
Method Article
Die Entwirrung der Funktion eines bakteriellen Effektor aus einem nicht-kultivierter Pflanzenschädling eine Hefe mit Zwei-Hybrid-Screen
In diesem Artikel
Zusammenfassung
Bakterielle Effektor-Proteine sind wichtig für den Aufbau erfolgreicher Infektionen. Dieses Protokoll beschreibt die experimentelle Identifizierung von proteinösen Bindungspartner eines bakteriellen Effektor-Protein in seiner natürlichen pflanzlichen Wirt. Die Identifizierung dieser Effektor - Wechselwirkungen über Hefe - Zwei-Hybrid - Screens ist ein wichtiges Instrument in entwirren molekularen Pathogenität Strategien werden.
Zusammenfassung
Die Entwirrung der molekularen Mechanismen der Krankheitsmanifestationen ist wichtig, Pathologien und Symptomentwicklung in der Pflanzenwissenschaft zu verstehen. Bakterien haben verschiedene Strategien entwickelt, ihre Wirts Stoffwechsel zu ihrem eigenen Vorteil zu manipulieren. Diese bakterielle Manipulation ist oft verbunden mit schweren Symptomentwicklung oder dem Tod der betroffenen Pflanzen. Die Bestimmung der spezifischen bakteriellen Moleküle verantwortlich für die Host-Manipulation ist ein wichtiger Bereich in der mikrobiologischen Forschung geworden. Nach der Identifizierung dieser Bakterien-Moleküle, die sogenannten "Effektoren", ist es wichtig, ihre Funktion aufzuklären. Ein einfacher Ansatz die Funktion eines Effektor zu bestimmen, ist seine proteinösen Bindungspartner in seinem natürlichen Wirt über eine Hefe-Zwei-Hybrid (Y2H) Bildschirm zu identifizieren. Normalerweise birgt der Host zahlreiche potentielle Bindungspartner , die nicht ausreichend durch ein beliebiges in silico - Algorithmus vorhergesagt werden kann. Es ist daher die beste Wahl, um perform einen Bildschirm mit dem hypothetischen Effektor gegen eine ganze Bibliothek von exprimierten Wirtsproteine. Es ist besonders schwierig, wenn der Erreger wie phytoplasma kultivierbaren ist. Dieses Protokoll bietet eine Schritt-für-Schritt-Anleitung für die DNA-Aufreinigung aus einem phytoplasma-infizierten holzig Wirtspflanze, die Verstärkung des potentiellen Effektor, und die anschließende Identifizierung der molekularen Interaktionspartner der Anlage mit einem Y2H Bildschirm. Obwohl Y2H Bildschirme häufig verwendet werden, gibt es einen Trend, diese Technik zu Biotech-Unternehmen auszulagern, die den Y2H Service zu einem Preis anbieten. Dieses Protokoll bietet Anweisungen, wie ein Y2H in jedem anständig ausgestatteten molekularbiologischen Labor unter Verwendung von Standardlabortechniken auszuführen.
Einleitung
Hefe - Zwei-Hybrid - Screens (Y2H) wurden 1 vor etwa 27 Jahren entwickelt und haben seitdem breite Anwendung in verschiedenen Gebieten verwendet worden , um spezifische Protein-Protein - Wechselwirkungen zu bestimmen , 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 . Die physikalische Wechselwirkung eines bakteriellen Effektor mit einem Host-Zielprotein ist oft Voraussetzung für die funktionelle Manipulation dieses Zielproteins. Diese Interaktionen analysieren hat in den Fokus der vielen verschiedenen Bereichen der Infektion 12 Biologie bewegt, 13, 14, 15,"> 16, 17, 18. Das Prinzip des Y2H Bildschirm ist , dass die Wechselwirkung der beiden Proteine in die Rekonstitution eines funktionellen Transkriptionsfaktors führt, der die Expression von Reportergenen antreibt. Die bekannte Effektor (der" Köder ") translatorisch verschmolzenen an die DNA-Bindungsdomäne (DBD) des Transkriptionsfaktors, und die möglichen Wechselwirkungspartner (die "Beute") an die Aktivierungsdomäne (AD) der jeweiligen Transkriptionsfaktor fusioniert ist. Da der Wechselwirkungspartner unbekannt ist, eine Bibliothek von potentiellen Interaktoren in die so genannte geklont "Beute-Bibliothek". Normalerweise diese Bibliothek durch Klonieren von cDNAs in einen geeigneten Beute-Vektor Expressionsplasmid codierend das AD hergestellt ist, die mit dem Köder-Vektor kodiert DBD. bei einer Wechselwirkung kompatibel ist, die rekonstituiert Transkriptionsfaktoren die Expression von Reportergenen, induzieren, die im allgemeinen das Wachstum von Hefe-Auswahl zu ermöglichen. dieIdentifizierung der Interaktionspartner wird durch die Sequenzierung der Beute Plasmid mit Plasmid-spezifischen Primer erreicht.
Bakterien haben verschiedene Strategien entwickelt , zu manipulieren und ihre Host-Stoffwechsel verwerten oder Abwehrmechanismen zu umgehen und Effektor - Molekülen über verschiedene Bakterien Sekretion Systeme 19, 20, 21 absondern. die Bindungspartner dieser bakteriellen Effektoren bestimmen, ist daher der erste Schritt in der Stellweges in dem Host zu identifizieren. Dies führt zu einem verbesserten Verständnis der spezifischen Mechanismen Pathogenität.
Y2H Bildschirme werden durchgeführt , bakterielle Effektor Interaktionen während phytoplasmoses zu identifizieren, und oft ist ein entscheidender Y2H anfänglichen Experiment zur weiteren Charakterisierung der molekularen Mechanismen Pathogenität in phytoplasma Forschung 22, 23. Doch das trafin den meisten Publikationen hod Beschreibung ist eher selten, und diese Techniken werden oft an Biotech-Unternehmen ausgelagert. Um die Aufmerksamkeit auf die Durchführbarkeit dieses Verfahrens zeichnen, dieses Protokoll liefert eine Schritt-für-Schritt-Informationen Interaktionspartner eines bakteriellen Effektor-Molekül in seinem natürlichen Wirt identifizieren.
Trotz der breiten Nutzung und den schnellen Vorlauf Ansatz von Y2H Bildschirme, darf nicht vergessen werden, dass bestimmte Interaktionen nicht in der Hefe-System auftreten könnten. Dies ist aufgrund der Tatsache , dass die Hefezelle als eine Art von "in vivo Reaktionsbehälter" mit bestimmten biologischen Einschränkungen verwendet. Mehrere Autoren haben die Vor- und Nachteile von Y2H und seine Derivate 11, 24, 25, 27 26, gerichtet. Allgemeine Überlegungen sind beispielsweise, dass die Hefezelle nicht die entsprechende bieten könnteGenexpression, (post-) translationale oder translocational Bedingungen für die jeweiligen Proteine untersucht. Dies kann auf dem Bildschirm zu falsch negativen Ergebnissen führen. Positive Wechselwirkungen können wiederum Artefakte und in der natürlichen Situation (dh im Fall von Effektoren in den geeigneten Wirt) auftreten , möglicherweise nicht. Es ist daher unerlässlich, um die Wechselwirkungen von der heterologen Hefe-Expressionssystem mit einem unabhängigen Interaktionstest in einem eng verwandten biologischen Systems zu bestätigen.
In dieser Studie wurden identifiziert die Bindungspartner der Effektor ATP_00189 aus dem nicht-kultivierbaren Pflanze Erreger Candidatus Phytoplasma mali (P. mali). Die Ergebnisse liefern wichtige Einblicke in die molekularen Mechanismen , die Symptomentwicklung von Apple proliferation zugrunde liegenden 28, eine Krankheit , die hohe wirtschaftliche Verluste in den betroffenen Apfelanbauregionen in Europa 29 verursacht.
Protokoll
1. Sammeln Wurzel und Blattproben von infizierten Apfelbäume
Hinweis: erregerspezifische DNA kann aus Wurzeln oder Blättern gereinigt werden. Der folgende Abschnitt enthält ein Protokoll für die Probenahme von beiden.
- Für die DNA-Präparation
- Probenentnahme und Vorbereitung von den Wurzeln
- Identifizieren Sie infizierte Bäume mit "Apple proliferation" -spezifische Symptome 30, 31 und Kontrolle Bäume , die symptomfrei sind. Mit sauberen Gartenschere schneiden Wurzelproben, die einen Durchmesser von 0,5 haben - 1 cm und eine Länge von etwa 5 cm. Cut-Proben von drei verschiedenen, entfernten Stellen des Wurzelsystems, und sie in angemessener Weise gekennzeichnet Plastiktüten.
- Halten Sie die Proben in einer Kühlbox mit gekühltem Wärmepackungen und lagern Sie sie im Kühlschrank bei 4 ° C bis zur Weiterverarbeitung.
Hinweis: Root-Architektur und Struktur in Bezug auf den physiologischen Zustand o variierenf Baum. Es ist wichtig, an drei verschiedenen Standorten zu probieren und nicht zu dünnen Würzelchen zu nehmen. Die Proben können für mehrere Tage bei 4 ° C gelagert werden, aber längere Lagerzeiten die Gefahr des Formens zu erhöhen. Moldy Proben können nicht für die DNA-Präparation verwendet werden.
- Halten Sie die Proben in einer Kühlbox mit gekühltem Wärmepackungen und lagern Sie sie im Kühlschrank bei 4 ° C bis zur Weiterverarbeitung.
- Spülen Sie die Wurzelproben mit Wasser, um den Boden zu entfernen. Setzen Sie sie in eine sterile Petrischale und mit einem sterilisierten Skalpell die Wurzel Epidermis und die Rinde zu entfernen.
- Reinigen Sie das Skalpell mit einem sauberen, fusselfreien Tuch abwischen, tauchen sie in 70% (v / v) Ethanol - Wasser - Lösung und Wärme sterilisieren es eine offene Flamme über (zB Bunsenbrenner). Rubbeln Sie an der Bast mit dem Skalpell, hacken sie in kleine Stücke, und aliquote 30-100 mg des gehackten Phloem in eine sterile 2,0 ml Reaktionsröhrchen. Lagern Sie die Proben bei -80 ° C für mehrere Monate.
- Identifizieren Sie infizierte Bäume mit "Apple proliferation" -spezifische Symptome 30, 31 und Kontrolle Bäume , die symptomfrei sind. Mit sauberen Gartenschere schneiden Wurzelproben, die einen Durchmesser von 0,5 haben - 1 cm und eine Länge von etwa 5 cm. Cut-Proben von drei verschiedenen, entfernten Stellen des Wurzelsystems, und sie in angemessener Weise gekennzeichnet Plastiktüten.
- Probenentnahme und Vorbereitung von den Blättern
- Identifizieren Sie eine infizierte und eine asymptomatische Kontrollbaum, eins in Schritt 1.1.1.1, beschrieben und zehn ungeschoren Blätter pro Baum pflücken. Ein Baum pro Bedingung ist ausreichend.
- Spülen Sie die Blätter mit Wasser und reinigen Sie die Oberflächen um 70% Spritzen (v / v) Ethanollösung. Legen Sie die Blätter in eine sterile Petrischale und sezieren die midrib (zentrale Vene) jedes Blatt mit einem sterilen Skalpell. Entfernen Sie die Epidermis und Rinde aus dem midrib, schneiden Sie den midrib in kleine Stücke, und aliquote 100 mg des midrib Gewebe in eine sterile 2,0 ml-Reaktionsröhrchen. Verwenden Sie die Proben sofort für die DNA-Herstellung oder bei -80 ° C für mehrere Monate.
Hinweis: Es ist bequem, das Pflanzenmaterial in Portionen von 100 mg zu aliquotieren und schließlich diese einzufrieren für die Lagerung. Jedes Aliquot kann direkt für die DNA-Präparation verwendet werden. Mit einem Gewicht tiefgefrorene Pflanzenmaterial ist umständlich, da das gefrorene Pflanzenmaterial Klumpen zu bilden neigt.
- Probenentnahme und Vorbereitung von den Wurzeln
2. (CTAB) -basierten DNA-Präparation
Hinweis: Die DNA kann für Pflanzenmaterial mit einer beliebigen Spalte-basierten DNA-Reinigungsverfahren gereinigt werden. In diesem Abschnitt wird eine CTAB-basiertes Verfahren zur DNA-Reinigung beschrieben. DNA - Reinigung basiert auf einem modifizierten Protokoll an anderer Stelle 32 beschrieben durchgeführt.
- Bereiten Sie die folgenden Puffer:
- CTAB - Puffer: Man löst 1% w / v Cetyltrimethylammoniumbromid (CTAB, M r: 364,45 g / mol), 100 mM Trishydroxymethylaminomethan (Tris, M r: 121,14 g / ml), 1,4 M Natriumchlorid (NaCl, M r: 58,44 g mol /) und 20 mM Ethylendiamintetraessigsäure-Dinatriumsalz (NaEDTA, M r: 372,24 g / mol) in Wasser und stellen sie sie auf pH 8,0.
- N-Lauroylsarcosin - Puffer: Man löst 10% w / v N-Lauroylsarcosin - Natriumsalz 33 (M r: 293,38 g / mol), 100 mM Tris und 20 mM NaEDTA in Wasser und den pH - Wert auf 8,0 einzustellen.
- Tris-EDTA (TE) Puffer: Man löst 10 mM Tris und 1 mM NaEDTA in Wasser und autoclave die Lösung.
- Ammonium - Acetat - Lösung: Eine 5 M Ammoniumacetat (M r: 77,0825 g / mol) Lösung in Wasser und im Autoklaven bei 120 ° C und 1,2 bar für 20 min.
- Mischungs 30-100 mg von frischen oder gefrorenen zerkleinerten Pflanzenmaterial (Schritt 1.1.2.2.) Mit 300 & mgr; l CTAB-Puffer, 30 ul N-Lauroylsarcosin-Puffer 6 & mgr; l Proteinase K (10 ug / ul) und 12 ul 2-Mercaptoethanol 34 (M r: 78,13 g / mol). Schütteln für 60 min bei 60 ° C. Lassen Sie die Mischung auf Raumtemperatur abkühlen, bevor Sie fortfahren.
- In 360 & mgr; l Ammoniumacetatlösung und kräftig mischen. Lassen Sie die Lösung für 5 min sitzen, und dann für 10 min bei Raumtemperatur bei 15.000 xg zentrifugiert. Den Überstand in ein sauberes Fläschchen und fügen 720 ul eiskaltem Isopropanol 35. Auszufällen die DNA für mindestens 30 min bei -20 ° C.
- Zentrifugieren der Mischung bei 15.000 × g für 30 min bei 4° C und den Überstand verwerfen. Waschen Sie das Pellet mit 500 ul eiskaltem 70% Ethanol und Spin es bei 4 ° C bei 15.000 × g für 5 min nach unten.
- Entsorgen Sie den Ethanolüberstand und sofort tauchen Sie das Fläschchen auf einem sauberen Papiertuch Resttropfen zu entfernen. Stellen Sie sicher, so viel Flüssigkeit wie möglich zu entfernen. Der Luft trocknen das Pellet für 15-20 Minuten oder für 2-3 Minuten in einem Vakuum-Konzentrator-Zentrifuge. Nicht zu trocknen das Pellet durch übermäßige Erwärmung oder verlängert Verdampfungszeiten.
- Resuspendieren des Pellets in 700 ul TE-Puffer und schließlich auf 37 ° C erwärmen Auflösen zu erleichtern. Fügen Sie 10 & mgr; l RNAse (10 ug / ul) und Inkubation für 15 min bei 37 ° C, bei 300 Upm Schütteln.
- In 700 ul Chloroform: Isoamylalkohol 36 24: 1 und gut mischen. Zentrifugieren Sie die Mischung bei 20.000 × g für 10 min bei 4 ° C und übertragen Sie die obere Phase des Stand in ein neues, sauberes Fläschchen.
- Auszufällen die DNA im Überstand durch Zugabe von700 ul 2-propanol 35 (Isopropanol, M r: 60,1 g / mol) und für mindestens 30 min bei -20 ° C inkubiert. Zentrifugieren Sie die Mischung bei 12.000 × g für 30 min bei 4 ° C und den Überstand verwerfen.
- Waschen Sie das Pellet mit 500 ul 70% Ethanol und Zentrifugation bei 12000 × g für 5 min bei 4 ° C. Trocknen Sie das Pellet wie in Schritt 2.5 beschrieben. Man löst es in 50 ul TE.
3. Detecting Candidatus Phytoplasma mali-spezifische DNA durch PCR
- Verdünnen Sie die DNA aus dem Pflanzenmaterial gereinigt 1:10 in Nuklease-freies Wasser und eine PCR mit erregerspezifischen Primern 37 laufen.
- Überprüfen Sie die Anwesenheit von P. mali-spezifische DNA in dem Pflanzenmaterial durch eine quantitative PCR - Protokoll mit Primer und spezifische Sonden für P. mali phytoplasma 37 verwendet wird . Prüfen Sie die Abwesenheit der anderen 16SrX phytoplasma Mitglieder , indem sie mit den jeweiligen Sonden für Ca die gleiche Durchführung der PCRnd. P. prunorum und für Cand. P. pyri DNA 37.
HINWEIS: Die DNA von einem nicht-infizierten Baum sollte auch getestet werden, um die Abwesenheit eines Pathogens in dieser Steuerung zu bestätigen. Bei diesem Schritt ist es wichtig, dass die DNA von anderen eng verwandten phytoplasma Spezies in der Probe nicht vorhanden ist, da dies das Risiko einer Verstärkung des Effektors von einer anderen Spezies als phytoplasma P. mali erhöhen würde. Eine gemischte Infektion mit einem anderen eng verwandten P. mali 16SrX Gruppe phytoplasma wird durch die Analyse der Probe mit den jeweiligen Sonden ausgeschlossen. Da die erwarteten Ergebnisse sollten für andere 16SrX phytoplasma negativ sein, ist es wichtig, die richtige positive Kontrollen umfassen, die PCR-Wirksamkeit zeigen.
- Überprüfen Sie die Anwesenheit von P. mali-spezifische DNA in dem Pflanzenmaterial durch eine quantitative PCR - Protokoll mit Primer und spezifische Sonden für P. mali phytoplasma 37 verwendet wird . Prüfen Sie die Abwesenheit der anderen 16SrX phytoplasma Mitglieder , indem sie mit den jeweiligen Sonden für Ca die gleiche Durchführung der PCRnd. P. prunorum und für Cand. P. pyri DNA 37.
4. Die Verstärkung des Potenzials Effektor Gene und Subklonierung in die Y2H Bait-Vektor
- Amplify das phytoplasmal Gen atp_00189 (nicht das Signalpeptid Teil umfasst) von P. MALI unter Verwendung der Primer 5-TCTCCTCCTAAAAAAGATTCTA-3 (vorwärts) und 5-TATTATTTATCTTTATTTTTTTCCTT-3 (rückwärts), mit Restriktionsschnittstellen für EcoRI und SalI am 5'- und den 3' - Enden, respectively. Reinige das PCR-Produkt mit einer spaltenbasierten DNA-Reinigungsverfahren.
- Klonen Sie das Amplifikat in den Y2H Bait - Vektor pLexA-N über EcoRI und Sall Ligatur.
HINWEIS: Hier 100 ng EcoRI und Sall linearisiert pLexA-N - Vektor mit 15 ng ähnlich verdaut atp_00189 PCR - Amplikon und 1 U T4-Ligase kombiniert wurde. Die Ligation wurde in einem Puffer durchgeführt , enthaltend 40 mM Tris-HCl (pH 7,9 bei 25 ° C), 10 mM MgCl 2, 10 mM Dithiothreitol (DTT, M r: 154.25 g / mol) und 0,5 mM Adenosintriphosphat (ATP, M r: 507,18 g / mol) in einem Gesamtvolumen von 10,0 & mgr; l bei 16 ° C über Nacht (14-20 h). Analysieren Sie die DNA aus dem nicht - infizierten Baum mit den gleichen Primer unspezifische Primer , um auszuschließen , die Bindung an Malus x domestica </ Em> DNA. - Transformieren 1 & mgr; l der Ligationsmischung in E. coli Zellen und elektro auswählen , für Kanamycin-resistente Klone auf Luria-Bertani (LB) Platten mit 50 ug / ml Kanamycinsulfat 38 ergänzt.
- Reinige die Plasmid - DNA aus den ausgewählten Bakterienklone mit einer spaltenbasierten Plasmid - Mini Vorbereitung Kit folgenden Anweisungen des Herstellers, und überprüfen Sie die erfolgreiche Integration und Ausrichtung des Einsatzes durch Sequenzierung 39.
5. Test zur Selbstaktivierung (Auto-Aktivierung) des Potential Effektor-Protein
- Bereiten Sie die folgenden Puffer und Wachstumsmedien:
HINWEIS: Die Nomenklatur für Hefe-Selektionsplatten Silbentrennung die Abkürzungen der fehlenden Aminosäuren in dem jeweiligen Medium mit einem "-" vor der Abkürzung. Zum Beispiel SD-trp-leu-his-Platten enthalten alle Aminosäuren, aber trp, leu und sein.- 10x Dropout-Mix: Wiegen200 mg L-Arginin - Monohydrochlorid (M r: 210,66 g / mol), 300 mg L-Isoleucin (M r: 131,17 g / mol), 269 mg L-Lysin - Monohydrat (M r: 164,21 g / mol), 200 mg L-Methionin (M r: 149,21 g / mol), 500 mg L-Phenylalanin (M r: 165,19 g / mol), 2 g L-Threonin (M r: 119,12 g / mol), 300 mg von L-Tyrosin (Mr: 181,19 g / mol), 200 mg L-uracil (112.09 g / mol) und 1,5 g L-Valin (M r: 117,15 g / mol). Man löst sie in 1 l doppelt destilliertem Wasser und im Autoklaven die Lösung. Die Lösung wird bei 4 ° C.
HINWEIS: Die jeweilige Dropout-Mix können auch gekauft vorgemischt werden. - 10x L-Adenin Ergänzung: Wiegen 100 mg L-Adenin Hemisulfatsalz (ade, M r: 184,17 g / mol) und löst ihn in 50 ml doppelt destilliertem Wasser. Filter sterilisieren, die Lösung mit einem 0,22-um Porenfilter.
- 10x L-Histidin Ergänzung: Wiegen 100 mg L-Histidin - Monohydrat (sein, M r : 209,63 g / mol) in 50 ml doppelt destilliertem Wasser und Filter sterilisieren mit einem 0,22-um Porenfilter.
- 10x L-Leucin - Ergänzung: Einwiegen von 500 mg L-Leucin (leu, M r: 113,17 g / mol) in 50 ml doppelt destilliertem Wasser und Filter sterilisieren mit einem 0,22-um Porenfilter.
- 10x L-Tryptophan Ergänzung: Wiegen 100 mg L-Tryptophan (trp, M r: 204,23 g / mol) in 50 ml doppelt destilliertem Wasser und Filter sterilisieren mit einem 0,22-um Porenfilter.
- SD-trp-leu-his-Ade-Medium und Platten: Man löst 0,67% w / v Yeast Nitrogen Base (ohne Aminosäuren) und 2% w / v D-Glucose - Monohydrat (M r: 198,17 g / mol) in doppel- destilliertes Wasser und Autoklaven. Für Agarplatten, fügen Sie 2% w / v Agar (mikrobiologische Qualität) vor dem Autoklavieren.
- Zur Herstellung von selektiven Platten, fügen 1x der Aminosäure-Stammlösung zur autoklaviert Sd-trp-leu-his-ade Medium. Wenn die Platten vorbereiten, stellen Sie sicher, dass der Agar auf ~ 50 ° C abgekühlt wird, bevor die Zugabedie Aminosäuren. Halten Sie die Stammlösungen steril.
- YPAD Medium: Man löst 1% w / v Hefeextrakt, 2% w / v Pepton (mikrobiologische Qualität), 0,004% w / v Adenin Hemisulfatsalz (M r: 184,17 g / mol) und 2% w / v Glucose Monohydrat Wasser und Autoklaven doppelt destilliert. Für YPAD-Agar-Platten, fügen Sie 2% w / v Agar vor dem Autoklavieren. 2x YPAD, verwenden Sie zweimal die Konzentrationen der oben genannten Zutaten vorzubereiten.
Anmerkung: (Optional) Aufgrund der hohen Konzentration von Glucose in dem Medium, 2x YPAD Medium dunkelbraun nach Autoklavierung wird. Als Alternative einer 40% (w / v) Glucose-Stammlösung durch Hindurchleiten durch ein 0,22-um-Filter hergestellt und sterilisiert werden kann. Der Filter-sterilisierte Glucosestammlösung wird dann zu dem Autoklaven behandelt 2x YPAD (fehlende Glucose) unter sterilen Bedingungen. Um Volumenfehler zu vermeiden, muss die 2x YPAD hergestellt werden, wenn man bedenkt, dass eine 10-fach Glucoselösung anschließend zugegeben wird. Das bedeutet, dass beispielsweise anstelle von 1 L of Medium nur 900 ml vorbereitet (mit allen Zutaten außer Glukose) und autoklaviert und dann 100 ml Glucose-Stammlösung zugegeben.
- 10x Dropout-Mix: Wiegen200 mg L-Arginin - Monohydrochlorid (M r: 210,66 g / mol), 300 mg L-Isoleucin (M r: 131,17 g / mol), 269 mg L-Lysin - Monohydrat (M r: 164,21 g / mol), 200 mg L-Methionin (M r: 149,21 g / mol), 500 mg L-Phenylalanin (M r: 165,19 g / mol), 2 g L-Threonin (M r: 119,12 g / mol), 300 mg von L-Tyrosin (Mr: 181,19 g / mol), 200 mg L-uracil (112.09 g / mol) und 1,5 g L-Valin (M r: 117,15 g / mol). Man löst sie in 1 l doppelt destilliertem Wasser und im Autoklaven die Lösung. Die Lösung wird bei 4 ° C.
- Lithiumacetat-vermittelte Hefetransformation zum Testen Selbstaktivierung
- Streak die Saccharomyces cerevisiae Reporterstamm NMY51 40 (NMY51: MATahis3Δ200 trp1-901 leu2-3,112 ade2 LYS2-: :( LexAOp) 4-HIS3 ura3: :( LexAOp) 8-lacZ ade2: :( LexAOp) 8-ADE2 GAL4) von einem gefrorene Glycerolstammlösung auf einem YPAD Agar-Platte und Inkubation für 48 h bei 30 ° C. Pick-Kolonien von dieser Platte und impfen 50 ml YPAD Medium. Lassen Sie die Hefe wachsen und die OD 600 messen regelmäßig das Wachstum zu überwachen.
- Lassen Sie die Hefe bis zu einer endgültigen OD 600 von 0,5-0,8 wachsen. Pelletieren Sie die Zellen, indem sie bei 700 xg für 5 min Zentrifugieren und Resuspendieren sie in 2,5 ml bidestilliertem, autoklaviert Wasser.
- Bereiten Sie sechs Aliquote mit 100 & mgr; l Hefesuspension und fügen 240 ul 50% W / v Polyethylenglykol 4000 (PEG, M r: 3.500-4.000 g / mol), 36 ul 1 M Lithiumacetat - Dihydrat (LiOAc, M r: 102,02 g / mol) und 25 ul 2% w / v Lachsspermien-DNA (aufgelöst). Bereiten Sie alle Reagenzien mit doppelt destilliertem, sterilem Wasser.
- Beschriften Sie die sechs Fläschchen mit dem Hefe - Suspension von "a" bis "f" und fügen Sie den folgenden Plasmidvektor - DNA: Negativkontrollen: (a) 1,5 ug Köderplasmid mit dem Effektor (pLexA-N-atp00189 28), (b) 1,5 ug leere Beute Plasmid pGAD-HA 41, (c) 1,5 ug leerer Köder Vektor pLexA-N 41, und (d) 1,5 ug leerer Köder Vektor pLexA-N und 1,5 ug leere Beute Plasmid pGAD-HA; Positivkontrollen: (e) 1,5 ug Positivkontrolle Köderplasmid DNA pLexA-p53 41 und 1,5 ug positiv Beute Plasmid pACT-larget 41; und die Selbstaktivierung Test: (f) 1,5 ug baes Plasmid mit dem Effektor (pLexA-N-atp00189) und 1,5 ug leere Beute Plasmid pGAD-HA.
- Mischen Sie die Reaktionen heftig und brüten sie für 45 min bei 42 ° C in einem Wasserbad. Pellet die Zellen für 5 Minuten bei 700 · g und den Überstand verwerfen. Resuspendieren der Zellen in 250 & mgr; l von 0,9% (w / v) Natriumchloridlösung (NaCl, M r: 48,44 g / mol) und verteilt 50 & mgr; l auf den folgenden selektiven Platten: SD-trp, Sd-leu, SD-trp- leu, SD-trp-leu-his, und SD-trp-leu-his-ade.
- Inkubieren der Platten für 3 - 4 Tage bei 30 ° C. Nach dem ersten Tag der Inkubation, dichten die Platten mit Kunststoffparaffinfilm die Platten vor dem Austrocknen zu verhindern.
- Überprüfen Sie das Wachstum der Hefe auf den Platten.
6. Testen Sie die Expression des Effektor
- Testen Sie den Ausdruck des Effektor durch Western - Blot - 42 mit einem Antikörper gegen das Lex-A - Tag , das am N-Terminus des Effektor gekoppelt ist , wennvon pLexA-N ausgedrückt.
Hinweis: Bedenken Sie, dass die translational fusioniert Lex-A-Tag etwa 24 kDa auf das tatsächliche Gewicht des Proteins von Interesse hinzufügt. Dies ist wichtig, wenn das Protein eine Größe in dem Western-Blot identifiziert.
7. Der Y2H Bildschirm
- Streak die pLexA-atp00189 (Effektor) transformierten NMY51 auf einem frischen SD-trp Platte und lassen Sie es für 2 wachsen - 3 Tage bei 30 ° C, bis rote Kolonien erscheinen.
- Impfen 3 ml SD-trp-Medium in einem kleinen mit einer roten Kolonie von der Agarplatte Anschüttelflasche und Inkubation über Nacht bei 30 ° C bei 120 unter Schütteln - 150 Umdrehungen pro Minute.
- Impfen 20 ml SD-trp in Schüttelkolben mit 1 ml der Übernachtkultur und lassen Sie es für 8 h wachsen.
- Stellen Sie die Kultur 0,2 600 = bis zu einer OD SD-trp Medium durch Zugabe und impfen 2x 100 ml in Schüttelkolben mit 10 ml der Starterkultur jeder. Wachsen über Nacht bei 30 ° C schüttelnd.
Hinweis: Stellen Sie sicher , dass die Hefe wächst nicht 6 bis OD00> 0,5 und verdünnt mit frischem SD-trp. - Messen Sie die OD 600 und das Pellet 120 OD 600 "Einheiten" . Wenn zum Beispiel eine OD 600 von 1,2 gemessen wird, Spin - down 100 ml, den Überstand verwerfen und das Pellet in 800 ml vorgewärmten 2x YPAD in einem Schüttelkolben mit einem magnetischen Rührstab.
- Spin down eine 2-ml-Aliquot, entfernen Sie den Überstand und das Pellet in Wasser. Stellen Sie sicher , dass die OD 600 der Hefesuspension zwischen 0,15 und 0,2 ist. Wenn nicht, stellen Sie mit 2x YPAD oder mehr Hefe aus der Nachtkultur hinzuzufügen.
Anmerkung: 2x YPAD dunkelbraun und die Ergebnisse der OD Messung beeinflussen können. Somit ist es notwendig, die Hefezellen in Wasser zu suspendieren, bevor die OD zu bestimmen. Als Alternative kann 2x YPAD durch Sterilisieren Glukose separat hergestellt werden, wie in dem Schritt 5.1.8 Note beschrieben, die die dunkle Färbung des Mediums verhindert.
- Spin down eine 2-ml-Aliquot, entfernen Sie den Überstand und das Pellet in Wasser. Stellen Sie sicher , dass die OD 600 der Hefesuspension zwischen 0,15 und 0,2 ist. Wenn nicht, stellen Sie mit 2x YPAD oder mehr Hefe aus der Nachtkultur hinzuzufügen.
- Inkubieren Sie die verbleibenden Hefekultur in einer Appropriately Größe Schüttelkolben (800 ml in einem 2 L Schüttelkolben oder dividieren 2 x 400 ml in zwei 1 l Schüttelkolben) und Inkubation bei 30 ° C, 120 bis 150 Umdrehungen pro Minute. Messen der OD 600 ungefähr alle 1,5 h bis zu einer OD 600 von 0,6 erreicht ist (Dauer 4 - 6 h). In der Zwischenzeit bereiten die Lösungen im nächsten Schritt beschrieben.
- Lithiumacetat-vermittelte Transformation
- Löse 2% w / v Lachssperma-DNA in Wasser und kochen 500 & mgr; l für 5 min in einem Wasserbad bei 100 ° C. Das Röhrchen auf Eis für 2 Minuten und wiederholen Sie den Erhitzungsschritt. Halten Sie die DNA auf Eis bis zur weiteren Verwendung.
- Bereiten Sie die folgenden Mischungen:
- TE / LiOAc Mischung: Mischen 3,08 ml 1 M LiOAc, 3,08 ml 100 mM Tris / 10 mM EDTA (pH-Wert: 7,5), und 21,84 ml sterilem doppelt destilliertem Wasser.
- PEG / LiOAc mix: Kombinieren 4,2 ml 1 M LiOAc, 4,2 ml mM Tris / 10 mM EDTA (pH: 7,5) und 33,6 ml 50% (w / v) PEG 4000.
- Zentrifugieren Sie die 800-ml Hefekultur (OD 600 = 0,6)bei 700 xg für 5 min, um die Zellen zu pelletieren. Entfernen Sie den Überstand und das Pellet in 200 ml sterilem doppelt destilliertem Wasser. Pellet die Zellen wieder bei 700 g für 5 min und den Überstand verwerfen. Hinweis: Bei Bedarf teilen sich die Suspension in mehrere Fläschchen für die Zentrifugation. Die Flüssigkeitsvolumina in 7.7.3. und 7.7.4. beziehen sich auf das gesamte Pellet abgeleitet aus 800 ml Suspension.
- Das Pellet in 16 ml TE / LiOAc Mix (siehe Schritt 7.7.1.1); Spin bei 700 xg für 5 min nach unten und den Überstand verwerfen. Das Pellet in 9,6 ml TE / LiOAc Mix.
- Bereiten Sie die folgende Reaktion mischt in geeigneter Größe Reaktion Polypropylen Behälter: 12 Flaschen mit 7 ug pGAD-HA-cDNA-Bibliothek Vektor, 100 & mgr; l von 2% Lachsspermien-DNA (siehe Schritt 7.7) und 2,5 ml PEG / LiOAc Mix. In 600 ul Hefe Zellsuspension aus Schritt 7.10 zu jedem der 12 Fläschchen und 1 min kräftig mischen.
- Inkubieren der Reaktionsmischung für 45 min bei 30 ° C in einem Wasserbad eind energisch alle 15 min mischen. In 160 & mgr; l DMSO zu jedem Fläschchen und kräftig mischen. Inkubieren der Mischung für weitere 20 min bei 42 ° C.
- Pellet die Zellen bei 700 xg für 5 min, den Überstand verwerfen und resuspendieren jedes Pellet in 3 ml 2x YPAD. Pool alle Zellen (mit insgesamt 36 ml aus den 12 Fläschchen) in einem 100-ml-Schüttelkolben und brüten die Hefe für 90 min bei 30 ° C und 120 Umdrehungen pro Minute.
- Pellet die Zellen für 5 Minuten bei 700 · g und den Überstand verwerfen. Das Pellet in 4,5 ml steriler 0,9% (w / v) NaCl und gut durch vorsichtiges Auf- und Abpipettieren mit einer 10-ml serologische Pipette mischen. Ziehen Sie 50 & mgr; l und bereiten das Zehnfache Verdünnungen 1 in 0,9% NaCl von 1:10 bis: 1000. Platte 100 & mgr; l jeder Verdünnung auf 90-mm-Petrischalen mit SD-trp-leu-Agar.
- Verbreiten Sie den Rest des unverdünnten Hefe Aufwirbelung auf 16- x 150 mm Durchmesser Petrischalen mit SD-trp-leu-his-ade Agar. In 3-AT an das Medium Selbstaktivierung zu reduzieren. Inkubieren Sie die SD-trp-leuPlatten für drei Tage, und die SD-trp-leu-his-Platten ADE für vier Tage bei 30 ° C.
HINWEIS: Die Klone, die auf den selektiven Platten erscheinen, sind (potentiellen) zusammenwirkenden Paare und die für den Köder in Wechselwirkung Partner pGAD-HA kodierenden Plasmid tragen. - Bestimmen Sie die Transfektionseffizienz durch die Kolonien der verschiedenen Reihenverdünnungen auf die SD-trp-leu-Selektionsplatten zu zählen. Übertragen Sie jeden Klon durch Kommissionierung und Ausstreichen der Kolonie mit einer sterilen Pipettenspitze auf frischem SD-trp-leu-his-ade-selektiven Platten. Inkubieren der Platten für 24 h bei 30 ° C. Wiederholen Sie jeden Tag diesen Schritt, bis insgesamt fünf Passagen erreicht ist.
8. Analyse der Klone aus der selektiven Platten
- Bereiten einer sterilen 2 ml-Reaktionsröhrchen mit 1 ml SD-trp-leu-his-ade für jeden Klon unter einer sterilen Haube. Lochen Sie ein Loch in jedes Röhrchen mit einer heißen Nadel und decken Sie das Loch mit einem Stück gasdurchlässige Versiegelung. Impfen jeder Flasche mit frischen Kolonie paaren rial von einem Klon (Schritt 7.17; verwenden Klone nach dem 5. Durchgang) und Inkubation für 24 h bei 30 ° C, bei 150 Umdrehungen pro Minute schütteln.
Hinweis: Es ist wichtig, Klone aus einer frischen Platte zu nehmen, weil Hefe von Platten für mehrere Tage bei 4 ° C ausreichend innerhalb von 24 h in flüssigem Medium wachsen nicht gespeichert genommen. - Pellet die Hefe für 5 min bei 4000 × g und den Überstand verwerfen. Das Pellet in dem entsprechenden Resuspensionspuffers (aus dem jeweiligen Minipräparationskit Plasmid-DNA) und übertragen sie in ein frisches Reaktionsgefäß 2,0 ml. 100 l Säure gewaschenen Glasperlen (425- bis 600 & mgr; m Durchmesser) und 5 min kräftig mischen.
- Fügen Sie die entsprechenden Puffer Lyse und fahren Sie mit dem Plasmidaufreinigung eine Plasmidpräparation Mini-Kit den Anweisungen des Herstellers folgen. Eine Elution der Plasmid-DNA mit 50 & mgr; l Wasser.
- Verwenden Sie die DNA aus Schritt 8.3 für eine Sequenzierungsreaktion mit der Beute Plasmid-spezifischen Primer, GAL4ADseqref "> 41 5 'ACCACTACAATGGATGATG -3'.
- Überprüfen Sie die Interaktion von de novo Cotransformieren 43, 44 der Köder und die Beute Vektor. Wählen Sie die transformierte Hefe auf SD-trp-leu-his-ade-Selektionsplatten.
Ergebnisse
Vor dem eigentlichen kann Y2H Bildschirm der Köder durchgeführt werden müssen für Selbstaktivierung getestet werden. Dies wird durch die Umwandlung der Köder Expressionsvektor zusammen mit dem leeren Beute Bibliothek Vektor und Überprüfung Wachstum auf selektiven Platten erreicht.
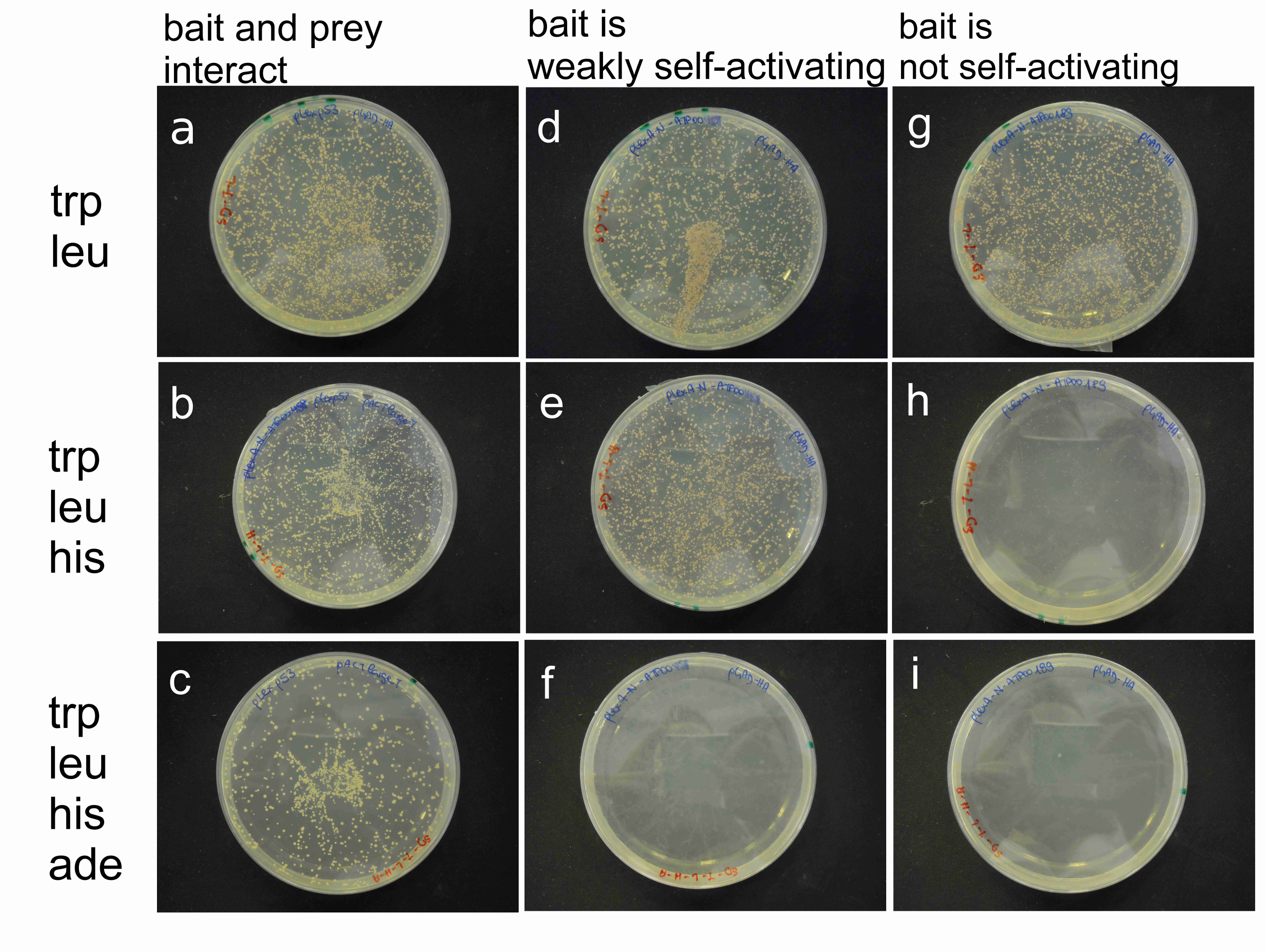
Um zu analysieren , ob die phytoplasmal Protein ATP_00189 selbstaktivierende ist, wurde die Selbstaktivierung Test in Abschnitt 5. Der Köderplasmid ergänzt den trp und die Beute Plasmid , das das leu Auxotrophie von S. cerevisiae NMY51 40 wie beschrieben durchgeführt. Eine erfolgreiche Co-Transformation wird also durch Wachstum auf selektiven Platten fehlen trp und leu gekennzeichnet. Die Wechselwirkung des Köder und Beute-Protein führt zu einer Komplementierung der seine und ade Auxotrophie von NMY51. Wenn Selbstaktivierung durch den Köder in der Abwesenheit einer Wechselwirkung auftritt, wächst die Hefe auf selektiven Platten seiner und ade fehlt. Starke und schwache SelbstAktivierung auftreten. Starke Selbstaktivierung des Köders wird durch Wachstum des co-transformierte Hefe auf trp-leu-his-ade abgereicherten Selektionsplatten gekennzeichnet. Schwache Selbstaktivierung führt zu Wachstum auf trp-leu-his aber nicht auf trp-leu-his-ade abgereicherten Selektionsplatten. Die richtige positive Kontrollen sind unverzichtbar für die Ergebnisse der Selbstaktivierungstest zu interpretieren. Eine Zusammenfassung der erwarteten Ergebnisse der Selbstaktivierungstest und deren Interpretation ist in Tabelle 1 und visualisiert in Abbildung 1 zur Verfügung gestellt.

Abbildung 1: Beispiel von Bait Selbstaktivierung Tests eines Bait Bevor ein Y2H Bildschirm durchführen. S. cerevisiae NMY51 wurde co-transformiert mit zusammenwirkenden Köder (pLEX-p53) und Beute (PACT larget) als eine positive Kontrolle (linkes Bild: ac), einem schwach selbstaktivierende plus eine leere Beute library - Vektor (Mitteltafel: df) und eine nicht selbstaktivierende Köder plus eine leere Beute Bibliothek Vektor (rechtes Bild: gi). Die Co-transformierten Hefe wurden auf SD - Platten fehlen trp und leu (oberes Bild: a, d, g), trp, leu und sein (Mitteltafel: b, e, h) und trp, leu, sein und ade (unterer Panel: c, f, i). Die Auswahl auf Medium ohne trp und leu ist eine positive Kontrolle für die erfolgreiche Co-Transformation, wie der Köder Vektor, der das trp Auxotrophie und die Beute Vektor der leu Auxotrophie von NMY51 (Wachstum auf SD-trp-leu) ergänzt. Im Falle einer Wechselwirkung zwischen Köder und Beute oder eine Selbstaktivierung des Köders wird der Reporterexpression von NMY51 eingeschaltet und ergänzt die His- und ade Auxotrophie (Wachstum auf SD-trp-leu-his-ADE). Ein schwaches Selbstaktivierung wird durch Wachstum auf SD-trp-leu-his - Platten (Mitteltafel, df) gekennzeichnet. Schwache Selbstaktivierung eines Köders müssen für die Analyse der Köder vor vermindert werdenin einem Y2H Bildschirm, zum Beispiel durch Zugabe von 3-AT zur selektiven Medien. Aminosäure Depletionen sind angegeben mit "-" in den jeweiligen Mediennamen. Bitte klicken Sie hier , um eine größere Version dieser Figur zu sehen.
{kind=link}
| pLexA-N-atp00189 | keiner | Kolonien | kein Wachstum | kein Wachstum | kein Wachstum | kein Wachstum |
| keiner | pGAD-HA | kein Wachstum | Kolonien | kein Wachstum | kein Wachstum | kein Wachstum |
| pLexA-N | keiner | Kolonien | kein Wachstum | kein Wachstum | kein Wachstum | kein Wachstum |
| pLexA-N | pGAD-HA | Kolonien | Kolonien | Kolonien | kein Wachstum | kein Wachstum |
| pLexA-p53 | PACT larget | Kolonien | Kolonien | Kolonien | Kolonien | Kolonien |
| pLexA-N-atp00189 | pGAD-HA | Kolonien | Kolonien | Kolonien | kein Wachstum | kein Wachstum |
Tabelle 1: Erwartete Ergebnisse in einem Bait Selbstaktivierungstest. Die Transformation von S. cerevisiae NMY51 ergibt in Differential Wachstum auf Selektivmedien basierend auf den Eigenschaften von cotransformiert Köder und Beute. Wachstum auf selektiven Platten wurde bei 30 ° C nach der Transformation von verschiedenen Vektor-Kombinationen (af) nach 72 h Inkubation ausgewertet. Schwache Selbstaktivierung wird durch das Wachstum der Hefe auf SD-trp-leu-his und starke Selbstaktivierung durch Wachstum auf SD-trp-leu-his-ade Auswahl plat gekennzeichnetes in Abwesenheit eines Interaktionspartner. Die pLexA-N kodierten phytoplasmal Effektor ATP_00189 zeigt keine Selbstaktivierung, die durch die Unfähigkeit des Bait-Vektor gekennzeichnet ist NMY51 transformiert in Abwesenheit seines und ADE zu wachsen.
Je nach dem Köder kann ein Y2H Bildschirm zahlreiche Hefe-Klone wachsen auf selektiven Platten gewinnen. Alle Klone müssen für mögliche Redundanzen analysiert und geprüft werden. Selbst wenn eine normalisierte cDNA-Bibliothek verwendet worden ist, ist es sehr wahrscheinlich, dass ein Interaktors in vielen verschiedenen Klone repräsentiert wird. In Abhängigkeit von der Bibliothek Klonierungstechnik ist es auch möglich, daß nur Fragmente des vollständigen Gens in einigen Beutevektoren eingefügt werden. Es ist daher ratsam, die de novo verstärken (von cDNA) und subklonieren das Gen voller Länge des Interaktors und die Interaktion zu testen in einer Eins-zu-Eins - Y2H Analyse (Abbildung 2).

Abbildung 2: Beispiel einer Wechselwirkung zwischen Köder und Beute in einem Y2H Experiment. Ein Hefe-Zwei-Hybrid (Y2H) Bildschirm wurde durchgeführt, und Plasmide, die aus positiven Interaktor Klone wurden gereinigt. Die Beute Vektor wurde sequenziert und die Host - Interaktionspartner Malus x domestica MdTCP24 und MdTCP25 wurden identifiziert. Als Negativkontrolle MdTCP34 (für die keine Interaktor in der Y2H Bibliothek Bildschirm identifiziert) wurde parallel subkloniert. Die volle Länge Gene wurden aus Apfel cDNA amplifiziert, subkloniert in dem Beute - Vektor (co-cistronically der Aktivierungsdomäne AD exprimierenden) und de novo co-transformiert mit dem Bait - Vektor des bakteriellen Effektor ATP_00189 an die DNA - Bindungsdomäne (BD) gekoppelt ausdrückt. Diese Zahl wird von 28 genommen. Bitte klicken Sie hier ein , um zu vergrößernVersion dieser Figur.
{kind=link}
Diskussion
Im Y2H Bildschirm wird das Potential Effektor-Protein mit verschiedenen hypothetischen interagierende Proteine (Schritt 7) co-exprimiert wird. Jeder wachsende Hefe Klon enthält den Köder, aber eine (potentiell) unterschiedliche Interaktor. Die interagierenden Proteine werden in einer cDNA-Bibliothek codiert wird, die in die Beute Plasmide kloniert wird. Der Köder und die Beute Plasmide jedes Co-cistronically-Code für einen Teil eines Hefe-Transkriptionsfaktors. Im Falle einer physikalischen Wechselwirkung zwischen Köder (Effektorzellen) und einer Beute plasmid-kodierten Interaktor die beiden Transkriptionsfaktor Teile (die DNA-Bindungsdomäne und die Aktivierungsdomäne) vereinigt sind, und die Expression des Reportergens induziert wird. Die Hefe in der Lage, auf Histidin und Adenin-verarmten SD-Selektionsplatten zu wachsen. Die Art der Beute cDNA-Bibliothek ist abhängig von der abgeschirmten Effektor und muss entsprechend gewählt werden. Die Beute und Köder Vektor, sowie der Hefestamm, müssen kompatibel sein. In diesem Bildschirm Bindung eine LexA DNA-Domäne und die Gal4 Aktivierung domain wurden als kompatibel Hefe-Transkriptionsfaktor-Einheiten eingesetzt. Die cDNA-Bibliothek kann individuell konstruiert werden, nach Maß, oder im Handel erhalten. Die Herstellung der cDNA Beute-Bibliothek ist nicht Teil dieses Protokolls. In diesem Protokoll wird ein selbst erstellter, normalisierte cDNA - Bibliothek aus der RNA von Malus x domestica Blätter verwendet wurde und kloniert in pGAD-HA 41. Der Effektor (Köder) Hefestamm exprimierenden wurde mit der Beute-Bibliothek transformiert, und Klone wurden auf Histidin und Adenin-verarmten SD-Selektionsplatten ausgewählt. Plasmid-DNA aus den Hefekolonien, ein säulenbasierte DNA Plasmid Mini Kit für Bakterien Extrakt in Kombination mit einer mechanischen Störung Schritt unter Verwendung von Glasperlen empfohlen, Hefe Lyse (Schritt 8) verbessern. In diesem Plasmid Reinigung wird der Köder und die Beute Vektor gereinigt in relativ geringen Konzentrationen gleichzeitig werden. Jedoch ist die Menge an Plasmid DNA genug, um das Wechselwirkungspartner durch Sequenzierung wit zu identifizierenhout vor Plasmid Ausbreitung (Schritt 8.4). Als Alternative gereinigte DNA in Schritt 8.4 kann in kompetente E. coli und ausgewählt mit Beute - Vektor-vermittelter Antibiotikaresistenz transformiert werden (zB das Ampicillin bei pGAD-HA). Die ausgewählte E. coli - Kolonien tragen nur die pGAD-HA Bibliothek Plasmid und High-Yield - Plasmid Reinigung kann mit diesen Klonen durchgeführt werden.
Auch die erfolgreiche Transformation von pLexA-N und pGAD-HA-Konstrukte ergänzt das Tryptophan und Leucin-Auxotrophie von NMY51. Eine Wechselwirkung zwischen dem Effektor und einem Protein (kodiert auf pGAD-HA) führt zur Aktivierung des Reportersystems in NMY51 Stämmen. Im Fall der Selbstaktivierung, NMY51 mit dem Effektor transformierte pLexA-N in Kombination mit dem leeren Bibliotheksvektor pGAD-HA wächst auf selektiven Platten fehlt trp-leu-his-ade aufgrund der unerwünschten Aktivierung des NMY51 Reportersystems exprimiert 40. Die Selbstaktivierung kann w seineak und in Abwesenheit von trp-leu-his oder die Aktivierung durch Wachstum stark sein kann und dadurch auf trp-leu-his-ade Platten 41 auftreten können. Selbstaktivierung kann einen massiven Hintergrund von falsch-positiven Klonen im eigentlichen Y2H Bildschirm verursachen. Um dies zu vermeiden, muss eine Selbstaktivierungstest mit dem Köder werden durchgeführt, in dem der Effektor-exprimierenden Vektor co-transformiert mit dem leeren Vektor-Bibliothek. In dieser experimentellen Einstellung, Reportergenexpression muss nicht induziert werden. Wenn Selbstaktivierung ( das heißt Wachstum auf selektiven Platten) in dem Test sichtbar ist, kann das Medium mit unterschiedlichen Konzentrationen von 3-Amino-1,2,4-triazol 45 (3-AT) ergänzt werden. 3-AT ist ein Inhibitor der imidazoleglycerol-phosphat - Dehydratase (HIS3), ein Enzym wichtig während Histidinbiosynthese 46. Die Supplementation mit 3-AT können die schwachen Effekte der Selbstaktivierung während Y2H Bildschirme 47, reduzieren48. Verschiedene Konzentrationen von 3-AT sollte getestet werden. In diesem Protokoll, 1-40 mM 3-AT wurden verwendet. Die niedrigste Konzentration, die Selbstaktivierung zur Unterdrückung führt sollten anschließend im Y2H Bildschirm verwendet werden. Die selektiven Platten des Selbstaktivierungstest kann nur dann, wenn die SD-trp-leu-Platten der eine ausreichende Anzahl von Klonen enthalten Cotransformation ausgewertet werden. Als Hinweis ≥ 500 Kolonien pro 90 mm (Durchmesser) Petrischale ausreichend sind. Um die genaue Transfektionseffizienz zu bestimmen, wird empfohlen, serielle Verdünnungen der co-transformierter Hefe herzustellen und sie auf SD-trp-leu-Platten zu verteilen.
Selbstaktivierung zu reduzieren, kann der Effektor in pLexA-C geklont werden, ein Köder-Expressionsvektor, der das LexA-Tag an den C-Terminus des Proteins verbindet. Die Orientierung der Lex-A - Tag kann Selbstaktivierung 24 dämpfen. Jedoch ist es nicht möglich, in jedem Falle zu verringern oder zu Selbstaktivierung abzuschaffen. Die Formation derrot oder rötlich Kolonien deutet auf eine schwache oder falsch-positive Interaktion. Der ade2 Reportergen von NMY51 wird nur aktiviert , wenn es um eine Interaktion von Protein-Protein kommt, was wiederum blockiert die Akkumulation eines roten Farbstoffes in diesem Hefestamm 40. In Abwesenheit einer Wechselwirkung sind NMY51 rötlich, während die Kolonien tragenden starke Interaktoren weiß sind. Die Beobachtung der Koloniefarbe auf den Selektionsplatten ist somit ein weiterer wichtiger Hinweis, um zu beurteilen, ob die jeweiligen Kolonien Echt- oder falsch-positiven Interaktoren.
Es ist schließlich notwendig, anzupassen oder bestimmte Testeinstellungen ändern in Bezug auf die Art der Köder und die Interaktion Eigenschaften. Mittlerweile wurden eine Reihe von Verbesserungen und derivative Techniken des gemeinsamen Y2H etabliert für die Analyse von sehr schwierig Protein-Interaktionen in verschiedenen Host-Systemen zu ermöglichen. Eine Überprüfung von Stynen et al. 49 Adressen and beschreibt verschiedene Aspekte der Y2H, seine Verbesserungen und Anpassungen und liefert somit wertvolle Informationen darüber, wie die entsprechende Interaktion Test zu wählen.
Y2H Bildschirme und abgeleitete Techniken weit verbreitet in verschiedenen Forschungsbereichen eingesetzt werden, wo auch immer Identifizierung von binären Protein-Interaktionen erforderlich ist. Selbst wenn kritische Kontrollen durchgeführt werden, wie beispielsweise Selbstaktivierungstests des Köders und ein-zu-eins - Wieder Transformationen der identifizierten interagierende Proteine, die Y2H anfällig falsche Ergebnisse liefern 26, 50, 51. Die Hefe als Modell für alle Interaktionsstudien nicht geeignet. Hefe nicht notwendigerweise eine zelluläre Umgebung dar, die die entsprechende posttranslationale Modifikation und Faltung für jedes 24 - Protein unterstützt. Weiterhin ist in der Y2H Einstellung werden die Proteine überexprimiert, und ihre Expression ist nicht die Kontrolledurch ihren natürlichen Promotor führte. Die Y2H zwingt die proteinösen Bindungspartner an den Zellkern, die nicht notwendigerweise ihre natürliche subzellulären Ziel. Die nativen zellulären Umstände der Interaktion kann somit nicht von der Hefe reflektiert werden und zu falsch negativen oder falsch-positiven Ergebnissen führen kann. Die meisten Y2H werden auf Hefe-Auxotrophie Komplementation als selektives Prinzip. Diese Ernährungs Auswahl wird durch eine hohe Empfindlichkeit aus, aber auf Kosten einer verringerten Selektivität im Vergleich zu anderen (beispielsweise chromogene reporter) -Assays 49. Wenn unreducible Selbstaktivierung (siehe oben) oder andere Beschränkungen auftreten , für eine bestimmte Effektorfunktion ist die Y2H kein geeigneter Test 25. In Tabelle 1 und Abbildung 1, die erwarteten Ergebnisse des Selbstaktivierungstest gegeben. Das Fehlen von realen Interaktionen (dh falsche Negative) durch Protein Toxizität, fehlerhafte translationale Protein verursacht werdenVerarbeitung, sterische Hinderung der Interaktion, unterrepräsentierte Interaktionspartner in der Beute Bibliothek, Membranlokalisierung des Köders oder fehlenden Komponenten , die für die Interaktion 24.
Es wird empfohlen, de novo verstärken das Volllängen - Gen des identifizierten interagierenden Proteins aus cDNA, subklonieren es in pGAD-HA, und führen eine Eins-zu-Eins - Wechselwirkungsassay die erzeugte pGAD-HA - Konstrukt in den Köder-exprimierenden durch Transformieren NMY51 Stamm. Dies ist notwendig, da die beobachtete Interaktors in dem Beute-Bibliothek nur ein Fragment eines größeren Proteins sein könnte. Jedoch ist die Information für die jeweiligen Volllängen-Gen nur zugänglich, wenn beträchtliche genomischen und transkriptomischen Sequenzdaten verfügbar sind. Das Transformationsprotokoll für das Selbstaktivierungstest hier beschrieben sind, können für eine solche Eins-zu-Eins-Assays angewendet werden, und die Wechselwirkung zwischen dem Bait und dem Interaktor reproduzierbar sein müssen.
Wechselwirkungen in einem Y2H Bildschirm identifiziert muss immer von einer anderen unabhängigen Technik bestätigt werden. Diese unabhängige Technik muss auf die natürliche Umgebung des eigentlichen Protein-Protein-Wechselwirkung näher sein. In dem Fall des phytoplasmal Effektor ATP_00189 wurde die Wechselwirkung mit den TCP - Transkriptionsfaktoren von Malus x domestica in planta mit bimolekularen fluoreszierendes Komplementation (BiFC) in Nicotiana benthamiana prüft Protoplasten 28 (nicht Teil des Protokolls). Der Effektor-Protein ATP_00189 wurde aus der Pflanze Erreger P. mali abgeleitet. In planta BIFC wurde somit ATP_00189 Wechselwirkungen mit den Malus x domestica Transkriptionsfaktoren zu überprüfen gewählt zuvor in dem Y2H 28 identifiziert. BiFC ist ein Protein-Protein-Interaktionsassay, der nicht die subzelluläre Translokation der interagierenden Proteine in den Zellkern, um das Reportersystem zur Aktivierung erfordert 52. Darüber hinaus ahmt die in planta - Expression und Modifikation Maschinen die Umgebung der natürlichen Interaktion zwischen Pflanze bakterielle Effektor in seiner Wirtspflanze. Allerdings ist ein globaler Bildschirm BIFC mit nicht durchführbar ist.
Es gibt mehrere Schritte in dem Protokoll, das sorgfältig behandelt werden müssen. Wenn das Gen von Interesse zu verstärken (Schritt 4), ist es wichtig zu bestimmen, dass Primer an Pflanzen-DNA binden. Somit DNA aus nicht-infizierten Pflanzen müssen als negative Kontrolle einbezogen. Die Sequenz des Gens von Interesse nicht enthalten die EcoRI und SalI-Restriktionsstellen, die für die gerichtete Klonierung des Einsatzes verwendet werden. Wenn die Sequenz diese Seiten enthält, müssen verschiedene Restriktionsenzyme für das Klonen ausgewählt werden. Hefe-Transformation ist eine zentrale Methode während dieses Protokolls. Die Transfektionseffizienz ist abhängig von der Kompetenz der Hefezellen, ihre Lebensfähigkeit, den Wachstumszustand, und die Qualität der Transfektion REAGent 53, 54. Für das vorgeschlagene Protokoll wird dringend empfohlen, Hefe zu verwenden, die aus frischen Platten nicht älter als zwei Wochen (gehalten bei 4 ° C), wenn die Transformationen durchführen. Oft wird das Wachstum der transformierten Hefe verzögert, wenn selektive Flüssigkulturen mit Kolonien aus alten Agarplatten inokuliert. Es ist auch hilfreich, um eine flüssige Starterkultur als Inokulum für den tatsächlichen Experimente zu verwenden und nicht die Hefe von der Platte direkt zu verwenden. Geringe Effizienz bei der Transfektion der Beute in den Köder-exprimierenden Hefe kann auf die Unterrepräsentation bestimmter Beuteproteine (potentielle Interaktionspartner) führen und den gesamten Bildschirm so schräg kann. In diesem Protokoll wird ein Hefe-Transfektionseffizienz von ~ 150.000 cfu / & mgr; g DNA transfiziert in der Y2H funktionierte gut.
die Mängel und Nachteile des Y2H Technik zu kennen und die Ergebnisse in einer kritischen und in geeigneter Weise zu interpretieren ist unverzichtbar, um die korr Zeichnungect Schlussfolgerungen. Y2H Assays und die Derivate werden seit vielen Jahren verwendet und haben viele Verbesserungen und Anpassungen in Bezug auf die verschiedenen Ködercharakteristik, die subzelluläre Lokalisierung der Wechselwirkung, die erwarteten Bindungspartner und andere Faktoren unterzogen, die für die Interaktion erforderlich sein könnten (siehe Y3H 55, 56, 57). Vor kurzem Array-basierten Y2H Bildschirme entwickelt worden, die für automatisierte Hochdurchsatz - Analyse zahlreicher Ködern gegen Millionen preys 58 ermöglichen. Die Zukunft liegt höchstwahrscheinlich in der Automatisierung und Hochdurchsatz-Ansätze zu diesem Test für die Aufklärung komplexer Signalwege und Interaktom in verschiedenen Forschungsbereichen zu ermöglichen.
Offenlegungen
Die Autoren erklären, dass sie keine finanziellen Interessen haben.
Danksagungen
Wir danken Christine Kerschbamer, Thomas Letschka, Sabine Oettl, Margot Raffeiner und Florian Senoner vom Versuchszentrum Laimburg und Mirelle Borges Dias Schnetzer von Dualsystems Biotech AG für die technische Unterstützung und Julia Strobl für das Manuskript Korrektur. Diese Arbeit wurde im Rahmen des APPL2.0 Projekt durchgeführt und wurde von der Autonomen Provinz Bozen / Bolzano, Italien und dem Südtiroler Apfelkonsortium teilweise finanziert.
Materialien
| Name | Company | Catalog Number | Comments |
| Cetyltrimethylammonium bromide (CTAB) | Applichem | A6284 | for DNA preparation |
| Trishydroxymethylaminomethane (Tris) | Applichem | A2264 | for media and buffer |
| Sodium chloride (NaCl) | Applichem | A2942 | for media and buffer |
| Ethylenediaminetetraacetic acid Disodium salt (NaEDTA) | Applichem | A2937 | for media and buffer |
| N-Lauroylsarcosin sodium salt | Applichem | A7402 | for DNA preparation |
| ammonium acetate | Sigma-Aldrich (Fluka) | 9688 | for DNA preparation |
| 2-mercaptoethanol | Applichem | A1108 | for DNA preparation |
| Isopropanol (2-Propanol) | Applichem | A3928 | for DNA preparation |
| Ethanol | Sigma-Aldrich (Fluka) | 51976 | for DNA preparation |
| RNAse | Applichem | A2760 | for DNA preparation |
| Chloroform:Isoamyl 24:1 | Applichem | A1935 | for DNA preparation |
| Proofreading Polymerase (e.g. iProof) | Bio-Rad | 203433 | for PCR |
| EcoRI | Thermo Scientific | ER0271 | for cloning |
| SalI | Thermo Scientific | ER0641 | for cloning |
| Column-based DNA Purification Kit (e.g. QIAquick) | Qiagen | 28104 | for cloning |
| Kanamycin Sulfate | Applichem | A1493 | for microbiological selection |
| 3-Amino-1,2,4-Triazole (3-AT) | Sigma-Aldrich | A8056 | for self-activation assay |
| L-Arginine Monohydrochloride | Applichem | A3680 | for yeast culture |
| L-Isoleucine | Applichem | A3642 | for yeast culture |
| L-lysine Monohydrate | Applichem | A3448 | for yeast culture |
| L-Methionine | Applichem | A3897 | for yeast culture |
| L-Phenylalanine | Applichem | A3464 | for yeast culture |
| L-Threonine | Applichem | A3946 | for yeast culture |
| L-Tyrosine | Applichem | A3401 | for yeast culture |
| L-Uracile | Applichem | A0667 | for yeast culture |
| L-Valine | Applichem | A3406 | for yeast culture |
| L-Adenine Hemisulfate Salt | Applichem | A1596 | for yeast culture |
| 0.22 µm Pore Filter | Sartorius | 16541 | for sterile filtration |
| L-Histidine Monohydrochloride | Applichem | A3719 | for yeast culture |
| L-Leucine | Applichem | A3496 | for yeast culture |
| L-Tryptophane | Applichem | A3410 | for yeast culture |
| Yeast Nitrogen Base | Sigma-Aldrich | 51483 | for yeast culture |
| D-Glucose Monohydrate | Applichem | A1349 | for yeast culture |
| Agar | Fisher Scientific | BP2641-1 | for yeast and bacteria culture |
| Peptone | Applichem | A2208 | for yeast culture |
| Polyethylene Glycol 4000 (PEG) | Applichem | A1249 | for yeast transformation |
| Lithium Acetate Dihydrate (LiOAc) | Applichem | A3478 | for yeast transformation |
| Salmon Sperm DNA | Applichem | A2159 | for yeast transformation |
| Antibody against Lex-A tag (e.g. Anti-LexA DNA binding region) | Millipore | 06-719 | for Western blot |
| Plasmid Miniprep kit (e.g. GenElute Plasmid Miniprep Kit) | Sigma-Aldrich | PLN350 | for plasmid purification from yeast and bacteria |
| Acid Washed Glass Beads | Sigma-Aldrich | G8772 | for plasmid purification from yeast |
| Ampicillin Sodium Salt | Applichem | A0839 | for microbiological selection |
| Yeast Extract | Applichem | A1552 | for yeast culture |
| Vacuum concentrator centrifuge (e.g. Concentrator 5301) | Eppendorf | Z368245 (Sigma) | for DNA preparation |
| Bait vector (e.g. pLexA-N) | Dualsystems | P01004 | for Y2H |
| Prey vector (e.g. pGAD-HA) | Dualsystems | P01004 | for Y2H |
| Control prey vector (e.g. pLexA-p53) | Dualsystems | P01004 | for Y2H |
| Control bait vector (e.g. pACT-largeT) | Dualsystems | P01004 | for Y2H |
| Electrocompetent E. coli (e.g. MegaX DH10B T1R Electrocomp Cells) | Invitrogen | C640003 | for cloning |
| Yeast reporter strain (NMY51:MATahis3Δ200 trp1-901 leu2-3,112 ade2 LYS2::(lexAop)4-HIS3 ura3::(lexAop)8-lacZ ade2::(lexAop)8-ADE2 GAL4, e.g. NMY51) | Dualsystems | P01004 | for Y2H |
| Plastic paraffin film (e.g. Parafilm M) | Sigma-Aldrich | P7793-1EA | for yeast and bacteria culture |
| Gas permeable sealer (e.g. BREATHseal) | Greiner | 676 051 | for yeast culture |
| PCR Cycler (e.g. T100 Thermal Cycler) | Bio-Rad | 1861096 | for PCR |
| quantitative PCR Cycler (e.g. CFX96 Touch Real-Time PCR Detection System) | Bio-Rad | 1855195 | for quantitative PCR |
| Microcentrifuge (e.g. Centrifuge 5417 R) | Eppendorf | 5417 R | general lab equipment |
| Centrifuge (e.g. Centrifuge 5804 R) | Eppendorf | 5804 R | general lab equipment |
| Nuclease-free water (DNAse-free, RNAse-free, Protease-free) | 5Prime | 2500010 | for PCR and DNA works |
| Scalpell | Swann-Morton | 0301 | for DNA preparation |
| Sterile filter (0.22 µm; Polyethersulfone) | VWR-International | 514-0073 (European Catalogue number) | for yeast and bacteria culture |
| Petri dishes Ø 90 mm | Greiner Bio-One | 633180 | for yeast and bacteria culture |
| Petri dishes Ø 150 mm | Greiner Bio-One | 639161 | for yeast culture |
| Photometer (e.g. BioPhotometer) | Eppendorf | 550507804 | for yeast culture |
| Photometer Cuvettes | Brand | 759015 | for yeast culture |
| Water Bath (e.g. Water Bath Memmert WB7) | Memmert | WB7 | for yeast transformation |
| Western Blot Equipment | Bio-Rad | diverse | for protein detection |
| dNTPs | 5Prime | 2201210 | for cloning |
| Shaker (Erlenmeyer) Flasks (100 mL; 1,000 mL; 2,000 mL) and breathable cotton plugs (e.g. Duran Erlenmeyer narrow-neck flasks) | Sigma Aldrich | Z232793, Z232858, Z232866 | for yeast and bacteria culture |
| Shaking Incubator (e.g. GFL 3031) | GFL | 3031 | for yeast and bacteria culture |
| Incubator | Binder | KB 53 (E3.1) | for yeast and bacteria culture |
| Ice Maker (e.g. Ice Maker IF 825) | Omniwash | IF 825 | general lab equipment |
| 0.2 mL, 1.5 mL and 2.0 mL Reaction Vials (Polypropylene) | Eppendorf | 0030.124.332 (0.2 ml); 0030.123.328 (1.5 ml); 0030.123.344 (2.0 ml) | general lab equipment |
| Pipettes and disposable pipette Tips (different volumina: 10 - 1,000 µL) | Eppendorf | diverse | general lab equipment |
| Spreader | Sigma-Aldrich | SPR-L-S01 | for yeast and bacteria culture |
| Vortex shaker (e.g. MS 3 Minishaker) | IKA | 3617000 | general lab equipment |
| T4-Ligase | Thermo Fisher | EL0011 | for cloning |
| Fridge (4 °C) (e.g. Liebherr Medline FKEX 5000) | Liebherr | 81.767.580.4 | general lab equipment |
| Freezer (-20 °C) (e.g. Liebherr Comfort Professional G5216) | Liebherr | G5216 | general lab equipment |
| Graduated Cylinder (e.g. Brand) | Sigma Aldrich | Z327352; Z327417; Z327441 | general lab equipment |
| Serological Pipettes (e.g. Fisherbrand) | Thermo Fisher | S55701 | for yeast and bacteria culture |
| Mechanical Pipettor (e.g. Pipetboy acu 2) | Integra-Biosciences | 155000 | general lab equipment |
| Cuvettes for Electroporation (1 mm) | Molecular Bio Products | 5510-11 | for bacteria transformation |
| Electroporator (e.g. Eporator) | Eppendorf | 4309000019 | for bacteria transformation |
| Gel and Blot imaging system (e.g. ChemiDoc MP System) | Bio-Rad | 1708280 | general lab equipment |
| Conical centrifuge tubes (Polypropylene) (50 mL) | VWR-International | 525-0403 (European Catalogue number) | general lab equipment |
| Conical centrifuge tubes (Polypropylene) (13 mL) | BD Falcon | 352096 | general lab equipment |
| Dithiothreitol (DTT; 1 M) | Thermo Scientific | P2325 | for ligation |
| adenosine triphosphate (ATP, e.g. ATP Solution (10 mM)) | Ambion | AM8110G (distributed by Thermo Scientific) | for ligation |
| Dropout mix without histidine, leucine, tryptophan, and adenine (DO, e.g. Yeast Synthetic Drop-out Medium Supplements) | Sigma-Aldrich | Y2021 Sigma | for yeast culture (ready-made mix) |
Referenzen
- Fields, S., Song, O. -K. A novel genetic system to detect protein-protein interactions. Nature. 340 (6230), 245-246 (1989).
- Dombrosky-Ferlan, P. -M., Corey, S. -J. Yeast two-hybrid in vivo association of the Src kinase Lyn with the proto-oncogene product Cbl but not with the p85 subunit of PI 3-kinase. Oncogene. 14, 2019-2024 (1997).
- Singh, R. Rice Mitogen-Activated Protein Kinase Interactome Analysis Using the Yeast Two-Hybrid System. Plant Physiol. 160, 477-487 (2012).
- Lee, H. -J., et al. Interaction of NADPH oxidase 1 with Toll-like receptor 2 induces migration of smooth muscle cells. Cardiovasc. Res. 99, 483-493 (2013).
- Del Bene, F., Tessmar-Raible, K., Wittbrodt, J. Direct interaction of geminin and Six3 in eye development. Nature. 427 (6976), 745-749 (2004).
- Kumar, A., Godwin, J. -W., Gates, P. -B., Garza-Garcia, A. -A., Brockes, J. -P. Molecular Basis for the Nerve Dependence of Limb Regeneration in an Adult Vertebrate. Science. 318 (5851), 772-777 (2007).
- Park, S. -Y., et al. Abscisic Acid Inhibits Type 2C Protein Phosphatases via the PYR/PYL Family of START Proteins. Science. 324 (5930), 1068-1071 (2009).
- Selyunin, A. S., et al. The assembly of a GTPase-kinase signalling complex by a bacterial catalytic scaffold. Nature. 469 (7328), 107-111 (2011).
- Uetz, P., et al. Herpesviral Protein Networks and Their Interaction with the Human Proteome. Science. 311 (5758), 239-242 (2006).
- Ushioda, R., Hoseki, J., Araki, K., Jansen, G., Thomas, D. Y., Nagata, K. ERdj5 Is Required as a Disulfide Reductase for Degradation of Misfolded Proteins in the ER. Science. 321 (5888), 569-572 (2008).
- Young, K. -H. Yeast two-hybrid: so many interactions, (in) so little time. Biol. Reprod. 58, 302-311 (1998).
- Krachler, A. M., Woolery, A. R., Orth, K. Manipulation of kinase signaling by bacterial pathogens. J. Cell Biol. 195 (7), 1083-1092 (2011).
- Hewezi, T., Baum, T. J. Manipulation of Plant Cells by Cyst and Root-Knot Nematode Effectors. Mol. Plant Microbe Interact. 26 (1), 9-16 (2012).
- McGhie, E. J., Brawn, L. C., Hume, P. J., Humphreys, D., Koronakis, V. Salmonella takes control: effector-driven manipulation of the host. Curr. Opin. Microbiol. 12 (1), 117-124 (2009).
- Kay, S., Bonas, U. How Xanthomonas type III effectors manipulate the host plant. Curr. Opin. Microbiol. 12 (1), 37-43 (2009).
- Navarro-Garcia, F., Serapio-Palacios, A., Ugalde-Silva, P., Tapia-Pastrana, G., Chavez-Dueñas, L. Actin Cytoskeleton Manipulation by Effector Proteins Secreted by Diarrheagenic Escherichia coli Pathotypes. Biomed. Res. Int. 2013, 22(2013).
- Bhat, B. S., Shanaz, E. Effectors-Role in Host-Pathogen Interaction. J. Agr. Env. Sci. 3 (2), 265-285 (2014).
- Bhavsar, A. P., Guttman, J. A., Finlay, B. B. Manipulation of host-cell pathways by bacterial pathogens. Nature. 449 (7164), 827-834 (2007).
- Natale, P., Brüser, T., Driessen, A. -J. -M. Sec- and Tat-mediated protein secretion across the bacterial cytoplasmic membrane-Distinct translocases and mechanisms. Biochimica et Biophysica Acta-Biomembranes. 1778 (9), 1735-1756 (2008).
- Costa, T. R. D., et al. Secretion systems in Gram-negative bacteria: structural and mechanistic insights. Nat Rev Micro. 13 (6), 343-359 (2015).
- Tseng, T. -T., Tyler, B. M., Setubal, J. C. Protein secretion systems in bacterial-host associations, and their description in the Gene Ontology. BMC Microbiol. 9 (1), 1-9 (2009).
- MacLean, A. M., et al. Phytoplasma effector SAP54 hijacks plant reproduction by degrading MADS-box proteins and promotes insect colonization in a RAD23-dependent manner. PLoS Biol. 12 (4), e1001835(2014).
- Sugio, A., Kingdom, H. N., MacLean, A. M., Grieve, V. M., Hogenhout, S. A. Phytoplasma protein effector SAP11 enhances insect vector reproduction by manipulating plant development and defense hormone biosynthesis. Proc Natl Acad Sci. 108 (48), 1254-1263 (2011).
- van Criekinge, W., Beyaert, R. Yeast Two-Hybrid: State of the Art. Biol Proced Online. 2 (1), 1-38 (1999).
- Bruckner, A., Polge, C., Lentze, N., Auerbach, D., Schlattner, U. Yeast two-hybrid, a powerful tool for systems biology. Int. J. Mol. Sci. 10 (6), 2763-2788 (2009).
- Serebriiskii, I., Estojak, J., Berman, M., Golemis, E. A. Approaches to Detecting False Positives in Yeast Two-Hybrid Systems. Biotech. 28 (2), 328-336 (2000).
- Golemis, E. A., Serebriiskii, I., Law, S. F. The yeast two-hybrid system: criteria for detecting physiologically significant protein-protein interactions. Curr. Issues Mol. Biol. 1 (1-2), 31-45 (1999).
- Janik, K., Mithöfer, A., Raffeiner, M., Stellmach, H., Hause, B., Schlink, K. An effector of apple proliferation phytoplasma targets TCP transcription factors - a generalized virulence strategy of phytoplasma. Mol. Plant Pathol. , (2016).
- Strauss, E. Microbiology. Phytoplasma research begins to bloom. Science. 325 (5939), 388-390 (2009).
- Kartte, S., Seemüller, E. Variable response within the genus Malus to the apple proliferation disease. J. Plant Dis. Protect. 95 (1), 25-34 (1988).
- Bovey, R. Observations and experiments on apple proliferation disease. Phytopath. Mediterr. 2 (3), 111-114 (1963).
- Schlink, K., Reski, R. Preparing High-Quality DNA From Moss (Physcomitrella patens). Plant. Mol. Biol. Rep. 20, 423(2002).
- Applichem. N-Lauroylsarcosine sodium salt. , https://www.applichem.com/fileadmin/datenblaetter/A7402_en_GB.pdf (2015).
- Applichem. 2-Mercaptoethanol. , https://www.applichem.com/fileadmin/datenblaetter/A1108_en_GB.pdf (2015).
- Applichem. 2-Propanol. , https://www.applichem.com/fileadmin/datenblaetter/A3928_en_GB.pdf (2015).
- Applichem. Chloroform:Isoamyl alcohol (24:1). , https://www.applichem.com/fileadmin/datenblaetter/A3928_en_GB.pdf (2015).
- Mehle, N., Nikolić, P., Gruden, K., Ravnikar, M., Dermastia, M. Real-time PCR for specific detection of three phytoplasmas from the apple proliferation group. Methods Mol. Biol. 938, 269-281 (2013).
- Applichem. Kanamycin sulfate. , https://www.applichem.com/fileadmin/datenblaetter/A1493_en_GB.pdf (2015).
- Smith, L. M., et al. Fluorescence detection in automated DNA sequence analysis. Nature. 321 (6071), 674-679 (1986).
- Lentze, N., Auerbach, D. Membrane-Based Yeast Two-Hybrid System to Detect Protein Interactions. Current Protocols in Protein Science. , Wiley Interscience. 1917-1952 (2008).
- Dualsystems Biotech AG. DUALhubrid Kit - Manual P01004 B06. www.dualsystems.com. , 1-44 (2012).
- Kurien, B. -T., Scofield, R. -H. Protein Blotting and Detection. , Humana Press. (2009).
- Gietz, R. D., Schiestl, R. H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protocols. 2 (1), 31-34 (2007).
- Yeast Transformation and Cloning. , JoVE Science Education Database. Cambridge, MA (USA). (2016).
- Sigma-Aldrich. 3-Amino-1,2,4-triazole. , http://www.sigmaaldrich.com/MSDS/MSDS/DisplayMSDSPage.do?country=IT&language=EN-generic&productNumber=A8056&brand=SIGMA&PageToGoToURL=http%3A%2F%2Fwww.sigmaaldrich.com%2Fcatalog%2Fproduct%2Fsigma%2Fa8056%3Flang%3Dit (2016).
- Ames, B. -N. The Biosynthesis of Histidine: d-erythro-Imidazole-Glycerol Phosphate Dehydrase. J. Biol. Chem. 228, 131-143 (1957).
- Joung, J. -K., Ramm, E. -I., Pabo, C. -O. A bacterial two-hybrid selection system for studying protein-DNA and protein-protein interactions. Prot Natl Acad Sci. 97, 7382-7387 (2000).
- Brennan, M. -B., Struhl, K. Mechanisms of Increasing Expression of a Yeast Gene in Escherichia coli. J. Mol. Biol. 136, 333-338 (1980).
- Stynen, B., Tournu, H., Tavernier, J., van Dijck, P. Diversity in genetic in vivo methods for protein-protein interaction studies: from the yeast two-hybrid system to the mammalian split-luciferase system. Microbiol. Mol. Biol. Rev. 76 (2), 331-382 (2012).
- Hengen, P. -H. Methods and reagents: False positives from the yeast two-hybrid system. Trends Biochem Sci. 22 (1), 33-34 (1997).
- Vidalain, P. -O., Boxem, M., Ge, H., Li, S., Vidal, M. Increasing specificity in high-throughput yeast two-hybrid experiments. Methods. 32 (4), 363-370 (2004).
- Pattanaik, S., Werkman, J. -R., Yuan, L. Bimolecular Fluorescence Complementation as a Tool to Study Interactions of Regulatory Proteins in Plant Protoplasts. Plant Transcription Factors: Methods and Protocols. Yuan, L., Perry, E. S. , Humana Press. Totowa, NJ. 185-193 (2011).
- Ito, H., Fukuda, Y., Murata, K., Kimura, A. Transformation of Intact Yeast Cells Treated with Alkali Cations. J. Bacteriol. 153 (1), (1983).
- Kawai, S., Hashimoto, W., Murata, K. Transformation of Saccharomyces cerevisiae and other fungi. Bioeng Bugs. 1 (6), 395-403 (2010).
- Bernstein, D. -S., Buetr, N., Stumpf, C., Wickens, M. Analyzing mRNA-protein complexes using a yeast three-hybrid system. Methods. 26, 123-141 (2002).
- Stumpf, C. -R., Opperman, L., Wickens, M. Analysis of RNA-Protein Interactions Using a Yeast Three-Hybrid System. Methods Enzymol. 449, 295-315 (2008).
- Cottier, S., et al. The yeast three-hybrid system as an experimental platform to identify proteins interacting with small signaling molecules in plant cells: potential and limitations. Front. Plant Sci. 2, 1-12 (2011).
- Häuser, R., Stellberger, T., Rajagopala, S. V., Uetz, P. Array-Based Yeast Two-Hybrid Screens: A Practical Guide. Two Hybrid Technologies: Methods and Protocols, Methods in Molecular Biology. , 21-38 (2012).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten