
Method Article
Visual condicionamiento clásico en madera hormigas
En este artículo
Erratum Notice
Resumen
Presentamos un protocolo para el condicionamiento de clásico de hormigas enjaezados que permite a los investigadores estudiar formación aprendizaje y la memoria visual a un nivel de análisis no es posible con el libre movimiento de personas.
Resumen
Varias especies de insectos se han convertido en sistemas modelo para estudiar la formación de la memoria y el aprendizaje. Aunque muchos estudios centran en mover libremente los animales, aplicación de paradigmas de condicionamiento clásico con insectos enjaezados los estudios han sido importantes para investigar las claves exactas que los individuos aprenden y los mecanismos neuronales subyacentes de aprendizaje y formación de la memoria. Aquí presentamos un protocolo para la evocación visual aprendizaje asociativo en madera hormigas a través del condicionamiento clásico. En este paradigma, las hormigas son aprovechadas y presentadas con una señal visual (una cartulina azul), el estímulo condicionado (CS), con una recompensa de azúcar apetitivo, el estímulo incondicional (Estados Unidos). Las hormigas realizan un reflejo de la extensión del maxilar Labium (MaLER), la respuesta incondicional (UR), que puede ser utilizada como una lectura para el aprendizaje. Entrenamiento consiste en 10 ensayos, separados por un intervalo de 5 minutos diferenciarlo (ITI). Las hormigas también se prueban para la retención de la memoria 10 minutos o 1 hora después del entrenamiento. Este protocolo tiene el potencial para permitir a los investigadores a analizar, en forma precisa y controlada, los detalles de la formación de la memoria visual y la base neural de la formación de memoria y el aprendizaje en hormigas de madera.
Introducción
Insectos se han utilizado como modelos para estudiar la memoria y el aprendizaje de formación1. Una veta especialmente exitosa de investigación implica el uso de condicionamiento clásico de animales restringidos, que permite un control preciso sobre las señales de ser aprendido y permite a los investigadores a investigar los mecanismos neurales que sustentan el aprendizaje y la memoria. La mayoría de los estudios se ha centrado en el condicionamiento de clásico apetitivo de trabajadores de la abeja, Apis mellifera. Los trabajadores de abeja están entrenados para asociar un CS con un Estados Unidos que evoca la Ronda Uruguay. En este paradigma, desarrollado originalmente por Takeda2 y Bitterman et al. 3, al UR es el reflejo de extensión probóscide (PER), los Estados Unidos son el azúcar y el CS es un olor. Las abejas aprenden la asociación entre el CS y los Estados Unidos y pueden formar una memoria a largo plazo de esta asociación.
El paradigma original utilizando el PER como la Ronda Uruguay se ha utilizado para desentrañar los caminos neuronales que sustentan el aprendizaje olfativo de las abejas4. Ha sido modificada en varias formas de aprendizaje de los diferentes estímulos sensoriales, incluyendo estímulos visuales5,6,7, incorporar aprendizaje aversivo mediante supresión de la por8 o la extensión de la picadura de prueba Reflejo (SER)9 como la Ronda Uruguay y para probar el aprendizaje en otras especies, como abejorros10 y moscas de la fruta11. Aunque con varias modalidades de formación de la memoria ha sido estudiada a través del condicionamiento clásico, visual aprendizaje es todavía difícil de observar desde este enfoque, incluso en especies muestran un alto grado de capacidad de aprendizaje visual durante la alimentación, tales como abejas.
Estudios recientes han aplicado un enfoque similar a los insectos que no tienen una probóscide, como langostas, que realizan la respuesta de apertura de Palp (POR)12y las hormigas, que realizan el reflejo de extensión maxilar Labium (MaLER)13. Esto ha revelado capacidades de aprendizaje específicos de fase que coincide con las estrategias de alimentación específicas de fase de los dos fenotipos diferentes langosta del desierto, el gregario y las formas solitarious, refuerzan la idea de que la formación de la memoria tiene que coincidir con necesidades ecológicas14. Además, estudios sobre el aprendizaje olfativo en hormigas ha demostrado semejanzas entre las hormigas y las abejas en la formación de la memoria y retención, con retención de memoria a largo plazo, 72 horas después del entrenamiento, siendo dependiente de proteína síntesis15.
Estas adaptaciones del paradigma original han permitido la formación de memoria y el aprendizaje a ser estudiado con muchas modalidades y en varias especies de modelo. Esto es esencial para la identificación de mecanismos de la memoria común de formación para los insectos, sino también para identificar las diferencias que reflejan la particular ecología de aprendizaje y la memoria en diferentes especies. El objetivo principal del protocolo que se describe aquí es proporcionar una manera para llevar a cabo experimentos de condicionamiento clásico un estímulo condicional visual con una especie de hormiga ampliamente estudiado, Formica rufa. Se emite desde el estudio del aprendizaje visual en hormigas de madera16, que también es una adaptación de los paradigmas de condicionamiento clásico visual.
Protocolo
1. mantenimiento de las colonias de Formica rufa
Nota: Las colonias de la hormiga de madera (Formica rufa, L.) utilizadas para este estudio fueron recogidas del bosque de Ashdown, Sussex, Reino Unido (N 51 4.680, E 0 1.800). En el Reino Unido, F. rufa colonias deben recogerse entre julio y agosto. Una buena parte de la jerarquía debe eliminarse, incluyendo varios cientos de trabajadores y cría, para la Colonia es sostenido y activo durante períodos más largos de tiempo (un año). Permiso debe obtenerse de las autoridades concernientes antes de retirarlo del nido.
- Casa de las colonias de la hormiga de madera en una caja amplia (unos 135 x 70 cm x 35 cm) con las paredes cubiertas con resina de politetrafluoroetileno (PTFE) (p. ej., Fluon) para evitar que las hormigas escapen, a 21 ° C, bajo 12 h luz: 12-h ciclo oscuro.
- Proporcionar agua y sacarosa (333 gr/L) a las hormigas, con dispensadores de agua. Para garantizar la que alimentación de las hormigas es rica en proteínas, alimentación de la Colonia con congelados grillos 2 x una semana. Rocíe el nido con agua todos los días para mantenerlo húmedo.
- Para mejorar las condiciones sanitarias de la Colonia, agregar resina de pino cuando es necesario; resina de pino reduce el parasitismo en las colonias de hormiga madera17. Coloque un recipiente pequeño (alrededor de 15 cm de diámetro y 5 cm de profundidad) en el nido hormigas pueden depositar a los miembros de la Colonia muerta en él.
- Mantener a las hormigas en las condiciones anteriores durante al menos 2 semanas después de recoger antes de intentar cualquier experimentos. Eliminar la sacarosa desde el nido a morir de hambre las hormigas 2 d antes de los experimentos, mejorando así su motivación.
2. selección y aprovechamiento de las hormigas
Nota: Para aprovechar las hormigas, es necesario un soporte a medida. Esto puede ser construido usando arcilla de modelaje y una cartulina con un corte de apertura conectado horizontalmente. La superficie superior de la cartulina debe ser parcialmente cubierta con la cera, permitiendo insectos pasadores para fijarse durante la fijación. Manejo cuidadoso es necesario para cada paso de este protocolo (incluyendo mantenimiento, transporte y experimento), pero particularmente al aprovechamiento de las hormigas, para no someter a altos niveles de estrés antes de la formación.
- Para seleccionar a las hormigas que están motivadas a comer, coloque un portaobjetos con una gota de sacarosa (200 g/L) dentro de una caja pequeña (unos 14 cm x 8 cm x 5 cm) con las paredes cubiertas con resina de PTF para evitar que se escapen. No seleccione las hormigas que llevan las hormigas muertas o madera del nido. Por el contrario, tomar las hormigas que están reptando por las paredes del nido, porque éstos son más propensos a ser forrajeros tratando de dejar el nido para buscar alimento.
- Coloque cada hormiga en la caja y esperar a ver si se alimenta de la caída de la sacarosa. Si lo hace, transferir inmediatamente a otra caja vacía para evitar la saciedad.
- Transfiera cada hormiga a un tubo separado. Coloque cada tubo en el congelador por 1-2 min, o en hielo picado hasta 5 min, para inmovilizar la hormiga.
- Lugar la hormiga inmovilizada en el soporte, a través del corte en la cartulina, por la articulación entre la cabeza y el tórax. Mantener la antena en el lado de la cabeza, con insectos pernos embebidos en la capa de cera sobre la cartulina.
- Utilice un elemento de calefacción a medida a la punta de un alfiler insectos a la cabeza de la hormiga, paralelo a la cartulina. Durante este procedimiento, no tocar las antenas con el elemento de calefacción del alambre para evitar incurrir en daños a ellos.
- Después la cera se seque, quite los pasadores de insectos con las antenas y retire con cuidado la hormiga del titular.
- Fijar el perno insectos manteniendo la hormiga en un modelado de barro cilindro y fijar un soporte plástico a medida debajo de él, asegurando la hormiga mantiene una postura de pie típico y que es libre de moverse todo el cuerpo excepción la cabeza.
- Mantener las hormigas enjaezadas en la oscuridad durante al menos 2 h antes de la formación.
3. formación y prueba
-
Configuración, el estímulo condicionado y el estímulo incondicionado
- Llevar a cabo los experimentos en una caja de acrílico blanca (50 x 50 x 50 cm), abierto en el frente para permitir el acceso para el experimentador. Registrar el comportamiento de la hormiga con una cámara.
- Coloque la cámara con un lente macro directamente encima de la hormiga, a través de un orificio en la superficie superior de la caja.
- Para reducir cualquier señales visuales extraños, mantenga la habitación en la oscuridad excepto una fuente de luz apuntando directamente a la parte superior de la caja de acrílico (dos bombillas de 26 W).
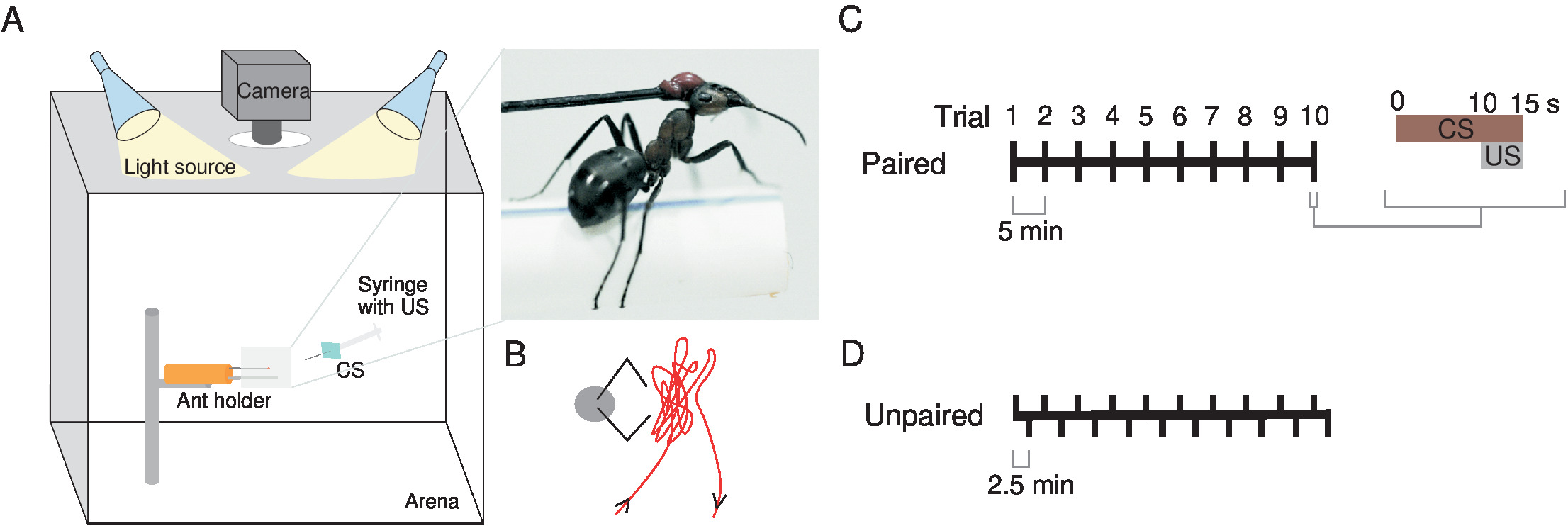
- Utilizar como una referencia visual (CS) un rectángulo de cartón de color azul brillante (espectro FiguraS1) (60 x 45 mm) unido en su centro a una aguja, conectada a una jeringa (2 mL) con el cual los Estados Unidos (sacarosa 200 g/L) manualmente se entrega a la hormiga (figura 1A y 1B ).
Nota: La solución de sacarosa se puede hacer con cualquier azúcar blanco, siempre que no tiene color y olor cuando está disuelto en agua. - Realizar dos tipos de formación en paralelo, emparejado y desapareado (véase la explicación abajo), en un orden aleatorio.
Nota: En este estudio, no prestamos atención a la naturaleza de la señal visual. Sólo la asociación entre la señal y la recompensa fueron tomada en cuenta. Color y forma no fueron probados, y es probable que un CS con otras características producirían resultados similares. Sin embargo, el color azul fue elegido porque las hormigas del género han demostrado ser sensibles a longitudes de onda que cubren el color azul18.
- Llevar a cabo los experimentos en una caja de acrílico blanca (50 x 50 x 50 cm), abierto en el frente para permitir el acceso para el experimentador. Registrar el comportamiento de la hormiga con una cámara.
-
Formación de pares
Nota: En la formación de pares, la CS y Estados Unidos se presentan en cada ensayo, asociados entre sí (figura 1).- Iniciar la grabación de la s hormigas 10 antes de presentar el CS para asegurarse de que las hormigas no espontáneamente realizan a MaLER antes de la presentación. Si las hormigas realizan a MaLER durante este período, posponer el juicio por unos segundos. Si alguna hormiga muestra este comportamiento continuamente, excluirla del análisis.
- Mueva la jeringa + CS frente a la hormiga para ~ 10 s, con la punta de la aguja se mantiene entre las hormigas y un máximo de 5 mm por encima y a cada lado. Durante este tiempo, mover la punta de la aguja lo más cerca posible a la cabeza de la hormiga pero sin tocar las antenas (figura 1B).
Nota: Movimiento fue incluido en la presentación del CS ya se ha demostrado para desempeñar un papel en el aprendizaje visual en las abejas6. - Aplique presión para exudar una sola gota de sacarosa desde la punta de la jeringa y coloque la gota de sacarosa junto a las antenas y partes bucales para permitir que la hormiga detectar la sacarosa. Permita que la hormiga a alimentar por cerca de 5 s.
Nota: La cantidad de hormigas de sacarosa consumida en cada ensayo no fue controlada en estos experimentos. La caída de la sucrosa en hormigas que pienso debe ser lo suficientemente grande como para que puedan alimentar libremente por cerca de 5 s. - Repita este procedimiento de 10 x, con un ITI de 5 minutos.
Nota: En estos experimentos, las hormigas siempre se gira a la derecha, con su ojo derecho cerca de la parte abierta de la caja de acrílico. Por lo tanto, el CS siempre se acercó a las hormigas de su lado derecho. Aunque esto no invalida el aprendizaje, estos experimentos pueden realizarse girando la mitad de las hormigas a la izquierda y la otra mitad a la derecha, para evitar cualquier efecto posible lateralización.
-
Formación impar
Nota: Este curso consiste en presentar a las hormigas el CS o los Estados Unidos por separado, estos dos estímulos que, por lo tanto, disociarse entre sí con el tiempo (figura 1).- Iniciar la grabación de la hormigas 10 s antes de cada ensayo.
- Presentamos el CS de la misma manera como en la formación de pares pero no entregan la sacarosa a la hormiga.
- Después de 2,5 min, entregar Estados Unidos directamente a las piezas bucales (también tocar las antenas) con una jeringa sin la CS adjuntadas.
- Comience con una prueba de CS y repite este procedimiento 10 x para cada tipo de ensayo, intercalando con un ITI de 2,5 min.
-
Prueba
Nota: Una prueba debe ser realizado solo 1 x cada 10 min o 1 h después del último juicio de formación; una prueba en la que la hormiga se presenta con el CS pero no con los Estados Unidos podría causar la extinción de la memoria asociativa formada anteriormente.- Iniciar la grabación de la s hormigas 10 antes de la prueba.
- Presentamos el CS a la hormiga para cerca de 10 s.
- Asegúrese de que la hormiga está siendo motivada para alimentación mediante la entrega de sacarosa después de la prueba.
4. Análisis y recopilación de datos
- Registrar el comportamiento de la hormiga, desde arriba, 10 s antes de cada ensayo y durante las presentaciones de CS y Estados Unidos. Asegúrese de que todas las pruebas y ensayos se registran para su posterior análisis.
- Utilizando los registros realizados durante el entrenamiento y prueba, anotar las respuestas de las hormigas durante los 10 s de presentación de CS.
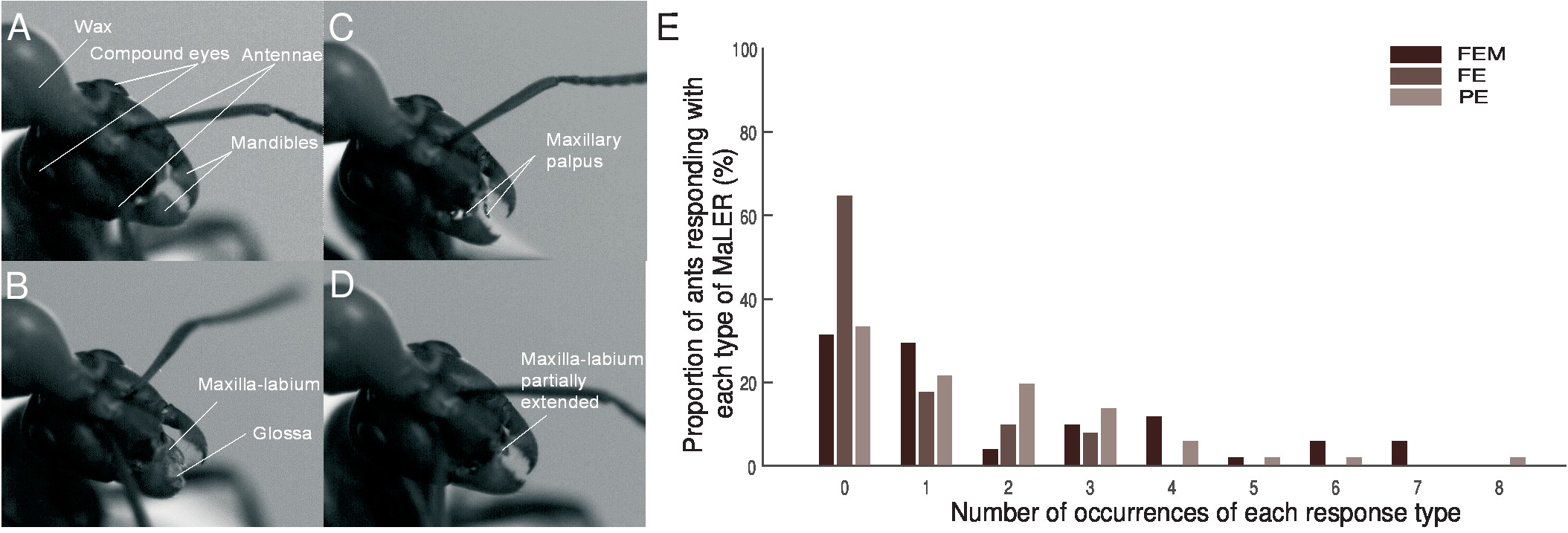
- Separar las respuestas de las hormigas durante la presentación de CS en tres tipos de comportamiento, dependiendo de la extensión y el movimiento de las piezas bucales: extensión completa con movimiento (FEM) como si alimentación, extensión completa sin movimiento (FE) o parcial (PE) de extensión de la palpos de labia de maxilar superior o maxilar (figura 2A - 2D). Para el análisis, el grupo todos los tipos de MaLER como respuesta única (Figura 2E).
- Excluir cualquier hormiga que no alimentan en cada prueba y prueba.
Nota: Para el análisis, se recomienda una prueba estadística que las hormigas individuales se tiene en cuenta, lo representa la variabilidad dentro de individuos. Al clasificar las respuestas de las hormigas durante el entrenamiento de forma binaria (1 para una respuesta positiva y 0 para ninguna respuesta), regresión logística con efectos mixtos es recomendable16,19. Para comparar la proporción de hormigas responder en cada ensayo de entrenamiento o prueba, un G-test o prueba exacta de Fisher se recomienda, dependiendo del número de hormigas analizados16,20.
Resultados
Durante los experimentos de condicionamiento clásico, el CS no debe inducir una respuesta espontánea de los animales. En los experimentos que hemos realizado, sólo el 3% - 4% de las hormigas realizan a MaLER en respuesta a la señal visual en el primer ensayo, antes de la formación. Hormigas que fueron sometidos a un entrenamiento apareado realizan MaLER cada vez más en respuesta a la CS (Figura 3A, regresión logística, N = 51, grados de libertad (df) = 507, z = 5.949, p < 0.01). El porcentaje de hormigas apareadas que responden a la CS plateaued alrededor 50%, a partir del tercer ensayo. Por el contrario, las hormigas no mostró significativas no aumentan en MaLER durante el entrenamiento (figura 3B, regresión logística, N = 29, df = 287, z = 0.758, p = 0,45). La ocurrencia de MaLER en respuesta a la señal visual fue significativamente mayor durante el emparejado de desapareado formación (regresión logística, N = 80, df = 796, z =-5.306, p < 0.01), que era cierto para cada prueba, excepto la primera (tabla 1).
Para examinar su corto y una memoria intermedia15, las hormigas se probaron 10 min o 1 h después del último juicio de formación. Para ambas pruebas, la proporción de hormigas realizar a MaLER en respuesta a la CS fue significativamente mayor al que habían experimentado el entrenamiento apareado en lugar impar (figura 3 y 3D; Tabla 1).
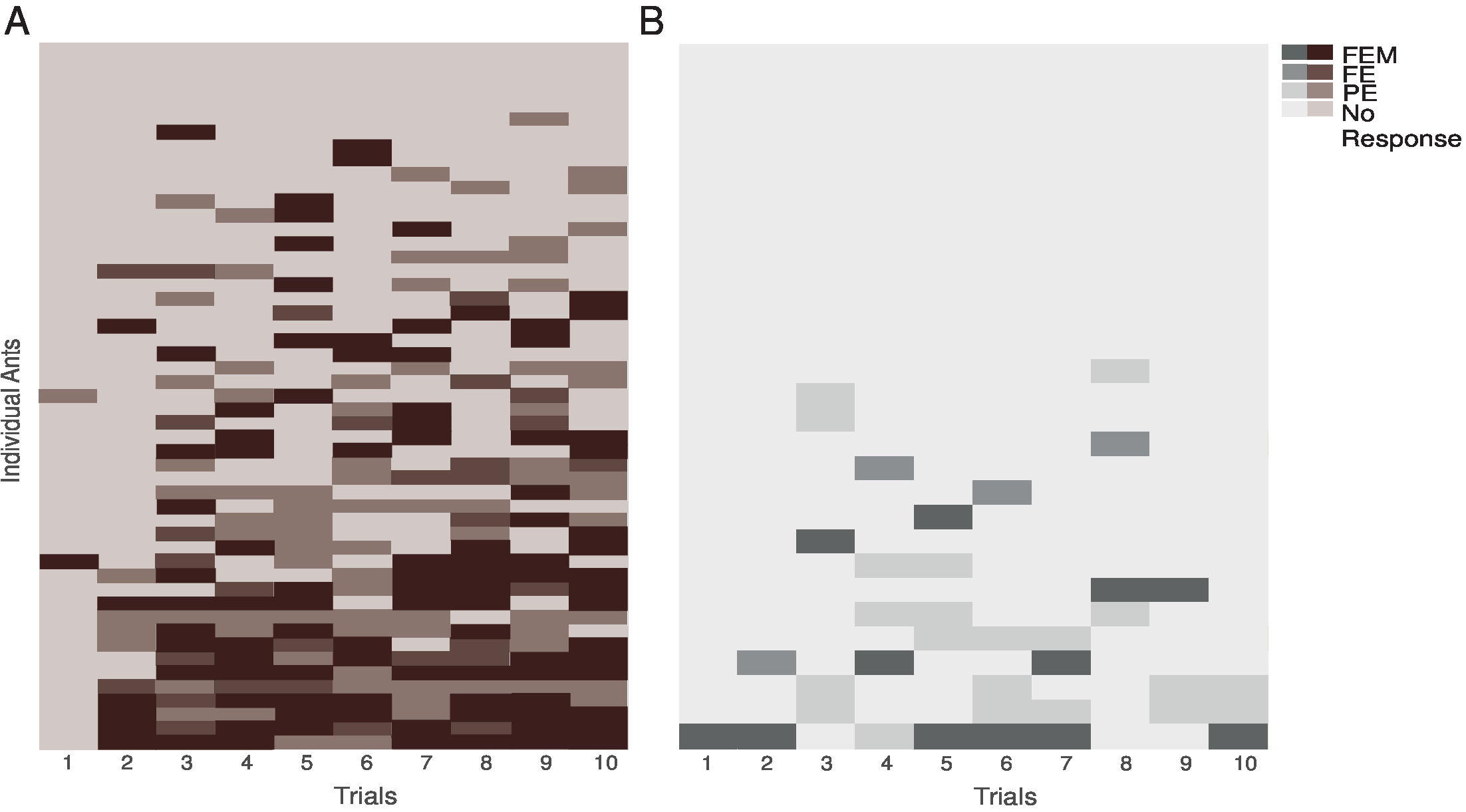
Durante las hormigas individuales, formación demostraron una variación sustancial en el número y tipo de MaLER, que muestran (Figura 4A y 4B). Sólo el 14% de hormigas pareadas respondió en cada ensayo de la segunda o tercera prueba adelante, mientras más hormigas se alternaron entre los ensayos en los que respondieron y en el que no lo hicieron. En los ensayos de entrenamiento en el que las hormigas respondieron, varía el grado en que extendió y movió sus piezas bucales. Por lo tanto, dividimos MaLER en tres tipos: FEM, FE o PE. Normalmente, las hormigas realizan FEM o PE más que FE. Sin embargo, sólo unas pocas hormigas constantemente realizan el mismo tipo de respuesta; en la mayoría de los casos, las hormigas demostraron poca consistencia en el tipo de MaLER que se presentaron (figura 4).

Figura 1 : Montaje experimental y esquema de formación. (A) fijar la hormiga con cera a un pin del insecto al modelado de un cilindro de arcilla (naranja) y el lugar de un soporte a medida por debajo de ella para permitir una postura naturalista. Coloque dentro de una caja de acrílico blanco iluminada por dos fuentes de luz, directamente debajo de una cámara. Uso como el estímulo condicionado (CS) de un cartón (cuadrado azul) unido a la jeringa que ofrece el estímulo de unconditioned (los E.E.U.U.), una recompensa de azúcar. El recuadro muestra una vista cercana de una hormiga en el soporte. (B) la punta de la jeringa de la aguja se mueve más cerca posible a la cabeza de la hormiga pero sin tocar las antenas como se muestra en este esquema. Emparejado de hormigas de tren a través de cada una (C) o (D) desapareado formación. Esta figura ha sido modificada desde Fernandes et al. 16. haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2 : La respuesta de la extensión del maxilar Labium de hormigas de madera. Fotogramas individuales de grabaciones de vídeo muestran movimientos de parte de boca de las hormigas durante el entrenamiento. (A) este panel no muestra ninguna respuesta. (B) este panel muestra una completa extensión (FE) de la labia del maxilar superior que termina en la glossa. (C) este panel muestra una extensión parcial (PE) con sólo el maxilar palpus visible. (D) este panel muestra una extensión parcial (PE) de las estructuras del maxilar labium. (E) hormigas en formación emparejado (N = 51) realizar una extensión completa con movimiento (FEM, marrón oscuro), una FE (marrón intermedio) o PE (marrón claro). Esta figura ha sido modificada desde Fernandes et al. 16. haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3 : Forma de hormigas memorias asociativas de un cue visual junto con una recompensa de azúcar de madera. (A) el porcentaje de hormigas emparejados (N = 51) realizar MaLER en respuesta a la presentación de CS significativamente incrementado a lo largo de la formación. (B) el porcentaje de hormigas realizar MaLER no aumentó significativamente a lo largo de la formación impar (N = 29). Hormigas fueron probados (C) 10 minutos (emparejados: N = 15; desapareado: N = 15) o (D) 1 hora (emparejados: N = 15; desapareado: N = 14) después del último juicio de formación. El porcentaje de respuesta a la CS durante y después del entrenamiento de pares o impar de hormigas están representadas en marrón o gris, respectivamente. Están representados los tres tipos de MaLER en oscuridad (FEM), media (FE) y tonos de luz (PE). Esta figura ha sido modificada desde Fernandes et al. 16. haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4 : Rendimiento individual de las hormigas durante el entrenamiento. Estos paneles muestran las actuaciones individuales de (A) las hormigas emparejadas y (B) desapareado hormigas. Están representados los tres tipos de MaLER en oscuridad (FEM), medio (FE) y marrón claro (PE) o gris. Esta figura ha sido modificada desde Fernandes et al. 16. haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
| Ensayo | N | DF | G (ajustado) | P |
| 1 | 80 | na | na | > 0.1 |
| 2 | 80 | 1 | 3,86 | < 0.05 |
| 3 | 80 | 1 | 8.41 | < 0,01 |
| 4 | 80 | 1 | 6.63 | < 0,01 |
| 5 | 80 | 1 | 8.41 | < 0,01 |
| 6 | 80 | 1 | 7.5 | < 0,01 |
| 7 | 80 | 1 | 10.69 | < 0,01 |
| 8 | 80 | 1 | 11,76 | < 0,01 |
| 9 | 80 | 1 | 17.13 | < 0,01 |
| 10 | 80 | 1 | 17.13 | < 0,01 |
| 10 min | 59 | 1 | 5.5 | < 0.05 |
| 1 h | 59 | 1 | 4.42 | < 0.05 |
Tabla 1: comparación de las respuestas de MaLER al CS entre las hormigas que había experimentado emparejado y desapareado entrenamiento, para cada ensayo y prueba El número de hormigas (N) grados de libertad (df), G-prueba de independencia (G) y p-valor se muestran. El primer ensayo se analizó con prueba exacta de Fisher. Esta tabla ha sido modificada de Fernandes et al. 16.
Discusión
Condicionamiento clásico es uno de los paradigmas más bien establecidos para estudiar el aprendizaje y la memoria. El protocolo que presentamos aquí es una adaptación del paradigma diseñado para miel abeja trabajadores2,3 y posteriormente utilizado con otras especies, tales como abejorros, moscas de la fruta, que también por una lectura para el aprendizaje de10 ,11y langostas y hormigas, que utilizan POR y MaLER, respectivamente12,13. Mediante este protocolo, es posible capacitar enjaezadas hormigas de madera para aprender la asociación entre una clave visual y una recompensa de azúcar y analizar la retención de este corto (10 minutos) y medio plazo (1 hora) a memoria16.
En cualquier experimento conductual, es necesario tener en cuenta pasos críticos que pueden reducir al mínimo la variabilidad en las respuestas de los animales. En el protocolo presentado aquí, se toman varias medidas para minimizar la variabilidad antes y durante el entrenamiento. Antes del comienzo de los experimentos, la Colonia tiene que ser muerto de hambre durante al menos dos días y hormigas deben seleccionarse basado en su disposición a comer de una gota de azúcar en la caja de la explotación. Seleccionar a las hormigas de esta manera se pretende maximizar las posibilidades de las hormigas de la formación que están motivados para alimentar a. Cuidado de manejo es también una consideración importante porque puede ayudar a reducir los niveles de estrés, que altera aprendizaje si es demasiado intenso21. Con este fin, hormigas deben ser anestesiadas con frío para permanecer inmóvil mientras se aprovecha, porque cualquier movimiento (para escapar) durante este procedimiento puede ser una fuente de estrés. Además, el contacto entre la hormiga y la cera debe ser mínimo, evitando el contacto entre las antenas y la cera caliente o alambre, que podría causar daños. Aunque estas observaciones no han sido analizadas formalmente, las antenas parecen moverse con un patrón específico durante el aprendizaje.
Durante los experimentos, cuidado entrega del azúcar también es importante para mantener las hormigas motivado. Otra vez, mientras esto no se ha analizado formalmente, la entrega de alimentos abrupta parece causar estrés adicional a la hormiga, que a su vez condujo a una falta de motivación y aprendizaje. Además, la sacarosa entregada durante el entrenamiento debe ser de una disminución de la concentración (200 g/L) para evitar la saciedad antes de finalizar el entrenamiento y pruebas. Esto permite MaLER a ser una respuesta incondicional del buen candidato porque, junto con una baja performance espontánea de la respuesta a la señal visual, también no saturar sobre ensayos. Por último, contrariamente a la mayoría condicionamiento clásico estudios2,3,5,6,7,8,9,10, 11 , 12 , 13, entrenamos una hormiga en el tiempo hasta el final del experimento, dejando en su lugar entre los ensayos en lugar de quitarlo para probar otra hormiga. Formación varias hormigas juntos parecía producir resultados más variables, que pueden ser debido a un aumento de tensión o conflicto entre información visual causada por el cambio completo del paisaje. Para reducir la duración de cada experimento, utilizamos un ITI de 5 minutos en vez del ITI de 10 minutos en la mayoría de los estudios de condicionamiento clásico16. Aunque todas estas consideraciones deben ayudar a garantizan que las hormigas están motivadas para alimentarse y aprender durante el entrenamiento, no puede evitarse cierta variabilidad. Se recomienda utilizar las hormigas que parecen tener un comportamiento social, apetitivo y locomoción normal y excluyendo las hormigas del análisis el momento no se alimentan de un ensayo de formación o una prueba.
La naturaleza de la CS no fue probada en este estudio. Aunque hemos utilizado un estímulo visual azul porque las hormigas del mismo género son sensibles a estas longitudes de onda de18, otros colores también pueden aprenderse en asociación con una recompensa. Se necesitarían otros experimentos para caracterizar completamente los colores siendo visto y aprendido en este montaje. Esto también es cierto para diferentes formas y tamaños de la pista visual. No hemos probado si sería suficiente para distinguir el estímulo visual presentado aquí a la distancia de los ojos de las hormigas se presentó en resolución espacial de las hormigas. Aunque los ojos compuestos de las hormigas de madera han sido descritos en términos de tamaño y número de facetas22, a nuestro conocimiento, su resolución espacial no se ha completamente descrito todavía. Sin embargo, esto ha sido calculado para Melophorus magoti23. Una caracterización similar de las hormigas madera, o los ojos de otros insectos probados contribuiría a una investigación clara de las características de la localización visual observado y aprendido por los animales. Además, se incluyeron movimiento al presentar el estímulo visual a la hormiga ya se ha demostrado para desempeñar un papel en el aprendizaje de la abeja durante el condicionamiento clásico6. Sin embargo, esto también no se comprobó en este estudio y, debido a la naturaleza de diferentes movimiento de insectos en comparación con pocos insectos voladores, las diferencias entre el condicionamiento clásico de visual de la abeja y la hormiga podrían ser observadas.
Por último, hemos podido examinar retención de memoria a largo plazo porque las hormigas no sobrevivió siendo aprovechado por tantas largos períodos después del entrenamiento. Sin embargo, en posteriores juegos de experimentos, hemos mantenido las hormigas vivo y motivado para comer y aprender a aprovechar y ellos toda la noche en un ambiente oscuro y húmedo (colocando una caja sobre ellos). Por lo tanto, este paradigma podría utilizarse para desentrañar la retención de memoria a largo plazo de las hormigas de madera, además de memoria a corto y medio plazo.
Con este simple procedimiento adaptado de paradigmas de condicionamiento clásico general, es posible estudiar la adquisición y retención de memoria visual en hormigas madera enjaezadas, que han sido estudiadas ampliamente en paradigmas con animales free-mover. Este paradigma tiene el potencial para ser utilizado para el análisis de la base neural del aprendizaje visual en un modelo muy bien establecido para la navegación de insectos.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Los autores agradecen a Tom Collett y Cornelia Buehlmann por compartir información valiosa con respecto a la colección y mantenimiento de las colonias de hormigas de madera. Los autores también agradecen a Justine Crevel para comentar sobre las versiones anteriores de este artículo.
Materiales
| Name | Company | Catalog Number | Comments |
| Fluon | Blades Biological Ltd, Edenbridge, UK | ACS 109; ACS 112; ACS 114 | For preventing insects from scaping |
| Crickets | Blades Biological Ltd, Edenbridge, UK | LZJ 217 | Given to the ant colonies as protein source |
| Natural Pine Rosin/Resin | Minerals-water Ltd, Rainham, UK | 500g | Given to the ant colonies for sanitation |
| Austerlitz Insect Pin | Fine Science Tools GmbH, Heidelberg, Germany | 26000-40 | For harnessing ants |
| High speed camera | Edmund Optics Inc., Barrington, USA | eo-13122M | MaLER recordings during training and testing |
| Macrolens | Cannon, Surrey, UK | EF 100 mm f/2.8 L Macro IS USM | MaLER recordings during training and testing |
| Software | IDS Imaging Development Systems GmbH | uEye64 | MaLER recordings during training and testing |
| Blue Cardboard | john smith's at Union Store, University of Sussex | JACK-PJM41358 | Constitutes de conditional stimulus |
| Syringe | Fisher Scientific LTD, Loughborough, UK | BD Plastipak 300185case; Product Code.12369289 | US and CS (attached) presentation |
| Needle (0.5 x 16 mm) | Fisher Scientific LTD, Loughborough, UK | BD Microlance 300600; Product Code:10442204 | US and CS (attached) presentation |
Referencias
- Chittka, L., Niven, J. Are bigger brains better?. Current Biology. 19 (21), R995-R1008 (2009).
- Takeda, K. Classical conditioned response in the honey bee. Journal of Insect Physiology. 6, 168-179 (1961).
- Bitterman, M. E., Menzel, R., Fietz, A., Schäfer, S. Classical conditioning of proboscis extension in honeybees (Apis mellifera). Journal of Comparative Psychology. 97 (2), 107-119 (1983).
- Menzel, R., Muller, U. Learning and memory in honeybees: from behavior to neural substrates. Annual Review of Neuroscience. 19 (1), 379-404 (1996).
- Hori, S., Takeuchi, H., Kubo, T. Associative learning and discrimination of motion cues in the harnessed honeybee Apis mellifera L. Journal of Comparative Physiology A. 193 (8), 825-833 (2007).
- Balamurali, G. S., Somanathan, H., De Ibarra, N. H. Motion cues improve the performance of harnessed bees in a colour learning task. Journal of Comparative Physiology A. 201 (5), 505-511 (2015).
- Mujagić, S., Würth, S. M., Hellbach, S., Dürr, V. Tactile conditioning and movement analysis of antennal sampling strategies in honey bees (Apis mellifera L). Journal of Visualized Experiments. (70), e50179 (2012).
- Wright, G., et al. Parallel reinforcement pathways for conditioned food aversions in the honeybee. Current Biology. 20 (24), 2234-2240 (2010).
- Vergoz, V., Roussel, E., Sandoz, J. C., Giurfa, M. Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex. PloS One. 2 (3), e288 (2007).
- Laloi, D., et al. Olfactory conditioning of the proboscis extension in bumble bees. Entomologia Experimentalis et Applicata. 90 (2), 123-129 (1999).
- Chabaud, M. A., Devaud, J. M., Pham-Delègue, M. H., Preat, T., Kaiser, L. Olfactory conditioning of proboscis activity in Drosophila melanogaster. Journal of Comparative Physiology A. 192 (12), 1335-1348 (2006).
- Simões, P., Ott, S. R., Niven, J. E. Associative olfactory learning in the desert locust, Schistocerca gregaria. Journal of Experimental Biology. 214 (15), 2495-2503 (2011).
- Guerrieri, F. J., d'Ettorre, P. Associative learning in ants: conditioning of the maxilla-labium extension response in Camponotus aethiops. Journal of Insect Physiology. 56 (1), 88-92 (2010).
- Simões, P. M., Niven, J. E., Ott, S. R. Phenotypic transformation affects associative learning in the desert locust. Current Biology. 23 (23), 2407-2412 (2013).
- Guerrieri, F. J., d'Ettorre, P., Devaud, J. M., Giurfa, M. Long-term olfactory memories are stabilised via protein synthesis in Camponotus fellah ants. Journal of Experimental Biology. 214 (19), 3300-3304 (2011).
- Fernandes, A. S. D., Buckley, C. L., Niven, J. E. Visual associative learning in wood ants. Journal of Experimental Biology. 221 (3), 173260 (2018).
- Christe, P., Oppliger, A., Bancala, F., Castella, G., Chapuisat, M. Evidence for collective medication in ants. Ecology Letters. 6 (1), 19-22 (2003).
- Aksoy, V., Camlitepe, Y. Behavioural analysis of chromatic and achromatic vision in the ant Formica cunicularia (Hymenoptera: Formicidae). Vision research. 67, 28-36 (2012).
- Bates, D. M. . lme4: Mixed-Effects Modeling with R. , (2010).
- Sokal, R. R., Rohlf, F. J. . Biometry (3rd edn). , (1995).
- Bateson, M., Desire, S., Gartside, S. E., Wright, G. A. Agitated honeybees exhibit pessimistic cognitive biases. Current Biology. 21 (12), 1070-1073 (2011).
- Perl, C. D., Niven, J. E. Differential scaling within an insect compound eye. Biology letters. 12 (3), 20160042 (2016).
- Schwarz, S., Narendra, A., Zeil, J. The properties of the visual system in the Australian desert ant Melophorus bagoti. Arthropod Structure & Development. 40 (2), 128-134 (2011).
Erratum
Formal Correction: Erratum: Visual Classical Conditioning in Wood Ants
Posted by JoVE Editors on 1/01/1970. Citeable Link.
An erratum was issued for: Visual Classical Conditioning in Wood Ants. The Acknowledgments section was updated, and a supplemental figure was added.
The Acknowledgments sections was updated from:
The authors thank Tom Collett and Cornelia Buehlmann for sharing valuable information regarding collection and maintenance of wood ant colonies. The authors also thank Justine Crevel for commenting on previous versions of this article.
to:
The authors thank Tom Collett and Cornelia Buehlmann for sharing information regarding collection and maintenance of wood ant colonies. The authors also thank Justine Crevel for commenting on previous versions of this article, and Nora Nevala for measuring the spectrum intensity of the visual stimulus. This work was supported by a BBSRC grant to JEN (grant number BB/R005036/1). All the data pertaining to this manuscript are published in the University of Sussex Research Data Repository online database (10.25377/sussex.5794386).
The following supplemental figure was added to the end of the Representative Results section:
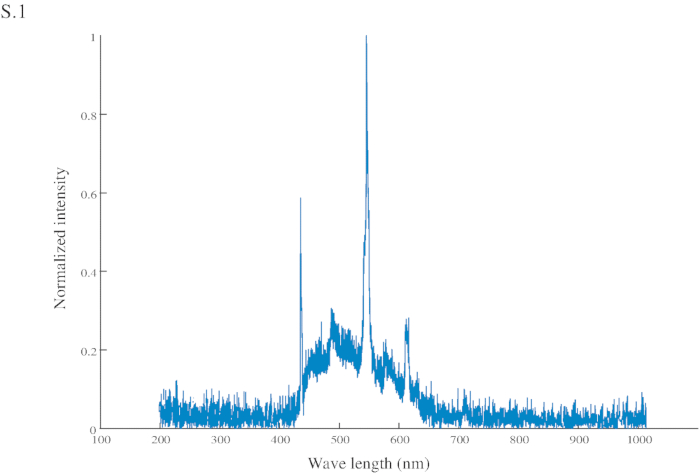
Figure S1: Normalized intensity of the conditional stimuli (CS). The CS has a peak intensity at 545 nm (in the green range) and another at 435 nm (in the blue range).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados