
Method Article
线虫生殖道内精子引导和运动的测定
摘要
精子必须成功地通过输卵管来受精卵母细胞。在这里, 我们描述了一种测量精子在雌雄同体子宫内的精子迁移的方法。该方法可以提供有关交配后精子在子宫内的分布的定量数据, 以及关于速度、定向速度和反转频率的定量数据。
摘要
成功的受精是有性生殖的基础, 但对引导精子进入女性生殖道内卵母细胞的机制了解甚少。虽然体外研究表明, 体内受精动物的精子可以对周围的各种线索做出反应, 但由于无法想象它们在女性生殖道内的行为, 这给理解精子的迁移带来了挑战和移动性。在这里, 我们描述了一种使用线虫的方法, 它克服了这一限制, 并利用了它们的透明表皮。被线粒体染料染色的线虫雄性与成年雌雄同体 (作为修饰的雌性) 交配, 并将荧光标记的精子沉积到雌雄同体子宫中。然后, 可以使用在活雌雄同体中使用 epi-fluorescence 显微镜直接跟踪标记精子的迁移和运动能力。在野生类型的动物中, 大约90% 的标记精子在子宫中爬行, 到达受精部位, 或精子。在交配后1小时内可以拍摄子宫图像, 以评估精子在子宫内的分布情况以及到达精子的精子百分比。或者, 可以在交配后立即拍摄延时图像, 以评估精子速度、定向速度和反转频率。这种方法可以与其他遗传和分子工具相结合, 可供虫识别新的遗传和分子机制, 这些机制对调节女性生殖道内的精子引导和活力很重要。
引言
精子 (称为精子) 通过女性生殖道向卵母细胞导航的分子机制尚不清楚, 但对有性生殖的基础却很重要。精子运动是高度动态的, 取决于强大的通信信号, 改变精子速度和方向运动 1,2,3,4,5,6,7.,8,9,10,11,12.线虫已成为研究精子体内运动的有力模型, 因为雌雄同体的透明表皮允许在单个细胞分辨率2,3下跟踪活精子, 8,10。本文的目的是为评估精子在雌雄同体子宫内的运动提供方法。
在精子和卵母细胞在外部环境 (即光细胞环境) 中相遇的动物物种中, 精子对卵母细胞分泌的趋化信号做出反应。这些信号引导精子运动的方向, 使它们更接近信号源4,6,11。然而, 对内部受精物种中精子运动的了解要少得多。一个主要的挑战是女性生殖道的结构, 在大多数物种中, 显微无法接触到女性生殖道。例如, 在人类、小鼠和猪身上进行的体外研究提供了证据, 证明精子的亚群可以对趋化因子、流体流动和热梯度1,5, 7,9做出反应。 12岁有了这些系统, 无法可视化和跟踪精子在体内的运动, 这对发现调节这些功能的关键机制的策略造成了严重的限制。
为了克服这些限制, 我们开发了一种方法, 利用线虫线虫直接显示精子在受精后, 测量个人精子在体内的迁移参数, 并测量精子数量的目标能力施肥地点。这些方法, 连同线虫分子和遗传工具集, 促进了化学信号分子和分子机制的发现, 调节精子的运动行为。例如, 基因屏幕可以在雌雄同体或雄性中进行, 以确定对体内有效精子运动至关重要的基因 13.分子可以注射到雌雄同体性腺, 以测试对精子活化、迁移速度和定向动力3的影响.此外, 所描述的方法可用于监测无赖精子迁移到异位体的位置, 并评估精子竞争10,14。
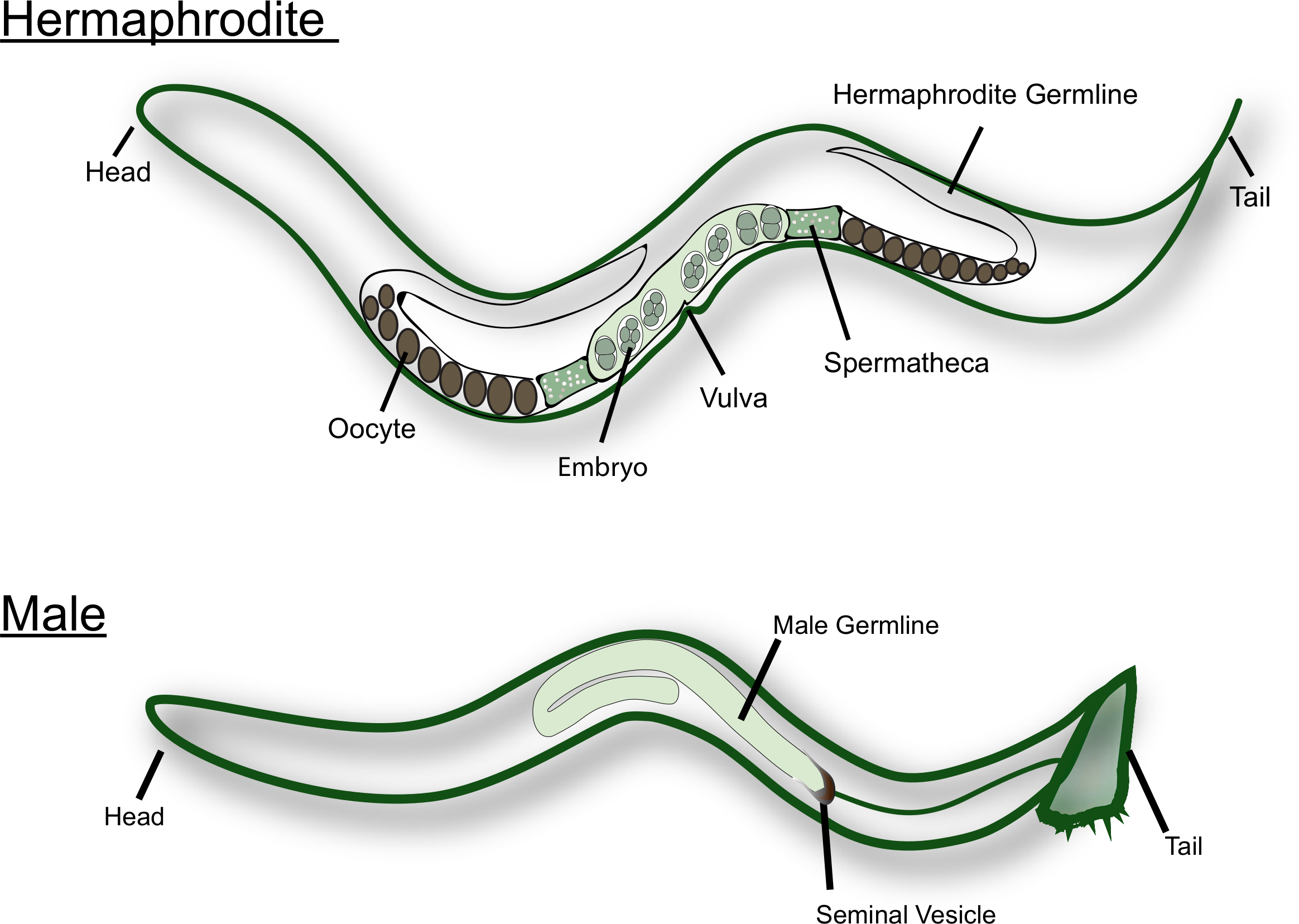
线虫在自然界中以雌雄同体和雄性的形式存在 (见图 1)。雌雄同体性腺有两个 u 形的手臂, 是彼此的镜像。在 L4 幼虫阶段, 最近端生殖细胞 (即精子附近的细胞) 发生精子发生。每个原代精母细胞进入减数分裂, 并产生四个单倍体精子细胞。这些精子细胞与第一个成熟的卵母细胞一起被推入精子细胞, 并经历精子发生 15.成虫从精子发生转变为生质。卵母细胞沿着性腺以装配线的方式成熟, 最成熟的卵母细胞位于性腺的近端, 旁边是精子。需要来自精子的 msp 信号来触发减数分裂的成熟和排卵16,17。男性c.线虫, 另一方面, 有一个 j 形的性腺, 只产生精子。精子细胞储存在精囊中。当与雌雄同体或雌配时 , 雄性会将尾巴附近的小刺插入外阴中。精子在射精过程中被激活 , 当它们接触18时。线虫的精子不会游泳, 因为它们没有鞭毛。相反, 它们在生殖道中爬行, 使用假足动物进行运动。众所周知, 男性精子, 这是较大的大小, 有一个竞争优势, 比雌雄同体精子14。
在这种方法中,雄性线虫作为精子捐献者, 并与成虫交配。成年男性被荧光线粒体染料染色, 以产生标记的精子。一旦通过雌雄同体沉积, 精子必须在子宫中的胚胎周围爬行, 走向精子或受精部位。线虫模型的透明表皮允许直接显示每个精子, 因为它通过女性生殖道导航。近年来, 我们的实验室已经成功地使用这种方法来证明一类 f 系列前列腺素在引导精子从外阴到精子1920 的重要性。控制其合成的分子机制仍在研究中。然而, 这种评估精子活力和迁移的方法极大地促进了控制体内受精动物精子和卵母细胞通信的关键参与者的识别。下面的协议描述了如何执行此检测。
研究方案
注: 本协议中的所有步骤均在室温 (~ 20-22°c) 或恒温孵化器中执行, 设定为16°c 或20°c。男性和雌雄同体线虫生长使用标准的培养条件和 na22 或 op50大肠杆菌作为食物来源21,22。在下面的过程中使用了野生 N2 雌雄同体和雾 2(q71)雄性。
1. 第1天: 选择 L4 级雌雄同体进行治疗
- 为了获得一致的结果, 所有雌雄同体都应作为积极繁殖的成虫进行同步。将20-30 级雌雄同体采摘到6厘米种子线虫生长培养基 (NGM) 板上。在20°c 下培养雌雄同体28-30小时。
请注意:只有12-15 个雌雄同体将用于交配。剩下的雌雄同体是多余的。
2. 第1天: 用荧光线粒体染料染色的男性
- 在未播种的 NGM 板的中心放置一个大肠杆菌 (食物点) 的点, 使男性染色板。要制作食物点, 请使用玻璃搅拌棒的末端, 将大肠杆菌从种子板的细菌草坪上刮掉, 并将其沉积在未播种的板上。圆点的直径应为 ~ 5-7 毫米。
- 在 DMSO 中混合 2Μl 1 Mm 线粒体染料 (见材料表) 和 10Μl mM 缓冲液 (3 克 kh 2 po4, 6g Na 2 hpo 4, 5 克氯化钠, 1 mM 1 mM 4, h2 o 至 1 l 后添加 mgso4 高压灭菌)。将所有的丝裂染料溶液输送到男性染色板上的食物点上。让盘子在黑暗中干燥 (~ 30分钟)。
请注意:三丝染料对光敏感。保护所有含有丝染的溶液、板材和蠕虫不受光线的影响。将 1 mM 库存存放在-20°c。 - 选择 ~ 100 1-3天的成年男性 23 到丝染染色食物点上的男性染色盘。用铝箔包裹板材, 在16°c 下孵育过夜。交配时, 每12-15 个雌雄同体使用 ~ 50-60 个雄性。如果需要超过100名男性, 制作更多的染色板, 以防止男性过度拥挤。
请注意:男性也可以通过在 M9 缓冲液中的10Μm 丝染溶液中孵育3小时的手表玻璃来染色。将蠕虫覆盖, 以防止蒸发和暴露。3小时后, 使用巴斯德移液将雄性转移到10厘米的种子 NGM 板上。转移尽可能少的丝裂染料溶液。将板材包裹在铝箔中, 在16°c 中孵育一夜。
3. 第2天: 约会
- 从第一天开始, 把被污染的雄性捡到一个新的, 种子 NGM 盘子上。把盘子放在黑暗中, 直到交配。这一步骤可确保去除男性周围多余的有丝分裂染料染色细菌。在交配板上携带过多的有丝分裂染料染色细菌可以染色雌雄同体组织。
- 通过将厚度的大肠杆菌混合物的 2Μl滴入未播种的 ngm 板, 制作配合板。让厚实的细菌干燥, 使交配点。雄性和雌雄同体将被转移到这个点交配。要制作厚实的大肠杆菌, 将隔夜大肠杆菌的3毫升旋转, 并在 mL 的1毫升中重新悬浮细菌颗粒。这种混合物可在4°C 下储存, 再利用长达6个月。
请注意:大肠杆菌溶液的厚度可以调整。如果溶液太薄, 雄性可能会从交配点上爬出来, 而不是聚集在上面交配。用大肠杆菌溶质制成的太厚的交配点可能会降低交配效率。 - 当步骤3.2 中的配合板干燥时, 将300Μl 的 1% (w/v) tricaine (tri)、300μl 的 0.1% (w/v) Tetrbisole (Tet) 和900μl M9 混合在一起。
请注意:将三卡因和 0.1% (w v) 作为引极剂储存在-20°c。避免反复冻融。 - 将 Tet/Tri 溶液的600μl 转移到手表玻璃上。
- 将第一天采摘的12-15 个雌雄同体转移到手表玻璃中的泰提-三溶液中。孵化 30分钟, 固定雌雄同体。保持手表玻璃覆盖, 以防止泰图三溶液蒸发。
请注意:重要的是, 在图像采集过程中, 两性的麻醉时间至少为 30分钟, 减少了产生运动蠕虫的时间, 这与成像有一定的作用。 - 雌雄同体在孵化时, 从步骤3.1 中选择50-60 个染色雄性, 进入交配点 (步骤 3.2)。将盘子存放在黑暗中, 直到第3.8 步。
- 在 Tet/tri 溶液中孵育30分钟后, 使用玻璃巴斯德移液器将固定化的雌雄同体从手表玻璃转移到未播种的 NGM 板上。去除尽可能多的液体, 让多余的液体干燥。
请注意:不要让雌雄同体过度干燥。一旦所有可见的液体都蒸发了, 就开始下一步。 - 将麻醉后的雌雄同体从未播种的 NGM 板转移到与染色雄性的交配点上。在黑暗中孵化 30分钟, 让雄性与雌雄同体交配。
- 交配30分钟后, 立即将雌雄同体安装以进行延时成像, 或将雌雄同体转移到一个新的种子 NGM 板上, 以休息 1小时, 然后再进行成像。
请注意:利用雌雄同体子宫的延时视频来量化精子速度和反转频率。仍然图像的子宫拍摄后1小时交配被用来量化精子的分布, 或 spem 引导。
4. 第2天: 安装用于可视化的蠕虫
-
在h2o中创建具有2% 琼脂糖的安装垫
注: 2% 琼脂糖可以散装制造, 并在玻璃试管中, 并在4°c 下储存。在需要时, 每个都可以在每次使用前进行微波微波, 并储存在热块中, 以防止其凝固。- 要制作安装垫, 请将三个玻璃显微镜幻灯片并排对齐, 并可触摸长边。将两块遮罩胶带放在两个外部幻灯片上。这些带有胶带的外玻璃幻灯片将作为支撑, 因此产生的琼脂糖垫的厚度将是 "两个胶带深"。
- 将融化的2% 琼脂糖 ~ 75μl 放置在中心滑梯上 (这是没有胶带的幻灯片)。立即将一个新的玻璃显微镜幻灯片放在琼脂糖的顶部。这张顶部玻璃幻灯片应垂直于其他幻灯片, 每一端停留在两个支撑幻灯片的磁带上。
- 让琼脂糖变硬 (约 30秒)。小心地从琼脂糖垫上滑动, 取出顶部玻璃滑梯。
- 将 Tet/tri 溶液的10-15μl 放置在2% 琼脂糖垫上。将交配的雌雄同体转移到垫子上。注意转移尽可能少的细菌。
- 在琼脂糖垫上的蠕虫上放置一个盖板。
5. 第2天: 图像采集设置
请注意:任何具有弹性荧光、10倍和60x 目标的直立错误镜, 以及数码相机都可以用来获取精子分布的图像。评估精子速度、定向速度和反转频率所需的软件能够获取延时图像。
-
匹配后1小时的图像采集
- 将幻灯片安装到显微镜舞台上。通过眼片, 使用红色荧光滤光片 (TRITC 过滤器) 的10倍目标扫描琼脂糖垫上的蠕虫。一旦发现蠕虫, 请短暂打开荧光灯, 看看蠕虫是否交配。如果精子在子宫内可见, 切换到60x 目标。
请注意:覆盖物上的60x 目标所产生的压力可能会损坏一些易碎的蠕虫, 导致肠道或性腺从动物身上挤压出来。使用10倍目标扫描成功交配可以最大限度地减少蠕虫暴露在增加的压力下。不要将交配的蠕虫暴露在长时间的荧光灯下。 - 利用微分干涉对比显微镜 (DIC), 定位蠕虫, 使外阴和一个精子。通过聚焦精子的中心来聚焦图像。检查 DIC 和 TRITC 通道的曝光情况。在 DIC 中, 内部蠕虫结构应该是清晰可见的。在 TRITC 中, 单个精子应该是可见的明显的点。
请注意:每一张图片都应该捕捉子宫从外阴到精子。如果子宫太长, 无法容纳在一张图像上, 可以拍摄两个单独的图像。没有必要在相同的曝光水平下拍摄所有图像。然而, 重要的是, 单个精子可以在荧光图像中进行区分和量化。 - 获取每个子宫的 DIC 和荧光图像。
- 重复步骤 5.1.1-5.1.3, 直到所有交配的雌雄同体已被成像。
- 将幻灯片安装到显微镜舞台上。通过眼片, 使用红色荧光滤光片 (TRITC 过滤器) 的10倍目标扫描琼脂糖垫上的蠕虫。一旦发现蠕虫, 请短暂打开荧光灯, 看看蠕虫是否交配。如果精子在子宫内可见, 切换到60x 目标。
-
拍摄延时视频
- 扫描琼脂糖垫, 并定位子宫内含有标记精子的雌雄同体, 如步骤5.1.1 和5.1。2
- 配置软件以获取 DIC 和 TRITC 通道中的延时图像。一般情况下, 延时图像是以15-30 的间隔拍摄的, 每个子宫10-20。
6. 量化
-
定量精子在交配后1小时拍摄的子宫图像上的分布
- 从一端的外阴和另一端的精子, 将子宫分成三分之二。这将表示三个区域。区域 1 (Z1) 包含外阴, 3 区 (Z3) 包含精子。
- 手动统计子宫每三分之一内的精子数量, 并将每个区域中的精子数量报告为整个子宫中精子总数的百分比。下面提供了一个示例。

注: 有时, TRITC 通道图像的信号强度需要调整, 以便图像中捕获的每个精子都可以看到和量化。
-
在延时图像中跟踪精子
请注意:在本文中, 我们使用 nis-元素软件进行分析。在下面的部分中, 我们给出了使用此软件手动跟踪精子的说明 (步骤 6.2.1) 以及开源软件 imagej函斐济 (步骤 6.2.2)。-
用 nis 元素追踪精子
- 打开具有要跟踪的延时序列的. nd2 文件。要开始跟踪, 请右键单击软件并选择"分析控制", 打开 "跟踪" 面板追踪就好
- 在跟踪面板中, 选择 "定义新投资回报率"。通过单击要跟踪的每个精子来定义每个感兴趣的区域 (ROI)。所选精子上将出现彩色标记。选择了所有 Roi 后, 单击"完成" 。
- 一旦确定了 Roi, 就移动到延时系列中的下一帧。将 ROI 标记拖动到图像中精子的新位置。继续这样做, 直到精子不能再被跟踪。将出现虚线, 将通过延时图像的所有帧连接 roi 标记放置的每个位置。
请注意:即使在野生动物中, 也应将第2区中的精子跟踪为第1区和第3区的精子, 它们往往以圆形的方式移动。 - 通过单击 "跟踪" 面板中的"导出" , 将所有可量化的数据 (例如路径长度、时间、xy 位置等) 从跟踪的精子导出到 excel 文档。
-
与斐济一起追踪精子
- 将延时系列中的 TRITC 通道图像转换为. tif 文件。将一个系列中的所有文件保存到一个文件夹中。
- 使用 "生物格式" 导入函数将图像导入到斐济。将图像从一个延时序列导入为一个超堆栈。
- 在斐济通过插件打开 TrackMate 24 追踪我们使用 Track配合特进行手动跟踪。将打开一个对角线框。
- 在斐济工具栏中选择 "跟踪方法" 工具。双击将被跟踪的精子。将出现一个带有虚线的绿色圆圈。这个圆圈可以通过在圆圈内点击并拖动到所需的位置来重新定位。通过同时按alt键和滚动鼠标, 可以更改此圆的大小。
- 设置了跟踪器的大小和位置后, 再次单击圆圈。虚线将变成一条实心绿线。同时命中shift键和l键打开跟踪模式。这将在斐济工具栏中显示。
- 移动到延时系列中的下一帧。要在新框架中设置跟踪精子的新位置, 请将鼠标悬停在新点上, 然后按a键。跟踪器现在将出现在新位置, 并将显示一条连接跟踪器已放置在以前帧中的位置的行。
- 完成跟踪后, 单击 "跟踪方法" 对话框中的"分析"以生成所需的数据。
- 若要计算速度, 请将精子的总路径长度除以经过的时间。
- 要计算矢量速度, 从指向精子的外阴开始, 通过子宫画一条线。测量精子从痕迹开始到结束沿着这条线迁移的距离。把这个距离除以经过的时间。负值表示精子已经从精子中迁移出去。
- 要记录反转频率, 请计算精子轨迹在三个连续的延时帧内产生小于90°的角度的次数。
-
用 nis 元素追踪精子
结果
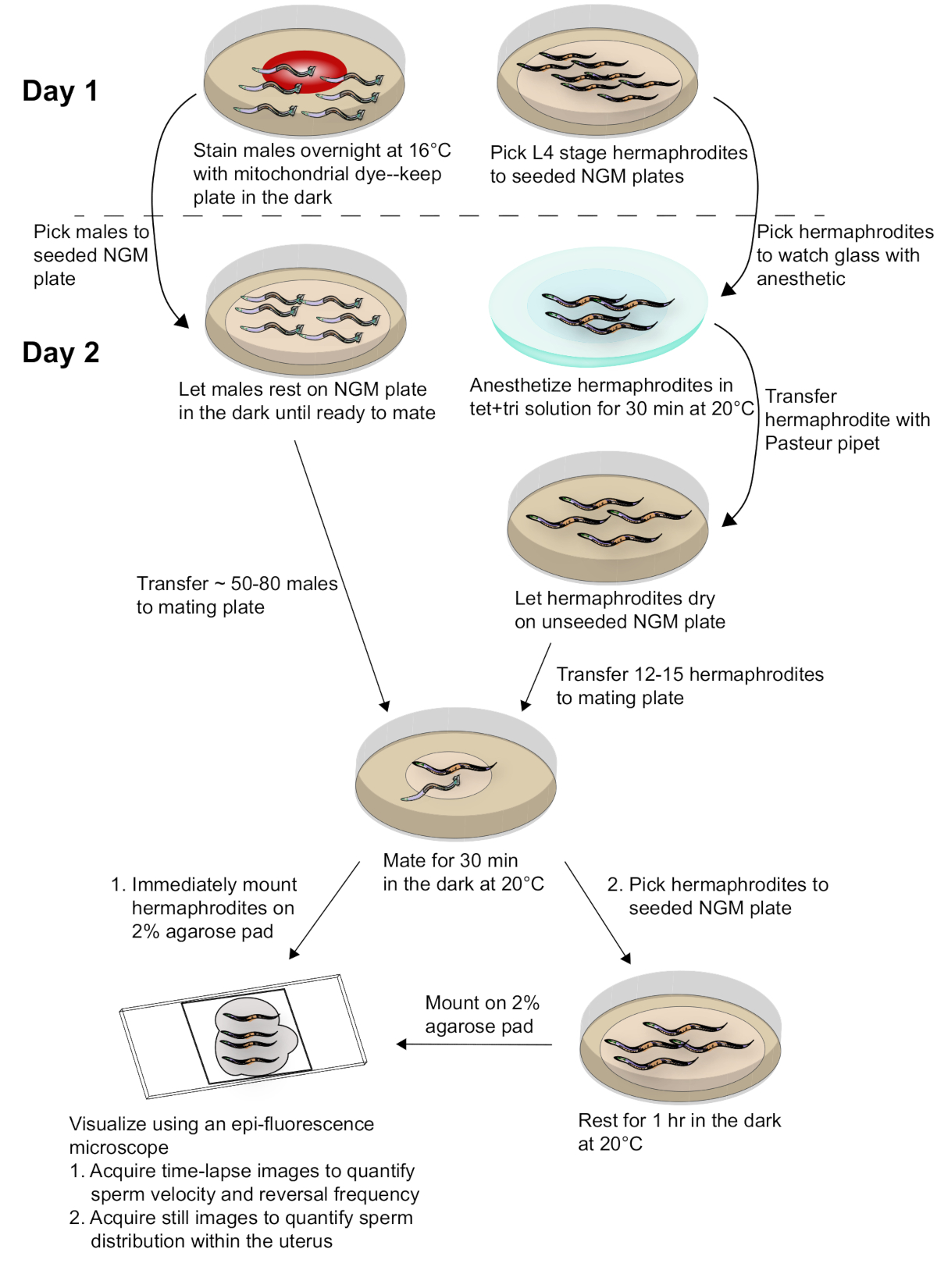
为了得到本文所描述的结果,雾 2(q71)雄性被染成丝片, 并与野人型 n2 雌雄同体交配。图 2提供了该方法的总体方案, 包括蠕虫制备、匹配和分析。正如电影 1所显示的, 成体雌雄同体生殖道有两个手臂, 它们是彼此的镜像。交配时, 标记的精子通过外阴沉积在雌雄同体子宫中。精子在子宫内发育中的胚胎周围向精子细胞移动, 在那里它们被储存到受精。当成虫的生殖细胞发育成卵母细胞时, 近端、最成熟的卵母细胞通过鞘细胞的对比被推入精子细胞。当卵母细胞在精子中时, 就会发生受精。
为了量化精子的分布和通过女性生殖道的迁移, 雌雄同体子宫被划分为三个区域 (电影 1, 图 3 a)。第1区跨越子宫的前三分之一, 从外阴开始。2区跨越子宫的中间三分之一, 而第3区跨越子宫的最后三分之一, 包括精子。使用野生类型、N2 雌雄同体和雾 2(q71)男性进行适当的精子引导, 应导致大约90% 的标记精子到达精子或3区 (图 3b)。 导致精子数量太少 (图 3C, 少于10-15 个精子) 或精子过多 (图 3C, 子宫中充满精子) 的精子不应计算在内。在导致精子少于10-15 的交配中, 3-4 个无赖精子可能会严重扭曲数据。同样, 当子宫完全充满精子时, 精子就不能适当地迁移。精子可能看起来分散在子宫内的一些突变体, 显示不良的精子引导表型。然而, 在这种情况下, 精子不应该填补子宫的每个缝隙, 如图3D 所示。性腺每个臂的定量被认为是一个样本, 或一个 n。
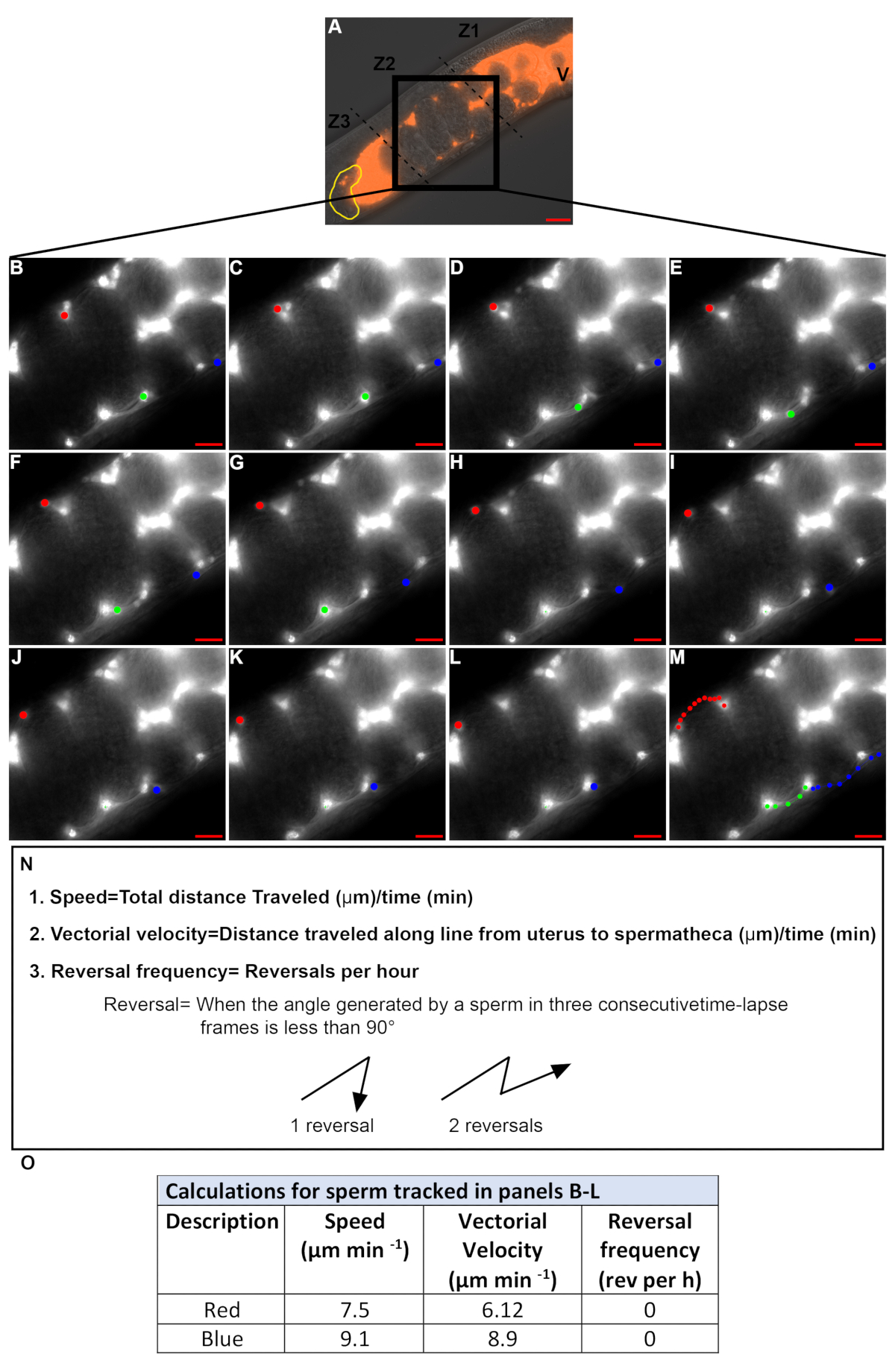
拍摄延时图像来量化精子速度和反转频率。只有第2区的精子才应被跟踪 (图 4a), 因为第1区和第3区 (电影 1) 中的精子即使在野生动物体内也倾向于以圆形模式移动。以15-30 秒的间隔拍摄的时间激光图像通常用于跟踪精子。只有精子, 可以遵循连续帧超过 2.5-3分钟的量化。在图 4B-m中, 以红点和蓝点为标记的精子满足这一标准, 而以绿点为标记的精子则不符合这一标准。因此,图4N 中定义的值被量化为以红点和蓝点为标记的精子 (图 4N), 而以绿点为标记的精子的值没有量化。

图 1:成虫和雄性的卡通.图中标注了主要的生殖结构。请点击这里查看此图的较大版本.
{kind=link}

图 2: 样品制备和数据采集示意图.用丝分裂染色染色的雄性与同步的成体雌雄同体交配。交配后立即拍摄交配雌雄同体的延时图像, 以获取精子速度和反转频率的数据。交配1小时后, 还会拍摄交配雌雄同体的图像, 以评估精子在子宫内的分布情况。有关详细信息, 请参阅文本。请点击这里查看此图的较大版本.
{kind=link}

图 3: 量化精子在雌雄同体子宫内的分布.(A).线虫雌雄同体子宫的示意图。V = 外阴, E = 胚胎, S = 精子, O = 卵母细胞, Z1-z3 = 用于测量精子分布的 Z1-z3 = 区域1-3。(B-D)。DIC + TRITC 仅合并 (左面板) 和 TRITC (右面板) 图像的野生型雌雄同体子宫1小时后, 交配到雾-2(q71) 男性染色的丝染。精子呈红色。黄色轮廓表示精子的位置。刻度杆: 20μm。Z1、Z2、Z3 在 B 中的定量表示每个区域中的百分比精子±标准偏差。C和d中的图像表示导致精子太少 (c) 或过多 (d) 以便量化的匹配。请点击这里查看此图的较大版本.
{kind=link}

图 4: 量化精子在子宫内迁移过程中的速度和反转频率.(A). dic + tritc 合并了含有荧光精子 (红色) 的雌雄同体子宫的图像。V = 外阴, 黄色 = 精子, Z1-z3 = 子宫的三个区域, 黑匣子: 区域2。(B-M)。延时 TRITC 通道图像放大到区域 2 (A 中的黑匣子)。图像是每隔20秒获得一次。在每个图像中跟踪3个单独的精子 (红色、绿色和蓝点)。面板m中的彩色点表示来自b-l的每个精子的路径。刻度杆 = 20μm。精子速度、矢量速度和反转频率的方程和定义。(O). 在b-l中通过红点和蓝点跟踪的精子的速度、矢量速度和反转频率。请点击这里查看此图的较大版本.
{kind=link}

电影 1: 电影精子运动和迁移.一种野生雌雄同体与染成丝染的雾 2 ( q71 )雄配。这部电影是在不同时间间隔拍摄的延时图像的组合。请点击这里观看此视频。(右键单击下载.
讨论
精子在卷曲的女性生殖道中导航和发现卵母细胞的能力对有性生殖至关重要。最近使用内部受精动物精子的研究表明, 它们积极响应各种环境暗示, 包括化学信号、流体流动和温度梯度1,5, 7,9,12. 然而, 这些观察主要是体外实验的结果, 对精子行为和生殖道内的交流了解甚少。获取体内精子迁移和运动数据的主要障碍之一是大多数女性生殖区缺乏可见性。我们在这里描述的使用线虫的方法克服了这一限制。正如代表性的结果所表明的, 透明的表皮允许在一个完整的活生物体中以单个细胞的分辨率对每个精子进行直接的可视化和跟踪。
线虫有两种性别。男性, 与 XO 基因型, 只产生精子, 并在这种方法, 被用作精子捐献者。雌雄同体, 具有 XX 基因型, 是修饰性的雌性。他们的性腺首先在第四幼虫阶段进行精子发生, 并在成年25切换到卵母细胞生成.本方法使用成虫, 其生殖组织为女性生殖道提供了一个模型。在这个分析中, 两性的利用使我们能够识别男性和女性的遗传和分子途径, 这些途径可能会调节精子的引导和活力。结合大量可用于线虫的遗传和分子技术, 该方法可以使人们对精子迁移和活力以及精子和卵母细胞的交流有新的认识。
除了协议部分中提供的细节外, 该协议中的几个关键步骤值得进一步考虑。
蠕虫
雾-2(q71)、他-5(e1490)或他 8(e1489 ) 突变雄性可以用来代替 n2 雄性。这些突变增加了培养中男性的频率 , 但不影响男配或精子功能13。女性, 如雾 2(q71)女性, 可以用来代替雌雄同体。然而, 雌性必须与雄性进行预配, 以便适当地发展卵母细胞。这种交配前受精卵的存在也确保了子宫足够长, 可以正确评估精子的分布。如果正在评估突变体或实验性雌雄同体, 则包括一个对照组。例如, 突变雌雄同体应与野生型 N2 雌雄同体配对, 作为检测中其他变量的对照。用含有质粒的细菌喂养的雌雄同体体, 用于 RNA 干扰检测, 也应使用含有空载体控制的细菌喂养。从精子受精的外阴到受精部位的精子可能会根据子宫中卵子的数量而变化 (即子宫随着卵子数量的增加而扩大)。如果对基因型进行比较, 选择子宫中含有相似数量的卵子的雌雄同体。不要选择含有孵化胚胎或移动幼虫的雌雄同体。可以执行一个时间课程, 以确定应检测雌雄同体的最佳年龄。
采摘雌雄同体和雄性
重要的是, 为这种检测选择的雌雄同体不是来自过度生长或饥饿的板块。食物和信息素提示调节 DAF-7 的表达, Tgf 的同源体。DAF-7 通路已被证明调节 f 系列前列腺素的合成, 这些前列腺素在引导精子进入精子 20方面发挥着重要作用。从过度拥挤或饥饿的盘子中挑选雌雄同体可能会导致精子指导不良与兴趣目标无关。盘子的密度似乎不会影响男性精子。然而, 年龄太小或太老的雄性可能会导致交配效率下降 (即交配板上有足够精子在子宫内量化的雌雄同体的百分比)。1-3岁的成年男性是最佳的此方法23。
抗性雌雄同体
在我们手中, Tetricole 和 Tricaine 的组合确保蠕虫被固定, 并在交配和图像采集过程中保持活着。其他麻醉剂, 如氮化钠, 也可以使用。然而, 氮化钠毒性很强, 条件需要标准化。不建议使用微珠、琼脂糖和微流体室的固定化技术, 因为它们会干扰交配。
男性染色
这份手稿中使用的丝裂染料, MitoTrackerCMXRos, 已被广泛用于标记精子, 以及其他线粒体, 在线虫.用这种丝分裂染料标记的精子是完全功能的, 保持其被激活, 迁移, 受精卵母细胞, 并产生可行的后代26,27。其他线粒体染料, 如罗丹明6g 和 dioc6 已被用来染色线虫线粒体 28,29. 然而, 这些染料的条件需要标准化, 以便在此检测中标记精子。除了线粒体标记机制外, 还可以使用 DNA 污渍 (如 Syto17) 来标记精子进行迁移检测.虽然这些标记技术是相对容易和快速执行, 转基因策略也可以用来产生精子, 表达荧光标记下的精子特定的启动子 31,32。
交配
过厚的交配点可能会降低交配效率。当将雄性和麻醉化的雌雄同体转移到交配点时, 应注意转移尽可能少的细菌。
制作琼脂糖垫并放置盖板
在琼脂糖垫中可以产生气泡。它们可能会在图像采集过程中折射光线, 或者在足够大的情况下导致蠕虫掉落, 从而无法获得图像。同样, 当盖板滑放在琼脂糖垫上时, 可能会沿着雌雄同体产生气泡。这些气泡折射光线并导致图像质量下降。实践将有助于减少气泡的发生。
量化
在量化精子分布时, 不同的 z-面通过蠕虫的子宫可能有轻微的差异, 精子分布。我们发现, 拍摄一张以精子为重点的图像, 可以给我们提供与平均多个 z 节获得的结果相似的可重复结果。我们建议将图像聚焦在精子的中心, 但根据实验者的需要, 可以稍微改变焦平面。然而, 至关重要的是, 所有图像都是以同样的方式拍摄的。此外, 重要的是只计算焦点精子。这是柜台在确定聚焦精子的标准时的自由裁量权。然而, 至关重要的是, 这些标准必须系统地适用于每一种量化的蠕虫。对于精子跟踪, 许多软件提供自动跟踪功能。然而, 我们发现手动跟踪的性能优于软件的自动跟踪算法, 主要有两个原因: 1) 在密闭空间内, 类似大小的原子核丰富, 使得软件很难区分单个精子并为每个精子创建定义的 Roi。2) 当精子进入和脱离焦点时, 它们的强度会发生变化, 使软件很难在长时间内跟踪精子。
披露声明
提交人没有利益冲突。
致谢
我们衷心感谢我们已故的导师迈克尔·米勒博士, 感谢他鼓舞人心和无私的指导, 并创造了这种方法, 作为更好地了解精子和卵母细胞沟通的工具。他的突然去世对他的家人、实验室和科学界来说都是巨大的损失。这项研究得到了国家卫生研究院 (R01GM085105 至 MAM 和 F30HD094446 至 MH) 的支持。内容完全由作者负责, 不一定代表国家卫生研究院的官方观点。
材料
| Name | Company | Catalog Number | Comments |
| Reagents and Material | |||
| 60 mm x 15 mm Petri dish | Fisher | FB0875713A | |
| Agar | Fisher | BP1423-500 | |
| Sodium Chloride | Fisher | S671-3 | |
| Peptone | Fisher | BP1420-500 | |
| Cholesterol | Sigma-Aldrich | C8667 | |
| LB broth, Miller | Fisher | 1426-2 | |
| Escherichia coli strain NA22 | Caenorhabditis Genetics Center (CGC) | NA22 | Either this or OP50 E. coli can be used for C. elegans maintenance and assay. Both may be purchased at the CGC |
| N2 | CGC | N2 | |
| fog-2(q71) | CGC | CB4108 | |
| Platinum wire 0.25 mm dia | Alfa Aesar | 10288 | |
| 5 3/4" Disposable Pasteur pipet | Fisher | 13-678-20A | |
| Watch glass | Fisher | 02-612A | |
| 5 mm Dia. Glass rod | Fisher | 50-121-5269 | |
| MitoTracker CMXRos (Mito-dye) | Fisher | M7512 | Shield from light, store at -20 °C |
| Monopostassium phosphate | Fisher | P285-500 | |
| Disodium phosphate | Fisher | S374-1 | |
| Magnesium sulfate | Fisher | M63-500 | |
| Dimethyl sulfoxide | Fisher | BP231-1 | DMSO |
| Aluminum foil | Fisher | 01-213-102 | |
| Ethyl 3-aminobenzoate methanesulfonate | Sigma | E10521-10G | Tricaine is the common name. Store in aliquotes at -20 °C. |
| Tetramisole hydrochloride | Sigma | L9756-5G | Store in aliquotes at -20 °C |
| Agarose | Fisher | BP1356-100 | |
| Coverslips | Fisher | 12-548-A | 18 x 18-1 |
| Frosted microscope slides | Fisher | 12-552-3 | |
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| 16 °C and 20 °C incubators | Fisher | 97-990E | Same model, set at different temperatures. |
| Upright Microscope with epi-fluorescence illuminator, camera, and 10x and 60x objectives | Nikon | ||
| Software with image acquisition and tracking capabilities | Nikon | NIS-elements AR | |
| Stereo-microscope | Nikon | SMZ800N | Any stereo-microscope that can be used to visualize C. elegans may be used with this protocol |
参考文献
- Boryshpolets, S., Perez-Cerezales, S., Eisenbach, M. Behavioral mechanism of human sperm in thermotaxis: a role for hyperactivation. Human Reproduction. 30 (4), 884-892 (2015).
- Edmonds, J. W., McKinney, S. L., Prasain, J. K., Miller, M. A. The gap junctional protein INX-14 functions in oocyte precursors to promote C. elegans sperm guidance. Developmental Biology. 359 (1), 47-58 (2011).
- Edmonds, J. W., et al. Insulin/FOXO signaling regulates ovarian prostaglandins critical for reproduction. Developmental Cell. 19 (6), 858-871 (2010).
- Espinal-Enriquez, J., Priego-Espinosa, D. A., Darszon, A., Beltran, C., Martinez-Mekler, G. Network model predicts that CatSper is the main Ca(2+) channel in the regulation of sea urchin sperm motility. Scientific Reports. 7 (1), 4236 (2017).
- Hunter, R. H., Nichol, R. A preovulatory temperature gradient between the isthmus and ampulla of pig oviducts during the phase of sperm storage. Journal of Reproduction and Fertility. 77 (2), 599-606 (1986).
- Hussain, Y. H., Guasto, J. S., Zimmer, R. K., Stocker, R., Riffell, J. A. Sperm chemotaxis promotes individual fertilization success in sea urchins. Journal of Experimental Biology. 219 (Pt 10), 1458-1466 (2016).
- Kantsler, V., Dunkel, J., Blayney, M., Goldstein, R. E. Rheotaxis facilitates upstream navigation of mammalian sperm cells. Elife. 3, e02403 (2014).
- Kubagawa, H. M., et al. Oocyte signals derived from polyunsaturated fatty acids control sperm recruitment in vivo. Nature Cell Biology. 8 (10), 1143-1148 (2006).
- Miki, K., Clapham, D. E. Rheotaxis guides mammalian sperm. Current Biology. 23 (6), 443-452 (2013).
- Ting, J. J., Tsai, C. N., Schalkowski, R., Cutter, A. D. Genetic Contributions to Ectopic Sperm Cell Migration in Caenorhabditis Nematodes. G3. 8 (12), 3891-3902 (2018).
- Yanagimachi, R., et al. Chemical and physical guidance of fish spermatozoa into the egg through the micropyledagger, double dagger. Biology of Reproduction. 96 (4), 780-799 (2017).
- Zhang, Y., et al. Generation of Gradients on a Microfluidic Device: Toward a High-Throughput Investigation of Spermatozoa Chemotaxis. PloS One. 10 (11), e0142555 (2015).
- Hoang, H. D., Miller, M. A. Chemosensory and hyperoxia circuits in C. elegans males influence sperm navigational capacity. PLoS Biology. 15 (6), e2002047 (2017).
- Hansen, J. M., Chavez, D. R., Stanfield, G. M. COMP-1 promotes competitive advantage of nematode sperm. Elife. 4, (2015).
- L'Hernault, S. W. Spermatogenesis. WormBook: The Online Review of C. Elegans Biology. , 1-14 (2006).
- Miller, M. A., et al. A sperm cytoskeletal protein that signals oocyte meiotic maturation and ovulation. Science. 291 (5511), 2144-2147 (2001).
- Greenstein, D. Control of oocyte meiotic maturation and fertilization. WormBook: The Online Review of C. Elegans Biology. , 1-12 (2005).
- O'Hagan, R., Wang, J., Barr, M. M. Mating behavior, male sensory cilia, and polycystins in Caenorhabditis elegans. Seminars in Cell & Developmental Biology. 33, 25-33 (2014).
- Hoang, H. D., Prasain, J. K., Dorand, D., Miller, M. A. A heterogeneous mixture of F-series prostaglandins promotes sperm guidance in the Caenorhabditis elegans reproductive tract. PLoS Genetics. 9 (1), e1003271 (2013).
- McKnight, K., et al. Neurosensory perception of environmental cues modulates sperm motility critical for fertilization. Science. 344 (6185), 754-757 (2014).
- Chaudhuri, J., Parihar, M., Pires-daSilva, A. An introduction to worm lab: from culturing worms to mutagenesis. Journal of Visualized Experiments. (47), (2011).
- Stiernagle, T. Maintenance of C. elegans. WormBook: The Online Review of C. Elegans Biology. , 1-11 (2006).
- Chatterjee, I., et al. Dramatic fertility decline in aging C. elegans males is associated with mating execution deficits rather than diminished sperm quality. Experimental Gerontology. 48 (11), 1156-1166 (2013).
- Tinevez, J. Y., et al. TrackMate: An open and extensible platform for single-particle tracking. Methods. , 80-90 (2017).
- Corsi, A. K., Wightman, B., Chalfie, M. A Transparent Window into Biology: A Primer on Caenorhabditis elegans. Genetics. 200 (2), 387-407 (2015).
- Sato, M., Sato, K. Degradation of paternal mitochondria by fertilization-triggered autophagy in C. elegans embryos. Science. 334 (6059), 1141-1144 (2011).
- Wang, Y., et al. Kinetics and specificity of paternal mitochondrial elimination in Caenorhabditis elegans. Nature Communications. 7, 12569 (2016).
- Wolke, U., Jezuit, E. A., Priess, J. R. Actin-dependent cytoplasmic streaming in C. elegans oogenesis. Development. 134 (12), 2227-2236 (2007).
- Mottram, L. F., Forbes, S., Ackley, B. D., Peterson, B. R. Hydrophobic analogues of rhodamine B and rhodamine 101: potent fluorescent probes of mitochondria in living C. elegans. Beilstein Journal of Organic Chemistry. 8, 2156-2165 (2012).
- Singson, A., Hill, K. L., L'Hernault, S. W. Sperm competition in the absence of fertilization in Caenorhabditis elegans. Genetics. 152 (1), 201-208 (1999).
- Wu, J. C., et al. Sperm development and motility are regulated by PP1 phosphatases in Caenorhabditis elegans. Genetics. 190 (1), 143-157 (2012).
- Seidel, H. S., et al. A novel sperm-delivered toxin causes late-stage embryo lethality and transmission ratio distortion in C. elegans. PLoS Biology. 9 (7), e1001115 (2011).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。