
Method Article
在 果蝇卵巢成人线粒体DNA复制的原位标记埃杜染色
摘要
Drosophila oogenesis continues to be exceptionally useful in the study of mitochondrial proliferation and inheritance. This manuscript describes a detailed protocol used to label the replicating mitochondrial DNA (mtDNA) in Drosophila adult ovaries with 5-ethynyl-2´-deoxyuridine (EdU), which facilitates uncovering mechanisms associated with mitochondrial inheritance that were previously debatable.
摘要
The mitochondrial genome is inherited exclusively through the maternal line. Understanding of how the mitochondrion and its genome are proliferated and transmitted from one generation to the next through the female oocyte is of fundamental importance. Because of the genetic tractability, and the elegant, ordered simplicity by which oocyte development proceeds, Drosophila oogenesis has become an invaluable system for mitochondrial study. An EdU (5-ethynyl-2´-deoxyuridine) labeling method was utilized to detect mitochondrial DNA (mtDNA) replication in Drosophila ovaries. This method is superior to the BrdU (5-bromo-2'-deoxyuridine) labeling method in that it allows for good structural preservation and efficient fluorescent dye penetration of whole-mount tissues.
Here we describe a detailed protocol for labeling replicating mitochondrial DNA in Drosophila adult ovaries with EdU. Some technical solutions are offered to improve the viability of the ovaries, maintain their health during preparation, and ensure high-quality imaging. Visualization of newly synthesized mtDNA in the ovaries not only reveals the striking temporal and spatial pattern of mtDNA replication through oogenesis, but also allows for simple quantification of mtDNA replication under various genetic and pharmacological perturbations.
引言
除核基因组中,每个的真核细胞还包含数千个在线粒体基质小环状DNA的拷贝。而线粒体DNA(mtDNA的)编码的电子传递链的必需亚基,大多数线粒体蛋白质,包括所有的因素对复制和线粒体DNA的转录是由核基因组编码。相反,下面的继承孟德尔定律的核基因组,动物线粒体基因组是通过母系遗传独占。因此,理解的mtDNA是如何增殖和从一代传递到下一个通过女性的卵母细胞是至关重要的。然而,有过线粒体DNA复制是如何在阴生殖调节继续争论。此外,广为接受的mtDNA瓶颈理论线粒体传输表明线粒体DNA中的原始生殖细胞的种群二次采样到一个相对较小的数都灵3G发展1 2。这也意味着,线粒体DNA的复制是时间和卵子发生过程监管空间。因此,种系发育过程中原位检测线粒体DNA复制的将促进线粒体遗传的机制的理解。
果蝇卵子提供了一个听话的基因系统来研究线粒体DNA复制和传播。在每两个果蝇卵巢,有卵腔室16-20独立串称为卵巢管3,其是产蛋的功能单元(参见图1A)。每个卵巢管包含卵子发生,其中,前尖端是由一个称为germarium结构的渐进线性组织。该germarium被进一步分成包含在不同发育阶段的生殖细胞的四个区域。在区域1,生殖系干细胞通过不对称分裂产生称为为c子细胞ystoblasts。区域2a包含已经完成了他们最后的分工cystoblasts。 Cystoblasts经过四轮分工生产16电池组。 16细胞仍然被称为环运河桥梁细胞质相互连接。仅其中的一个单元承诺分化为卵母细胞,而其他15发展为多倍体护士细胞。必不可少的胞质结构,被称为fusome,建议用环管的形成,以促进,确定囊肿极性和护士细胞-卵母细胞的相互作用4,5。作为孢囊向区域2b移动,囊肿结构跨越germarium的整个宽度,并变得更加与毛囊细胞相关联。该germarium结构结束与包含第一个初露头角的卵室区3。随后,将蛋室在germarium,它通过卵巢管在14形态不同阶段进行的后端装配。卵母细胞的生长依赖于滋养细胞,WHICH转运蛋白,mRNA和经由环管进入卵母细胞内膜结构( 例如 ,高尔基)。期间果蝇卵子发生,人们发现,每个16细胞囊肿内的线粒体的一个级分与fusome相关联,通过环管移动,并且被输送到单个卵母细胞在称为巴尔比亚尼体6大的质量。这种现象提出了有助于通过女性卵巢线粒体遗传的质量控制。
线粒体DNA合成的检测依赖于标记的DNA前体掺入到细胞DNA中。传统上,胸苷5-溴-2'-脱氧尿苷的核苷类似物(BrdU)标记用于标记在组织培养细胞7,8- mtDNA的复制。然而,基于抗体的BrdU标记显示出若干局限性,特别是对于整个贴装组织染色。 BrdU标记的一个主要缺点是,它需要的DNA变性,以暴露所述的BrdU表位,因此,它可以被抗BrdU抗体进行检测。检体已苛刻的变性条件如化学品( 如,盐酸或甲醇和乙酸的混合物),热或用DNase消化,这可能会破坏试样的结构和下列染色步骤9复杂化下处理, 10。
这里,另一种胸苷类似物5-乙炔基-2'-脱氧尿苷(EDU)被用来标记在果蝇成年卵巢复制线粒体DNA。这种方法是快速,高度敏感。的EdU DNA复制过程中容易地结合到细胞DNA中。下面检测是基于一个"点击"反应中,铜(Ⅰ)催化的末端炔基和荧光叠氮化物10之间的共价反应。由于反应不需要检体的变性,它允许良好的结构保存。此外,染料的大小zide只有1/500的一种抗体分子10,使整体安装组织的快速,高效的渗透。我们已经用这种方法果蝇卵子发生过程中检测线粒体DNA的复制,发现在果蝇卵巢11 germarium区一个惊人的空间格局,这导致我们提出了一个相关的复制线粒体DNA选择性的继承机制。我们在座的果蝇卵巢线粒体DNA复制的标签的EdU了详细的方案。为了证明该协议的应用,我们还测试了线粒体DNA突变体果蝇的线粒体DNA复制( 公吨:COI T300I)11,以及与线粒体解偶联剂,其消散线粒体膜电位和潜在破坏线粒体DNA复制的多样的治疗。
研究方案
1.组织收集和解剖
- 在含干酵母每个小瓶,文化10只成年雌性苍蝇与10名男性2-3天。
注意:保持良好的喂养雌性果蝇会提高整体素质和卵巢的生产产量和促进清扫。 - 麻醉雌性蝇上的二氧化碳(CO 2)的飞垫。
- 下的立体镜,放置逆转录介质上的解剖垫(补充有10%胎牛血清(FBS)的施耐德的果蝇介质)数滴。所有进行解剖过程中的媒介,以保持组织健康地活着。
- 抓住一个女养肥飞在其下端胸部精尖鼻钳。使用另一套镊子,直到腹部组织暴露在苍蝇的极端后轻轻拉扯。从分离其他组织( 如胆)的两个卵巢。
注:每个卵巢显示一个不透明的结构,是撰写16-20连接卵巢管Ð。 - 为了提高试剂的渗透,通过拉开他们,并传递钳提示在每间卵巢管几次打开卵巢。保留卵巢内连接时下列程序,以尽量减少组织损失的卵巢管。
- 马上转移卵巢含有500微升施耐德的果蝇培养基含10%FBS的1.5ml微量管中。
- 重复清扫,收集每个离心管中10-15卵巢。
2.标签的EdU
- 吸在每个管的介质,并用500μl含有7微米阿非迪霉素10%FBS的施耐德的果蝇介质的替换。
注意:阿非迪霉素用于通过抑制DNA聚合酶α,而不影响线粒体DNA复制7,12到方框核DNA的合成。它是可以提高阿非迪霉素浓度最多到70微米,以达到更好的抑制作用。 3-30毫阿非迪霉素的DMSO储备溶液可以储存在黑暗中在-20℃下长达6个星期。 - 保持卵巢健康,确保它们下溶液浸渍,同时改变介质。通过步2.6使用带有10%FBS施耐德的果蝇网上平台。
- 孵育卵巢在RT上轻轻旋转台式摇杆3小时。
- 进行药物治疗, 例如,线粒体解偶联剂羰氰化物4-三氟甲氧基苯腙(FCCP),添加药物的适当的浓度( 例如,10μMFCCP)到培养基中之后阿非迪霉素治疗的2小时。继续孵育另外1小时。
- 有或没有药物删除含有阿非迪霉素网上平台。简要地冲洗中等卵巢(阿非迪霉素是没有必要的)的两倍。
- 加入1ml含有10μM的EdU和7微米阿非迪霉素,继续incubating的室温2小时。储存在-20℃下的10毫米的EdU原液(在DMSO中)。
- 删除含有的EdU和阿非迪霉素的媒介。用培养基(无阿非迪霉素)洗涤两次,每次3分钟。
3.组织固定和透
- 制备在磷酸盐缓冲盐水(PBS,pH7.4)中4%的多聚甲醛溶液中。
注:我们使用存储在预刻划安瓿市售的多聚甲醛。打开新的安瓿并在使用前稀释至4%。
注意:甲醛是有毒的;它应与皮肤和眼睛的保护在通风橱处理。 - 用4%多聚甲醛的卵巢在室温20分钟,轻轻转动。
- 除去固定剂并且在1ml的PBS中的3%BSA的清洗卵巢两次,每次5分钟。
- 通透的组织,取出洗净解决方案,并在PBS加入1毫升中含0.5%的Triton X-100。孵育在室温20分钟。
- 除去透化溶液并在1ml洗两次的在PBS中的3%BSA中。
4.检测的EdU
注意:的EdU检测是基于"点击"化学,铜(Ⅰ)催化[3 + 2]环加成13,这增加了荧光叠氮化物向的EdU的末端炔基,并且荧光分子进行后续检测。由于铜(I)中,这是很容易氧化成非催化铜(II)物种,需要以催化反应,建议减少铜(II)硫酸盐原位得到的Cu(I)。即,铜(II)硫酸盐是在还原剂的存在下使用,例如抗坏血酸(这里的"缓冲添加剂"),以生成铜(I)。
- 在实验之前,准备的Alexa Fluor叠氮化物的工作溶液(的Alexa Fluor 488叠氮化物,的Alexa Fluor 555叠氮化物或其他人,这取决于优选的荧光团,其中被命名为"染料叠氮化物"以下),埃杜反应缓冲液和的EdU缓冲添加剂根据制造商的说明。
- 每瓶70微升DMSO染料叠氮化物。储存在-20℃下长达1年的工作溶液中。
- 通过用去离子水稀释10倍的EdU反应缓冲液制备新鲜的EdU反应缓冲液。
注意事项:使用后,存放在4℃的任何剩余1X解决方案。 1倍溶液为长达6个月是稳定的。 - 通过在2ml去离子水将粉末充分溶解使的EdU缓冲添加剂的10倍的原液。
注意:此原液是长达1年,在-20℃的稳定。如果该溶液开发了棕褐色,它已经降低,并应被丢弃。
- 准备这些EDU反应鸡尾酒新鲜每次和准备的15分钟内使用。实现可重复的结果,确保反应组分保持在相同的比率。
- 使新鲜的1X的EdU稀释在去离子水中的10倍的原液(在步骤4.1.3的方法制备)1:10缓冲添加剂溶液。使用这个Solution在同一天。
- 由下列成分以组合制备1毫升的EdU反应鸡尾酒:860微升1×的EdU反应缓冲液,40微升硫酸铜 ,2.5微升染料叠氮化物,将100μl1×的EdU缓冲添加剂。
注意:该成分以确保最佳性能增加是重要的。
- 用3%BSA的除去PBS中,加入0.5毫升的EdU反应鸡尾酒到每个管中。孵育在室温下,轻轻旋转30分钟。保持避光样本。
- 取出的EdU反应鸡尾酒并用1ml的PBS中的3%BSA的洗一次。
- 用1毫升的PBS洗一次样品。
5.抗体标记
注:的EdU染色后进行抗体标记。如果没有额外的染色需要,可以进行直接安装和成像。重要的是在样品中所有以下程序避光。
- 除去洗涤溶液。在1毫升含有0.2%BSA和0.1%的Triton X-100的PBS 30分钟阻断溶液阻断卵巢。
- 除去封闭溶液,并用封闭液稀释的第一抗体( 例如,小鼠ATP合酶亚基α抗体,1:1000稀释)代替。在4°CO / N孵育在黑暗中。
- 用1ml封闭溶液3次,每次10分钟的洗样品。为了最大限度地减少的背景下,洗两次,每30分钟。除去封闭溶液。
- 在RT 2小时:(200稀释例如,山羊抗小鼠的Alexa Fluor 568二级抗体,1)与封闭溶液稀释的二级抗体孵育。
注:使用不同的颜色比一个耦合到的EdU。 - 重复步骤5.3执行的洗涤步骤。
- 用1毫升的PBS冲洗一次,以去除去污剂。
6.安装和影像
- 小心取出所有的PBS和IMMediately覆盖用50μl固定培养基(有或没有DAPI)卵巢。
注意:使用而卵巢管被彼此分离不变硬的安装平台。卵巢可以存储在安装介质在4℃下进行长达一周。 - 切枪头的末端,用它小心卵巢转移到一个载玻片。
- 完全独立的每个卵巢管的体视显微镜下精尖鼻钳。在后端和舞台14或成熟的卵子取出室连接的组织。年轻的蛋商会是在前端和透明。
- 对准镊子卵腔室,使它们不相互重叠。
- 慢慢降低对样品的顶部上的#1.5玻璃盖玻片。允许安装介质聚合在室温数小时。用透明的指甲油密封。
- 图像滑动第二天,或储存于4℃之前成像。
- 共聚焦MI下可视化具有63X油浸物镜croscope。捕获三维的z堆栈的图像。
注意:有较强的教育栏目后期蛋商会和背景荧光信号在上皮鞘。当检查在特定germarium的EdU标记,应避免由重叠或靠近其他组织或后期蛋商会germariums。
结果
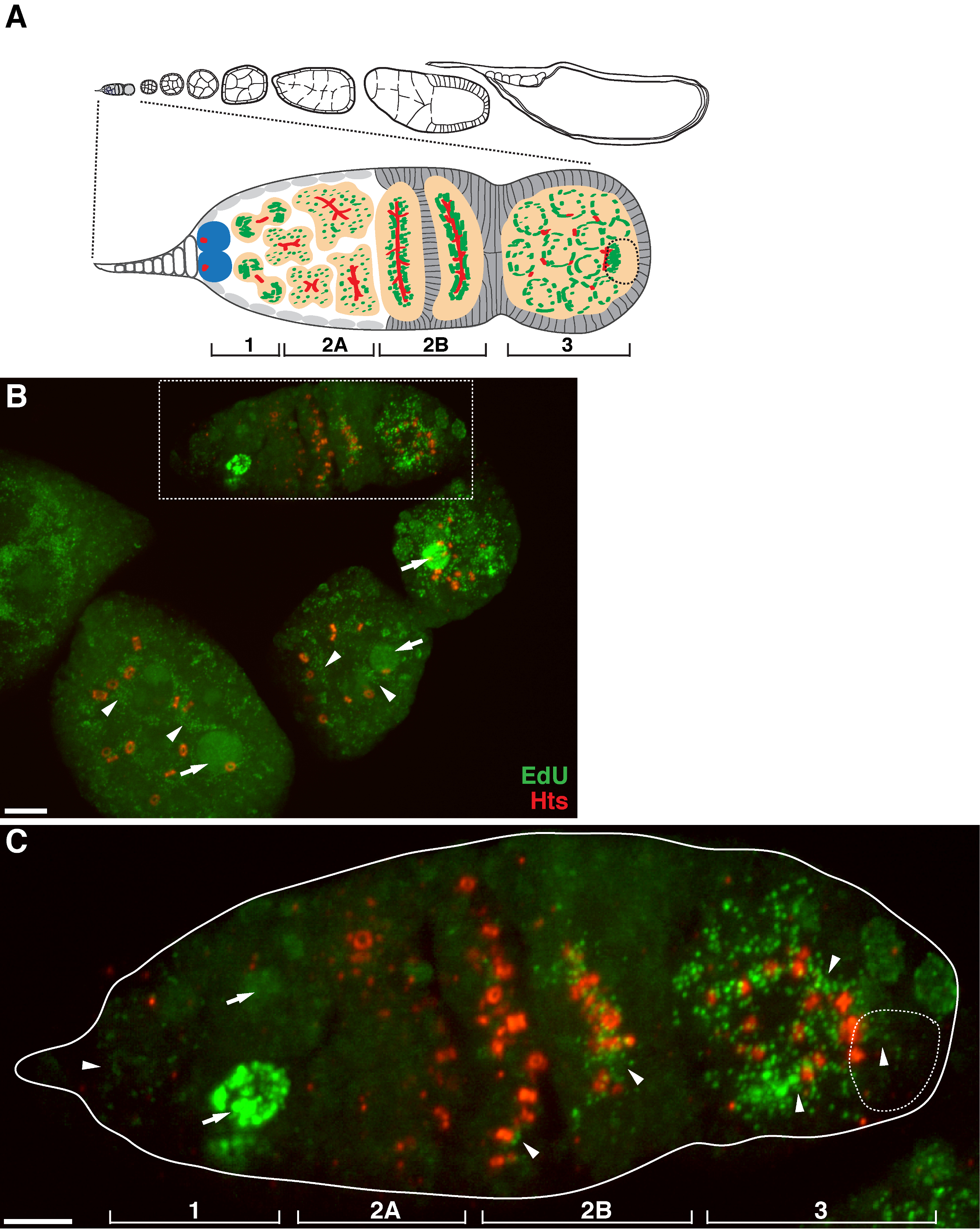
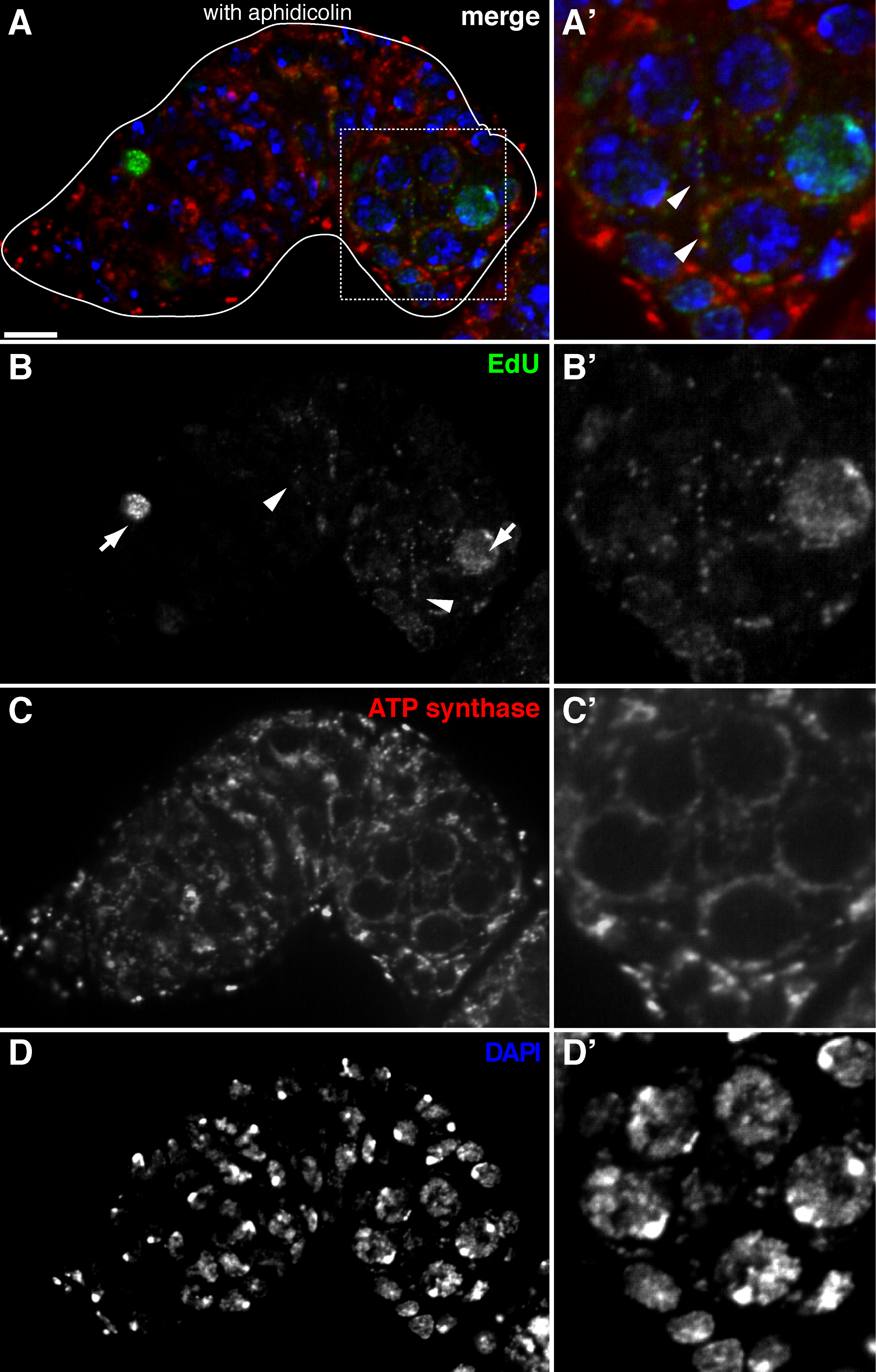
上述协议允许与线粒体( 图1B-C)相关联的点状结构,这表明果蝇卵子中的线粒体DNA复制的可视化。所述的EdU泪点与由染色ATP合酶α亚基( 图2)标记的线粒体本地化。所观察到的信号是在用溴化乙锭11,对线粒体DNA复制14抑制剂治疗的卵巢缺席,验证这些泪点确实标签复制mtDNA.Aphidicolin用于抑制核DNA染色,而不会影响线粒体DNA复制( 图2)。无阿非迪霉素治疗,紧张的EdU信号标记细胞核,并几乎没有观察到mtDNA泪点( 图3)。然而,在阿非迪霉素的存在下,核掺入显着降低,并观察到与线粒体有关的许多泪点。
有一个在后germarium蛋室( 图1B)线粒体DNA的复制水平高。然而,值得注意的是,线粒体DNA复制显示在germarium的空间格局。如通过的EdU泪点的数字表示,有线粒体DNA复制在germarium的区域1中等水平,但几乎没有的EdU掺入区域2A( 图1C)。作为囊肿在germarium向下移动到区域2B,线粒体DNA复制恢复,并在区域2B的后部囊肿的EdU泪点的数目比在区域2A( 图1C)高得多。具体而言,密集的EdU掺入集中在环运河和fusome结构,由胡力邵泰(HTS)蛋白染色。线粒体DNA保持在较高的水平在germarium的区域3复制( 图1C)
为了证明特异性的协会集成电路的基因或与线粒体增殖治疗, 果蝇卵巢可经受基因操作或药物治疗。我们治疗卵巢用不同浓度FCCP,一个典型的线粒体protonophore,消散线粒体膜电位。作为对照,DMSO对线粒体DNA的复制( 图4A)没有影响。高剂量FCCP(10μM)几乎完全耗尽整个germarium( 图4D)线粒体DNA的复制。尽管如此,FCCP(2或5微米)的低浓度对线粒体DNA复制在区域1中的区域2B和3( 图4B-C)的影响较小,但抑制复制,表明区域2B和3是线粒体破坏更敏感,或它们保持相对较慢的复制动力学。上述结果表明,线粒体DNA复制与线粒体活性相关。特别是,germarium不同地区的反应不同线粒体小鬼airment。

果蝇卵子发生过程 图1. 线粒体DNA的复制。(A) 果蝇卵巢管的图和germarium的放大图。在卵巢管示出由左到右,前至后的蛋室中,连续的发育阶段。在germarium,所述fusome(红),生殖系干细胞(蓝色),线粒体(绿),未来的卵母细胞(虚线,由定位,fusome结构和线粒体簇识别),显影囊肿(桃)和四个发育区域示。 (B)埃杜标记野生型卵巢管和抗体对HTS-RC,为环运河和fusome标记代表Z堆叠投影。在阿非迪霉素的存在下,所述的EdU被掺入线粒体DNA(箭头)和细胞核(箭头)。比例尺,10微米。( 三 ) 乙方在盒装区域概述germaria的放大图。四个发育区域表示。比例尺,5微米11。 请点击此处查看该图的放大版本。
{kind=link}

图2. 在通过的EdU掺入用阿非迪霉素治疗可视果蝇 germarium线粒体DNA复制(A) - (D)示出的EdU掺入野生型germarium的代表性共聚焦部(绿色,b)中,线粒体,标记被ATP合酶α亚基染色(红色,C),以及核,标记用DAPI染色(蓝色,D)的预孵育DNA聚合酶α抑制剂阿非迪霉素。 (A&#39 ;-D')中流露出的EdU掺入(B方框区域的放大图像(A)"),线粒体(C')和核(D')。在阿非迪霉素的存在下,核掺入(箭头)被降低和许多泪点中线粒体(箭头)内本地化。比例尺,10μm以下。这个数字已经从11修改。 请点击此处查看该图的放大版本。
{kind=link}

图3.线粒体DNA复制是在未经阿非迪霉素治疗果蝇 germarium几乎检测不到(A) - (D)示出的EdU掺入(绿色)的野生型germarium的代表性共聚焦部分,线粒体,标记由ATP合酶α亚基染色(红色),以及核,标有在不存在的DNA聚合酶α抑制剂阿非迪霉素的DAPI染色(蓝色)。 (A'-D'),(A)中方框区域的放大图像显示掺入的EdU(B'),线粒体(C')和核(D')。阿非迪霉素无,勉强被检测到强烈的EdU信号标签核(箭头),和线粒体DNA泪点。比例尺,10μm以下。这个数字已经从11修改。 请点击此处查看该图的放大版本。
{kind=link}

图4.线粒体解偶联剂损害线粒体DNA的复制。说明野生型germarium代表Z堆栈项目DMSO处理(A)或在2微米(B)中,5微米(C),10μM的EdU掺入期间(D)的浓度的线粒体解偶联剂FCCP。注意,在与2微米FCCP(B)的处理过的区域2B中的受损的EdU标记,并在这两个区域2B和3与5微米FCCP处理(在盒子中概述)。四个发育区域被示出。箭头,核DNA;箭头,线粒体DNA。比例尺,10微米。这个数字已经从11修改。 请点击此处查看该图的放大版本。
{kind=link}
讨论
的EdU标记是一种新颖的和有效的方法来检测在增殖细胞,它是基于在新合成的DNA的掺入和核苷类似物的染色DNA的合成。此方法优于在于它是更快和高度敏感的BrdU标记方法。更重要的是,它允许良好的结构保存和高效率的EdU染料渗透整个安装组织9,10。在历史上,以BrdU标记一个更好的选择,使用期间的S相学习核DNA复制的EdU标记细胞周期。阿非迪霉素是DNA聚合酶α的抑制剂,它是用于在S期12 15核DNA复制的主要聚合酶。线粒体DNA复制是通过DNA聚合酶γ,这是不敏感的阿非迪霉素处理进行。因此,与前和的EdU孵育期间阿非迪霉素治疗显著抑制的EdU掺入核DNA。这应该另外,阿非迪霉素可以是数周稳定的,如果适当地存储,并且阿非迪霉素在抑制核DNA复制的效果是在我们手中变量。用强大的核DNA标记的卵巢管和蛋商会应排除进一步的数据分析。
线粒体DNA复制可以容易形象化为在细胞质中,这也提供了一个简单的方法通过计数的EdU泪点的数量归一化到细胞质的总体积来定量线粒体DNA复制水平泪点。成像软件可以应用到自动识别在显微图像,这对大型数据集计算分析特别有用的EdU泪点。然而,应采取预防措施,因为核DNA复制的不完全抑制可以导致在染色体上不同基因座的的EdU掺入和显示为在细胞核内泪点。另外,在其他情况下,埃杜的强度掺入REPLIcating线粒体DNA可能是微弱的,而背景和噪声可能会很高。因此,对于自动图像分析的各个参数应仔细限定。它也建议,图像应经过培训的眼睛进行检查,以确保适当的EdU泪点被确定。
果蝇卵子发生期间新合成的线粒体DNA的可视化提供了一个机会来研究线粒体DNA复制如何生理或病理条件下的监管,通过开展在经受各种药理或遗传扰动的苍蝇实验。 COI T300I 11:在先前的研究中,在一个线粒体DNA突变体果蝇公吨进行的EdU掺入法。此外,为了破坏线粒体膜电位,卵巢用各种浓度的线粒体解偶联剂FCCP或2,4-二硝基苯酚(DNP)的EdU之前掺入处理。根据实验的目的和药物的特点,不同的方法可能是有效的传递采纳。对于成年果蝇,药物可以作为蒸气(例如,乙醇和可卡因)16,17或药物可以注射到腹部,在那里它迅速扩散在整个主体18。最常见的做法是,药物加入到飞食品或蔗糖/药物饱和滤纸。例如,微管组装,秋水仙碱的抑制剂,被卵巢切除19之前供给到果蝇2-3天。因此,为了评估药物递送方法,选择适当的浓度是很重要的。
为了确保在果蝇卵巢线粒体DNA复制的成功的成像,有几个关键的步骤必须认真执行。最重要的是,夹层和的EdU掺入过程中保留了卵巢的生存能力和健康是必要的(步骤1和2)。需要被加热至室温与FBS施耐德的果蝇中使用前。每个人都应该尽量减少蛋室和解剖工具或枪头之间的直接接触。卵巢应在解决方案沉浸任何时候,以避免脱水。处理不当的组织可以很容易导致晕倒或无荧光信号。在组织安装步骤,卵巢管应该从彼此分离并摊开在显微镜载玻片。确保蛋室不堆叠在彼此的顶部。我们注意到,鸡蛋室超出14期小显示掺入的EdU。此外,由于大尺寸,后期鸡蛋商会往往会造成邻近的年轻蛋商会是失焦。因此,建议在后期蛋室被丢弃。
在这里,我们提供的标签在果蝇卵巢成年线粒体DNA复制了详细的方案。此方法允许线粒体DNA复制的简单量化在各种遗传学和药理学扰动并且将成为潜在的解剖发育线粒体生物合成和线粒体DNA遗传机制非常有用。
披露声明
The authors have nothing to disclose.
致谢
We thank K. Delaney for comments on the manuscript. This work was supported by the National, Heart, Lung, and Blood Institute (NHLBI) Intramural Program.
材料
| Name | Company | Catalog Number | Comments |
| Schneider’s Drosophila medium | Invitrogen | 21720-024 | |
| FBS | Invitrogen | 10100-147 | |
| Pair of Dumont #5 Forceps | Fine Science Tools | 11252-20 | |
| Aphidicolin | Sigma | A0781 | Aliquot after dissolving in DMSO. Avoid repetitive thawing and freezing. Protect from light. May be used within 6 weeks after dissolving. |
| FCCP | Sigma | C2920 | |
| DMSO | Sigma | D2650 | |
| Paraformaldehyde, 16% EM grade | Electron Microscopy Sciences | 15710 | Formaldehyde is toxic; it should be handled in a fume hood with skin and eye protection. |
| PBS | KD Medical | RGF-3190 | |
| BSA | Sigma | A7030 | |
| Triton X-100 | Sigma | T9284 | |
| Click-iT EdU Alexa Fluor 488 Imaging Kit | Invitrogen | C10337 | EdU, CuSO4, Alexa Fluor 488 azide, EdU reaction buffer and Edu buffer additive are included |
| Mouse ATP synthase subunit α antibody, (15H4C4) | MitoSciences | Ab14748 | 1:1,000 dilution |
| Mouse Hts antibody (clone RC) | Developmental Studies Hybridoma Bank (DSHB) | hts RC | 1:1,000 dilution |
| Goat anti-mouse Alexa Fluor 568 secondary antibody | Invitrogen | A-11004 | 1:200 dilution |
| Vectashield mounting medium with DAPI | Vector Laboratories | H-1500 | |
| Glass coverslips, #1.5 22 mm x 22 mm | Fisher Scientific | 12-541-B | |
| Microscope slide | Fisher Scientific | 22-038-103 | |
| Nail polish | Elf | Many of the pigments used in nail polishes are fluorescent and leach into specimens. Only clear nail polish should be used. |
参考文献
- Taylor, R. W., Turnbull, D. M. Mitochondrial DNA mutations in human disease. Nat Rev Genet. 6 (5), 389-402 (2005).
- Wallace, D. C., Chalkia, D. Mitochondrial DNA genetics and the heteroplasmy conundrum in evolution and disease. Cold Spring Harb Perspect Biol. 5 (11), a021220 (2013).
- Spradling, A. C. . The development of Drosophila melanogaster. , (1993).
- Riechmann, V., Ephrussi, A. Axis formation during Drosophila oogenesis. Curr Opin Genet Dev. 11 (4), 374-383 (2001).
- Lin, H., Yue, L., Spradling, A. C. The Drosophila fusome, a germline-specific organelle, contains membrane skeletal proteins and functions in cyst formation. Development. 120 (4), 947-956 (1994).
- Cox, R. T., Spradling, A. C. A Balbiani body and the fusome mediate mitochondrial inheritance during Drosophila oogenesis. Development. 130 (8), 1579-1590 (2003).
- Davis, A. F., Clayton, D. A. In situ localization of mitochondrial DNA replication in intact mammalian cells. J Cell Biol. 135 (4), 883-893 (1996).
- Iborra, F. J., Kimura, H., Cook, P. R. The functional organization of mitochondrial genomes in human cells. BMC Biol. 2, (2004).
- Rakic, P. Neurogenesis in adult primate neocortex: an evaluation of the evidence. Nat Rev Neurosci. 3 (1), 65-71 (2002).
- Salic, A., Mitchison, T. J. A chemical method for fast and sensitive detection of DNA synthesis in vivo. Proc Natl Acad Sci U S A. 105 (7), 2415-2420 (2008).
- Hill, J. H., Chen, Z., Xu, H. Selective propagation of functional mitochondrial DNA during oogenesis restricts the transmission of a deleterious mitochondrial variant. Nat Genet. 46 (4), 389-392 (2014).
- Lentz, S. I., et al. Mitochondrial DNA (mtDNA) Biogenesis: Visualization and Duel Incorporation of BrdU and EdU Into Newly Synthesized mtDNA In Vitro. Journal of Histochemistry & Cytochemistry. 58 (2), 207-218 (2010).
- Tornoe, C. W., Christensen, C., Meldal, M. Peptidotriazoles on solid phase: [1,2,3]-triazoles by regiospecific copper(i)-catalyzed 1,3-dipolar cycloadditions of terminal alkynes to azides. J Org Chem. 67 (9), 3057-3064 (2002).
- Horwitz, H. B., Holt, C. E. Specific inhibition by ethidium bromide of mitochondrial DNA synthesis in physarum polycephalum. J. Cell Biol. 49, 546-553 (1971).
- Huberman, J. A. New views of the biochemistry of eucaryotic DNA replication revealed by aphidicolin, an unusual inhibitor of DNA polymerase alpha. Cell. 23 (3), 647-648 (1981).
- McClung, C., Hirsh, J. Stereotypic behavioral responses to free-base cocaine and the development of behavioral sensitization in Drosophila. Curr Biol. 8 (2), 109-112 (1998).
- Moore, M. S., et al. Ethanol intoxication in Drosophila: Genetic and pharmacological evidence for regulation by the cAMP signaling pathway. Cell. 93 (6), 997-1007 (1998).
- Dzitoyeva, S., Dimitrijevic, N., Manev, H. Gamma-aminobutyric acid B receptor 1 mediates behavior-impairing actions of alcohol in Drosophila: adult RNA interference and pharmacological evidence. Proc Natl Acad Sci U S A. 100 (9), 5485-5490 (2003).
- Koch, E. A., Spitzer, R. H. Multiple effects of colchicine on oogenesis in Drosophila: induced sterility and switch of potential oocyte to nurse-cell developmental pathway. Cell Tissue Res. 228 (1), 21-32 (1983).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。