
Method Article
点击化学可视化植物木动力学: 双标记是幸福!
摘要
极乐, 一个研究木动力学的双重标记协议被开发了。使用合成单体记者和 SPAAC 和 CuAAC bioorthogonal 单击反应的顺序组合, 此方法为深入分析生物木中的底的因素铺平了道路.
摘要
木质素是地球上最普遍的生物大分子之一, 也是纤维素生物质的主要成分。这种酚醛聚合物在高等植物的生长和生活中起着重要的结构和保护作用。虽然调节木过程的复杂机制在体内强烈影响许多植物衍生产品的工业稳定, 但科学界仍有很长的路要走去破译它们。在一个简单的三步工作流程中, 本文所提出的双标记协议使 bioimaging 对植物组织的活跃 lignifying 区进行了研究。第一步包括两个独立的化学记者的新陈代谢的合并, 两个本地 monolignols 的代理人, 产生木质素的 H 和 G 单位。在加入生长的木质素聚合物之后, 每位记者通过 bioorthogonal SPAAC/CuAAC 点击反应的顺序组合, 然后用自己的荧光探针进行特殊标记。结合木质素的自发荧光, 这一方法导致三色的植物细胞壁内木质素定位图的生成, 共焦荧光显微镜和提供精确的空间信息的存在或缺乏活动木机械的规模, 植物组织, 细胞和不同的细胞壁层。
引言
在过去的二十年里, 《化学记者战略》已经成为一个强有力的两步法来研究非基因编码生物分子的动力学和功能的方法。1,2,3在该策略中, 生物的一个合成模拟器 (即化学报告器) 首先由生物体进行代谢, 然后是一个化学探针 (e. g, 荧光荧光)共焦显微成像) 是通过 bioorthogonal 点击化学与合并后的记者进行共价键连接的。探针必须快速地和具体地反应对被介绍的化学制品的修改, 而对存在于活系统中的任何生物分子是惰性的。在许多方面, 这种方法克服了常见的 bioconjugation 技术的局限性, 通过使用高度特定的点击化学结扎从而提供了机会跟踪代谢物或大分子以前无法进入在生活系统中4,5,6。
尽管这种强大的方法在细菌和动物细胞中的快速增长的普及, 报告描述它在植物生物学中的使用是惊人的很少和远在7,8,9,10, 11,12。我们特别感兴趣的是在植物中应用这一策略来研究木质素的形成, 这是地球上最普遍的生物大分子之一, 也是纤维素生物质的主要成分。13,14木质素是一种在高等植物的生长和生活中起着重要的结构和保护作用的酚醛聚合物。
它一般由三 4-hydroxyphenylpropanoid 基: H (p羟基苯基), G (guaiacyl) 和 S (syringyl) 单位分别从三 "monolignols" (p-coumaryl, 松柏和 sinapyl 醇) 获得通过细胞胞浆中的丙烷通路合成 (图 1)。在被出口到细胞壁之后, monolignols 被氧化到自由基由过氧化物或漆之后他们接受纯化学的根本耦合反应对聚合对木质素聚合物, 过程被称为木。15,16虽然木强烈影响许多植物衍生产品的工业稳定, 但科学界仍有很长的路要去破译木的复杂机制。

图 1: 植物细胞中的木过程.Monolignols 是 biosynthesized 苯丙氨酸在胞。在被出口到细胞壁之后, monolignols 被氧化到自由基由过氧化物或漆之后他们接受纯化学的根本耦合反应对聚合对木质素聚合物, 过程被称为木。请单击此处查看此图的较大版本.
{kind=link}
虽然报告关于使用 bioorthogonal 反应为糖分析是许多,2,3,17他们的应用例子到其他类型的生物分子是较少。bioorthogonal 化学用于木质素 bioimaging 的用途最近才被 Tobimatsu et al.所开创8在拟南芥中, 提供有关将松柏醇替代物纳入木质素聚合物的信息, 其中形成 G 单位、8、9 , 从而展示了概念的证明化学报告策略在这方面是适用的。使用 CuAAC 也说明了使用不同的松柏醇衍生物的几个月后, 布考斯基et al。9但是, 木质素也包含了 H 和 S 单元, 对木过程的更深理解需要更多地了解如何将所有的 monolignols 纳入聚合物中, 以及哪些因素可以控制其组成。这一领域的新进展目前依赖于开发有效的方法, 在生活系统中同时跟踪多个化学记者。尽管一些关于糖的文章在近几年打下了基础18,19,20,21,22, 双标记方法仍然是一个主要的挑战bioorthogonal 化学如果一个可重现的单标记点击协议很难开发, 那么两个独立的化学记者之间的两个相互兼容的 bioorthogonal 反应的双重标记方法更是难上加难。这方面的几个例子, 率先采用了应变促进叠氮化-炔烃加 (SPAAC) 和烯烃-嗪逆电子要求翅果-桤木 (DA存货) 的反应, 研究糖在动物细胞。然而, 我们认为, 由于木质素的结构特征 (由富电子取代的桂型单体组成, 可与电子差反应, 所以在这个应用中, 可能无法保证 DAinv 反应的 bioorthogonality。烯烃, 如嗪探头用于 DAinv 反应), 这可能产生非特定的标签。此外, DA "库存" 反应需要综合难以访问的化学处理, 以及笨重和亲油性, 从而提高了化学品的注册、运输和/或本地化的可能性.记者在体内可能会受到影响。当我们认为, 后者方面是特别相关的, 在点击化学方法的情况下研究木, 我们选择了一个不同的方向, 并制定了一个 Bioorthogonal 结扎成像顺序策略 (极乐) 使用结合应变促进叠氮化-炔烃加 (SPAAC) 和铜催化叠氮化-炔烃加 (CuAAC)在体内.23
这两个反应确实是两个主要的 bioorthogonal 点击反应, 已被用于迄今, 特别是在最近发表的木质素成像的几个例子。8,9我们的双标记策略允许在一个单体的报告器和一个终端炔烃上使用一个叠氮化基团, 这两个化学处理是 i) 不对生物相关结构和 ii) 非常小的大小 (图 2).因此, 这些合成的修改对研究中的生物的物理化学性质的影响降到最低, 从而减少了自然和天然单体基材在运输方面可能存在的差异,代谢率在新陈代谢的并网阶段。虽然 SPAAC 和 CuAAC 的结合似乎非常直观, 乍一看, 它是我们的知识, 只有第二个例子的双重标记使用此策略和第一次应用的结构以外的糖。12,23

图 2: 极乐双标记策略.化学记者 hAZ和 g monolignols 是带有本地 h 和 g 的标记类似物.它们首先被引入外源喂养 (步骤 1) 的细胞壁的生长木质素聚合物中。Cyclooctyne-和叠氮化-功能化荧光探针, 然后依次结扎到合并的记者通过 bioorthogonal 点击化学: SPAAC 反应 (步骤 2) 是高度具体的 HAZ单位, 并在后面的 CuAAC 反应 (步骤 3), 它是特定于 G 的单元 (步骤 3), 从而允许两个记者的特定本地化独立于同一示例中.请单击此处查看此图的较大版本.
{kind=link}
我们首先设计并验证了叠氮化标签的单体记者HAZ ( p-coumaryl 醇的代用物和木质素 H 单元的前驱体), 然后制定了极乐双标记策略, 它与以前报告的炔烃标记的G与、9 (松柏醇的代用物和木质素 G 单位的前驱体)。在这个可复制的协议开发了和测试在胡麻, 一个经济上重要植物种类, 双重新陈代谢的并网 HAZ和G在木质中首先达到在顺序 SPAAC/CuAAC 之前标签.在这里, 标记的HAZ单元是第一次明确标记通过SPAAC 结扎的 cyclooctyne 功能化的荧光, 其次是 CuAAC 介导结扎第二个荧光探针在标记G单位。本方法用于研究植物细胞壁内木过程的动态变化, 并可应用于体内, 用于茎横断面、活茎及不同植物种类的幼苗。
研究方案
注: 液体和固体½ MS 介质必须事先准备好, 如补充表 1中所述。
1. 植物培养
-
2月老亚麻植物的栽培
- 播种亚麻 (亚麻亚麻L.) 在塑料花盆中使用盆栽堆肥的种子。
- 在生长室生长亚麻在22° c 以光周期 16 h/8 h 天或夜。
- 1月后向植物提供垂直支撑, 直到2月大。
-
2周老亚麻幼苗的培养
- 将12亚麻籽裹在一块乳酪布上, 用橡皮筋固定, 制成一捆。
注: 在层流烟气罩下进行无菌条件下的下一步步骤。 - 将种子捆放在以前的蒸压250毫升玻璃瓶中。
- 在瓶子里加70毫升的70% 乙醇, 轻轻搅拌1分钟。
- 小心地把乙醇迁, 同时把包留在瓶子里。
- 加入100毫升2% 次氯酸钠, 轻轻搅拌10分钟。
- 用迁去除次氯酸钠溶液, 同时把包留在瓶子里。
- 重复步骤 1.2. 5-1. 2.6。
- 加入100毫升无菌超纯水, 搅拌1分钟。
- 重复步骤 1.2.8 3 次, 在增加冲洗时间每漂洗 (5, 10 和 15 min)。
- 温热固体½ MS 培养基, 将其倒入无菌的植物培养盒中, 高度为2厘米, 让它冷却直至凝固。
- 用无菌钳将每种种子巧妙地放在固体培养基上。
- 关闭无菌箱。
- 将盒子转移到22° c 的生长室, 光周期为 16 h/8 小时/夜, 并种植亚麻幼苗2周。
- 将12亚麻籽裹在一块乳酪布上, 用橡皮筋固定, 制成一捆。
2. 化学记者的新陈代谢的合并
注: 三实验模型如下: i) 代谢标记的茎横断面, ii) 全茎和 iii) 植物幼苗。每个协议是一个生物复制和数量可以适应所需的生物复制的数量。单体解决方案是根据在实验前表 2中所述的库存解决方案编写的。库存解决方案可以在-20 ° c 的几个月内存储。HAZ: g 和或H: g比例可进行调整以适合所有定制的实验设计。
- 化学报告员加入2月老亚麻茎横断面
- 准备一个300µL 的化学记者解决方案, 包含10µM 的G 与和10µM 的HAZ 在液体无菌½ MS 培养基中。
- 涡HAZ/G 和解决方案, 并将其传输到一个具有微的48井板的井中。
- 准备一个300µL 负控制解决方案, 包含10µM 的G和10µM 的H在液体无菌½ MS 培养基。
- 涡流H/G解决方案, 并将其传输到同一48井板的第二个井。
- 用新的剃刀刀片将一个2月大的亚麻植株的茎秆切成10厘米的土壤。
- 使用刀片 (约150-250 µm 厚) 精心准备50徒手横断面的茎秆。
- 立即将横断面放置在½ MS 培养基中的玻璃杯中。
- 目视检查并选择完整的整个剖面, 并在填充了HAZ/G 与解决方案的井中随机分布10个剖面。
- 对填充了H/G解决方案 (作为负控制) 的井重复步骤2.1.8。
- 重复步骤2.1.8 为第三井填充300µL 的½ MS 培养基 (作为背景控制, 以调整荧光基线在随后的数据处理的共聚焦显微镜图像)。
- 在20° c 为 20 h 的生长腔中, 在连续光中孵育板。
- 用微去除每个井中的单体溶液。
- 添加500µL ½ MS 介质到每一个井, 轻轻搅拌10分钟, 并删除这个冲洗解决方案与微。重复4次, 立即进行极乐标记 (3 节)。
- 化学记者加入2月老亚麻全茎
- 在液体无菌½ MS 培养基中, 准备一个5毫升的化学记者解决方案, 其中包含10µM 的G 和10µM 的H 。
- 涡HAZ/G 和解决方案, 并将其传输到20厘米高的玻璃试管中, 并带有吸管。
- 在液体无菌½ MS 培养基中, 制备一个5毫升负控制溶液, 包含10µM 的G和10µM 的H 。
- 涡流H/G解决方案, 并将其传输到20厘米高的玻璃试管中。
- 准备第三个装有5毫升½ MS 介质的玻璃试管 (作为背景控制, 在随后的共焦显微镜图像数据处理过程中调整荧光基线)。
- 用新的剃刀刀片将一个2月大的亚麻植株的茎秆切成10厘米的土壤。丢弃根系统和下部的茎, 并继续到下一步与植物茎上部。
- 将整个切茎的底座浸入包含HAZ/G 对中解决方案的管道中。将茎在其孵化液中立即放在根部系统除去后, 以防止其干燥。
注: 如果该方法适用于较年轻的/较早的植物和/或其他物种, 单体溶液的体积必须根据植物的大小/年龄和茎的直径来调整。 - 将阀杆固定在垂直支承上, 使其保持平直。
- 密封玻璃管与膜, 以避免蒸发的解决方案。
- 对带有H/G解决方案的负控制示例重复步骤 2.2. 6-2. 2.9。
- 对含有½ MS 介质的荧光背景控制样品, 重复步骤 2.2. 6-2. 2.9。
- 用氖光在连续光中孵育每茎20小时。
- 在 20 h 新陈代谢的并网以后, 去除在hAZ/G 和解决方案中孵育的词干, 并用½ MS 介质冲洗。
- 使用刀片切割和丢弃的底部1厘米的茎, 然后准备一个1厘米高的缸从基地的剩余茎。
- 仔细准备20手绘横向横截面 (约150-250 µm 厚) 从这个气缸, 并把它们立即在一个充满½ MS 介质的手表玻璃。
- 把500µL 的½ MS 培养基放在一个48井板的井中。
- 目视检查和选择完整的整个横断面, 并随机分布其中的 10, 在充满了½ MS 解决方案。
- 重复步骤 2.2. 13-2. 2.17, 用于与H/G解决方案一起孵化的负控制阀杆。
- 重复步骤 2.2. 13-2. 2.17 的荧光背景控制杆孵育与½ MS 培养基。
- 小心地在每个井中删除解决方案与微。
- 添加500µL ½ MS 介质到每一个井, 轻轻搅拌10分钟, 并删除这个冲洗解决方案与微。重复4次, 立即进行极乐标记 (3 节)。
- 化学记者成立2周老亚麻幼苗
注: 以下步骤均在无菌条件下进行。- 在液体无菌½ MS 培养基中, 准备一个2毫升的化学记者解决方案, 其中包含10µM 的G 和10µM 的H 。
- 涡HAZ/G 和解决方案, 并将其传输到20厘米高的玻璃试管中, 并带有吸管。
- 在液体无菌½ MS 培养基中, 制备一个2毫升负控制溶液, 包含10µM 的G和10µM 的H 。
- 涡流H/G解决方案, 并将其传输到20厘米高的玻璃试管中。
- 准备第三个20厘米高的玻璃试管, 内含2毫升的½ MS 培养基 (作为背景控制, 在随后的共焦显微镜图像数据处理过程中调整荧光基线)。
- 用长的镊子 (1.2 节) 巧妙地从固体½ MS 培养基中去除2周老的亚麻幼苗。
- 将幼苗轻轻地转移到包含HAZ/G 和解决方案的管道中, 确保其根浸入解决方案中。
- 用塑料瓶盖将玻璃管关上以避免蒸发。
- 重复步骤 2.3. 6-2. 3.8 为负控制管与 H/G 溶液。
- 对含有½ MS 介质的荧光背景控制管重复步骤 2.3. 6-2. 3.8。
- 在20° c 的生长腔中, 在连续光照下孵育每个幼苗20小时。
- 巧妙地去除在HAZ/G 和解决方案中孵育的幼苗, 并用½ MS 介质将其冲洗干净。
- 微妙地准备20写意横断面 (近似150-250 µm 厚) 的下胚轴。
- 把500µL 的½ MS 培养基放在一个48井板的井中。
- 目视检查和选择完整的整个横断面, 并随机分布其中的 10, 在充满了½ MS 解决方案。
- 重复步骤 2.3. 12-2. 3.15 为负控制苗与H/G解决方案一起孵化。
- 重复步骤 2.3. 12-2. 3.15 的荧光背景控制幼苗与½ MS 培养基。
- 小心地在每个井中删除解决方案与微。
- 添加500µL ½ MS 介质到每一个井, 轻轻搅拌10分钟, 并删除这个冲洗解决方案与微。重复4次, 立即进行极乐标记 (3 节)。
3. SPAAC 和 CuAAC 对植物横断面样品的双重荧光标记
注: 极乐标记协议是相同的所有3实验模型 (第2节)。
-
应变促进叠氮化-炔烃加 (SPAAC) 标记
- 在液体无菌½ MS 培养基中制备1毫升的 SPAAC 溶液, 其中含有5µM 的 DBCO PEG4-5/6-6g 110。漩涡的解决方案, 并保持在黑暗中。
- 在新陈代谢的并网协议的最后的洗涤的步以后 [2.1.13, 2.2.21 或2.3.19 根据被选择的实验设计], 去除½ MS 媒介并且增加300µL SPAAC 解答每井与微。
- 通过覆盖铝箔或将它放在一个盒子里, 来屏蔽48井板的光。
- 在室温下, 在黑暗中轻轻搅动1小时的机械振动筛板。
-
铜催化叠氮化-炔烃加 (CuAAC) 标记
- 洗净每井4次的步骤 2.1.13, 同时保持在黑暗中的板块。
- 准备1毫升的 CuAAC 溶液, 其中含有2.5 毫米的抗坏血酸钠, 0.5 毫米丘索4和5µM 5-TAMRA-PEG3-叠氮化物在液体无菌½ MS 培养基中。漩涡的解决方案, 并保持在黑暗中。
注意: 此 CuAAC 解决方案必须在使用前进行新的准备, 并且不能在4° c 的几天内存储。 - 每井取出½ MS 介质, 并使用微直接加300µL 的 CuAAC 溶液。
- 在室温下, 在黑暗中轻轻搅动1小时的机械振动筛板。
- 使用微删除 CuAAC 解决方案。
- 使用微, 添加500µL ½ MS, 在黑暗中搅拌10分钟, 然后删除这个洗涤液。重复两次。
- 添加500µL 7:3 甲醇/水溶液, 在黑暗中搅拌1小时, 然后删除解决方案
注意: 甲醇通过皮肤接触或吸入而中毒, 被指定为人类致癌物。它的使用需要化学通风罩和手套。 - 添加500µL ½ MS, 在黑暗中搅拌10分钟, 然后删除解决方案。重复4次。
4. 在显微镜幻灯片上安装样品
- 将5滴安装介质放在玻璃显微镜幻灯片上。
- 小心存放在幻灯片上的每个HAZ/G和标记的跨节。
- 将指甲油涂抹在幻灯片的两面。
- 用片盖住滑梯。小心避免气泡。
- 使用纸巾去除多余的安装介质。
- 在所有4边上用指甲油密封样品。让指甲油在室温下干燥 (约30分钟)。
- 对负控制H/G跨节重复步骤 4.1-4.6。
- 重复步骤 4.1-4.6 的荧光背景控制横截面。
- 将幻灯片存放在4° c 的黑暗中, 直到在共聚焦显微镜下观察。
注意: 准备好的幻灯片可以存储几个星期到几个月, 这取决于准备的质量。如果一个新的观察是在储藏后, 确保样品没有干涸。
5. 用荧光共焦显微镜进行图像采集
- 按照制造商的指示, 按正确的顺序打开共焦显微镜系统的所有必要单元。
- 选择理想的目标, 最大数值孔径和适应激发和发射波长 (例如, obj 60x, 1.2。自发荧光通道: λex 405 毫微米, λem 450 nm/25;SPAAC HAZ 通道: λex 488 nm、λem 525 nm/25;CuAAC G 与通道: λex 561 nm、λem 595 nm/25;顺序模式)。
- 选择在软件中生成图像所需的参数 (需要12或16位, 像素大小适应奈奎斯特标准)。
- 在显微镜台上放置HAZ/G 幻灯片, 并将样品置于物镜下。
- 观察并找到所需的植物组织区域。
-
获取高质量的双标记 HAZ/G 与示例的高品质共焦图像.
- 选择 Z 轴中最荧光的平面。
- 调整增益, 偏移和激光功率, 以达到最佳的信号分布沿整个灰色水平范围为每个通道 (自发荧光, HAZ 和G和)。同样的参数必须适用于所有后续的收购。
- 启动所选映像的获取并保存文件。对样品的每个相关区域重复。
注: 如果需要, 还可以获取 3D Z 堆栈重建。
- 使用相同的设置, 获取标签样本的图像只孵化在½ MS, 以确定荧光水平 (如2节所述的萤光背景控制)。
- 使用相同的设置, 获取与本机 monolignols H/G一起孵化的负控制样品的图像, 以确定样品的非特定荧光粘附。
- 将数据处理应用于所获得的图像, 在减负控制图像和 hAZ/g 与图像中的背景自发荧光信号。
结果
通过使用提出的极乐协议涉及合成单体类似物和 bioorthogonal 点击化学, 它是可能的可视化动力学的木过程在活植物。与木质素可视化技术 (如组织化学染色, immunolocalization 或自发荧光) 不同, 这种 "双击" 协议专门针对在代谢合并过程中产生的木质素的从头步骤和区分它从先前存在的木质素样品与三重通道细胞成像的共焦荧光显微镜 (图 3)。H AZ-使用 DBCO-PEG4-5/6-6g 110 的激发和发射波长来检测单元, 在 SPAAC 步骤 (λex501 nm/λem 526 nm, 绿色通道), 而g在 azidefluor 545 探测器的特征波长上同样检测到 g 的合并终端炔烃标记在 CuAAC 步骤中进行λex 546 nm/λ em 565 nm, 红色通道);第三个通道对应于预先存在的木质素, 这是检测使用其固有的自发荧光在 405 nm (蓝色通道)。这种方法导致在植物细胞壁内生成三色的木质素定位图, 提供精确的空间信息, 说明器官不同组织间存在或缺乏活性木机械 (图3A), 在同一组织中的不同单元格类型 (图 3B-c) 和同一单元格的不同壁层内 (图 3D) 之间。换言之, 此方法允许我们通过将 "活动" 木站点 (HAZ和G与通道) 区分开来, 并将它们与早期形成木质素的区域进行区别.植物发育阶段 (自发荧光通道)。此外, 与自发荧光相比, 该技术的灵敏度大大提高, 而且可以检测到少量的新鲜合成木质素 (图 3B)。

图 3: 在亚麻茎中加入单体化学记者的成像.(a) 一半的手工制作的横断面的亚麻茎, 重建的图片。(B) 封闭的第一层区分木质部。左面板: 木质素自发荧光 (蓝色, 405 nm),中间面板: 合并木质素自发荧光 (蓝色) 和 HAZ荧光 (绿色, 526 nm) 通道,右面板: 合并木质素自发荧光 (蓝色) 和 G荧光 (红色, 565 nm) 通道。箭头表示第一个标记的细胞壁从形成层说明增加的灵敏度的极乐与荧光。(C) 在不同的次生木质部和 (D) 的细胞类型上贴上标签, 封闭在次生木质部细胞上, 说明同一细胞壁不同层的标记变化。左面板: 合并木质素自发荧光 (蓝色) 和 HAZ萤光 (绿色, 526 nm) 通道, 中间面板: 合并木质素自发荧光 (蓝色) 和 G 与 (红色, 565 nm) 通道,右面板: 合并木质素自发荧光 (蓝色), HAZ (绿色) 和 G 与 (红色) 荧光通道.HAZ和 G 和 co 本地化以黄色描述.请单击此处查看此图的较大版本.
{kind=link}
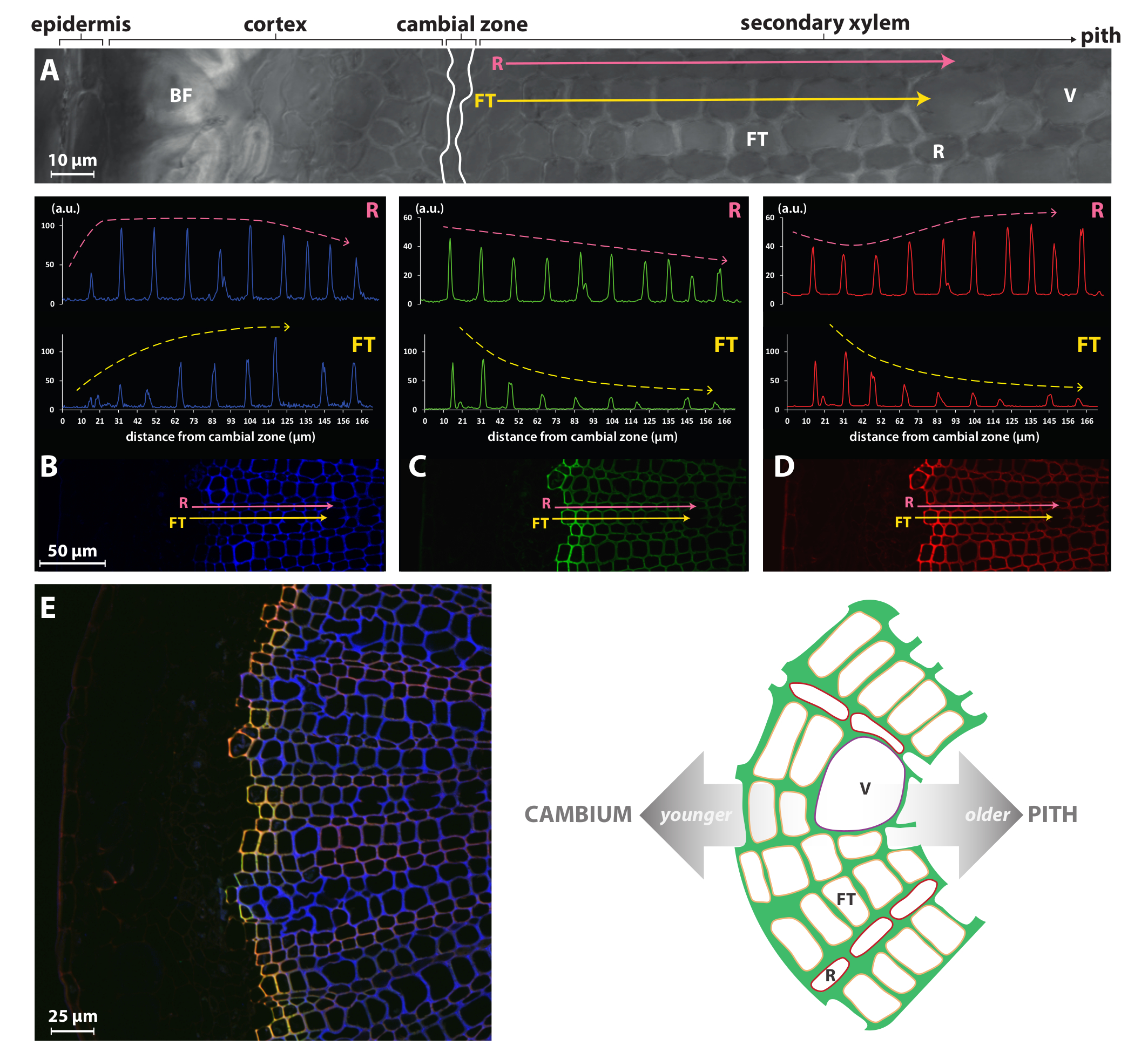
化学报告策略, 如此处介绍的极乐方法或以前由 Tobimatsu et al.发布的单标记程序因此, 8可以比以前可访问的方法 (如免疫或组织化学染色) 进行更细致的研究, 同时实现非常简单。例如,图 4说明了在亚麻次生木质部的纤维胞/容器 (FT) 中, HAZ/G的信号强度在最初的几个细胞壁中是非常高的, 但在较旧的细胞中逐渐减少。这个剖面与自体荧光的相对, 并表明最大木是非常迅速达到在前两到三英尺的细胞层从形成。相比之下, 射线 (R) 细胞似乎继续木到其发展的后期阶段, 因为HAZ/G与在所有细胞的壁上都更加恒定, 从形成层到髓.这并不奇怪, FT 和 R 细胞类型显示对比木动力学, 因为这些细胞类型有不同的生物学角色与相关的差异, 他们的发展计划。罚球确实是细长的成熟的管, 并迅速死亡, 留下死的空细胞与厚的化墙, 发挥关键作用, 在机械支持和垂直运输的水和矿物质, 而窄行的 R 细胞组成的木质部光线在其成熟和功能状态下是活的, 没有厚的化壁。

图 4: 特定于单元格的单体报告员在亚麻木质部中成立.亮场共焦显微镜观察 (a) 部分的手绘横断面从亚麻茎。粉红色 (射线薄壁细胞, R) 和黄色 (纤维胞细胞, FT) 箭头指示向量跨越从层区向髓和扫描木质素自发荧光在 405 nm (B), HAZ荧光在 526 nm (C) 和 G荧光在 565 nm (D)。(E) 次生木质部的视图。左面板: 合并木质素自发荧光 (蓝色)、HAZ (绿色) 和 G 与 (红色) 荧光通道.HAZ和 G 和 co 本地化以黄色描述.右面板: 亚麻茎部次生木质部结构示意图。V、船只;FT, 纤维胞;R, 射线薄壁细胞。此图已从狮子et al.中修改23请单击此处查看此图的较大版本.
{kind=link}
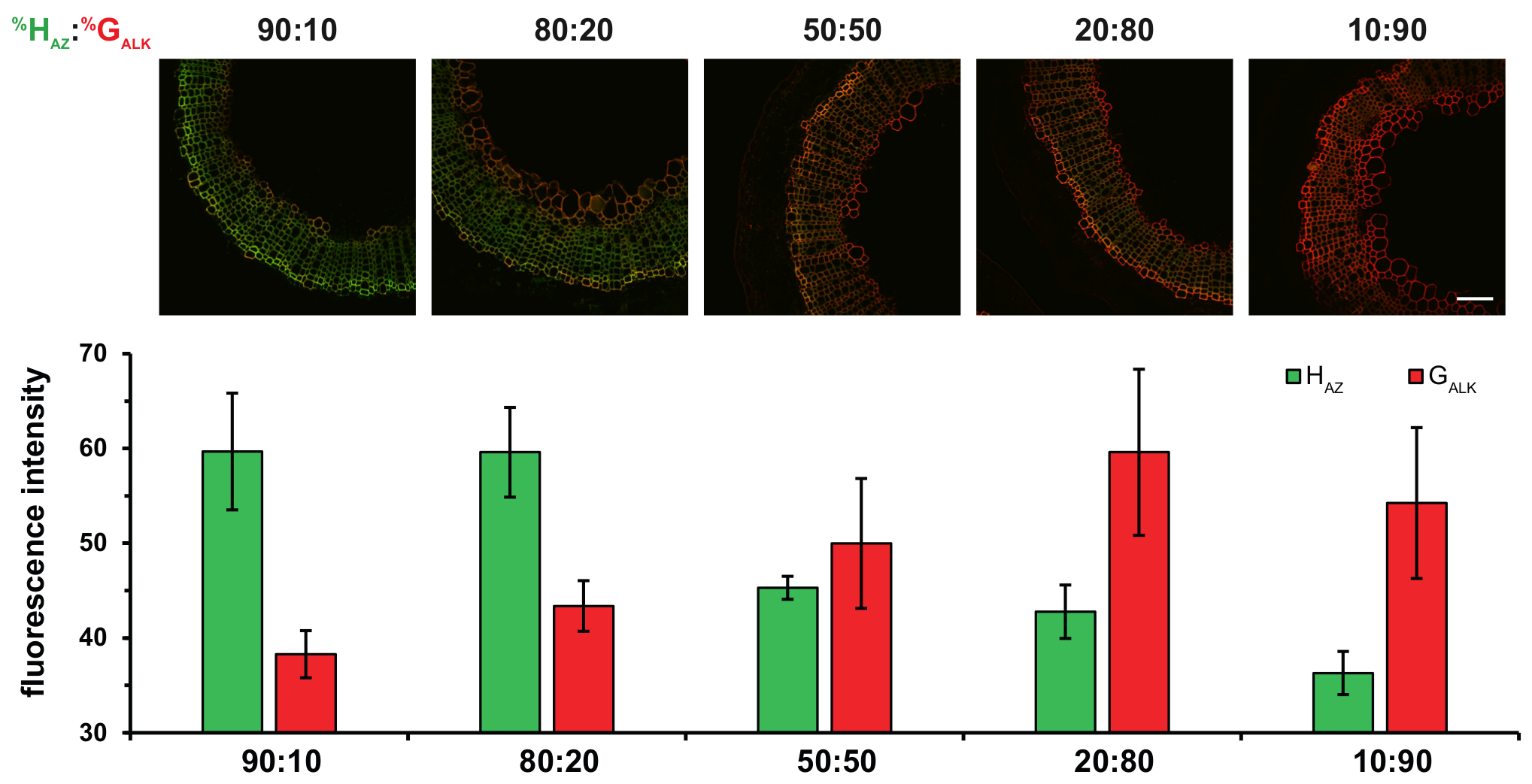
此外, 使用两个不同的化学记者对应的 H 和 G 单位的两个 bioorthogonal 反应在极乐协议允许更多的定量结果得到。多路复用标签策略, 如极乐, 可以提供更多的洞察力, 控制单体的并网比率在不同的条件, 并有助于破译难以捉摸的木过程。如果木中 H、G 和 S monolignols 的相对百分比确实是根据植物的种类、组织、年龄或环境条件而变化的, 那么调节这一成分的错综复杂的机制还没有完全被理解。双贴标技术是研究调节木质素成分的一些参数的有力途径。例如, 更改HAZ: G与极乐的比值使我们能够证明次生木质部组织中的木质素成分直接依赖于细胞壁内的单体可用性, 而不是过氧化物酶/漆酶特异性 (图 5)。

图 5: 用不同比例的 HAZ和 G 对中来孵化亚麻茎段的效果.HAZ: 在每个数字列的上方都给出了 G 与% 的比率.顶部: 合并的 HAZ和 G 和通道.底部: 直方图表示 HAZ的平均荧光强度和 different% 比率的 G 对. 数值以灰度级平均荧光强度的平均值表示. 缩放栏 = 100 µm. 此图已从 狮子 et al.中修改23请单击此处查看此图的较大版本.
{kind=link}
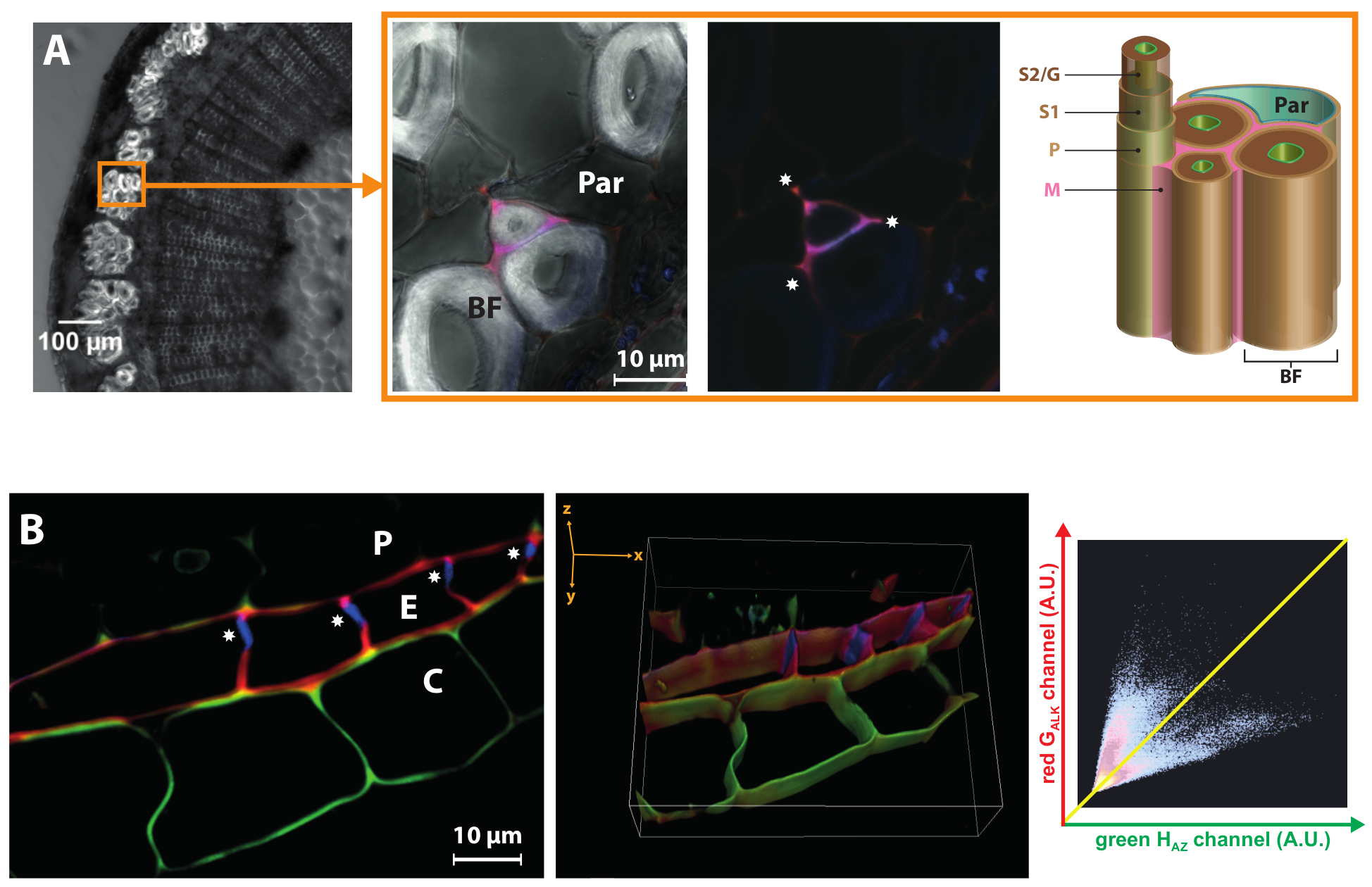
亚麻是为它的麻纤维 (BF) 被用于制造纺织品, 豪华纸或环境友好的复合材料。他们的工业稳定的一个重要方面是, 它们在其细胞壁中含有非常低的木质素, 其特征是极厚的 S2/G 次层19,24。极乐可以突出木在同一细胞壁的不同层之间的动态差异。图 6A显示HAZ和G通讯报告器合并仅限于某些但并非所有麻纤维的单元格角和中鳞片/原细胞壁.木在厚麻纤维二次细胞壁层中的总缺失甚至当单体化学记者外部提供时, 表明它们的 hypolignified 状态是由于缺乏适合酶介导的氧化和 monolignols 加入到一个不断增长的聚合物链, 而且它不只是由于转录调控的单体生物合成基因先前报告的25。这一观察也与亚麻过氧化物酶基因在具有化麻纤维的亚麻化学lbf1突变体的外茎组织中上调的事实相关.26。

图 6: 极乐突显细胞壁下部结构-或层特定的差异.(a) 左面板: 亚麻茎段的亮场图像。圆圈表示纤维束。右面板: 接近麻纤维。合并的 HAZ和 G 与木通道 (带有和不亮场) 显示, 该信道仅限于单元格角 () 和某些麻纤维的中间鳞片/原细胞壁.  BF, 麻纤维;Par, 薄壁细胞;M, 中间鳞片;P, 原细胞壁;S1, 二级细胞壁第一层;S2/G, 次生细胞壁的次生层/凝胶层。(B) 2D 切片和3D 重建亚麻根内皮区共焦 z 叠加变焦。凯氏条 () 只显示了自发荧光, 不包含 HAZ或 G "对".关联的 fluorogram 显示一个 G 与/HAZ反关联: 高绿色荧光与低红色信号 (皮层) 和反之亦然 (内皮) 相关。柱 (P), 内皮 (E), 皮质 (C)。请单击此处查看此图的较大版本.
BF, 麻纤维;Par, 薄壁细胞;M, 中间鳞片;P, 原细胞壁;S1, 二级细胞壁第一层;S2/G, 次生细胞壁的次生层/凝胶层。(B) 2D 切片和3D 重建亚麻根内皮区共焦 z 叠加变焦。凯氏条 () 只显示了自发荧光, 不包含 HAZ或 G "对".关联的 fluorogram 显示一个 G 与/HAZ反关联: 高绿色荧光与低红色信号 (皮层) 和反之亦然 (内皮) 相关。柱 (P), 内皮 (E), 皮质 (C)。请单击此处查看此图的较大版本.
{kind=link}
最后, 这两种颜色的方法也可以提供有价值的信息的 "木状态" 细胞壁子结构在不同发育阶段和植物器官以外的茎。例如, 亚麻根的内皮的特点是在发育的早期阶段, 其径向和横向细胞壁存在凯氏带。这一带疏水性生物, 由 suberin 和/或木质素, 防止水和溶质的根采取被动进入通过体, 迫使他们通过通过等离子膜通过彼此路线, 从而有助于植物根系的选择性吸收能力。当应用到亚麻根时, 我们的策略表明, HAZ和 G 胚被合并在凯氏带的切向壁中, 也包含在缺少 ( 图 6B) 的径向壁的部分。然而, 单体记者在凯氏带本身的完全缺席表明, 它是成熟的在这个发展阶段, 而其他墙壁仍然是进一步生物沉积的地点。z 叠重构的3D 视图清楚地带凯氏带到光。有趣的是, 毗邻内皮的一些皮质细胞的壁也被证明能够优先合并 HAZ (从而表明在亚麻根皮质中存在木质素/suberin), 而不是 G 和.联合定位分析显示了 HAZ和 G 对位之间的反相关关系, 这表明在这两种相邻的单元格类型中存在特定于细胞的壁结构/酶机制.
补充表 1: 固体½ MS 培养基的制备请单击此处下载此文件.
补充表 2: 准备化学报告员库存解决方案请单击此处下载此文件.
讨论
如前所述, 本文介绍的双标记极乐协议是 SPAAC/CuAAC 组合在体内1223的第一个示例之一。每一个步骤都经过了彻底的优化和验证, 这是非常重要的, 这两个点击化学标签反应顺序执行是尊重 (即, SPAAC 第一, 其次是 CuAAC)。所有交叉控件都显示, 在应用极乐协议时, 每个标记步骤都是特定的23 : 执行 SPAAC 步骤首先导致HAZ叠氮化函数的高度选择性标记。cyclooctyne-官能化荧光通过 [3 + 2] 加反应与快速动力学。一旦HAZ单元被标记, CuAAC 步骤就需要铜 (I) 催化激活G炔烃, 通过与叠氮化-氟545探头的反应生成三唑链.相反, 相反的顺序 (即, CuAAC 第一, 其次是 SPAAC) 不应使用, 因为它导致G与和 HAZ单位交叉耦合, 这与荧光结扎和诱发巨大的信号损失.此外, 还必须强调中间洗涤步骤的必要性, 以避免非特定的染色。
结果表明, 本方法可用于各种生物实验设计。《极乐标记协议》首次应用于亚麻茎秆 (大约150-250 µm 厚) 的徒手横断面上, 这是以前切割和孵化的 monolignols。虽然这种设计的优势是最大限度地减少所需数量的化学记者 (随着孵化量的减少) 和促进统计复制的生产, 它不是严格说来, 一个在体内系统, 并在一些的情况下, 可能不反映所有方面的真实时空木动力学。在第二个实验设计中, 我们因此改编了极乐协议的方法, 以前用于研究标记 monolignols 在松树和银杏27中的加入。在这种方法中, 植物的根和茎是物理分离的, 整个茎的基部在单体溶液中孕育, 这可能被称为 "花瓶" 的方法。在留下茎期望 (潜伏期) 时间之后, 横断面被切开和极乐协议执行。这使我们可以证明 (i), 经改良的 monolignols 是通过活干运输的, 并在细胞壁内生长的木质素聚合物中加入, (ii), 本地化模式与横截面的结构基本相同。方法.这类实验的优点是在实际的活体细胞中进行, 允许更长的实验和更深入的研究, 但需要更多的化学报告。最后, 极乐协议也用于亚麻植物幼苗, 代表一个真正的生活植物模型, 其中化学记者必须通过根部吸收, 然后才被输送到茎。虽然这种模式具有明显的优势, 在活植物中进行, 在实践中, 它是有限的幼苗和不真正适合调查木动力学的老植物的实际原因 (长潜伏期, 高化学记者数量)。然而, 这三实验设计是互补的, 都有他们的利弊有关的实际方面和生物学意义上的类型的生物问题要回答。
为了研究亚麻的木动力学, 我们的协议具有很强的适应性, 不仅在生物实验设计方面, 而且在其他植物种类和器官/组织的应用方面也是如此。例如, 极乐可以很容易地转移到拟南芥或杨树属, 更适合研究与敲出或击倒突变体的各种基因。原则上, 与我们的方法的双重标记研究也可以扩展到其他生物分子通过使用植物细胞壁聚合物的二个不同的被改进的前体-包括所有三主要 monolignols 或他们的新陈代谢的前体并且各种各样构成多糖基质的单糖。自成立以来, bioorthogonal 化学确实主要是通过代谢性寡糖工程 (教育部)4、5、17、28来研究糖/多糖但令人惊讶的是, 目前仅有很少的植物生物学应用,7,8,9,10,11.就反应的相容性而言, 木质素的研究确实是一个复杂的案例, 因为两个化学记者都被纳入同一网状聚合物中。 未标记的Haz-g与跨链路编队的可能性是由于3D 内的 g与和H单元的空间接近而克服的主要问题。木质素的结构23, 如果两个化学记者没有合并到同一类型的生物或任何给定单元的同一空间区域, 则可能不存在此限制。
在更广泛的范围内, 极乐方法可以应用于细菌或动物模型中的任何双色荧光成像研究, 分别使用两个具有叠氮化物和末端炔烃标记的化学记者。
披露声明
作者声明他们没有竞争的金融利益。
致谢
我们感谢研究联盟 FRABio 和 TisBio 成像平台 (Univ. 里尔, CNRS, FR 3688, FRABio, BiochimieStructurale et Fonctionnelle des 组合 Biomoléculaires), 为提供有利于实现这一目标的技术环境工作.
材料
| Name | Company | Catalog Number | Comments |
| (E)-4-(3-(2-(2-(2-azidoethoxy)ethoxy)ethoxy)prop-1-en-1-yl)phenol (HAZ) | Synthesized as in Lion et al. Cell Chem. Biol. 2017, 24, 3, 326-338 | ||
| (E)-4-hydroxy-3-propargyloxycinnamyl alcohol (GALK) | Synthesized as in Lion et al. Cell Chem. Biol. 2017, 24, 3, 326-338 | ||
| 2% sodium hypochlorite | |||
| 20 cm high glass tube | |||
| 250 mL Schott glass bottle | |||
| 48-well Plate | |||
| 5/6-TAMRA-PEG3-Azide | Jena Bioscience | CLK-AZ109-1 | |
| Aluminium foil | |||
| Cheese cloth | |||
| Compost containing clay | |||
| Coniferyl alcohol (G) | Sigma Aldrich | MFCD00002922 | |
| Copper (II) sulfate pentahydrate | |||
| DBCO-PEG4-5/6-Carboxyrhodamine 110 | Jena Bioscience | CLK-A127-1 | |
| Milli-Q Ultrapure water | |||
| Eppendorf 1,5 mL | |||
| EtOH | |||
| Flax seeds (L. usitatissimum L.) | |||
| Fluoromount-G™ Slide Mounting Medium | Electron Microscopy Sciences | 17984-25 | |
| Glass coverslip | |||
| Glass microscope slide | |||
| Growth chamber | CLF-Plant Climatics | For 2-week-old plants culture | |
| Growth chamber | Angelantoni Life Sciences | For 2-month-old plants culture | |
| Magenta plant culture box | For 2-week-old seedling culture | ||
| Methanol | Toxic (SGH02, SGH06, SGH08), work with gloves under a hood | ||
| Micropipette | |||
| Nail polish | |||
| Nikon A1R confocal microscope | Nikon | ||
| Orbital shaker | |||
| Parafilm | |||
| p-Coumaryl alcohol (H) | Carbosynth | FC145653 | |
| Plastic cap | |||
| Plastic pipette | |||
| Plastic pot | For 2-month-old plants culture | ||
| Razor blade | |||
| Rubber band | |||
| Sodium Ascorbate | |||
| Sterile clamp | |||
| Vertical support | |||
| Vortex | |||
| Reagents for liquid and solid ½ MS medium | |||
| KH2PO4 | |||
| KNO3 | |||
| NH4NO3 | |||
| MgSO4.7H2O | |||
| CaCl2.2H2O | |||
| MnSO4.H2O | |||
| ZnSO4.7H2O | |||
| H3BO3 | |||
| KI | |||
| Na2MoO4.2H2O | |||
| CuSO4.5H2O | |||
| CoCl2.6H2O | |||
| Na2EDTA.2H2O | |||
| FeSO4.7H2O | |||
| Thiamine.HCl | |||
| Pyridoxine.HCl | |||
| Glycine | |||
| Nicotinic acid | |||
| Myo-inositol | |||
| Saccharose | |||
| MES hydrate | |||
| Agar |
参考文献
- Grammel, M., Hang, H. C. Chemical reporters for biological discovery. Nat Chem Biol. 9 (8), 475-484 (2013).
- Prescher, J. A., Bertozzi, C. R. Chemistry in living systems. Nat Chem Biol. 1 (1), 13-21 (2005).
- Sletten, E. M., Bertozzi, C. R. Bioorthogonal chemistry: fishing for selectivity in a sea of functionality. Ange Chemie (Int Ed. Engl). 48 (38), 6974-6998 (2009).
- Chang, P. V., et al. Metabolic labeling of sialic acids in living animals with alkynyl sugars. Ange Chemie (Int Ed. Engl). 48 (22), 4030-4033 (2009).
- Gilormini, P. A., et al. A sequential bioorthogonal dual strategy: ManNAl and SiaNAl as distinct tools to unravel sialic acid metabolic pathways. Chem. Commun. 52 (11), 2318-2321 (2016).
- Mbua, N. E., et al. Abnormal accumulation and recycling of glycoproteins visualized in Niemann-Pick type C cells using the chemical reporter strategy. Proc Natl Acad Sci U S A. 110 (25), 10207-10212 (2013).
- Anderson, C. T., Wallace, I. S., Somerville, C. R. Metabolic click-labeling with a fucose analog reveals pectin delivery, architecture, and dynamics in Arabidopsis cell walls. Proc Natl Acad Sci U S A. 109 (4), 1329-1334 (2012).
- Tobimatsu, Y., et al. A click chemistry strategy for visualization of plant cell wall lignification. Chem. Commun. 50 (82), 12262-12265 (2014).
- Bukowski, N., et al. Development of a clickable designer monolignol for interrogation of lignification in plant cell walls. Bioconjugate Chem. 25 (12), 2189-2196 (2014).
- Pandey, J. L., et al. A versatile click-compatible monolignol probe to study lignin deposition in plant cell walls. PLOS ONE. 10 (4), e0121334 (2015).
- Pandey, J. L., et al. Investigating biochemical and developmental dependencies of lignification with a click-compatible monolignol analog in Arabidopsis thaliana stems. Front Plant Sci. 7, (2016).
- Zhu, Y., Wu, J., Chen, X. Metabolic labeling and imaging of N-linked glycans in Arabidopsis thaliana. Ange Chemie (Int Ed. Engl). 55 (32), 9301-9305 (2016).
- Boerjan, W., Ralph, J., Baucher, M. Lignin biosynthesis. Annu Rev Plant Biol. 54, 519-546 (2003).
- Weng, J. -. K., Chapple, C. The origin and evolution of lignin biosynthesis. New Phytol. 187 (2), 273-285 (2010).
- Mottiar, Y., Vanholme, R., Boerjan, W., Ralph, J., Mansfield, S. D. Designer lignins: harnessing the plasticity of lignification. Curr Opin Biotechnol. 37, 190-200 (2016).
- Rinaldi, R., et al. Paving the way for lignin valorisation: recent advances in bioengineering, biorefining and catalysis. Ange Chemie (Int Ed). 55 (29), 8164-8215 (2016).
- Wratil, P. R., Horstkorte, R., Reutter, W. Metabolic glycoengineering with N-acyl side chain modified mannosamines. Ange Chemie (Int Ed. Engl). 55 (33), 9482-9512 (2016).
- Feng, L., et al. Bifunctional unnatural sialic acids for dual metabolic labeling of cell-surface sialylated glycans. J Am Chem Soc. 135 (25), 9244-9247 (2013).
- Dumont, M., et al. Plant cell wall imaging by metabolic click-mediated labelling of rhamnogalacturonan II using azido 3-deoxy- d - manno -oct-2-ulosonic acid. Plant J. 85 (3), 437-447 (2016).
- Niederwieser, A., et al. Two-color glycan labeling of live cells by a combination of Diels-Alder and click chemistry. Ange Chemie Int Ed. 52 (15), 4265-4268 (2013).
- Späte, A. -. K., et al. Exploring the potential of norbornene-modified mannosamine derivatives for metabolic glycoengineering. Chem Bio Chem. 17 (14), 1374-1383 (2016).
- Späte, A. -. K., et al. Rapid Labeling of metabolically engineered cell-surface glycoconjugates with a carbamate-linked cyclopropene reporter. Bioconjugate Chem. 25 (1), 147-154 (2014).
- Lion, C., et al. BLISS: a bioorthogonal dual-labeling strategy to unravel lignification dynamics in plants. Cell Chem Biol. 24 (3), 326-338 (2017).
- del Río, J. C., et al. Structural characterization of guaiacyl-rich lignins in flax (Linum usitatissimum) fibers and shives. J Agric Food Chemist. 59 (20), 11088-11099 (2011).
- Huis, R., et al. Natural hypolignification is associated with extensive oligolignol accumulation in flax stems. Plant Physiol. 158 (4), 1893-1915 (2012).
- Chantreau, M., et al. Ectopic lignification in the flax lignified bast fiber1 mutant stem is associated with tissue-specific modifications in gene expression and cell wall composition. The Plant Cell. 26 (11), 4462-4482 (2014).
- Terashima, N., Ko, C., Matsushita, Y., Westermark, U. Monolignol glucosides as intermediate compounds in lignin biosynthesis. Revisiting the cell wall lignification and new 13C-tracer experiments with Ginkgo biloba and Magnolia liliiflora. Holzforschung. 70 (9), 801-810 (2016).
- Noel, M., et al. Probing the CMP-sialic acid donor specificity of two human β-d-galactoside sialyltransferases (ST3Gal I and ST6Gal I) selectively acting on O- and N-glycosylproteins. Chembiochem: Eur J Chem Biol. , (2017).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。