
Method Article
La disección y de montaje lateral de embriones de pez cebra: Análisis para el Desarrollo de la médula espinal
En este artículo
Resumen
Los procesos de desarrollo tales como la proliferación, el patrón, la diferenciación, y guía de axones se pueden modelar fácilmente en la médula espinal pez cebra. En este artículo se describe un procedimiento de montaje para los embriones de pez cebra, que optimiza la visualización de estos eventos.
Resumen
La médula espinal pez cebra es un modelo de investigación eficaz para la investigación del sistema nervioso por varias razones. En primer lugar, genética, transgénicos y de genes enfoques desmontables pueden ser utilizados para estudiar los mecanismos moleculares que subyacen desarrollo del sistema nervioso. En segundo lugar, las grandes garras de embriones de desarrollo sincronizados proporcionan grandes tamaños de muestras experimentales. En tercer lugar, la claridad óptica del embrión de pez cebra permite a los investigadores visualizar progenitoras, gliales, y poblaciones neuronales. Aunque los embriones de pez cebra son transparentes, espesor de la muestra puede impedir la visualización microscópica efectiva. Una razón para esto es el desarrollo en tándem de la médula espinal y el tejido somite suprayacente. Otra razón es la bola yema grande, que todavía está presente durante los períodos de la neurogénesis temprana. En este artículo, nos demuestran microdisección y la eliminación de la yema de embriones fijos, lo que permite la visualización microscópica, preservando el tejido circundante somito. Nosotros also demostrar montaje semipermanente de embriones de pez cebra. Esto permite la observación de neurodesarrollo en los ejes dorso-ventral y anterior-posterior, ya que conserva la tridimensionalidad del tejido.
Introducción
Visualización de la médula espinal en el pez cebra se inhibe por un número de factores. Debido al espesor de los somitas suprayacentes y la ubicación interna de la médula espinal, se requiere una distancia considerablemente larga de trabajo para alta resolución celular. El balón ha yema (que todavía está presente durante las primeras etapas de la neurogénesis) aumenta aún más la distancia de trabajo necesario, y se daña fácilmente con la presión de un cubreobjetos. Además, los restos de yema dañado prohíbe la visualización clara de los tejidos. Aunque son posibles secciones transversales en el eje (DV)-dorso-ventral, que no permiten fácilmente la visualización simultánea en el eje antero-posterior (AP) 1.
Para superar estos obstáculos, los embriones se disecan y se montaron en portaobjetos. Este procedimiento proporciona varias ventajas. En primer lugar, embriones de pez cebra se encuentran fácilmente con la cara lateral hacia arriba, lo que facilita la visualización eje AP. En segundo lugar, la eliminación de la yema de BALl disminuye la distancia de trabajo requerido, y limita los escombros. En tercer lugar, este procedimiento de montaje permite por tanto fluorescente y microscopía de campo claro. En cuarto lugar, los embriones montados son estables durante meses a 4 ° C, lo que permite la visualización espécimen prolongado. Por último, la progresión del desarrollo se da en que los segmentos anteriores son más maduros que los segmentos posteriores. Con el fin de asegurar que por etapas hemisegments espinales emparejados se comparan entre embriones, que recubre el tejido somite se utiliza como una guía. Por ejemplo, la décima somite más posterior se superpone a la décima hemisegment espinal, que es equivalente de desarrollo en embriones de etapa con ajuste. Este procedimiento de montaje de los embriones intactos permite una fácil identificación de los somitas.
Utilizamos la genética y genes tecnologías knock-down para estudiar los mecanismos de guiado de los axones en la médula espinal en desarrollo. En particular, se ha determinado que se requieren robo2, ROBO3 y DCC para comisural Asce PrimariaNding (COPA) pathfinding axón. Usando el procedimiento de montaje descrito aquí, hemos sido capaces de examinar el crecimiento ventral, cruce la línea media, ancho de comisura, dorsal y anterior 2,3 de crecimiento. Este procedimiento de montaje también se puede aplicar a DV o AP patrón de la médula espinal. Hemos utilizado este procedimiento para determinar los papeles dispares de efectores aguas abajo de Wnt en DV patrón de la médula espinal. El uso de este montaje, hemos sido capaces de obtener la resolución de los marcadores de DV en el nivel de células individuales, y determinar los cambios de modelado minutos como consecuencia de la alteración de recepción de señales Wnt 4,5. Este procedimiento de montaje también permite el cálculo del índice mitótico a través de anti-fosfohistona 3 o etiquetado BrdU (proliferación de progenitores) 5 diferenciación.
Protocolo
1. Montaje de los embriones (al término de técnica de visualización seleccionado, como la inmunocitoquímica, hibridación in situ, etc)
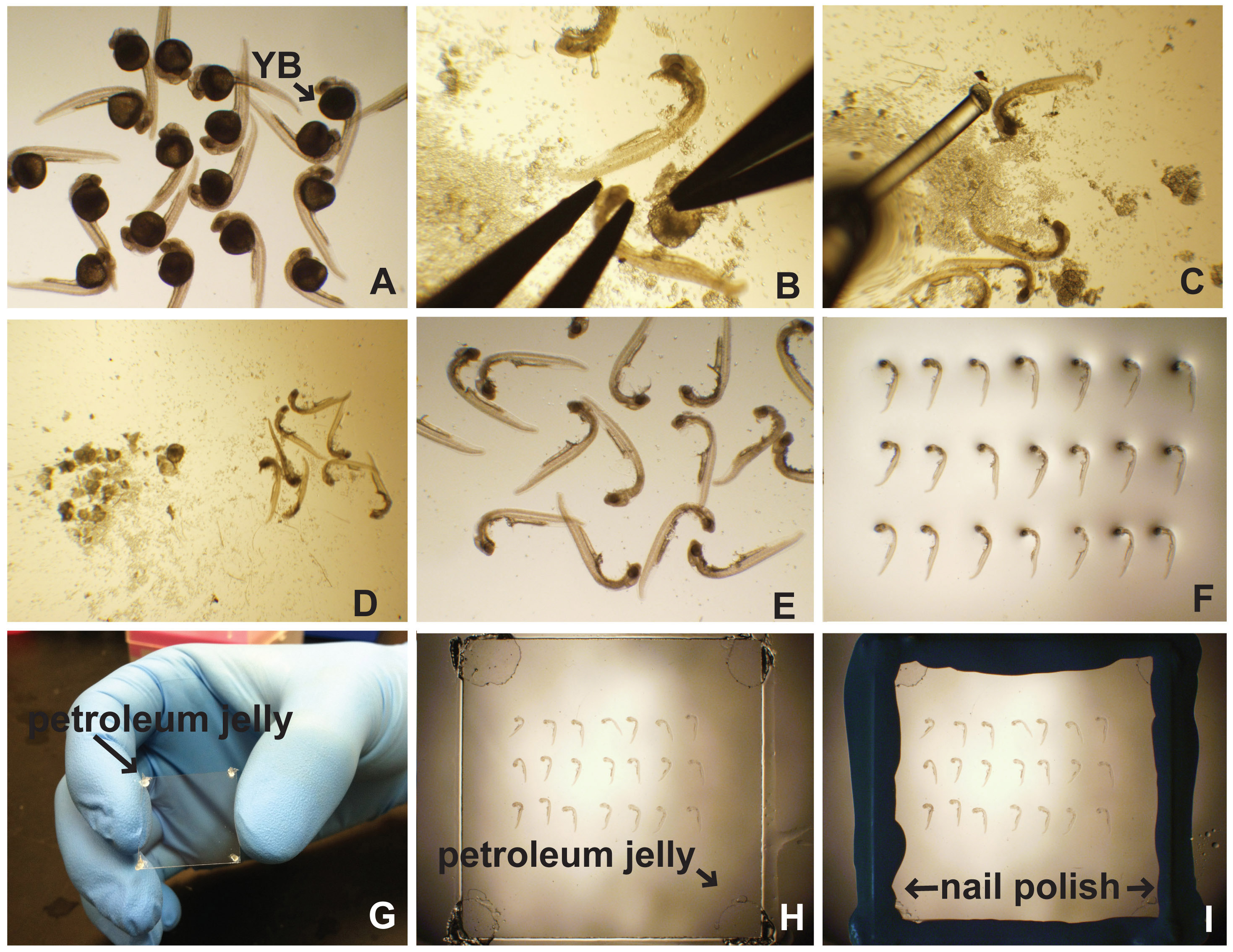
- Coloque los embriones de un plato Petri de 35 mm llena con tampón de elección (Figura 1A). La elección de tampón se basa en el procedimiento de etiquetado. Típicamente tampones basados en PBS son adecuados.
- Bajo un microscopio de disección, y el uso de fórceps, asegurar la cabeza con un par de pinzas mientras tira de la yema de distancia con el otro par (Figura 1B). Alternativamente, use un alfiler de insectos en un soporte de pasador de insectos para tirar suavemente la yema.
- Usando el poker embrión (línea de pesca (0,41 mm de diámetro) pegado a cualquiera de un tubo capilar o una pipeta Pasteur) mover los embriones a una parte de la placa de Petri que no tiene una gran cantidad de restos de yema (figuras 1C-D).
- Utilizando una pipeta Pasteur de vidrio, aspirar los embriones en tan poco líquido como sea posible, y gtemente pipetear ellos en un portaobjetos (Figura 1E).
- Suavemente absorber y expulsar el exceso de líquido utilizando un laboratorio toallita. Evite el contacto con los embriones.
- Añadir una gota de medio de montaje a los embriones. Para etiquetas fluorescentes, utilice un reactivo Antifade de elección. Para señales colorimétricas, 70% de glicerol se puede utilizar como un medio de montaje.
- Usando el poker embrión, oriente los embriones en sus lados en filas (Figura 1F).
- Añadir una "pizca" de vaselina o grasa de alto vacío a cada esquina de un cubreobjetos (Figura 1G). Esto evita el daño del embrión a partir de una compresión excesiva.
- Coloque el cubreobjetos (lado vaselina hacia abajo) en la parte superior de los embriones (Figura 1H).
- Golpear suavemente el cubreobjetos en cada esquina hasta que los medios de montaje (70% de glicerol o reactivo de montaje anti-fade) está tocando el cubreobjetos (Figura 1H).
- Si es necesario, agregar material de montaje con la pipeta justo al lado del coverslip, usando una pipeta en el intervalo de 20-200 ml. Se mecha bajo.
- El uso de un laboratorio de limpiar, limpiar completamente todo el cubreobjetos. Asegúrese de no aplicar presión en el cubreobjetos, ya que podría dañar los embriones. Esta área debe estar seca y no tienen vaselina o medio de montaje sobre el mismo.
- Utilice el esmalte de uñas para sellar alrededor del borde del cubreobjetos (Figura 1I).
Resultados
Cuando elucidar los mecanismos que subyacen a diversos eventos de desarrollo tales como patrón celular, la diferenciación, y guía de axones, es importante ser capaz de visualizar las células en el contexto de su tejido. Defectos dorsoventral de modelado típicamente se presentan como un desplazamiento ventral o dorsal en el dominio de la expresión de factores de transcripción en el pax, nkx o familias dbx 4. A veces, los cambios pueden ser sutiles, que comprende sólo unos diámetros celulares 4. Análisis de corte transversal permite este tipo de análisis. Sin embargo, reactivos con las señales más débiles, o secciones transversales que no son perpendiculares al eje de AP pueden limitar la interpretación. Además, el análisis de la sección transversal no toma en cuenta fácilmente si los cambios persisten en el eje AP, y se basa en la disponibilidad de un criostato. Esta limitación se elude mediante vistas laterales de la médula espinal. Como se observa en la Figura 2B, Nkx6.1 ARNm es dISTRIBUIDO en aproximadamente la mitad ventral de la médula espinal en 24 horas después de la fecundación (HPF), dentro del dominio progenitora. Las células progenitoras son distinguibles de las células mitóticas mensaje debido a la presencia de la placa de piso, que se encuentra en la parte medial de la médula espinal. Visto con la óptica DIC, las células individuales son claramente distinguibles, y un límite definido entre la expresión y nonexpressing células es evidente.
La salida del ciclo de células progenitoras se acompaña de cambios en la expresión génica. Por ejemplo, todas las neuronas post-mitóticas expresan HuC / D, que es detectable con 1,5 inmunocitoquímica. sondas de ARNm para los islotes, gata, y la familia VSX etiquetar las neuronas post-mitóticas específicos de posición coherente y número dentro de cada hemisegment médula espinal 6. Además, los receptores de orientación axón tales como los de la familia de robo se expresan en poblaciones restringidas de neuronas postmitóticas (Figura 2C) 2,7,8. Montaje lateral de embriones permite conteos precisos de las neuronas post-mitóticas. Del mismo modo, los progenitores que continúan dividiendo pueden cuantificarse con anti-fosfato histona 3 inmunocitoquímica, así como el etiquetado BrdU. Además, la muerte celular se puede evaluar con TUNEL etiquetado 4-6.
A las 24 HPF, los axones de las neuronas siguientes postmitóticas pueden visualizarse con varios métodos, y se distinguen en el único nivel celular: Dorsal Lateral Ascendente (Dola), comisurales Primaria ascendente (COPA), comisurales Secondary ascendente (COSA), ventral longitudinal Descendente (Veld), Kolmer-Agdur (KA), comisurales bifurcar / longitudinales (COB / L), circunferencial Ascendente (CIA), circunferencial Descendente (CID), y Unipolar comisural Descendente (CM subyacente), Rohon-Barba (RB), las neuronas motoras ( M) 9. Los axones se extienden desde estas neuronas en la parte dorsal, ventral, anterior y posterior direcciones. Ellos rama, cruzan la línea media, y salen tanto de la dorsaly ventral de la médula espinal. Cuando se combina con microscopía confocal de barrido láser, estos diversos comportamientos celulares son evidentes en los embriones montados lateralmente, usando inmunocitoquímica y proteínas fluorescentes codificadas genéticamente, tales como GFP 2. En la Figura 2D, se utilizó ZNP-1 inmunocitoquímica para etiquetar motoneuronas en desarrollo. Motoneuronas salen de la médula espinal ventral para inervar la musculatura que rodea el desarrollo.

Figura 1. La disección y lateral de montaje de embriones de pez cebra. (A) Después del procesamiento para inmunofluorescencia, hibridación in situ, etc, los embriones se colocan en un plato de Petri de 35 mm llena de PBT (PBS con 0,5% de Triton X-100) o PTW (PBS con 0,1% de Tween-20). La yemabola (YB) es evidente. (B) La yema se separa por asegurar la cabeza del embrión seguida de disección cuidadosa. (C) Un póquer embrión (línea de pesca pegado a una pipeta Pasteur) se utiliza para embriones separados y escombros yema de (D). Embriones limpiados se pipetean sobre un portaobjetos de microscopio (E) y alineados (F). Jalea (G) Petróleo se aplica a las esquinas de la cubreobjetos. (H) El cubreobjetos se coloca suavemente en la parte superior de los embriones en medio de montaje. Polaco (I) Nail se utiliza para sellar los bordes del cubreobjetos. Haga clic aquí para ver la imagen más grande .
{kind=link}

< strong> Figura 2. Análisis de los patrones de expresión de genes y axón pathfinding en montado lateralmente embriones de pez cebra. (A) Dibujo de un embrión de pez cebra en 24 HPF. En BD, aproximadamente cuatro hemisegments por encima de la extensión del saco vitelino se visualizan (área del recuadro). (B) Nkx6.1 se expresa en la médula espinal ventral, incluyendo la placa de piso (FP). (C) ROBO3 se expresa en las neuronas postmitóticas (PMN). La placa de suelo y progenitor zona no es evidente en este plano focal más lateral. En B, C, la médula espinal está delimitado por un soporte en la parte izquierda de la imagen. (D) microscopía confocal se utilizó para la imagen ZNP-1 de inmunofluorescencia, que las etiquetas de los axones motores que salen de la médula espinal ventral. En todas las imágenes, es anterior a la izquierda y dorsal es hacia arriba. A larga distancia de trabajo 40x, la inmersión en agua (NA 0.8) lente se utilizó (3,3 mm).oad/50703/50703fig2highres.jpg "target =" _blank "> Haga clic aquí para ver la imagen más grande.
Discusión
Montaje lateral del pez cebra fija ayudas embriones visualización del desarrollo de la médula espinal. A través de la eliminación de la pelota yema de huevo, se reduce la distancia de trabajo requerido para imágenes microscópicas excepcional. Nuestros resultados representativos se limitan a la etapa 24 HPF, sin embargo, esta técnica se puede utilizar tan pronto como 18 HPF, aunque embriones anteriores son más difíciles de diseccionar debido al tamaño del embrión y la fragilidad del tejido. Esta técnica también es aplicable a los embriones de más de 24 HPF. Ayudado por la claridad óptica de la embriones de pez cebra, la capacidad de realizar pantallas adelante genéticos, genética inversa, y enfoques transgénicos, varios procesos de desarrollo pueden ser investigados. Esto incluye los índices mitóticos de progenitores neuronales con marcadores como BrdU y anti-fosfato histona marzo 4 a 6 y los patrones a través de la expresión de marcadores progenitoras neuronales y gliales en el pax, nkx, dbx, y familie oligs 1,4-6. Los reactivos que reportan determinación glial y neuronal (como la proteína glial fibrilar ácida (GFAP) y HuC / D) 1,4,5,10 se puede utilizar. Diferenciación subtipo neuronal puede ser analizada a través de la expresión de los islotes, vsx, engrailed y genes gata 1,4-6. Y, por último, el análisis de la orientación axón es posible a través de anticuerpos tales como anti-tubulina acetilada, 3A10 y ZNP-1 2,11,12. Aunque otros sistemas modelo de vertebrados (ratón y pollo) se utilizan para estudiar el desarrollo de la médula espinal, sólo el permiso de pez cebra visualización simultánea de todos los ejes de desarrollo en el embrión intacto.
La limitación obvia de este enfoque es que los embriones se fijan, que excluye de imágenes en vivo. De hecho, la eliminación de (o daño a) los resultados de yema de huevo en la rápida muerte embrionaria, por lo que no es posible reducir la distancia de trabajo en embriones vivos utilizando esta técnica. Sin embargo, una gran ampliación de la médula coimágenes rd en embriones vivos es posible. Con el fin de preservar la yema durante de imágenes en vivo, los embriones se pueden colocar en las diapositivas de depresión, que proporcionan un espacio más grande entre la muestra y el cubreobjetos. Alternativamente, varios cubreobjetos de 22 mm x 22 mm se pueden pegar a cada diapositiva de un 3 en 1 x en portaobjetos de microscopio. Los cubreobjetos sirven como un "puente" para el cubreobjetos encima. Si los embriones son mayores de 18 HPF, tricaína se utiliza para anestesiar a los embriones para evitar el movimiento. Si bien se requiere una distancia de trabajo de 0,288 mm para los embriones deyolked en 24 HPF, cuando se trabaja con embriones vivos incrustados en agarosa, con yemas intactas, una lente microscópica con una distancia de trabajo de 3,3 mm es suficiente. Alternativamente, cultivos de explantes derivados a partir de embriones deyolked pueden ser visualizados utilizando una técnica de inmovilización coágulo de plasma 13. Varias poblaciones de células se pueden observar a través del uso de proteínas fluorescentes codificadas genéticamente, tales como GFP, mCherry, o el fotoconductortinte convertibles Kaede (por nombrar algunos). Además, las células marcadas con fluorescencia dentro de embriones quiméricos también se pueden ver con este enfoque 14.
Una técnica de montaje consistente que es estable durante meses es una herramienta crítica para los biólogos celulares y de desarrollo. Esta técnica sencilla y reproducible, lo que permite una fácil comparación entre los diferentes ensayos experimentales. Además, la alineación de los embriones en filas permite fácilmente la identificación de los embriones específicos para su posterior análisis.
Divulgaciones
No existen intereses económicos que compiten por cualquiera de autor.
Agradecimientos
Skidmore Facultad Donaciones para el Desarrollo financió la preparación y publicación de este manuscrito.
Materiales
| Name | Company | Catalog Number | Comments |
Petri dishes 35 mm x 10 mm | VWR | 25373-041 | |
| Dumont Forceps #3 | Fisher Scientific | NC9839169 | |
| Cover glass | Fisher Scientific | 12-541-B22X22-1.5 | |
| Slides | Fisher Scientific | 12-550-343 | |
| SlowFade Gold | Fisher Scientific | S36936 | |
| ProLong Gold | Life Technologies | P36934 | |
| Petroleum Jelly | any grocery store |
| |
| Loop Holders | VWR | 80094-482 | |
| Insect Pins | Fine Science Tools | 26002-10 | |
| Nickel Plated Pin Holders | Fine Science Tools | 26016-12
| |
Olympus Stereomicroscope | Olympus | SZ61 |
Referencias
- Gribble, S. L., Nikolaus, O. B., Dorsky, R. I. Regulation and function of Dbx genes in the zebrafish spinal. Dev. Dyn. 236 (12), 3472-3483 (2007).
- Bonner, J., et al. Midline crossing is not required for subsequent pathfinding decisions in commissural neurons. Neural Dev. 7 (1), 18 (2012).
- Ross, A. B. J. Activation of Wnt signaling using Lithium Chloride: Inquiry-Based Undergraduate Laboratory Exercises. Zebrafish. , (2012).
- Bonner, J., et al. Proliferation and patterning are mediated independently in the dorsal spinal cord downstream of canonical Wnt signaling. Dev. Biol. 313 (1), 398-407 (2008).
- Gribble, S. L., et al. Tcf3 inhibits spinal cord neurogenesis by regulating sox4a expression. Development. 136 (5), 781-789 (2009).
- England, S., et al. Roles of Hedgehog pathway components and retinoic acid signalling in specifying zebrafish ventral spinal cord neurons. Development. 138 (23), 5121-5134 (2011).
- Challa, A. K., Beattie, C. E., Seeger, M. A. Identification and characterization of roundabout orthologs in zebrafish. Mech Dev. (1-2), 101-101 (2001).
- Lee, J. S., Ray, R., Chien, C. B. Cloning and expression of three zebrafish roundabout homologs suggest roles in axon guidance and cell. 221 (2), 216-230 (2001).
- Downes, G. B., Waterbury, J. A., Granato, M. Rapid in vivo labeling of identified zebrafish neurons. Genesis. 34 (3), 196-202 (2002).
- Kim, H., et al. Notch-regulated oligodendrocyte specification from radial glia in the spinal cord of zebrafish embryos. Dev. Dyn. 237 (8), 2081-2089 (2008).
- Sylvain, N. J., Brewster, D. L., Ali, D. W. Zebrafish embryos exposed to alcohol undergo abnormal development of motor neurons and muscle fibers. Neurotoxicol. Teratol. 32 (4), 472-480 (2010).
- de Soysa, T. Y., et al. Macondo crude oil from the Deepwater Horizon oil spill disrupts specific developmental processes during zebrafish embryogenesis. BMC Biol. 10 (40), (2012).
- Langenberg, T., Brand, M., Cooper, M. S. Imaging brain development and organogenesis in zebrafish using immobilized embryonic explants. Dev Dyn. 228 (3), 464-474 (2003).
- Deschene, E. R., Barresi, M. J. Tissue Targeted Embryonic Chimeras: Zebrafish Gastrula Cell Transplantation. J. Vis. Exp. (31), (2009).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados