olio di silicone caricato al punto 2.4.5 all'interno dello strato di cellule somatiche, mantenendo le PGC caricate nell'ago. Ciò garantisce che i PGC appena caricati siano adiacenti ai PGC raccolti in precedenza senza alcun materiale intermedio tra di loro.
- Dopo aver completato la raccolta delle PGC dagli embrioni nella fila di sinistra, separare il più possibile le PGC dal tuorlo e dagli altri contaminanti. Per raggiungere questo obiettivo, depositare tutte le PGC nell'ago sulla superficie di un embrione e rimuovere eventuali tuorli o altri contaminanti in un altro embrione vicino.
- Successivamente, raccogli i PGC dagli embrioni nella riga di destra. Combina i PGC raccolti dalle righe destra e sinistra.
- Applicazione di un agente crioprotettivo (CPA) ai PGC
- Dopo aver lavato l'ago con CPA in una goccia, caricare CPA fresco in un'altra goccia nell'ago e aggiungere CPA alle PGC depositate sull'embrione. Il volume della CPA dovrebbe essere equivalente a quello delle PGC.
- Rimuovere la maggior quantità possibile di CPA dal cluster di PGC 1-2 s dopo l'aggiunta di CPA. I PGC si restringono leggermente e diventano di forma quadrata subito dopo l'aggiunta di CPA.
- Svuotare l'ago e poi caricare l'olio di silicone per 5 secondi o più. Caricare tutti i PGC raccolti e quindi caricare nuovamente l'olio di silicone per 5 s o più. I PGC sono ora inseriti tra due strati di olio di silicone (Figura 2B).
NOTA: È importante rimuovere il più possibile tuorlo, CPA e altri contaminanti.
- PGC criopreservanti
- Aprire il rubinetto a tre vie (Figura 1C) e quindi staccare l'ago dal micromanipolatore. Tamponare l'olio dalla superficie dell'ago con carta velina morbida. Non tocchi direttamente la punta dell'ago con il fazzoletto.
- Fissare l'ago a un portaaghi e bloccarlo in posizione alla base utilizzando del nastro in vinile (Figura 1H). Applicare un'etichetta sul tubo di supporto.
- Congelare il supporto con l'ago rivolto verso il basso immergendolo in azoto liquido. Non rilasciare il supporto fino a quando il liquido non smette di frizzare fuori dal rack.
- Conservare il supporto in un serbatoio di stoccaggio dell'azoto liquido nell'area della fase liquida, non nell'area della fase vapore.
3. Scongelamento e trapianto di PGC
- Raccolta, decoroninazione e allineamento di embrioni da mosche ospiti agagametiche
- Raccogliere e decoronionare gli embrioni dai moscerini ospiti agametici seguendo il passaggio 2.
- Allineare gli embrioni dell'ospite agametico di stadio 5 su un vetrino per trapianto. Tuttavia, questa volta, orientare il posteriore verso destra (il lato da manipolare) e il ventrale verso l'alto (Figura 5). Allineare circa 30 embrioni in due file in 20 minuti.
- Durante l'allineamento degli embrioni, azionare un umidificatore per 2-10 minuti se l'umidità della stanza lo richiede (Tabella 1). L'umidità ideale è compresa tra il 30% e il 40%, ma può variare a seconda delle condizioni termiche.
- Scongelamento e trapianto di PGC in embrioni ospiti
- Per scongelare rapidamente i PGC crioconservati, infilare il supporto contenente l'ago nella soluzione EBR 1x a temperatura ambiente con l'ago rivolto verso il basso e tenerlo immerso per 10 s.
- Posizionare il vetrino per trapianti sul tavolino di un microscopio. Collegare l'ago congelato al supporto del capillare e portare il primo embrione nella fila di sinistra e la punta dell'ago sullo stesso piano focale.
- Utilizzando un obiettivo 20x, spostare delicatamente la punta dell'ago sulla superficie dell'estremità posteriore dell'embrione.
- Pungolare delicatamente l'esterno di ogni embrione e assicurarsi che torni lentamente alla sua forma originale. Il pungolo confermerà che la pressione interna dell'embrione non è né troppo alta né troppo bassa.
- Muovi delicatamente l'ago e penetra in un embrione dal polo posteriore.
- Depositare delicatamente circa 10-20 PGC appena all'interno del polo posteriore, precisamente tra la membrana vitellina e lo strato cellulare somatico dell'embrione. Evitare di depositarli nello strato cellulare somatico. Se il liquido perivitellino fuoriesce dall'embrione, aspirare il liquido fuoriuscito nell'ago e rimuoverlo.
- Ritrarre l'ago dall'embrione. Ripetere i passaggi 3.2.5 e 3.2.6 per gli embrioni successivi.
4. Incubazione di embrioni e ripristino dei ceppi di donatori
- Prelevare gli embrioni che non ricevono PGC trapiantati e incubare gli embrioni rimanenti in una camera umidificata (Figura 1G) a 25 °C.
- A partire da 24 ore dopo il trapianto e il prima possibile dopo la schiusa, utilizzare le pinze per prelevare e trasferire le larve schiuse in fiale standard di cibo per Drosophila e incubare a 25 °C.
- Per ravvivare il ceppo, incrociare femmine e maschi appena emersi (Figura 6).
NOTA: Gli ospiti agamitici consentono di ripristinare l'intero genoma in una sola volta senza passare a ceppi di cromosomi bilanciatori. La coesistenza di maschi agametici in una fiala non avrà importanza perché le femmine, anche se accoppiate con loro, non mostrano risposte post-accoppiamento a lungo termine, inclusa una diminuzione della ricettività alla riaccoppiamento18,19.
Risultati
L'efficienza del trapianto di PGC crioconservato è stata riportata da Asaoka et al.13 ed è riportata nella Tabella 2 per il trapianto di PGC crioconservati per 1 giorno o più in azoto liquido. Il tasso di schiusa è stato di 168/208 embrioni trapiantati (80,8%) e la vitalità da embrione ad adulto è stata di 87/208 (41,8%). La frequenza di mosche fertili è stata di 28/87 (32,2%). Questa frequenza non differiva tra le PGC crioconservate per 8-30 giorni e quelle crioconservate per 31-150 giorni (20/57 vs. 8/30, G' = 0,63, p >0,1, d.f. = 1). Il numero medio di progenie per coppia è stato di 77,2 ± 7,1 (n = 18, 28-122), indicando la capacità delle PGC crioconservate di diventare cellule staminali germinali. Dei 26 aghi, 10 non hanno prodotto progenie fertile, 7 aghi hanno prodotto 1 progenie fertile, 7 aghi hanno prodotto 2 progenie fertile e 2 aghi hanno prodotto 3 o 4 progenie fertile. Il numero medio di mosche fertili per ago è stato di 1,1 ± 0,2. Sulla base di questi dati, con una confidenza del 95%, 11 aghi sono sufficienti per produrre 6 o più progenie, in cui è probabile che siano inclusi almeno una femmina e un maschio.
Negli esperimenti di cui sopra, abbiamo utilizzato embrioni che esprimono l'mRNA ovo-A in PGC (nanos>ovo-A, OvoA_OE embrioni) come ospite agagametico. Su 669 femmine F1 e 720 maschi F1 prodotti da coppie nanos>ovo-A trapiantate, non c'era alcun escaper derivato dalle PGC ospiti. Diversi mutanti di Oskar (osk) sono anche sensibili alla temperatura agagametico20,21. Poiché non è più disponibile un mutante osk con un'elevata vitalità omozigote e il fenotipo agagametico, abbiamo ricreato il mutante missenso osk[8] 20 mediante editing genomico assistito da CRISPR/Cas9. Questi moscerini erano completamente agametici (0 fuggi su 230 femmine e 192 maschi) a 25 °C, ma alcuni fuggivano a 23 °C (1 su 248 femmine e 1 su 290 maschi). nanos>ovo-A sono quindi raccomandati come embrioni ospiti agagametici. Sia UASp-ovo-A che nanos-Gal4 stock 13 saranno presto disponibili presso il KYOTO Drosophila Stock Center.

Figura 1: Attrezzatura necessaria. (A) Un sistema di micromanipolatori per raccogliere e trapiantare cellule. i) microscopio invertito, ii) micromanipolatore meccanico, iii) siringa, iv) supporto capillare, v) rubinetto a tre vie, vi) umidificatore e vii) stereomicroscopio. (B) Una siringa. (C) Un rubinetto a tre vie e tubi in silicone collegano una siringa e un supporto capillare. (D) Un ago e un supporto capillare sono collegati a un micromanipolatore. (E) Una coppetta per la raccolta degli embrioni con una piastra per la raccolta degli embrioni (6 cm di diametro, 7,7 cm di altezza). (F) Un filtro a rete in acciaio inossidabile. (G) Un contenitore utilizzato come camera umida con un vetrino. Per mantenere l'umidità, posizionare la carta bagnata sul fondo e chiudere il coperchio. (H) Un porta-aghi con un ago per la crioconservazione. (I) Una rastrelliera per la crioconservazione e una scatola con aghi. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Un vetrino per la raccolta di PGC e un ago per crioconservazione. (A) Un vetrino per la raccolta di cellule germinali primordiali (PGC) rivestito di colla. Gli embrioni decoroniati sono allineati su due file e orientati con la parte anteriore a destra (il lato da manipolare) e il lato ventrale rivolto verso l'alto. Viene apposta una cornice per la piscina embrionale, vengono depositate due gocce di soluzione di agenti crioprotettivi (CPA) e la piscina viene riempita con olio di silicone. (B) Un ago dovrebbe contenere la minor quantità possibile di tuorlo e altri contaminanti. I PGC sono inseriti tra due strati di olio siliconico quando crioconservati in azoto liquido. Fare clic qui per visualizzare una versione più grande di questa figura.
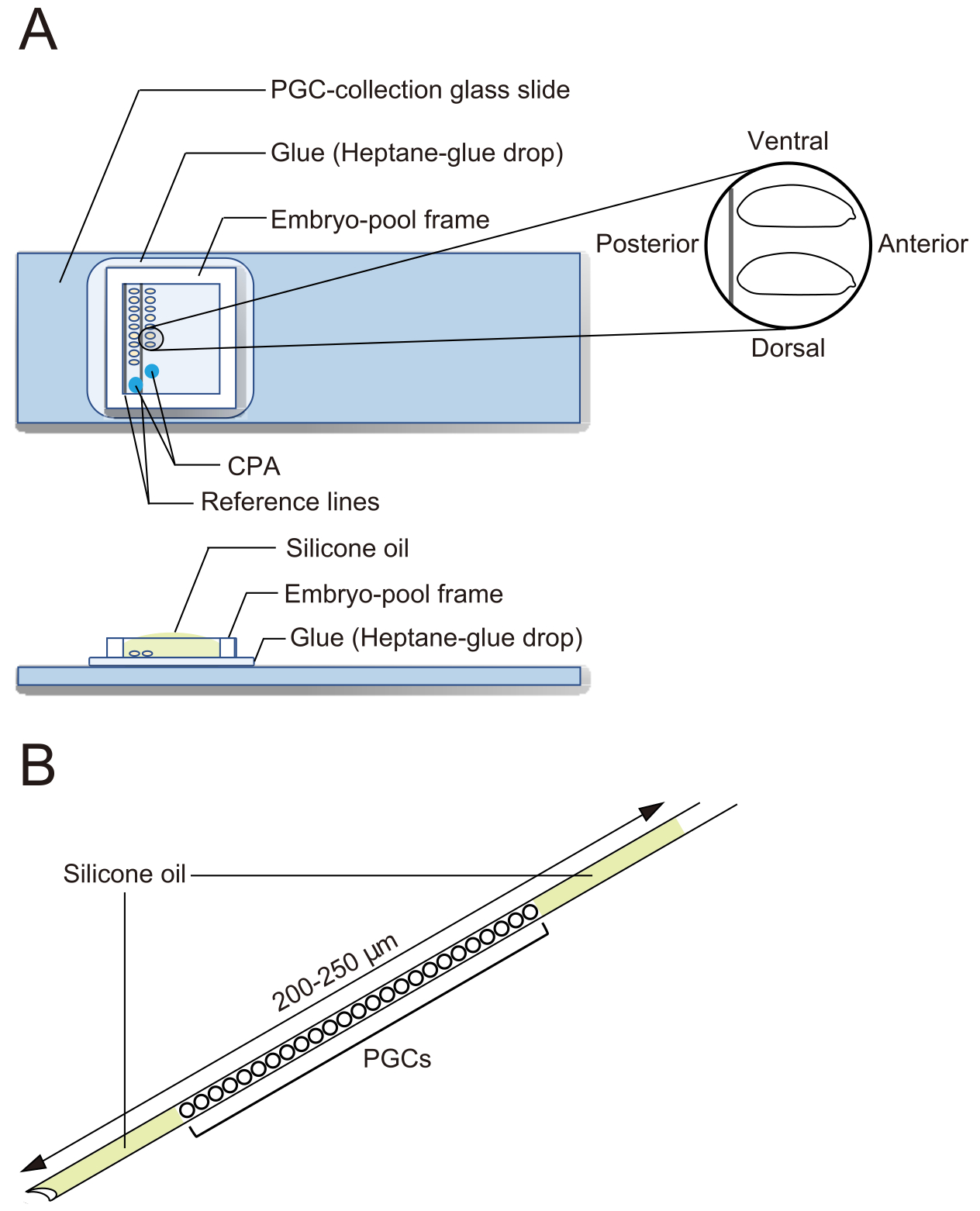{kind=link}

Figura 3: Realizzazione dell'ago. Metodo di lucidatura della punta in tre fasi per realizzare un ago con una dimensione del foro appropriata e una punta affilata. Fare clic qui per visualizzare una versione più grande di questa figura.
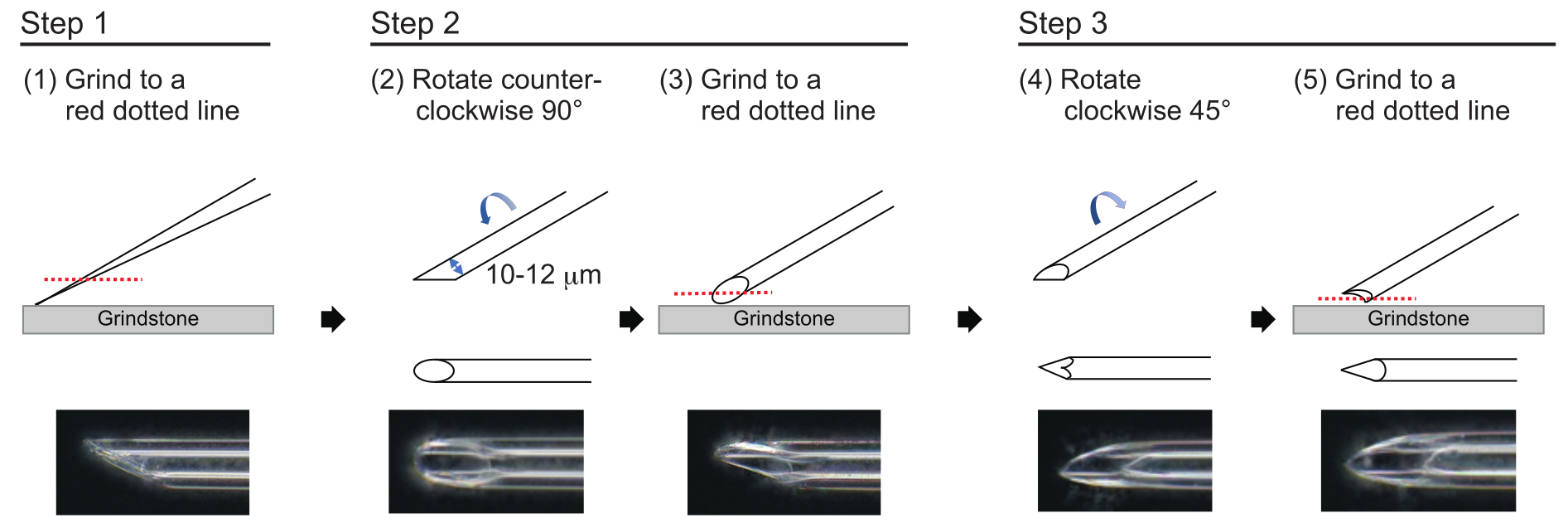{kind=link}

Figura 4: Schema di raccolta degli embrioni. Dopo due pre-ritiri, di solito raccogliamo tre o quattro volte al giorno. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Allineamento dell'embrione ospite. Allineamento degli embrioni ospiti su un vetrino. Fare clic qui per visualizzare una versione più grande di questa figura.
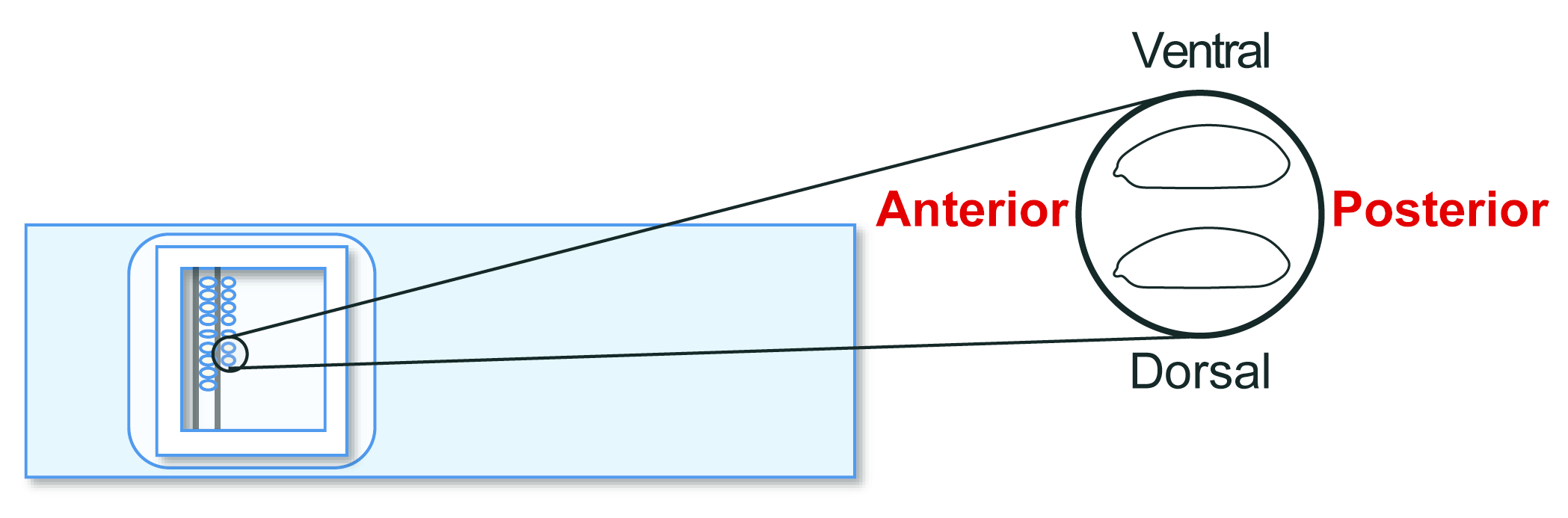{kind=link}

Figura 6: Una panoramica del metodo di crioconservazione PGC. Una panoramica di tutti i passaggi seguiti per effettuare la crioconservazione delle cellule germinali primordiali (PGC). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Umidità dell'ambiente | |||
| < 30% | ~ 30% | > 30% | |
| Allineare gli embrioni ospiti (~20 minuti) | Utilizzare un umidificatore per 2 - 10 minuti | Utilizzare un umidificatore a intermittenza per 1 minuto | Non utilizzare un umidificatore |
| Scongelare le PGC donatrici | Non applicabile | Non applicabile | Non applicabile |
| PGC essiccati all'aria | Omettere questo passaggio | Omettere questo passaggio | 5 minuti |
| Applicare l'olio di silicone | Non applicabile | Non applicabile | Non applicabile |
| PGC per trapianti | Non applicabile | Non applicabile | Non applicabile |
| Tutti questi passaggi dovrebbero essere completati in 50 minuti. | |||
Tabella 1: Essiccazione degli embrioni durante l'allineamento embrionale e lo scongelamento delle PGC.
| Ceppo del donatore | Periodo di crioconservazione | Numero di embrioni trapiantati (A) | Numero di larve schiuse (B) (schiusa, B/A) | Numero di adulti chiusi (C) (vitalità da uovo ad adulto, C/A) | Numero di adulti fertili (D) (frequenza di mosche fertili, D/C) |
| Visualizzazione del materiale M17 | 8 - 30 giorni | 134 | 108 (80.6%) | 57 (42.5%) | 20 (35.1%) |
| Visualizzazione del materiale M17 | 31 - 150 giorni | 74 | 60 (81.1%) | 30 (40.5%) | 8 (26.7%) |
| M17: imbardata; TM6B, P{Dfd-GMR-nvYFP}4, Sb[1] Tb[1] ca[1]/ Pri[1] | |||||
Tabella 2: Efficienza del trapianto di PGC crioconservato. Questa tabella è stata modificata da13. Tutti i dati provengono da ospiti agagametici.
Discussione
Un fattore critico per il successo nella crioconservazione e nella rianimazione delle PGC è l'utilizzo di embrioni buoni. Le femmine giovani (ad esempio, da 3 a 5 giorni) dovrebbero essere utilizzate per la raccolta degli embrioni. Sia gli embrioni donati che quelli ospiti vengono valutati mediante ispezione microscopica e vengono utilizzati solo quelli allo stadio di blastoderma (stadio 5)12. Per la raccolta di PGC, di solito allineiamo circa 40 embrioni donati in un periodo di 20 minuti e raccogliamo PGC da circa 30 embrioni nella fase iniziale 5; Gli embrioni più vecchi e difettosi non vengono utilizzati. Dopo la crioconservazione e lo scongelamento, i PGC dovrebbero mantenere la loro forma; I PGC si rompono in una conservazione infruttuosa. Anche gli embrioni ospiti dovrebbero essere allo stadio 5 e avere una pressione interna moderata; Gli embrioni dovrebbero tornare lentamente alla loro forma originale dopo una leggera sollecitazione. Gli embrioni troppo essiccati e non sufficientemente essiccati non si svilupperanno normalmente dopo il trapianto. Poiché il trapianto eterosessuale di PGC non riesce a produrre gameti in Drosophila 5,10, il trapianto di PGC da embrioni di donatori multipli in embrioni ospiti ha maggiori probabilità di produrre adulti fertili. A tal fine, di solito raccogliamo PGC da circa 30 embrioni per ago.
Come crioprotettori, abbiamo provato glicole etilenico, dimetilsolfossido e glicerolo insieme al saccarosio a varie concentrazioni. Abbiamo determinato che l'EBR contenente il 20% di glicole etilenico e 1 M di saccarosio è il migliore13; tuttavia, l'uso di diversi crioprotettori può migliorare la conservazione delle PGC22.
Questo metodo di crioconservazione richiede competenze specializzate nella gestione delle PGC e sono necessarie circa 6 settimane di formazione per raccogliere e trapiantare comodamente le PGC. Per valutare e migliorare la competenza delle abilità, questo può essere suddiviso in sei fasi di addestramento: 1) allineamento degli embrioni su un vetrino, 2) controllo di un manipolatore, 3) trapianto di PGC da un embrione in un altro embrione senza crioconservazione, 4) trapianto di PGC da 10 o più embrioni in 5-10 embrioni, 5) trapianto di PGC dopo l'applicazione di CPA e 6) trapianto di PGC dopo il congelamento-scongelamento. Ogni passaggio può richiedere 1 settimana. Gli obiettivi a breve termine della fase 3 sono un tasso di schiusa del 40%, una vitalità da embrione ad adulto del 10%-20% e una frequenza di mosche fertili del 20%.
La crioconservazione PGC richiede strumentazione costosa e personale altamente qualificato. Pertanto, questo metodo potrebbe non essere adottato da molti laboratori. Tuttavia, l'attuale metodo PGC presenta diversi aspetti importanti. In primo luogo, le PGC sono molto più piccole degli embrioni e sono molto permeabili ai crioprotettori. Al contrario, la permeabilità crioprotettiva è fortemente limitata dagli strati cerosi degli embrioni di Drosophila, che è il problema più grave nella crioconservazione degli embrioni. In effetti, studi precedenti hanno fatto grandi sforzi per trovare una finestra temporale in cui gli embrioni hanno un alto tasso di sopravvivenza e uno strato di cera più sottile. Il secondo riguarda lo sviluppo e la variazione morfologica tra i ceppi. Le PGC vengono raccolte dagli embrioni in stadio 5 (2 ore e 30 minuti e 20 minuti dopo la deposizione delle uova), mentre la crioconservazione degli embrioni viene eseguita sugli embrioni in stadio 16 (14-22 ore dopo la deposizione delle uova). Gli embrioni sono, quindi, molto più vecchi e mostrano una variazione di ceppo molto maggiore nella finestra temporale ottimale per la crioconservazione rispetto alla crioconservazione PGC. In effetti, la frequenza degli ospiti che producevano progenie derivata da donatori non variava tra i cinque ceppi studiati da Asaoka et al.13, sebbene gli ospiti non fossero agametici. Inoltre, le PGC hanno il potenziale per essere utilizzate in applicazioni di ingegneria genetica, come l'editing del genoma 14,15,16.
Divulgazioni
Gli autori non hanno conflitti di interesse da dichiarare.
Riconoscimenti
Ringraziamo il KYOTO Drosophila Stock Center per i ceppi di mosca. Ringraziamo anche la signora Wanda Miyata per l'editing in lingua inglese del manoscritto e il Dr. Jeremy Allen di Edanz (https://jp.edanz.com/ac) per aver curato una bozza di questo manoscritto. Questo lavoro è stato sostenuto da sovvenzioni (JP16km0210072, JP17km0210146, JP18km0210146) dall'Agenzia giapponese per la ricerca e lo sviluppo medico (AMED) a T.T.-S.-K., sovvenzioni (JP16km0210073, JP17km0210147, JP18km0210145) da AMED a S.K., una sovvenzione (JP20km0210172) da AMED a T.T.-S.-K. e S.K., una sovvenzione per la ricerca scientifica (C) (JP19K06780) della Japan Society for the Promotion of Science (JSPS) a T.T.-S.-K., e una sovvenzione per la ricerca scientifica in aree innovative (JP18H05552) da JSPS a S.K.
Materiali
| Name | Company | Catalog Number | Comments |
| Acetic acid | FUJIFILM Wako Pure Chemical Corporation | 017-00256 | For embryo collection |
| Agar powder | FUJIFILM Wako Pure Chemical Corporation | 010-08725 | For embryo collection |
| Calcium chloride | FUJIFILM Wako Pure Chemical Corporation | 038-24985 | For EBR solution |
| Capillary | Sutter Instrument | B100-75-10-PT | BOROSILICATE GLASS; O.D: 1.0mm, I.D: 0.75mm , length: 10cm, 225Pcs |
| Capillary holder | Eppendorf | 5196 081.005 | Capillary holder 4; for micromanipulation |
| Chromic acid mixture | FUJIFILM Wako Pure Chemical Corporation | 037-05415 | For needle washing |
| CPA solution | 1x EBR containing 20% ethylene glycol and 1M sucrose | ||
| Double-sided tape | 3M | Scotch w-12 | For glue extracting |
| Ephrussi–Beadle Ringer solution (EBR) | 130 mM NaCl, 5 mM KCl, 2 mM CaCl2, and 10 mM Hepes at pH 6.9 | ||
| Ethanol (99.5) | FUJIFILM Wako Pure Chemical Corporation | 057-00451 | For embryo collection |
| Ethylene glycol | FUJIFILM Wako Pure Chemical Corporation | 054-00983 | For CPA solution |
| Falcon 50 mm x 9 mm bacteriological petri dish | Corning Inc. | 351006 | For embryo collection |
| Forceps | Vigor | Type5 Titan | For embryo handling |
| Grape juice | Asahi Soft Drinks Co., LTD. | Welch's Grape 100 | For embryo collection |
| Grape juice agar plate | 50% grape juice, 2% agar, 1% ethanol, 1% acetic acid | ||
| Heptane | FUJIFILM Wako Pure Chemical Corporation | 084-08105 | For glue extracting |
| Humidifier | APIX INTERNATIONAL CO., LTD. | FSWD2201-WH | For embryo preparation |
| Inverted microscope | Leica Microsystems GmbH | Leica DM IL LED | For micromanipulation |
| Luer-lock glass syringe | Tokyo Garasu Kikai Co., Ltd. | 0550 14 71 08 | Coat a plunger with silicon oil (FL-100-450CS);for micromanipulation |
| Mechanical micromanipulator | Leica Microsystems GmbH | For micromanipulation | |
| Micro slide glass | Matsunami Glass Ind., Ltd. | S-2441 | For embryo aligning |
| Microgrinder | NARISHIGE Group | Custom order | EG-401-S combined EG-401 and MF2 (with ocular lens MF2-LE15 ); for needle preparation |
| Microscope camera | Leica Microsystems GmbH | Leica MC170 HD | For micromanipulation |
| Needle holder | Merck KGaA | Eppendorf TransferTip (ES) | For cryopreservation |
| Potassium chloride | Nacalai Tesque, Inc. | 28514-75 | For EBR solution |
| Puller | NARISHIGE Group | PN-31 | For needle preparation; the heater level is set to 85.0-98.4, the magnet main level to 57.8, and the magnet sub level to 45.0. |
| PVC adhesive tape for electric insulation | Nitto Denko Corporation | J2515 | For embryo-pool frame |
| Silicon oil | Shin-Etsu Chemical, Co, Ltd. | FL-100-450CS | For embryo handling |
| Sodium chloride | Nacalai Tesque, Inc. | 31320-05 | For EBR solution |
| Sodium hypochlorite solution | FUJIFILM Wako Pure Chemical Corporation | 197-02206 | Undiluted and freshly prepared; for embryo breaching |
| Sucrose | Nacalai Tesque, Inc. | 30404-45 | For CPA solution |
Riferimenti
- Brüschweiler, W., Gehring, W. A method for freezing living ovaries of Drosophila melanogaster larvae and its application to the storage of mutant stocks. Experientia. 29, 134-135 (1973).
- Steponkus, P. L., et al. Cryopreservation of Drosophila melanogaster embryos. Nature. 345, 170-172 (1990).
- Mazur, P., Cole, K. W., Hall, J. W., Schreuders, P. D., Mahowald, A. P. Cryobiological preservation of Drosophila embryos. Science. 258 (5090), 1932-1935 (1992).
- Zhan, L., Li, M. G., Hays, T., Bischof, J. Cryopreservation method for Drosophila melanogaster embryos. Nat Comm. 12, 2412 (2021).
- Van Deusen, E. B. Sex determination in germ line chimeras of Drosophila melanogaster. Development. 37 (1), 173-185 (1977).
- Breen, T. R., Duncan, I. M. Maternal expression of genes that regulate the bithorax complex of Drosophila melanogaster. Dev Biol. 118, 442-456 (1986).
- Schupbach, T., Wieschaus, E. Germline autonomy of maternal-effect mutations altering the embryonic body pattern of Drosophila. Dev Biol. 113, 443-448 (1986).
- Irish, V., Lehmann, R., Akam, M. The Drosophila posterior-group gene nanos functions by repressing hunchback activity. Nature. 338, 646-648 (1989).
- Hülskamp, M., Schröder, C., Pfeifle, C., Jäckle, H., Tautz, D. Posterior segmentation of the Drosophila embryo in the absence of a maternal posterior organizer gene. Nature. 338, 629-632 (1989).
- Steinmann-Zwicky, M., Schmid, H., Nöthiger, R. Cell-autonomous and inductive signals can determine the sex of the germ line of Drosophila by regulating the gene Sxl. Cell. 57 (1), 157-166 (1989).
- Stein, D., Roth, S., Vogelsang, E., Nüsslein-Volhard, C. The polarity of the dorsoventral axis in the drosophila embryo is defined by an extracellular signal. Cell. 65 (5), 725-735 (1991).
- Kobayashi, S., Yamada, M., Asaoka, M., Kitamura, T. Essential role of the posterior morphogen nanos for germline development in Drosophila. Nature. 380, 708-711 (1996).
- Asaoka, M., et al. Offspring production from cryopreserved primordial germ cells in Drosophila. Comm Biol. 4 (1), 1159 (2021).
- Blitz, I. L., Fish, M. B., Cho, K. W. Y. Leapfrogging: primordial germ cell transplantation permits recovery of CRISPR/Cas9-induced mutations in essential genes. Development. 143 (15), 2868-2875 (2016).
- Koslová, A., et al. Precise CRISPR/Cas9 editing of the NHE1 gene renders chickens resistant to the J subgroup of avian leukosis virus. Proc Natl Acad Sci U S A. 117 (4), 2108-2112 (2020).
- Zhang, F. Efficient generation of zebrafish maternal-zygotic mutants through transplantation of ectopically induced and Cas9/gRNA targeted primordial germ cells. J Genet Genom. 47 (1), 37-47 (2020).
- Campos-Ortega, J. A., Hartenstein, V. Stages of Drosophila Embryogenesis. The Embryonic Development of Drosophila. , (1997).
- Manning, A. A sperm factor affecting the receptivity of Drosophila melanogaster females. Nature. 194, 252-253 (1962).
- Kubli, E. Sex-peptides: seminal peptides of the Drosophila male. Cell Mol Life Sci. 60, 1689-1704 (2003).
- Lehmann, R., Nüsslein-Volhard, C. Abdominal segmentation, pole cell formation, and embryonic polarity require the localized activity of oskar, a maternal gene in drosophila. Cell. 47 (1), 141-152 (1986).
- Kiger, A. A., Gigliotti, S., Fuller, M. T. Developmental genetics of the essential Drosophila Nucleoporin nup154: allelic differences due to an outward-directed promoter in the P-element 3′ end. Genetics. 153 (2), 799-812 (1999).
- Rienzi, L. F., et al. Perspectives in gamete and embryo cryopreservation. Semin Reprod Med. 36 (5), 253-264 (2018).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati