
Method Article
Ratón Rombencéfalo
En este artículo
Resumen
Neuronas embrionarias nacen en la zona ventricular del tubo neural, pero migran a alcanzar los objetivos adecuados. Branchiomotor facial (FBM), las neuronas son un modelo útil para estudiar la migración neuronal. Este protocolo describe la cultura wholemount ex vivo de hindbrains embrión de ratón para investigar los mecanismos que regulan la migración FBM.
Resumen
Neuronas embrionarias nacen en la zona ventricular del cerebro, pero posteriormente migran a nuevos destinos para alcanzar los objetivos adecuados. Por lo tanto, Descifrar las señales moleculares que guían la migración de manera cooperativa neuronal en el cerebro embrionario es importante entender cómo las complejas redes neuronales se forman que luego sostener la vida postnatal. Branchiomotor facial (FBM), las neuronas en el cerebro posterior embrión de ratón migran desde rombómero (R) 4 caudalmente para formar los núcleos faciales apareados en la región derivada-R6 de la parte posterior del cerebro. Aquí le ofrecemos un protocolo detallado para wholemount cultivo ex vivo de hindbrains embrión de ratón adecuados para investigar las vías de señalización que regulan la migración FBM. En este método, hindbrains de embriones E11.5 ratón se disecan y se cultivaron en una preparación de libro abierto en insertos de cultivo celular de 24 horas. Durante este tiempo, las neuronas FBM migran caudalmente hacia R6 y pueden estar expuestos a anticuerpos función de bloqueo y los pequeños moleculES en los medios de cultivo o perlas de heparina cargadas con proteínas recombinantes para examinar las funciones para las vías de señalización implicadas en la orientación de la migración neuronal.
Introducción
Neuronas embrionarias nacen en la zona ventricular del cerebro, pero posteriormente migran a nuevos destinos para llegar a las regiones de destino apropiados que se encuentran a una gran distancia. El correcto posicionamiento de los cuerpos celulares neuronales en lugares apropiados a lo largo de los ejes dorso-ventral y anterior-posterior del cerebro en desarrollo es esencial para el cableado correcto, la supervivencia, y la función de estas neuronas después de la etapa migratoria 1-4. De manera similar a los mecanismos moleculares que controlan axón orientación 5-7, se cree conjuntos combinatorios de las señales de atracción y repulsión para guiar las neuronas que migran 1,8. Sin embargo, debido a las interacciones de múltiples tipos de células, las señales que controlan la migración neuronal han sido estudiados con menos intensidad que los involucrados en el direccionamiento del axón, que se puede estudiar la célula autónoma. El cerebro posterior desarrollo de los vertebrados se ha utilizado en varios estudios recientes para entender los mecanismos moleculares y celularesde la migración neuronal, por ejemplo en la chica, el ratón y el pez cebra 1-4,9. Este órgano contiene varios tipos diferentes de neuronas, incluyendo varios subtipos de precerebellar y neuronas motoras 5,7,10,11.
Motorneuron Rombencéfalo nacen en la zona ventricular cerca de la placa del piso, y se diferencian en subconjuntos específicos de acuerdo a su rhombomere de origen 1,12. El branchiomotor facial (FBM), las neuronas se generan en rombómero (R) 4 en el cerebro posterior y se extienden sus axones dorsalmente a través de un punto de salida R4 en el segundo arco branquial para inervar los músculos faciales 2,9,13. Neuronas FBM de pez cebra y ratones proporcionan excelentes modelos para estudiar los mecanismos moleculares y celulares de la migración neuronal en un proceso que se visualiza fácilmente, debido a que estas neuronas reproducible translocan su cuerpos celulares en un proceso espaciotemporalmente bien definida. En ratones, las neuronas migran FBM primera cauDally través de R5 y entonces tanto en sentido caudal y ventral para alcanzar su posición final en el lado pial del cerebro posterior en el territorio de r6, donde forman los núcleos apareados del nervio craneal VII (Viin) 10,11,14. En el pez cebra, las neuronas migran ventralmente FBM inicialmente y luego cambian de dirección en el límite de la R4-R5 continuar migrando hacia la superficie pial de una manera dependiente de la laminina 4,12,15,16. Esta migración procede durante un período de varios días en el desarrollo y se puede dividir en fases de la migración tangencial y radial, lo que permite la identificación de moléculas que median estos dos procesos distintos. En contraste, las neuronas FBM de la parte posterior del cerebro de embriones de pollo permanecen en R4 3,13,17-19.
Durante su migración, las neuronas FBM se pueden identificar, al igual que otros tipos o neuronas motoras, a través de su expresión del factor de transcripción de los islotes homoeodomain 1 (ISL1) 14. Por lo tanto, Mayoristasla tinción de inmunofluorescencia emount o hibridación in situ para este marcador en diferentes etapas de desarrollo revela la corriente migratoria distinta de FBM somas se extiende desde R4 a R6 en el pez cebra o el ratón 4,15,16. Por otra parte, los reporteros transgénicos fluorescentes tales como ISL1-GFP se han utilizado como herramientas adecuadas para visualizar la migración de neuronas en el pez cebra FBM 3,17-19. Además de su idoneidad para la formación de imágenes, muchos investigadores han estudiado la migración de las neuronas FBM en el desarrollo de pez cebra, porque sus embriones de vida libre se pueden manipular fácilmente con técnicas de trasplante de células y compuestos farmacológicos aplicados directamente al agua del acuario. En contraste, el embrión de ratón desarrolla encerrado en el útero, lo que impide la implantación de perlas que transportan las señales de orientación o la administración de anticuerpos de la función de bloqueo que no cruzan la barrera placentaria. Además, los compuestos farmacológicos administrados a la madre embarazada puede tener sinefectos secundarios deseados que pueden afectar indirectamente la embriogénesis. Eludir esta limitación, hemos desarrollado un método ex vivo para la cultura posterior del cerebro del ratón todo que es compatible con la migración de las neuronas FBM y la supervivencia durante 24 horas después de la explantación 7,16. Este método permite la manipulación farmacológica fácil, la implantación de los granos que llevan las señales de orientación o la administración de anticuerpos de la función de bloqueo y también podría ser adaptado para estudiar la migración de otros subtipos neuronales en el cerebro posterior en diferentes etapas de desarrollo.
Protocolo
1. Opcional: Preparar Beads Heparina Affi-gel (perlas de gel) para FBM Ensayo de atracción
NOTA: preparar perlas de gel de al menos 1 día antes de comenzar el procedimiento de explante.
- Lavar 100 l de gel de heparina suspensión de perlas con PBS estéril durante 20 min en un rodillo a temperatura ambiente (RT).
- Perlas de pellets en una centrífuga de mesa durante 5 minutos a 13.000 x g. Añadir PBS estéril y repetir el proceso de lavado de 4x.
- Después del lavado final, quitar PBS y remojar los granos en un pequeño volumen de una solución estéril que contiene la proteína recombinante de elección, teniendo cuidado de cubrir las perlas con la solución. Este protocolo utiliza 100 ng / l VEGF165 humana recombinante en PBS para reproducir un experimento publicado previamente 16.
- Incubar las perlas de gel de heparina con la solución de proteína recombinante durante un mínimo de 12 horas y un máximo de 1 semana en un rodillo a 4 º C.
2. Revestimiento de la Cultura Insertars
Explantes cerebro posterior se cultivan en la cultura Corning inserta con un tamaño de poro de 8 micras o inserciones equivalentes. Insertos de cultivo se pueden reutilizar después de la finalización del protocolo, siempre que se lavan con agua destilada, esterilizados con etanol, y se almacenan en etanol al 70% hasta que se necesite.
NOTA: Los siguientes pasos deben llevarse a cabo en una campana de flujo en condiciones estériles.
- Preparar los medios de cultivo de explante que consiste en medio Neurobasal suplementado con B27 (20 l / ml), glucosa (6 mg / ml) y penicilina / estreptomicina (5 mg / l).
- Cultura de lavado inserta con PBS estéril durante 5 min y seco durante 5-10 min bajo la campana de flujo.
- Coloque un inserto de cultivo en cada pocillo individual de una placa 12well. Nota: Las inserciones pueden necesitar un pequeño empujón para encajar perfectamente en el pozo.
- Cubrir los insertos de cultivo con 10-20 mg / ml de laminina de ratón en medio Neurobasal y colocarlos en una incubadora de cultivo de tejidos (37 º C, 5% de CO 2) durante 1 hora. Nota: revestimiento debe llevarse a cabo en el día de la explantación.
3. Disección de Hindbrains de E11.5 embriones de ratón
- Cull temporizado ratón hembra embarazada con un procedimiento éticamente aprobado el día embrionario (E) 11,5 y colocar el útero que contiene los embriones en un plato de plástico de 100 mm con medio L15 enfriado en hielo.
NOTA: Todos los pasos de disección se deben realizar en L15 enfriado en hielo. - Utilizando un microscopio de disección y Dumont relojero fórceps número 5, rasgar la pared muscular del útero para exponer los embriones, suelte cada embrión, cortar el cordón umbilical, y cuidadosamente quite el saco vitelino.
- Usando una pipeta Pasteur de plástico con una abertura de gran calibre, transferir cada embrión en una placa de plástico limpio con L15 enfriado en hielo.
- Con unas pinzas Dumont, decapitar al embrión justo por encima de las extremidades anteriores. Si el experimento requiere el genotipado de los embriones, recolectar muestras de tejido para el aislamiento del ADN genómico (por ejemplo, una pequeña piece del saco vitelino o punta de la cola 16,20).
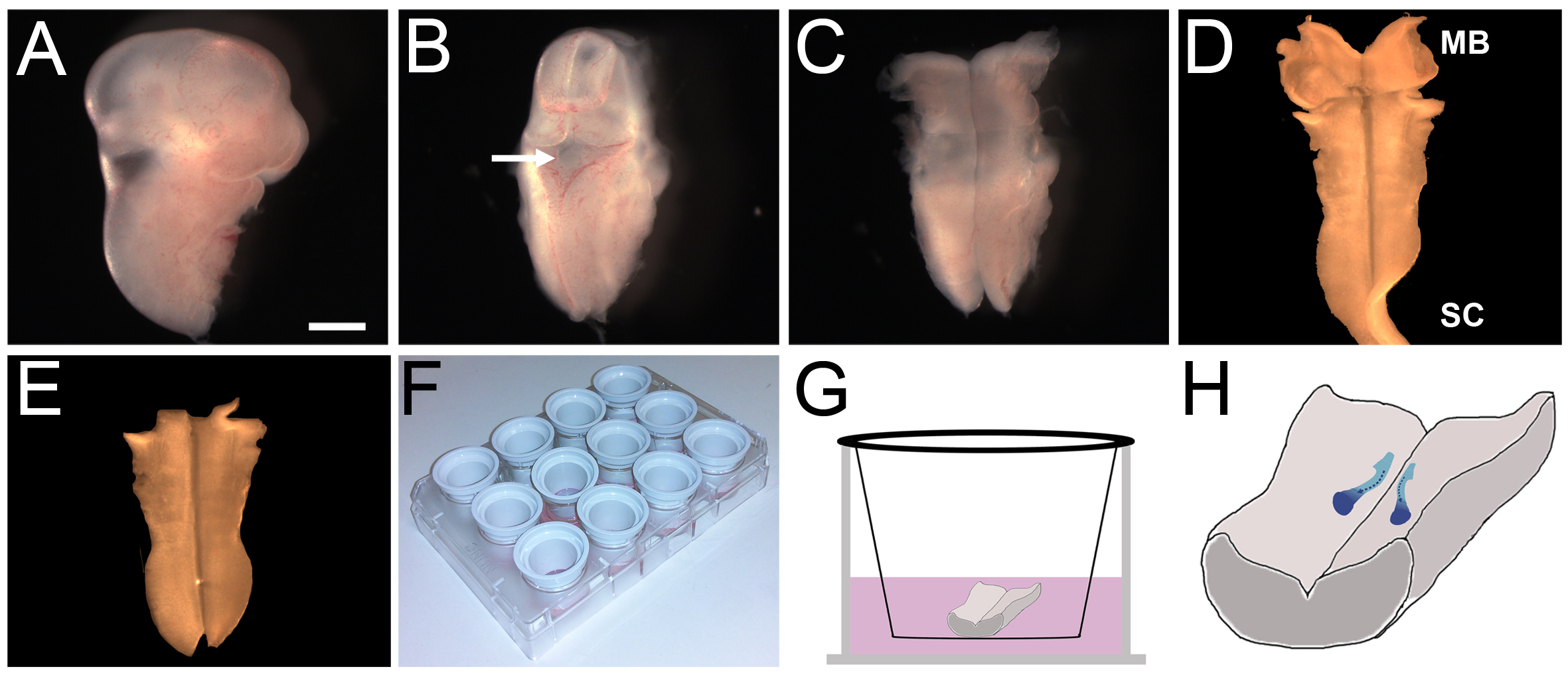
- Gire la parte dorsal de la cabeza y de identificar el 4 º ventrículo, que está cubierto por una delgada capa de tejido (Figura 2B). Perforar con cuidado el roofplate y empezar a pelar lejos caudalmente a lo largo de la línea media sobre la parte posterior del cerebro posterior y la médula espinal, y el rostral sobre el cerebro medio. El cerebro posterior ahora debe ser expuesto (Figura 2C).
- Burlarse cuidadosamente lejos la mesénquima de la cabeza restante y cualquier meninges que se adjuntan a la parte pial de la parte posterior del cerebro (Figura 2D).
- Retire el tejido del cordón del cerebro medio y la médula para que el cerebro posterior se despliega y se acueste en una preparación a libro abierto (Figura 2E).
- El uso de un gran calibre Pasteur plástico pipeta, transferir cada cerebro posterior diseccionado a una placa de 12 pocillos que contiene helado L15 y guardarlos en hielo hasta que todos hindbrains se han disecado.
- El uso de un de gran calibre Pasteur plástico pipeta, transferir un pecadogle cerebro posterior a un plato vacío, manteniendo una preparación de libro abierto, cara ventricular hacia arriba, en una gota de aproximadamente 100 l L15.
- Transferencia de unos pocos microlitros de las perlas de gel de heparina incubadas a la misma gotita. Nota: Debido a que los tamaños de los granos son variables, es aconsejable transferir alrededor de 10 cuentas y luego elegir un tamaño óptimo para el trasplante en el cerebro posterior.
- Hacer un pequeño desgarro en el tejido de cerebro posterior y insertar cuidadosamente 1-3 perlas de gel en el tejido cerebro posterior en el nivel de R5 / 6, a mitad de camino entre la línea media y el borde lateral de la parte posterior del cerebro, disminución de los mismos en el tejido de modo que el talón se coloca justo debajo de la superficie posterior del cerebro.
NOTA: El procedimiento de disección puede tardar entre 5-20 min / cerebro posterior, dependiendo de la experiencia, y puede prolongarse durante un período prolongado si camadas son grandes, en todo caso, hindbrains deben estar en la cultura de no más de 3 horas post-mortem para la buena resultados.
4. RombencéfaloExplante Cultura
- Retire la placa que contiene los insertos de cultivo de la incubadora, aspirar la solución de revestimiento laminina.
- Coloque un inserto de cultivo en una placa de cultivo separado lleno de helado L15 y, con un diámetro ancho Pasteur plástico pipeta, transferir cada lado ventral posterior del cerebro para arriba sobre la inserción del cultivo (Figura 2G). El cerebro posterior debe quedar completamente plana sobre la membrana de inserción.
- Levante con cuidado la inserción del cultivo de la antena y aplíquelo varias veces en un pañuelo de papel limpio para eliminar el exceso de líquido. Este proceso asegura que el cerebro posterior se adhiere a la inserción de la cultura en una preparación a libro abierto plana. Si el cerebro posterior se queja, su tejido puede irreversiblemente crecer juntos.
- Llene la placa de 12 pocillos original con 500 l de medios de cultivo precalentado y coloque el inserto de nuevo en este pozo. Ajustar cuidadosamente el volumen con otro 400-600 l de los medios de comunicación para cubrir sólo la parte posterior del cerebro, lo que garantiza que el cerebro posterior no flotafuera de la membrana. Si flota, vuelva al paso 4.4 y repetir hasta que la parte posterior del cerebro se mantiene unida a la membrana.
- En esta etapa, es posible añadir inhibidores biológicos de interés para los medios de comunicación para estudiar su efecto sobre la migración de las neuronas FBM.
NOTA: Si va a implantar cuentas o administrar otros tratamientos, se recomienda mantener al menos 2 explantes de control bajo condiciones normales de crecimiento por experimento para asegurar que el experimento ha sido creado con éxito. - Incubar explantes durante 24-30 horas en un incubador de cultivo de tejidos (37 º C, 5% de CO 2).
5. Wholemount Inmunofluorescencia tinción de Rombencéfalo explantes
- Aspirar los medios de comunicación de cada pocillo, enjuagar en PBS y fijar, por 2 horas a 4 ° C con agitación suave en frío de hielo 4% de formaldehído (recién preparada o recién descongelado 4% de paraformaldehído disuelto en PBS). Nota: No intente retirar cerebro posterior de la inserción del cultivo antes de la fijaciónes completa.
- Enjuague 3 veces con PBS. Retire con cuidado los hindbrains de los insertos de cultivo utilizando pinzas Dumont. Algunos explantes son difíciles de pelar, pero por lo general se pueden levantar aplicando presión suave a través de la expulsión repetida PBS de una pipeta.
- Transfiera los hindbrains a 2,0 ml tubos de fondo redondo para el etiquetado de inmunofluorescencia. Permeabilizar hindbrains durante 30 min a temperatura ambiente (TA) en PBS que contenía 0,1% de Triton X-100 (PBT) con suave de rodillo.
- Incubar durante 1 hora a TA en PBT que contiene 10% de suero de cabra normal inactivado por calor con balanceo suave.
- Incubar explantes con balanceo suave a 4 ° C durante 5 días con el anticuerpo primario específico para ISL1, diluido 1:100 en PBT que contiene 1% de suero de cabra normal inactivado por calor.
- Lave los explantes a TA 4x con PBT durante 15 min cada uno.
- Incubar explantes con suave balanceo a temperatura ambiente durante 3 h con fluoróforo conjugado de cabra anti-ratón (por ejemplo, Alexa Fluor 488 cabra antiratón, diluido 1:200) en PBT que contiene 1% de suero de cabra normal inactivado por calor.
- Lave los explantes 4x a TA con PBT durante 15 min cada uno con suave balanceo.
- Postfix los explantes en formaldehído al 4% durante 30 minutos a temperatura ambiente.
- Cubra una placa de vidrio con 3 capas de cinta aislante color negro y los impuestos especiales con un bisturí a una pequeña plaza de la cinta en capas para crear un bolsillo para los explantes, como alternativa, utilizar un portaobjetos de vidrio depresión.
- Monte cada cerebro posterior en el reactivo SlowFade en un bolsillo y cubrir con un cubreobjetos de vidrio, evitando cuidadosamente para atrapar burbujas de aire y la imagen usando un microscopio confocal de barrido láser.
NOTA: Como alternativa a la inmunotinción, la hibridación in situ con ribosondas que reconocen FBMS (es decir ISL1 o PHOX2B) puede ser utilizado para visualizar las neuronas FBM 16,21.
Resumen de los pasos y el calendario
~ 14 días: el apareamiento para obtener embarazos E11.25 temporizado
Opcional: Bead preparación (PROTOCOLO DEl 1): ~ 2 horas, el día antes del aislamiento del embrión
Preparar insertos de cultivo y medios de comunicación (Protocolo 2): ~ 30 min, antes del aislamiento del embrión
Aislamiento de embriones y la disección posterior del cerebro (pasos 3.1 a 3.4): ~ 10 min / embriones
Disección Rombencéfalo (pasos 3.5 a 3.7): ~ 5-10 min / rombencéfalo
Procedimiento de explante (pasos 3.8 a 3.9): ~ 5-10 min / rombencéfalo
Opcional: la implantación de bolas (pasos 3.10-3.11): ~ 5-10 min / rombencéfalo
Cultura explante (paso 4.7): 24 horas
Fijación para la tinción de anticuerpos (paso 5.1): 2 hr
Procedimiento de tinción y de imagen (Protocolo 5): 5 días
Resultados
En esta sección se ilustra ejemplos de los resultados que se pueden obtener mediante el estudio de FBM migración de neuronas en el cerebro posterior del ratón a través de la cultura ex vivo. Se demuestra que las neuronas FBM en hindbrains explantadas de día 11 embriones de ratón primera experimentan una migración tangencial (Figura 3 A) y luego comienzan a montar el núcleo facial del motor (Figura 3B), similar a su comportamiento en el útero (ver Figura 1). Además, demuestran que la implantación de un cordón VEGF165-empapado atrae neuronas FBM (Figuras 3C y 3D), como se muestra anteriormente 16. Es importante destacar que, este protocolo permite estudiar la migración FBM en la ausencia de vasos sanguíneos o los vasos factores derivados que pueden influir en la migración FBM en el útero, porque la vasculatura nonperfused degenera en la cultura 16. De este modo, en el etiquetado de glóbulos inespecífico observado cuando se utiliza el anticuerpo ISL1 ratón en reciénhindbrains aislados de ratón (Figura 1) ya no está presente en el tejido posterior del cerebro después de 24 horas en cultivo (Figuras 3A-F). Finalmente, se muestran dos ejemplos de hindbrains que no fueron explantados correctamente y por lo tanto contienen neuronas FBM en una distribución anormal, ya sea porque el cerebro posterior no fue explantó muy pronto después del aislamiento del embrión (Figura 3E) o porque el tejido de cerebro posterior plegado en el transwell ( Figura 3F).

Figura 1. FBM migración neuronal confocal z-stack de hindbrains wildtype ratón después ISL1 immunolabeling wholemount y flatmounting;.. La línea media posterior del cerebro está indicado con un asterisco en todos los paneles (A) superficie ventricular de un cerebro posterior E11.5 en el área que contiene neuronas FBM ISL1-positiva (flecha), lo que demuestra su migración tangencial de R4 a R6;. la posición de R4, R5 y R6 se indica (A ') superficie pial de la mitad de la misma cerebro posterior en el área que contiene el Anlage . de uno de los núcleos FBM emparejados (indicados con Viin), así como otras poblaciones de neuronas ISL1-positivo (B) la superficie ventricular de un cerebro posterior E12.5 neuronas que contienen FBM que están migrando tangencialmente (flecha); la punta de flecha indica un ejemplo de un vaso sanguíneo que contiene las células circulantes que se inespecíficamente etiquetados por la reacción cruzada del anticuerpo secundario anti-ratón utilizado para detectar el anticuerpo IgG de ratón ISL1. (B ') superficie pial de la mitad de la misma rombencéfalo E12.5, que contiene uno de los núcleos FBM emparejados. La línea media se indica con un asterisco en cada panel. La barra de escala (todas las hojas): 200 m. V, ventral, P, de la pía madre./ 51397fig1highres.jpg "target =" _blank "> Haga clic aquí para ver la imagen más grande.

Figura 2. E11.5 ratón disección posterior del cerebro y vivo la cultura ex. (AE) Los pasos claves en el protocolo de disección de cerebro posterior E11.5; barra de escala:. 1 mm (A) Jefe del embrión después de que se corta, junto con el resto del embrión en el nivel de la extremidad anterior (B) La parte rostral del. la cabeza se eliminó y el resto de la tejido de la cabeza de modo que el 4 º ventrículo (flecha) estaba orientado hacia arriba. (C) El techo de la 4 ª ventrículo se desprendió, y el cerebro posterior fue expuesto por pelado tejido debajo de la parte posterior del cerebro de distancia rostral y caudal. (D) Se eliminó la membrana pial (nótese que en este examplio, algunos médula espinal cervical (SC) de tejido ha permanecido unido a la parte posterior del cerebro). (E) El exceso de cerebro medio (MB) y la médula espinal (SC) de tejido se ha eliminado para retener sólo el cerebro posterior. (insertos F) Cultura se recubrieron con laminina y se colocan en una placa de cultivo de tejidos de 12 pocillos. (G) Cada cerebro posterior se colocó sobre un inserto y se cubre con los medios de comunicación. (H) Representación esquemática de la trayectoria seguida por la migración de las neuronas FBM (azul) durante 24 horas de cultivo. Haga clic aquí para ver la imagen más grande.
{kind=link}

Figura 3. Ratón cerebro posterior ex vivo la cultura. (A, B) Un cerebro posterior E11.5 se cultivó durante 24 horas y immunofluoresc temente etiquetados para ISL1 para ilustrar FBM la migración de neuronas en un explante; tanto ventricular (A) y (B) de la pía madre lados de la parte posterior del cerebro se muestran (C, D) hindbrains littermate E11.5 se cultivaron en presencia de heparina implantado grano empapado. en PBS (C) o VEGF165 (D); en cuenta que las neuronas FBM migran hacia y en la perla VEGF165, y por lo tanto la corriente de migración ampliado aún más caudalmente en comparación con el lado no tratado de la misma rombencéfalo o el cerebro posterior que contiene la bola de control (E. , F) Ejemplos de explantes insatisfactorios cerebro posterior E11.5, en la que CESM no han emigrado de r4 (E), o en el que el tejido cerebro posterior doblado durante el cultivo (F). La línea media se indica con un asterisco en cada panel. La barra de escala (para todos los paneles): 200 m. V, ventral, P, de la pía madre."> Haga clic aquí para ver la imagen más grande.
Discusión
Este protocolo describe la cultura wholemount de hindbrains E11.5 ratón en un sistema transwell para estudiar la migración de las neuronas FBM. Este protocolo permite a neuronas motoras del cerebro posterior ratón para mantenerse con vida y la migración por un período de 24 horas, lo que permite la manipulación ex vivo. Este método tiene numerosas ventajas experimentales para los investigadores que tratan de identificar los mecanismos moleculares y celulares de la migración neuronal. Considerando que los ensayos de migración tradicionales explante pequeñas piezas de tejido neural en la matriz en placas de cultivo y permiten la observación de las neuronas individuales, ya que responden a estímulos exógenos, una gran ventaja del ensayo Transwell es su idoneidad para manipular las neuronas que migran dentro del entorno de órganos anfitrión y por lo tanto una más contexto fisiológico. Es importante destacar que las sustancias se pueden aplicar fácilmente a los explantes in vivo del cerebro posterior ex para probar su efecto sobre la migración neuronal, evitando los posibles efectos secundarios asociados con administesonar estas sustancias a un ratón embarazada. Por último, el modelo in vivo ex también permite el ensayo de sustancias que no cruzan la barrera placentaria, tales como anticuerpos función de bloqueo. Debido a estas ventajas, el cerebro posterior cultivo ex vivo proporciona un método alternativo y complementario a la utilización de embriones de pez cebra, que pueden ser tratados con moléculas pequeñas solubles en agua en el agua del acuario, o en el útero de la electroporación de cerebros de embriones, que requiere el uso de especializada equipo y es más difícil de dominar que la técnica de cultivo descrito aquí. Otra ventaja del protocolo descrito aquí es su susceptibilidad a la implantación de perlas empapadas en proteína recombinante u otros reactivos, por lo tanto, permite la aplicación de un método embriológico estándar desarrollado para manipular los embriones de pollo a un modelo de ratón de la migración neuronal. En particular, el modelo in vivo ex cultivo puede ser aplicado a hindbrains de ratones modificados genéticamente defectuosativa en moléculas específicas implicadas en la migración neuronal, tales como receptores de crecimiento o el factor de orientación, y en combinación con implantes de talón para probar si se pierde la capacidad de respuesta a los ligandos. Además de la manipulación farmacológica, el protocolo in vivo ex cultura también podría ser adaptado para electroporar vectores de expresión que podrían manipular la expresión de genes de interés; métodos apropiados para la electroporación se han descrito previamente 22,23. Este protocolo también puede estar adaptado para visualizar la migración de las neuronas por microscopía de lapso de tiempo en explantes de cerebro posterior de los ratones transgénicos que contienen las neuronas FBM marcados con fluorescencia, por ejemplo ISL1-Cre; Rosa26Yfp 21. Finalmente, este protocolo también se puede usar para estudiar otros tipos de neuronas que migran en el cerebro posterior, tales como los que forman la oliva inferior, aunque esto requeriría el uso de hindbrains en las etapas embrionarias de edad avanzada y puede requerir la cultura para un máximo de 48 h, dependiendo de la viabilidad neuronal Ex vivo.
Los pasos críticos y solución de problemas
Para el éxito de este protocolo, es crucial que los embriones se recogen temprano en E11.5 día, más cerca de E11.25, cuando FBM la migración de las neuronas recién ha comenzado. Sin embargo, no siempre es posible coger embriones en esta etapa del desarrollo debido a la variabilidad de apareamiento natural de los ratones, y, en consecuencia, puede haber algo de variabilidad en la extensión de FBM migración entre diferentes experimentos. La variabilidad en la migración FBM también se puede observar si el experimento no se completa dentro del marco de tiempo asignado, alrededor de 3 horas, como se puede ver en la figura 3E. Tejido Rombencéfalo de embriones de ratón E11.25 es delicado. Cuando la disección y durante todo el procedimiento de explante, es importante no romper el tejido cerebro posterior en las áreas de R4-R6 donde se encuentran las neuronas FBM. Debido a la naturaleza delicada de la Tesisproceso exión, y debido a la velocidad a la que hindbrains se colocan en la cultura influye en los resultados, el procedimiento puede tardar un par de la práctica funciona de dominar, en particular, antes de que se utilizaron muestras preciosas o reactivos. Por último, es importante que el tejido de cerebro posterior se coloca en una configuración libro abierto en la inserción del cultivo, porque el plegado de la parte posterior del cerebro durante el cultivo de tejido evitará la migración normal de FBM (véase la figura 3F).
Divulgaciones
Ninguno de los autores tiene intereses o conflictos de intereses en competencia.
Agradecimientos
MT es apoyado por una beca de doctorado [ref. 092839/Z/10/Z] y CR por un nuevo Premio Investigador [ref. 095623/Z/11/Z] de la Wellcome Trust.
Materiales
| Name | Company | Catalog Number | Comments |
| Eppendorf round-bottomed reagent tube, 2.0 ml (Safe-Lock) | VWR | 211-2120 | |
| Cell culture plates, 12-well | Thermo Scientific | 150628 | |
| Plastic cell culture dish, 100 mm | Thermo Scientific | 150288 | |
| 15 mm Netwell insert with Mesh Polyester Membrane | Corning | 3477 | |
| Watchmaker forceps, no. 5 | Dumont | 91150-20 | |
| Phosphate buffered saline | Sigma | P4417 | |
| Laminin mouse protein | Life Technologies | 23017-015 | |
| Primary antibody, Isl1 | Developmental Hybridoma Bank | 39.4D5 | Dilution 1/100 |
| Alexa Fluor 488-conjugated goat anti mouse secondary antibody | Life Technologies | A11029 | Dilution 1/200 |
| Neurobasal medium | Life Technologies | 21103 | |
| B27 supplement (50x) | Life Technologies | 17504-044 | |
| Leibovitz’s L-15 | Life Technologies | 21083-027 | |
| Penicillin/ streptomycin | Life Technologies | 15070 | |
| Glucose | VWR | 101174Y | |
| Heat-inactivated goat serum | Sigma | G9023 | |
| Triton X-100 | Sigma | T8787 | |
| Paraformaldehyde | Sigma | P6148 | |
| Slowfade Antifade Kit | Life Technologies | S-2828 | Alternative mounting solutions may be used |
| Microscope slides | VWR | 631-0912 | |
| Cover glass | VWR | 631-0137 | |
| Affi-Gel Heparin Beads | Biorad | 153-6173 | Glass, latex, or coloured beads may be used alternatively |
| Recombinant human VEGF165 | R&D Systems | 293-VE | Resuspend in PBS, store aliquots at -80 °C |
| Stereo Microscope, Leica MZ16 | Leica | ||
| Confocal laser scanning microscope LSM710 | Zeiss |
Referencias
- Lumsden, A., Keynes, R. Segmental patterns of neuronal development in the chick hindbrain. Nature. 337 (6206), 424-428 (1989).
- Chandrasekhar, A. Turning heads: development of vertebrate branchiomotor neurons. Dev. Dyn. 229 (1), 143-161 (2004).
- Mapp, O. M., Wanner, S. J., Rohrschneider, M. R., Prince, V. E. Prickle1b mediates interpretation of migratory cues during zebrafish facial branchiomotor neuron migration. Dev. Dyn. 239 (6), 1596-1608 (2010).
- Glasco, D. M., Sittaramane, V., Biol, D. e. v. .., et al. . The mouse Wnt/PCP protein Vangl2 is necessary for migration of facial branchiomotor neurons, and functions independently of Dishevelled. 369 (2), 211-222 (2012).
- Marillat, V., Sabatier, C., et al. The Slit Receptor Rig-1/Robo3 Controls Midline Crossing by Hindbrain Precerebellar Neurons and Axons. Neuron. 43 (1), 69-79 (2004).
- Dickson, B. J. Molecular mechanisms of axon guidance. Science. 298 (5600), 1959-1964 (2002).
- Vivancos, V., Chen, P., et al. Wnt activity guides facial branchiomotor neuron migration and involves the PCP pathway and JNK and ROCK kinases. Neural Dev. 4, 7 (2009).
- Guan, K. -. L., Rao, Y. Signalling mechanisms mediating neuronal responses to guidance cues. Nat. Rev. Neurosci. 4 (12), 941-956 (2003).
- Guthrie, S. Patterning and axon guidance of cranial motor neurons. Nat. Rev. 8 (11), 859-871 (2007).
- Garel, S., Garcia-Dominguez, M., Charnay, P. Control of the migratory pathway of facial branchiomotor neurones. Development. 127 (24), 5297-5307 (2000).
- Gavalas, A., Ruhrberg, C., Livet, J., Henderson, C. E., Krumlauf, R. Neuronal defects in the hindbrain of Hoxa1, Hoxb1 and Hoxb2 mutants reflect regulatory interactions among these Hox genes. Development. 130 (23), 5663-5679 (2003).
- Grant, P. K., Moens, C. B. The neuroepithelial basement membrane serves as a boundary and a substrate for neuron migration in the zebrafish hindbrain. Neural Dev. 5, 9 (2010).
- Lumsden, A. Segmentation and compartition in the early avian hindbrain. Mech. Dev. 121 (9), 1081-1088 (2004).
- Tsuchida, T., Ensini, M., et al. Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes. Cell. 79 (6), 957-970 (1994).
- Qu, Y., Glasco, D. M., et al. Atypical cadherins Celsr1-3 differentially regulate migration of facial branchiomotor neurons in mice. J. Neurosci. 30 (28), 9392-9401 (2010).
- Schwarz, Q., Gu, C., et al. Vascular endothelial growth factor controls neuronal migration and cooperates with Sema3A to pattern distinct compartments of the facial nerve. Genes Dev. 18 (22), 2822-2834 (2004).
- Stockinger, P., Maître, J. -. L., Heisenberg, C. -. P. Defective neuroepithelial cell cohesion affects tangential branchiomotor neuron migration in the zebrafish neural tube. Development. 138 (21), 4673-4683 (2011).
- Wanner, S. J., Prince, V. E. Axon tracts guide zebrafish facial branchiomotor neuron migration through the hindbrain. Development. , (2013).
- Higashijima, S., Hotta, Y., Okamoto, H. Visualization of cranial motor neurons in live transgenic zebrafish expressing green fluorescent protein under the control of the islet-1 promoter/enhancer. Neurosci. 20 (1), 206-218 (2000).
- Laird, P. W., Zijderveld, A., Linders, K., Rudnicki, M. A., Jaenisch, R., Berns, A. Simplified mammalian DNA isolation procedure. Nucleic Acids Res. 19 (15), 4293 (1991).
- Srinivas, S., Watanabe, T., et al. Cre reporter strains produced by targeted insertion of EYFP and ECFP into the ROSA26 locus. BMC Dev. Biol. 1 (1), 4 (2001).
- Nichols, A. J., O'Dell, R. S., Powrozek, T. A., Olson, E. C. Ex utero electroporation and whole hemisphere explants: a simple experimental method for studies of early cortical development. J. Vis. Exp. (74), (2013).
- Holland, P. J., George, A. M., Worrell, L. T. C., Landsberg, R. L. In vitro electroporation of the lower rhombic lip of midgestation mouse embryos. J. Vis. Exp. (66), (2012).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados