
Method Article
Flujo de trabajo de tomografía de matriz para la adquisición dirigida de información de volumen mediante microscopía electrónica de barrido
En este artículo
Resumen
Describimos la preparación de cintas de secciones seriadas y su recolección en soporte de transferencia grande para su uso como muestras de tomografía de matriz, junto con procedimientos de imágenes automatizados en un microscopio electrónico de barrido. El protocolo permite la detección, la recuperación y la obtención de imágenes dirigidas de eventos locales y raros, y la adquisición de grandes volúmenes de datos.
Resumen
La microscopía electrónica se aplica en biología y medicina para obtener imágenes de detalles celulares y estructurales a resolución nanométrica. Históricamente, la microscopía electrónica de transmisión (TEM) proporcionó información sobre la ultraestructura celular, pero en la última década, el desarrollo de los microscopios electrónicos de barrido (SEM) modernos ha cambiado la forma de mirar dentro de las células. A pesar de que la resolución de TEM es superior cuando se necesitan detalles estructurales a nivel de proteína, la resolución SEM es suficiente para la mayoría de las preguntas relacionadas con la biología celular a nivel de orgánulos. El avance en la tecnología permitió soluciones automáticas de adquisición de volumen, como la imagen de cara de bloque serie (SBF-SEM) y el haz de iones enfocado SEM (FIB-SEM). Sin embargo, hasta el día de hoy, estos métodos siguen siendo ineficientes cuando la identificación y navegación a áreas de interés son cruciales. Sin los medios para la localización precisa de las áreas objetivo antes de la obtención de imágenes, los operadores necesitan adquirir muchos más datos de los que necesitan (en SBF-SEM) o, lo que es peor, preparar muchas cuadrículas e imaginarlas todas (en TEM). Proponemos la estrategia de "cribado lateral" mediante Tomografía Array en SEM, que facilita la localización de áreas de interés, seguida de imágenes automatizadas de la fracción relevante del volumen total de la muestra. Las muestras de tomografía de matriz se conservan durante la obtención de imágenes y se pueden organizar en bibliotecas de secciones listas para imágenes repetidas. Se muestran varios ejemplos en los que el cribado lateral nos permite analizar detalles estructurales a los que es increíblemente difícil acceder con cualquier otro método.
Introducción
A pesar de la importancia de las técnicas relacionadas con los EM, el esfuerzo requerido para dominarlas mantiene todo el campo restringido a un pequeño número de especialistas. Una dificultad significativa es la identificación y recuperación de una Región de Interés (ROI) en las muestras conservadas para EM. La apariencia de la misma muestra difiere considerablemente cuando se analiza por microscopía óptica y después del procesamiento para la observación EM. Los cambios para las muestras preparadas químicamente incluyen la contracción de la muestra anisotrópica después de los pasos de deshidratación (~ 10% en cada dimensión) y la pérdida de fluorescencia cuando se usa osmio en el protocolo de fijación y tinción(Figura 1A). Para la seccionamiento ultrafino, las muestras se incrustan en resinas epoxi o acrílicas utilizando diferentes estrategias(Figura 1B). Para obtener resultados exitosos de esta preparación, toda la muestra debe fraccionarse en piezas que no excedan de 1 mm x 1 mm. Para cumplir con los requisitos estándar de observación de las condiciones de observación de la Microscopía Electrónica de Transmisión (TEM), esta pequeña porción de la muestra se secciona aún más a rodajas de 50-150 nm de espesor. Las imágenes en escala de grises resultantes muestran la organización del tejido y la estructura del orgánulo de una fracción diminuta de toda la muestra con mayor detalle que cualquier otra técnica de microscopía(Figura 1C). Un conjunto de datos TEM típico proporciona información 2D, teóricamente extrapolada para comprender los procesos que ocurren naturalmente en un espacio 3D en células y tejidos. La Figura 1D presenta el desafío de la adquisición de volúmenes ultraestructurales: si se secciona un cubo de lado de 1.000 μm a 50 nm de espesor, se requerirán 20.000 secciones para cubrir todo el volumen; para un cubo lateral de 500 μm, será de 10.000 secciones. Para cubrir un volumen de 50 μm x 50 μm x 50 μm, podrían ser necesarias 1.000 secciones "solamente". Obtener este volumen manualmente es prácticamente imposible y extremadamente difícil de realizar con automatización. Si, además de la profundidad de la muestra, necesitamos cubrir toda la superficie de tales cubos hipotéticos, la cobertura de 1 μm2 de superficie a una resolución razonable se convierte en un problema logístico grave(Figura 1E). Mientras que para los extraordinarios proyectos a gran escala, como los enfoques de conectómica, el gran número de secciones es crucial, para la mayoría de los proyectos EM "mundanos", generar más secciones requeridas para la observación presenta una desventaja significativa.
Existen varios métodos para adquirir información ultraestructural 3D: microscopía electrónica de transmisión de seccionamiento en serie (TEM), tomografía TEM, tomografía de matriz (AT), microscopía electrónica de barrido de imágenes faciales en bloque serie (SBF-SEM) y microscopía electrónica de barrido de haz iónico enfocado (FIB-SEM). Las principales diferencias entre estos métodos son la estrategia de seccionamiento y si la adquisición de imágenes está acoplada a la sección de generación1. En el seccionamiento en serie TEM, las secciones secuenciales se recogen en cuadrículas de ranuras, las imágenes TEM se generan a partir de estas secuencias y se alinean2,3,4,5. En la tomografía TEM, las series de inclinación de secciones de 150-300 nm en una cuadrícula, y cuando se acoplan a la seccionamiento en serie, proporcionan una resolución muy alta, aunque relativamente pequeñavolúmenes 6,7,8. El enfoque AT utiliza secciones físicas con diversas formas manuales y semiautomáticas de recolección de secciones en soportes relativamente grandes, como cubiertas de vidrio, obleas de silicio o una cinta especial. Para la adquisición de imágenes, el soporte se analiza en SEM, con diversas estrategias de adquisición de imágenes están disponibles9,10,11,12,13,14,15. Para SBF-SEM, la seccionamiento físico se logra utilizando un mini-microtomo con un cuchillo de diamante engastado directamente dentro de la cámara SEM, con la imagen SEM generada a partir de la superficie del bloque de resina16,17,18,19. Para FIB-SEM, la fuente iónica elimina las capas delgadas de la muestra, seguida de imágenes automáticas de la superficie expuesta por el SEM20,21. La tomografía TEM y el AT generan secciones físicas, que se pueden volver a visualizar si es necesario, mientras que FSBF-SEM y FIB-SEM eliminan la sección después de la imagen. Una combinación reciente de secciones físicas fotografiadas por un SEM multihaz proporciona una combinación de métodos que resuelve el problema del "cuello de botella" de la velocidad de adquisición de imágenes22. Cada una de estas técnicas ha revolucionado la forma en que se pueden obtener y analizar los datos em, y cada enfoque tiene sus impactos prácticos relacionados con una pregunta de investigación determinada.
Dada la naturaleza de la preparación y la escala de las dimensiones ultraestructurales, no es sencillo predecir dónde se encuentra una estructura objetivo específica en el bloque de muestra (Figura 1D,E). Una solución para la localización del ROI es grabar las imágenes de todo el bloque a la resolución deseada desde el principio. Las estructuras de interés pueden estar en el volumen de datos adquirido cuando se está lejos del microscopio. El tiempo de adquisición y el manejo de datos asociados con esta estrategia son problemáticos. Es deseable reducir la cantidad de datos registrados, especialmente si los ROI son mucho más pequeños que el bloque tisular, es decir, si los objetos de interés son tipos específicos de células (no órganos completos). Diferentes técnicas correlativas de luz y microscopía electrónica (CLEM) pueden tener éxito cuando la fluorescencia se conserva y localiza antes o después de la preparación dentro de la misma muestra23,24 ,25,26,27,28,29. Sin embargo, muchas estructuras celulares son reconocibles incluso sin correlación de fluorescencia, solo basada en la ultraestructura conocida. Para estos casos, creemos que la tomografía de matriz de cribado lateral proporciona una compensación de equilibrio entre el esfuerzo invertido en la localización del ROI y la calidad de la información ultraestructural. Usando esta estrategia, un subconjunto de secciones en la oblea se examinan dentro de intervalos regulares, que se pueden establecer en función del tamaño y la naturaleza del ROI. Una vez que se encuentran los ROI, la adquisición de datos se configura en una serie continua de secciones que comienzan antes y terminan después de la sección de anclaje, recopilando la información relevante de manera específica.
Presentamos protocolos para AT que simplifican y aceleran la adquisición de regiones o eventos de interés en numerosas secciones y producen volúmenes de imágenes mejor alineados. El cribado lateral y la adquisición multipaso producen datos con muy alta resolución en regiones específicas con precisión. El procedimiento que describimos aborda varios desafíos de la adquisición de datos 3D EM, ya que prevé: compatibilidad con una amplia gama de especímenes sin cambiar fundamentalmente el flujo de trabajo de preparación de muestras; localización específica para seccionamiento y adquisiciones de SEM; reducción del tiempo y el esfuerzo durante la configuración; imágenes de regiones en múltiples secciones con mejor alineación de los volúmenes resultantes; y un procedimiento suave de costura y alineación para compilar diferentes imágenes en una imagen de mosaico cosida. Elegimos demostrar la fortaleza de nuestro método con varias muestras de proyectos publicados y en curso. Creemos que este enfoque puede facilitar significativamente la generación y adquisición de datos EM específicos, incluso para investigadores con experiencia limitada en EM.
Protocolo
NOTA: Los métodos de preparación de muestras se describen en otra parte14,30,31y no se tratan en esta publicación. En resumen, los ejemplos mostrados se fijaron químicamente con glutaraldehído, se posfijaron con 1% de OsO4,luego se trataron con acetato de uranilo acuoso al 1% antes de incrustarlo en la resina incrustada. Alternativamente, las muestras se pueden preparar utilizando congelación a alta presión, se pueden congelar con acetato de uranilo al 0,1% en acetona e incrustarse en resina acrílica. Los bloques de muestra se prepararon utilizando un método de incrustación plana que permite una visión clara de la muestra, facilitando su orientación para la sección (Figura 1B).
1. Procedimiento de generación de arreglos de discos
- Orientación y recorte de muestras integrados
- Usando el binocular / microscopio, identifique y marque el ROI en la superficie del bloque incrustado rascando ligeramente la superficie del bloque con una cuchilla de afeitar. Esto ayudará a orientar la muestra dentro del ultramicrotomo y reducirá la superficie de seccionamiento.
- Sujete la muestra en el soporte del ultramicrotomo (Figura 2A).
- Recorte la resina alrededor de la muestra, primero con la hoja de afeitar (recorte en bruto) y continúe con la herramienta de recorte de diamante (recorte fino; Figura 2A). Use cuchillos con una inclinación del borde de 20 ° o 90 ° para asegurarse de que las superficies superior e inferior del bloque sean paralelas al filo de corte del cuchillo.
NOTA: Este paso es fundamental para la seccionamiento en serie y la adquisición de cintas rectas. - Mezcle xileno y pegamento en proporción 3:1. Con una pestaña unida a un palillo de dientes, aplique esta mezcla en los bordes superior e inferior del bloque recortado. Deja que se seque bien.
- Prepare el soporte de montaje de la matriz A (es decir, obleas de silicio).
- Corte la pieza de la oblea con una herramienta de escisión de obleas (EMS), ajustando su tamaño al objetivo del proyecto. 2 cm x 4 cm es conveniente tanto para observaciones de luz como de microscopía electrónica.
- Limpie la oblea en agua destilada para deshacerse de los escombros.
- Descarga de resplandor/plasma limpia la superficie con equipo estándar. Los parámetros precisos dependerán de la máquina utilizada. Comience con los parámetros para la descarga de las rejillas y ajuste empíricamente el tiempo. Este paso es crucial para una buena distribución de las secciones en el soporte mientras las secciones se están secando y no debe omitirse.
- Preparar el soporte de montaje B de las matrices (es decir, cubiertas de vidrio)
- Si planea experimentos de inmunomarcado multiplex, transfiera las muestras en un cubrehojas para detectar mejor la señal de fluorescencia. Para mejorar la adhesividad de la capa de cubierta, utilice un procedimiento detallado de recubrimiento de gelatina que se describe anteriormente9.
- Aumente la conductividad recubriendo las diapositivas con el óxido de indio-estaño (ITO) u oro por evaporación. Mantenga los cubrehojas preparados en un ambiente limpio. Descarga con resplandor las muestras como se describe en el punto 1.2.3.
2. Seccionamiento de la muestra
- Preparación del cuchillo AT
NOTA: Para generar las matrices, utilice un cuchillo modificado (ATS), diseñado para facilitar la adquisición de cintas largas. Los cuchillos Histo-Jumbo o cuchillos similares concebidos para la generación de las secciones en soportes grandes también pueden servir para la seccionamiento AT.- Fije la aguja en la parte inferior del cuchillo con la cinta adhesiva espumosa y perfore (Figura 2B). Coloque el cuchillo ATS en el soporte del ultramicrotomo a 0°. Ajuste el borde de la cuchilla en paralelo a la superficie del bloque utilizando el procedimiento estándar.
- Lleve el bloque recortado al borde del cuchillo en una posición lista para la seccionamiento.
- Coloque la oblea/ funda dentro del recipiente del cuchillo y llénela con agua al mismo nivel que el filo del cuchillo. Deje que el filo de diamante del cuchillo se humidifique correctamente y, si es necesario, extraiga el agua con la jeringa adjunta (Figura 2C).
- Seccionamiento y transferencia de arreglos de discos
- Ajuste el microtomo a los parámetros de corte deseados. Con la cuchilla ATS, el rango de 50-100 nm y una velocidad de corte de 0.6-1 mm / s son recomendables. Iniciar la sección (Figura 2Di).
- Obtenga una cinta de la longitud que cubrirá un volumen z específico y detenga la colección. Dependiendo del tamaño del bloque, la homogeneidad del tejido y el tipo de resina, la cinta será relativamente recta (Figura 2D-ii). Muchas muestras no producirán cintas rectas, a pesar del esfuerzo invertido y el tipo de cuchillo que se utiliza.
- Dependiendo del objetivo final, haz una cinta larga o varias cintas cortas alineadas una al lado de la otra. Un soporte de 2 cm x 4 cm puede sostener convenientemente de 100 a 1,000 secciones. La disposición de la cinta en el escalón de soporte es un paso que requiere destreza y manos firmes. Sin embargo, la curva de aprendizaje para esta habilidad se adquiere rápidamente.
- Separe la cinta del filo del cuchillo con una punta limpia y no pegajosa de una pestaña pegada en el palillo (Figura 2Diii).
- Usando la pestaña, mueva suavemente la cinta por encima del centro del medio de soporte. En este punto, use cloroformo o pluma calefactora para estirar las secciones si es necesario. Sin embargo, recuerde que esta manipulación puede inducir la rotura de la cinta y la deformación.
- Comience a drenar el agua tirando de la jeringa. Para cintas más delicadas o una retracción de agua más lenta, deje que el agua gotee separando la jeringa de la manguera. Cuando el nivel de agua esté bajando al nivel de la oblea, controle la cinta y reposicionarla si es necesario, empujando suavemente la cinta hacia el centro. Después de asentar las cintas en la superficie de la oblea, continúe drenando hasta que el agua restante se retraiga por completo de la cuenca.
NOTA: Si no usa el cuchillo ATS de retracción de agua inferior, reduzca cuidadosamente el agua de los lados del cuchillo ATS para no inducir turbulencias. - Deje que las secciones en el soporte dentro del baño se sequen por completo. Dependiendo de la hidrofobicidad del soporte y del nivel de humedad ambiental, el agua se evaporará a una velocidad diferente(Figura 2Div). Es importante dejar que la muestra se seque lentamente para disminuir o evitar por completo todos los pliegues de la muestra.
- Transfiera la muestra seca a una caja bien cerrada para protegerla de la contaminación por suciedad y colóquela en un horno de 60 °C durante al menos 30 minutos. Si es necesario, contramante secciones que utilizan metales pesados para mejorar el contraste general.
- Al final del procedimiento, limpie con precaución el cuchillo siguiendo las instrucciones del fabricante.
NOTA: La transferencia de secciones múltiples o en serie en una oblea (Figura 2E) en comparación con la transferencia en una rejilla de ranura (Figura 2F) cambia la experiencia de preparación EM por completo. La seccionamiento y la recopilación ininterrumpidos de las secciones en soporte único reducen la cantidad de seccionamiento y los errores relacionados con la recopilación de secciones frecuentes en la sección en serie. El equivalente a 100 secciones de 1000 μm x 500 μm recogidas en una oblea corresponde a 33 rejillas de ranura(Figura 2G).
3. Observación de la muestra
NOTA: En esta sección se describen los pasos del flujo de trabajo implementados mediante un software disponible comercialmente (consulte la Tabla de materiales). Se puede utilizar cualquier software de adquisición de imágenes en cualquier SEM equipado con los detectores correctos, sin embargo, las acciones específicas del usuario diferirán y, a menudo, serán más manuales.
- Adquiera un mapa general de imágenes SEM que revele las ubicaciones de las secciones en la oblea. Una imagen de cámara óptica incorporada ayuda a definir un mosaico de imágenes SEM que cubre una cinta de secciones o todas las secciones. Cree el mosaico haciendo clic y arrastre con el ratón a la derecha en la imagen de la cámara de la muestra e inicie la Adquisición automática.
NOTA: Los ajustes de imagen muy gruesos, es decir, el tamaño de píxel de 1-2 μm y el tiempo de permanencia de 1 μs son suficientes. Este proceso se ilustra en Material Complementario (páginas 2-7). - Localice secciones con la función detección automática del Buscador de secciones o localícelas manualmente. Las posiciones y contornos de las secciones se recuperan automáticamente con este software basado en la coincidencia de imágenes. Para ilustrar este proceso, véase Material complementario (páginas 8 a 15).
- En caso de que las imágenes de resumen no muestren claramente los ROI, adquiera imágenes de mayor resolución de las secciones. Utilice la función Vista previa de sección para crear y adquirir imágenes automáticamente. Este proceso se ilustra en Material Complementario,páginas 16-18. La configuración de imágenes debe elegirse de acuerdo con la naturaleza y el tamaño del ROI que el usuario busca.
- Para encontrar la configuración óptima, active Live Imaging en el software de control del microscopio y navegue a un ROI. Modifique la configuración de imágenes hasta que las imágenes muestren el ROI claramente, pero la adquisición de imágenes no es excesivamente larga.
- Definir regiones de imagen
NOTA: Se proponen varias estrategias diferentes.- Si solo es necesario crear imágenes de unas pocas secciones, use las imágenes de las secciones creadas hasta ahora para navegar a las secciones relevantes y use el Visor ampliable que muestra todas las imágenes adquiridas en sus ubicaciones relativas originales para ver todas las secciones. Una vez que se encuentra una sección que debe ser fotografiada en alta resolución, cree una región de imagen con click-drag. Elija la configuración de imágenes de alta resolución y almacene la configuración en una plantilla. Reutiliza esta plantilla para otras secciones.
- Para encontrar eventos raros particularmente pequeños o difíciles de detectar, use el enfoque de detección lateral. Cree manualmente una región de imágenes con ajustes de imágenes de alta resolución en cada décima sección o en una sección por cinta y adquiera las imágenes. Revise las imágenes y marque las secciones que contienen el ROI en el software o tome nota.
- A partir de secciones que contienen el ROI, navegue hacia adelante y hacia atrás a través del conjunto de secciones y cree regiones de imágenes de alta resolución en las mismas ubicaciones relativas durante el tiempo que la estructura siga siendo visible en la sección. Esto se puede hacer manualmente o utilizando el procedimiento descrito en los siguientes pasos.
- Adquiere imágenes en más de diez secciones consecutivas. Haga clic en Iniciar refinamiento de posición después de ampliar la imagen para aumentar la precisión de las ubicaciones de sección registradas como se describe en el manual. Al hacerlo, disminuye la variabilidad de posición de las series de imágenes. El procedimiento se ilustra y explica en las páginas 19-21 del Material Complementario.
- Haga clic y arrastre en cualquier sección para definir una región de imagen mientras mantiene presionada la tecla Alt y seleccione Crear matriz de conjunto de mosaicos en el menú contextual que se abre cuando se suelta el botón del mouse. Luego, el software crea regiones de imágenes en la misma ubicación relativa en todas las secciones que se han encontrado o marcado anteriormente. Es posible limitar las imágenes a un rango específico de secciones con el control deslizante Section Span.
NOTA: Este procedimiento se puede repetir con cualquier número de regiones de imagen y, por lo tanto, permite grabar muchas imágenes pequeñas de alta resolución en lugar de grabar una sola imagen más grande en cada sección. - Una vez creado, configure el recuento de píxeles, el tamaño de píxel, el diseño de mosaico, el tiempo de permanencia de píxeles, etc. en cada serie de imágenes según sea necesario. Una vez que se crean y configuran las series de imágenes, todas se enumeran en una señal de trabajo.
- Configurar funciones automáticas e iniciar la adquisición de imágenes
- Cree una serie de imágenes independiente para las funciones automáticas utilizando el mismo método que se describe en el paso anterior. Mueva la serie de imágenes a una posición en la sección que contenga estructuras de alto contraste.
- Establezca la serie de imágenes en 1024 x 884 píxeles y elija un tamaño de píxel correspondiente a la resolución más alta utilizada en la serie de imágenes configurada en los pasos anteriores. En la lista de funciones automáticas, marque Enfoque automático y Estigmatizador automático.
- Seleccione Por sección en los controles de la secuencia de adquisición y asegúrese de que la imagen de funciones automáticas es el primer elemento de la lista. Inicie la adquisición de la imagen haciendo clic en el botón Ejecutar situado junto a la señal de trabajo. Estos procedimientos se ilustran en Material Complementario,páginas 22-23.
NOTA: No es necesario preenfocar manualmente en cada sección. Durante la sesión de grabación, cada vez que el microscopio avanza a una nueva sección, las funciones automáticas se ejecutarán antes de que todas las demás imágenes se graben en esta sección.
4. Alineación y análisis de datos
- Exportación de datos
- Asegúrese de que los datos se guardan en formato .tif, por lo que no es necesario una función de exportación dedicada. Ordene los datos en una estructura de carpetas que corresponda directamente a las capas y elementos del árbol de capas.
- Una vez que se hayan grabado los mosaicos de imágenes, utilice la función StitchAll para coser automáticamente todos los mosaicos.
- Alineación de pilas y recorte en Fiji
NOTA. Muchos paquetes de software (gratuitos y comerciales) se pueden utilizar para trabajar con datos de array tomography. Los pasos a continuación se muestran con el programa de código abierto Fiji32 porque está ampliamente disponible y contiene todas las funciones requeridas.- Importe una pila de imágenes (o imágenes cosidas) a Fiji como una pila virtual.
- Si es necesario normalizar el contraste/brillo, elija Contraste mejorado... en el menú Proceso. Establezca Píxeles saturados en 0,1 o menos y marque Procesar todos los sectores.
- En el menú Plugins, selecciona Registro | Alineación lineal de la pila con SIFT.
- Elija Rígido o Afín en el menú desplegable Transformación esperada. De lo contrario, mantenga la configuración predeterminada. Inicie la alineación haciendo clic en Aceptar.
NOTA: La carga de los datos como pila virtual permite a Fiji manejar pilas de cualquier tamaño. La salida de la alineación se crea en RAM; sin embargo, esto puede limitar el tamaño máximo de las pilas que se pueden procesar. En ese caso, utilice Register Virtual Stack Slices, que es una implementación de carpeta a carpeta del mismo algoritmo de registro. Una vez completado el registro, cargue los datos de salida como una pila virtual. - Recorte la pila de imágenes haciendo clic en Recortar para que contenga solo el ROI.
- Guarde la pila como una sola imagen .tif o una serie de imágenes .tif.
NOTA: Los pasos críticos de la tomografía de matriz se muestran en la Figura 3.
Resultados
Los siguientes ejemplos tienen como objetivo demostrar la versatilidad de los flujos de trabajo recomendados. Las ilustraciones de estudio de caso son proyectos para los que tuvimos dificultades para obtener resultados satisfactorios con cualquier otra técnica. Elegimos Drosophila adulto para ilustrar los desafíos típicos que uno podría encontrar con numerosos tipos de muestras. Este órgano tubular de aproximadamente 6 mm de largo, 500-1000 μm de sección transversal, se divide en diferentes regiones con una función única y composición celular (Figura 4A)33. Dependiendo de la orientación de la sección, las dimensiones del perfil intestinal y su apariencia en la sección varían. Las secciones orientadas transversalmente o longitudinalmente son relativamente grandes, y solo se pueden colocar un par en una sola cuadrícula TEM(Figura 2F). Solo se puede obtener una pequeña porción del tejido en el FIB, y para el SBF-SEM, la dificultad es similar a cualquier muestra no homogénea. AT proporciona una compensación eficiente para el análisis de dichas muestras y la incrustación plana facilita la localización del ROI. El recorte cuidadoso del exceso de resina alrededor del área seleccionada(Figura 4B)es importante para la recolección eficiente de las matrices del área relevante(Figura 4C). Cientos de secciones se pueden recolectar en una sola oblea de forma secuencial o aleatoria(Figura 4D). Dependiendo de la pregunta de investigación, la selección y adquisición de muestras requerirá una estrategia diferente, que arbitrariamente dividimos en varios escenarios. Para ilustrar los diferentes escenarios presentados de una manera más específica, elegimos varios estudios de caso de los diferentes proyectos de investigación.
Análisis de numerosas estructuras grandes distribuidas aleatoriamente en el rango de 1-10 μm (Figura 4E)
Con frecuencia, se requieren datos ultraestructurales para validar una hipótesis que surgió de varios enfoques experimentales, comparando una condición estándar y alterada experimentalmente. En estos casos, varias secciones suelen recopilarse aleatoriamente en cuadrículas y filtrarse para localizar e imaginar las áreas de interés. Esta táctica suele ser menos sistemática y limitada a un pequeño número de secciones analizadas. Sugerimos grabar resúmenes de decenas / cientos de secciones de resolución media de una cinta dada (Figura 4D). Para las secciones típicas de 70 nm, 200 secciones abarcarán aproximadamente 14 μm, que contendrán numerosas celdas, ya sea total o parcialmente, completadas en media hora. Como primer paso, se registra la descripción general de baja resolución de toda la cinta, y la descripción general ayuda a omitir las secciones que muestran artefactos de preparación (por ejemplo, pliegues, suciedad). Después, la adquisición se puede realizar de forma manual o automática directamente en partes seleccionadas de la sección, o en una sección completa, utilizando imágenes individuales o en mosaico, seguidas de costuras (Figura 4E). Después, las imágenes del área seleccionada se pueden adquirir utilizando parámetros de alta resolución. Por ejemplo, las mitocondrias, los núcleos y las microvellosidades pueden beneficiarse de un método estadísticamente mejorado(Figura 4Ei-iii).
Análisis de múltiples estructuras pequeñas y escasamente distribuidas en el rango de 500-1.000 nm (Película suplementaria 1)
En este escenario, el ROI no se puede identificar simplemente en un escaneo general de bajo aumento, y se necesitan imágenes de alta resolución. En las muestras TEM convencionales, el tedioso acercamiento y alejamiento de la sección es necesario hasta que se encuentre la característica requerida. A menudo, la obtención de imágenes de varias ubicaciones independientes en múltiples muestras es estadísticamente más relevante que la generación de un solo gran volumen. En tales casos, la complejidad de la adquisición manual crece exponencialmente. Aunque varias soluciones TEM permiten las adquisiciones automáticas o la detección de múltiples rejillas, el tamaño de la red y los desafíos de seccionamiento en serie con frecuencia hacen que el enfoque sea incompatible para muchas muestras. Para casos similares, generamos un mapa completo de resolución media de un ROI general en múltiples secciones a una resolución suficiente para identificar las estructuras de interés. Durante este paso de cribado lateral, es aconsejable saltar varias secciones a la vez, con el objetivo de golpear al menos una parte de la estructura de interés cuando se aborda al azar. Esto dependerá en gran medida de las dimensiones generales de la estructura: por ejemplo, si el tamaño total de la estructura es de 500 nm y las secciones tienen un grosor de 50 nm, al menos nueve secciones secuenciales seguidas probablemente contendrán una parte de la estructura de interés. De esta manera, el salto de 6-7 secciones será eficiente para encontrar muchos tipos diferentes de estructuras en múltiples áreas. La adquisición automática de los mapas de mosaico resueltos de las secciones seleccionadas permite una cuidadosa selección de estas secciones después de su adquisición. Una vez que se adquiere un mapa de alta resolución de este tipo, se pueden recortar varios ROI o utilizarlos para definir áreas de imágenes locales adicionales en los ROI(Película suplementaria 1). Golgi, centriolos, uniones, microtúbulos, diferentes tipos de vesículas son buenos ejemplos de las estructuras que podrían beneficiarse de este escenario (Película complementaria 1).
Análisis de grandes ROI escasamente distribuidos en muestras grandes (Figuras 4F-4H)
Este escenario involucra eventos raros, que con frecuencia se describen como "una aguja en un pajar" en la que el problema no está en la identificación del ROI sino en su localización. Para muchas muestras, el enfoque correlativo no es una opción válida, sin embargo, con frecuencia el ROI tiene una ultraestructura reveladora y, cuando se localiza, se puede identificar con alta confiabilidad. Para estas muestras, es esencial aplicar la adquisición multinivel, comenzando con las muestras preseleccionadas con decenas a cientos de secciones a resolución media. En el software utilizado aquí, hay dos estrategias diferentes para obtener conjuntos de imágenes de múltiples secciones: Grabar las imágenes de vista previa a una resolución más alta o adquirir un conjunto de mosaicos de matriz con la configuración adecuada(Figura 4F). Diferentes tipos de células especializadas en el intestinode Drosophila se distribuyen aleatoriamente (por ejemplo, células madre, enteroendocrinas) y se seccionan delgadas en orientación aleatoria. Sin embargo, se pueden distinguir visualmente después de examinar las imágenes obtenidas utilizando parámetros de alta resolución, ya sea de secciones individuales o como una colección de imágenes en serie(Figura 4G). Después de la alineación, las pilas se pueden representar utilizando diferentes soluciones de software(Figura 4H,Película suplementaria 2).
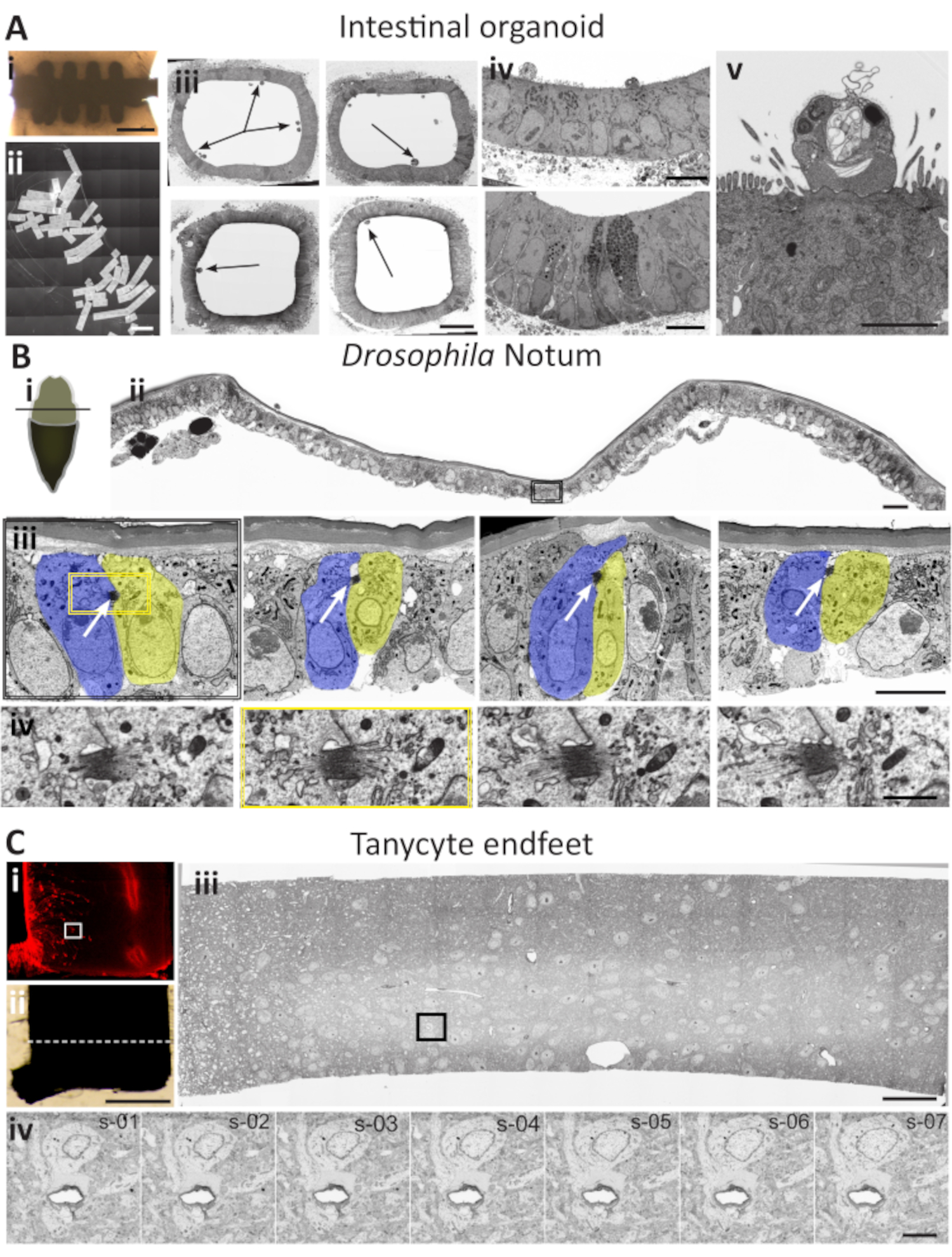
Escenario 1: Organoides intestinales (Figura 5A)
Los organoides se están convirtiendo rápidamente en una de las herramientas más vanguardistas de las ciencias de la vida modernas. Este modelo de órganos derivados de células madre 3D casi fisiológicos hace posible un estudio preciso de una variedad de procesos biológicos in vivo, incluida la renovación de tejidos, la respuesta a los medicamentos y la medicina regenerativa. Los minitubos intestinales34 recientemente introducidos abren una nueva generación de tecnología organoide, que se asemeja mucho a la fisiología de los tejidos in vivo, la composición del tipo celular y la homeostasis, lo que permite amplias perspectivas para el modelado de enfermedades, la interacción huésped-microbio y el descubrimiento de fármacos. Sin embargo, cuando se requiere la caracterización ultraestructural, la localización de diferentes tipos de células en tejidos tan grandes utilizando muestras aleatorias puede ser un desafío. Además, en los ensayos de "infección" variable, es crucial asegurarse de que el análisis revele diferentes etapas del desarrollo que afectan al tejido. Para tales estudios, la cobertura estadísticamente significativa de la muestra es central, pero difícil de lograr utilizando el enfoque tradicional de TEM en la red. El escenario AT-1 es beneficioso en tales casos: se pueden generar muchas secciones secuenciales en una oblea(Figura 5Aii)y examinarse utilizando parámetros de baja resolución para localizar las áreas generales de interés(Figura 5Aiii;flechas). Estas áreas pueden ser objeto de análisis posteriores utilizando parámetros de adquisición avanzados(Figuras 5Aiv y Figura 5Av). Cuando se detecta una estructura relevante (normalmente un grupo de 5-10 secciones una vez en cada 100-300 secciones), es fácil concentrarse en cada una de las estructuras de interés y adquirir imágenes individuales manualmente o utilizar las funciones de automatización para adquirir volúmenes de imágenes en múltiples secciones.
Escenario 2: Drosophila pupal notum ( Figura5B)
Estudiar la división celular y los mecanismos que controlan la progresión a través del ciclo celular es crucial para comprender los procesos estándar y alterados en los organismos multicelulares. La información que existe se deriva con frecuencia de sistemas unicelulares; sin embargo, esta solución carece del contexto crítico de las interacciones 3D entre las células de un tejido. Una monocapa unicelular del notum, la parte posterior en desarrollo de la larva de Drosophila, es un modelo perfecto para la interacción entre las células epiteliales en general y la división celular en particular35. Es un modelo establecido para estudios de interacciones moleculares y celulares utilizando la combinación de los datos disponibles por microscopía fluorescente y manipulaciones genéticas. La abscisión, el último paso de la división celular, asegura la separación final entre dos células en división, y caracterizar los cambios estructurales que ocurren durante la abscisión es esencial para nuestra comprensión de la mitosis. Sin embargo, las divisiones mitóticas en el notum no son fáciles de localizar a nivel ultraestructural: las células son relativamente grandes, en comparación con la zona de abscisión(Figura 5B). La relación entre el tamaño total de la zona de abscisión y la superficie de la sección a cubrir es grande (Figura 5Bi). Aunque es posible localizar la zona de abscisión utilizando los métodos TEM o SBF-SEM36,la tarea es laboriosa. Con este escenario, las imágenes generales automáticas de resolución media de los saltos de 20-40 secciones se pueden utilizar para localizar las células en división (Figura 5Bii). Cuando se identifican tales células, las secciones sirven como ancla para un examen más detallado de las secciones en las cercanías, y se pueden encontrar y seleccionar numerosas células en división para un análisis más detallado. De esta manera, la zona de abscisión puede ser localizada e fotografiada en su totalidad(Figura 5Biii). Dependiendo de la pregunta, se pueden recopilar imágenes individuales de alta resolución o secuencias de 3-7 imágenes para cubrir la profundidad de la estructura(Figura 5Biv).
Escenario 3: Neuronas tanicitarias de ratón (Figura 5C)
El ratón proporciona un modelo bien establecido para el desarrollo del cerebro y está bien documentado en diferentes niveles, incluso por EM. A pesar de que se han utilizado ampliamente diferentes métodos automatizados de cara de bloque en serie para estudiar el tejido cerebral, hay casos en los que la TA se adapta mejor para recopilar los datos necesarios. El hipotálamo es un modelo de neurociencia bien establecido, una parte del cerebro que contiene múltiples funciones de tipo neuronal. Los tanicitos hipotalámicos representan un subconjunto particular de células ependimogliales que recubren la parte inferior del tercer ventrículo, con procesos inusualmente largos (hasta 300 μm) y grandes patas finales (~5 μm)37. Esto los hace inconvenientes para el análisis por métodos TEM o FIB. La tarea se complica aún más cuando varios tanicitos independientes necesitan ser localizados y analizados. Uno de los enfoques para facilitar esta tarea puede ser la focalización semi-correlativa, en la que el mapa de fluorescencia se obtiene de las muestras marcadas fluorescentemente antes de fijar e incrustar para EM. El seccionamiento se realiza en el área capturada combinando la información posicional de la muestra de fluorescencia y la réplica de plástico plano incrustado. Después de eso, se puede usar el escenario AT 3: las imágenes de mosaico de visión general de alto nivel se generan para revelar las regiones con grupos de pies finales de tanicitos. Posteriormente, las funciones de automatización en el software se utilizan para configurar la adquisición de secuencias de las imágenes de una o varias áreas en una sola imagen o modo en mosaico. Estas imágenes se pueden analizar por separado, como pilas alineadas o renderizadas después de eso.
La potencia del método AT permite la "actualización" relativamente fácil de los datos de 2D a 3D: los mapas están disponibles desde la adquisición primaria y los volúmenes se pueden obtener del área seleccionada y sus alrededores. La pila resultante se puede alinear y posteriormente renderizar. Es esencial determinar de antemano qué resolución y calidad de imagen son necesarias para encontrar ROI. Las imágenes deben permitir reconocer el ROI, pero no más allá de este valor porque el tiempo de adquisición escala proporcionalmente al tiempo de permanencia de píxeles y al cuadrado inverso del tamaño de píxel.

Figura 1: Desafíos de la preparación de muestras EM y adquisición de volumen. (A) La pérdida de fluorescencia y contracción ocurre debido a una alta concentración de metales pesados y deshidratación durante la preparación de la muestra. (i) Un dibujo esquemático de un espécimen observado bajo LM (ii) el mismo espécimen preparado para EM, que se vuelve completamente opaco y pierde alrededor del 10% de su volumen. (B) La incrustación de muestras se realiza típicamente utilizando resinas epoxi o acrílicas. Los bloques tradicionales (i) se pueden utilizar con éxito para muestras homogéneas que no requieren una orientación particular. Los bloques planos (ii) son útiles cuando es esencial apuntar y orientar bajo el microscopio, un área precisa destinada a la seccionamiento, por ejemplo, en muestras no homogéneas o procedimientos de microscopía correlativa. (C) De todo el volumen de la muestra, sólo una fracción limitada se representa en una sola sección de 50 nm, proporcionando una imagen 2D de una muestra 3D, a menudo en una orientación desconocida. (D) Para ilustrar el problema de la grabación de volúmenes demasiado grandes frente a la orientación precisa, elegimos tres cubos concéntricos con las caras de 1000, 500 y 50 μm organizados para incluir una muestra hipotética de 1000 x 500 x 500 μm (granate oscuro). Si estos cubos de muestra hipotéticos se seccionan completamente con segmentos de 50 nm, para cubrir todo el volumen, se requerirá un total de 20,000, 10,000 y 1,000 cortes, y 800 tb, 100 tb y 100 gb, en consecuencia (resolución de imagen de 5 nm x 5 nm x 50 nm, datos de 8 bits). Esto muestra la importancia de planificar la adquisición de datos EM solo para adquirir el volumen mínimo necesario. (E) Cubrir una gran superficie de muestra en alta resolución presenta un problema similar al de gran volumen. Mosaicar varias imágenes de alta resolución en una es una solución útil para tal problema. Sin embargo, para cubrir una superficie de 1 mm x 1 mm utilizando el marco 2024 x 1048 en un aumento de 10,000x se requerirá una gran cantidad de baldosas, que pueden llegar a ser difíciles de coser. Además, si las secciones se comprimen o distorsionan de forma variable durante el corte, las pilas de datos resultantes se vuelven casi imposibles de alinear. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: El flujo de trabajo para la generación directa de las matrices de secciones en soporte grande. (A) Para el recorte ajustado utilizando la herramienta de recorte, los bloques se aseguran dentro del soporte de microtomo. Este paso ayuda a asegurar lados paralelos de un bloque y también reduce la resina vacía alrededor de la muestra. (B) Un cuchillo modificado para las adquisiciones de secciones AT. Un barco grande facilita la transferencia de las secciones y su manipulación durante la sección de muestras y la transferencia en el soporte. Una gran cuenca permite la manipulación de las secciones; el sistema de drenaje limita el movimiento de las cintas durante el paso de drenaje, el fondo plano hace que el secado gradual del soporte sea confiable. (C) El cuchillo, listo para seccionar con una oblea descargada de resplandor colocada en el fondo de la cuenca y nivelada con agua hasta los bordes. La construcción del cuchillo mantiene la aguja incrustada, sin interferir con el soporte. (D) Generación de matrices, vista superior en el área de seccionamiento del microtomo. (i) Las primeras secciones suelen ser fáciles de obtener, ya que se adhieren entre sí y forman una cinta regular. (ii) Cuando se agregan más secciones a la cinta y se hace más larga, la cinta pierde su estabilidad y con frecuencia se curva. Es crucial mantener las pistas de secuencia organizadas en secuencia en preparación para el paso de adquisición de imágenes. (iii) Cuando una cinta de secciones alcanza la longitud deseada, se separa cuidadosamente del filo de la navaja usando una pestaña. iv) El agua se drena de la cuenca; la oblea permanece en el interior hasta que está completamente seca al aire. Este paso es esencial, ya que ayuda a enderezar las secciones y evitar la formación de los micro pliegues. La oblea se coloca en el horno a 60 °C durante al menos 30 minutos para fijar las secciones en el soporte. (E) Ejemplo de obleas con las secciones transferidas. Aunque es conveniente obtener cintas rectas y precisas, las muestras reales evitan la formación de tales cintas ideales en la mayoría de los casos. Sin embargo, incluso las cintas "descuidadas" son muy informativas para la gran cantidad de casos, y la importancia de la cinta "ordenada" dependerá de una estrategia de investigación para la cual se recopilen las secciones. Barra de escala 1 cm. (F) Cuadrículas de ranura de ejemplo con las secciones serie. Incluso cuando muchas secciones se recogen en una cuadrícula, sigue siendo una pequeña fracción de lo que se puede recolectar en una sola oblea. La habilidad requerida para dominar la transferencia de las secciones en una cuadrícula (cuadrícula de ranura en particular) representó un cuello de botella significativo para dominar la preparación de muestras de microscopía electrónica. Barra de escala 500 μm. (G) Independientemente del método de recolección de secciones que se haya utilizado, las fortalezas del enfoque AT son la relativa facilidad de generación de secciones secuenciales, en comparación con la recolección en la red. Si se considera un bloque de muestra de 1000 μm x 500 μm, no hay problema para colocar alrededor de 100 secciones en una oblea (i) de 2 cm x 4 cm. Las secciones del mismo tamaño en una cuadrícula de ranura se ajustarán solo a tres secciones / cuadrícula como máximo (ii). Proporcionamos una imagen a escala para mostrar cuántas cuadrículas podrían ser necesarias para cubrir el mismo número de secciones, sin mencionar la dificultad de recopilar secciones en serie en la cuadrícula. Barra de escala = 1 cm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3:Pasos críticos del flujo de trabajo de la tomografía de matriz. Esquema del flujo de trabajo para la adquisición desatendida de pilas de imágenes de alta resolución. Todos los pasos preparatorios están automatizados (símbolos de engranajes verdes) y no requieren que ninguna acción se realice manualmente, sección por sección. Las pilas de imágenes se pueden alinear en cualquier software de análisis de imágenes capaz de alinear automáticamente de forma rígida o afín. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Tres escenarios de adquisición con intestino adulto de Drosophila como modelo de demostración. (A) dibujo de un intestino medio de Drosophila diseccionado, con tres regiones principales designadas por diferentes colores: anterior, medio y posterior. (B) Bloque plano recortado en el que un intestino está orientado para la sección transversal. Tenga en cuenta que la cantidad de resina vacía se equilibra cuidadosamente alrededor del tejido que contiene la región de interés (rectángulo blanco). (C) Las secciones seriales transversales están flotando en la superficie del agua dentro de la cuenca del cuchillo AT. Todas las imágenes se adquirieron en modo SEM de electrones secundarios utilizando el detector de espejo con el contraste inverso. (D) Imagen de mosaico cosido de secciones seriales transversales en una oblea. La barra de escala es de 1000 μm. (E) Sección transversal a través del intestino de Drosophila. Barra de escala 20 μm. La imagen es un mosaico cosido de 7 x 7 imágenes de rango medio. Recuadros: imágenes de mayor aumento y resolución de las regiones específicas de interés: (ii) núcleo, (iii) borde de cepillo y (i) mitocondrias. Barra de escala de 5 μm para todos. (F) Una imagen de rango medio de una sección transversal a través del intestino que se dirige a la ubicación de las células en desarrollo (cuadrado). La barra de escala es de 20 μm. (G) La matriz objetivo de secciones en serie recopiladas en función del área localizada durante el análisis de la sección presente en el panel F. Barra de escala es de 10 μm. (H) El modelo 3D se representa en función de la secuencia de pila de 50 secciones obtenida de la adquisición en serie objetivo en el panel G. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Estudios de caso para los escenarios de aplicación de AT. (A) Localización de diferentes estructuras celulares en el organoide intestinal. i) Microchip de silicio integrado. Barra de escala = 200 μm. (ii) Una imagen de mosaico cosida de 127 secciones transversales a través de la parte central del chip. Barra de escala = 1500 μm (iii) Cuatro imágenes de baja resolución de una sección transversal completa a través de la porción del organoide intestinal. Las flechas apuntan al sitio potencial de interés. La barra de escala es de 20 μm. (iv) Diferentes ROI, dirigidos a micrografías de baja resolución seleccionadas para su posterior análisis. Barra de escala = 10 μm. (v) Imagen de alta resolución de la célula infectada de interés. El análisis de la misma región en las secciones adyacentes puede proporcionar información 3D específica si es necesario. Barra de escala = 5 μm. (B) Localización de cuerpo medio en Drosophila melanogaster pupal notum. (i) Una vista esquemática de una pupa de Drosophila diseccionada. Notum expuesto para la disección (beige) después de quitar la parte de la cutícula protectora (marrón). La línea negra designa la dirección de la sección (ii) Una sección transversal a través del área presentada en el diagrama. La imagen combina imágenes SEM de alta resolución tomadas secuencialmente de 3x7 cosidas a un panel de mosaico. El rectángulo negro delimita el área que contiene una célula en división. La barra de escala es de 15 μm. (iii) Una imagen ampliada en la celda en división del panel ii. A este aumento y resolución, el cuerpo medio es evidente (flechas blancas). Se analiza toda la sección para encontrar las células en división. Saltar entre diferentes cintas de secciones en intervalos de 20-30 secciones durante el paso de cribado lateral permite localizar numerosos pares de células en división. La barra de escala es de 5 μm (iv) cuando se localiza una célula en división, imágenes secuenciales del cuerpo medio recogidas de cuatro secciones alrededor del cuerpo medio delimitadas por un cuadrado amarillo en el panel (iii). La barra de escala es de 1 μm. (C) Localización de los pies finales de tanycytes en el hipotálamo de ratón. (i) Imagen de fluorescencia de una rodaja de vibratomo. Los tanicitos expresan tdTomato proteína fluorescente (rojo). Un rectángulo blanco delimita el área de interés. Barra de escala 500 μm. (ii) La misma sección del vibratomo preparada para EM se recortará cuidadosamente alrededor del área de interés, basándose en la correlación indirecta de la información fluorescente del panel (i). La línea blanca punteada representa el área de seccionamiento ultrafino. La barra de escala es de 50 μm para ambos paneles. (iii) Sección transversal a través de la rebanada del vibratoma en el área de interés. La imagen de mosaico SEM se compone de 75 imágenes cosidas. Varias secciones son objeto de cribado lateral y se toman imágenes con parámetros similares. Las secciones se analizan "fuera de línea" para encontrar el ROI: los pies finales del tanicito. El rectángulo negro representa el área que contiene los extremos del tanicito. Esta sección servirá como ancla para un análisis más profundo. La barra de escala es de 15 μm. (iv) Imagen de alta resolución y alto aumento de los pies finales de los tanicitos que rodean un vaso sanguíneo. Después de la localización inicial del ROI en una sección, la secuencia z se recoge de las secciones adyacentes aguas arriba a la sección de anclaje (panel iii). Barra de escala 5 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

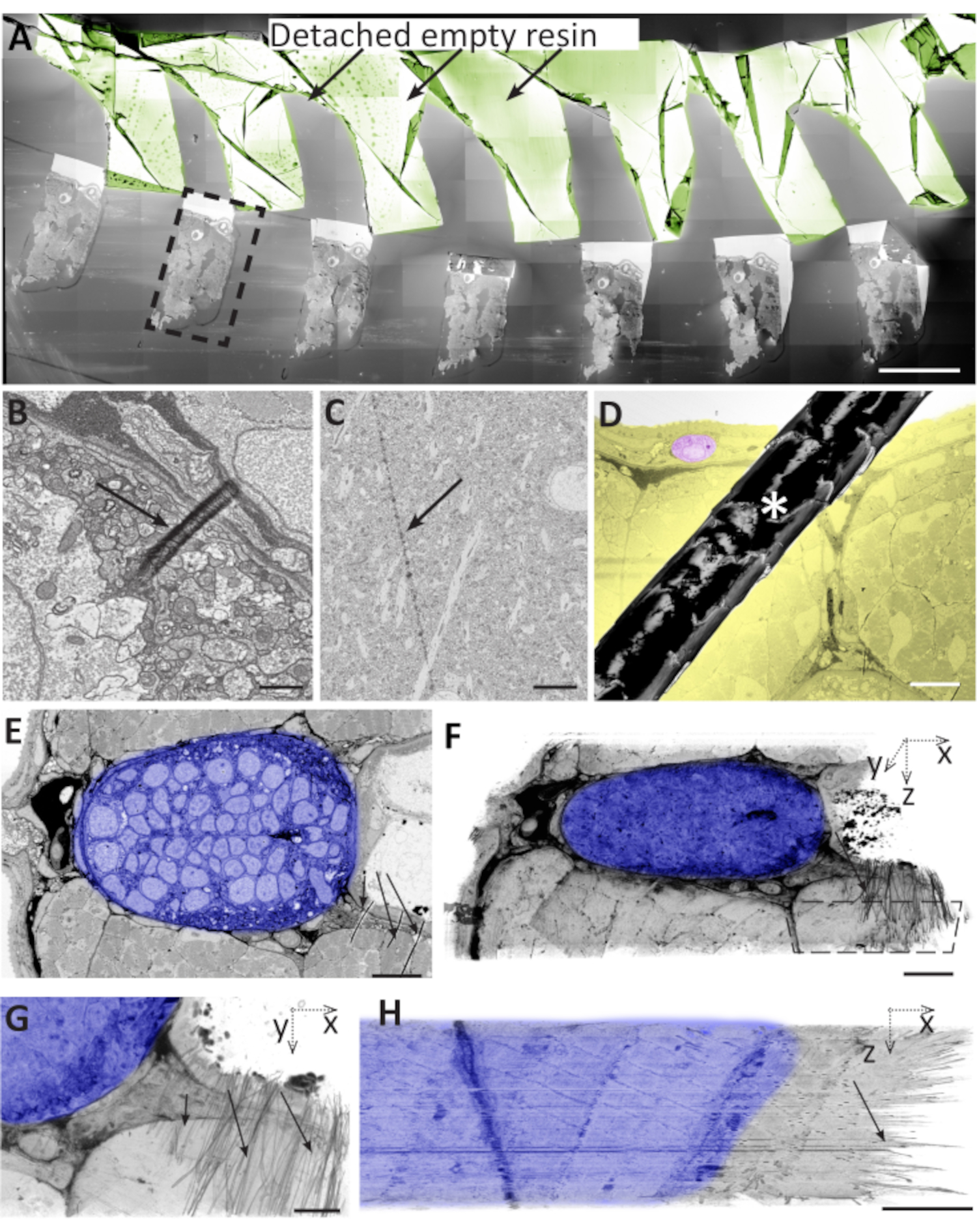
Figura 6: Los problemas durante la ultramicrotomía, la recolección de secciones y el almacenamiento de secciones pueden conducir a artefactos. ( A )Seccionesde muestras cerebrales en una oblea. La mayor parte de la resina vacía se desprendió del tejido y se dobló sobre sí misma (sepia). El cuadro negro discontinuo designa un área que se utiliza para contener toda la sección. Barra de escala 500 μm. (B) Pliegue pequeño y local en la superficie de una sección de pez cebra de 50 nm de espesor. Barra de escala 1 μm. (C) Marca de cuchillo en la superficie de una sección cerebral de ratón de 70 nm. Barra de escamas 5 μm. (D) El pelo (asterisco) en la superficie de la oblea que cubre parcialmente una sección muscular del pez cebra. En amarillo, el tejido es el objetivo para el análisis. En rosa, una célula utilizada para servir como referencia para el tamaño del área afectada. Barra de escala 50 μm. (E) Pez cebra notocorda con arrugas en la parte inferior derecha (flechas negras), donde el tejido neural denso (sombreado en azul) bordea el tejido muscular más blando y la resina vacía (flechas negras). Barra de escala 10 μm. (F) Segmentación del volumen de una pila de 50 imágenes como en E, mostrando que esta región mostró arrugas en la mayoría de las secciones. El polígono discontinuo delinea el área que se muestra en la barra de escala G. 10 μm. (G) Vista XY de la misma segmentación de volumen que en F, mostrando las arrugas como trazos negros cortos en la mitad derecha del bloque. Tenga en cuenta que la alineación de la pila en las partes restantes del tejido no se ve afectada por las arrugas. Barra de escala 5 μm. (H) Proyección XZ de la misma área que en G, mostrando las arrugas en las 50 secciones. Barra de escala 5 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Película complementaria 1: Una imagen de montaje de alta resolución de una sección transversal a través del intestino medio anterior de Drosophila. Imagen de mosaico de una imagen SEM SE-MD invertida. Se adquirieron automáticamente 352 mosaicos de imágenes separados a una resolución de 5 nm y se cosieron para presentar toda la sección transversal. Es posible hacer zoom para obtener más detalles y obtener una cobertura exhaustiva de los datos, utilizando la misma imagen. Uniones estrechas, microtubuli, diferentes tipos de vesículas pueden ser al hacer zoom. La barra de escala es de 10 μm. Haga clic aquí para descargar esta película.
Película complementaria 2: Representación de células intestinales de Drosophila. Cincuenta imágenes de mosaico alineadas de las secciones en el área de las células intestinales en división. Representación IMOD de los bordes celulares (azul, turquesa y naranja) y núcleos (blanco). Haga clic aquí para descargar esta película.
Materiales complementarios. Haga clic aquí para descargar este archivo.
Discusión
La microscopía electrónica da una idea de la ultraestructura de las células y los organismos, para lo cual a menudo es deseable obtener imágenes de estructuras de interés en su contexto de 3 dimensiones. A pesar de las numerosas tácticas de EM para el análisis ultraestructural, todavía no existe una solución "estándar de oro". La razón principal es la gran variedad de muestras, muchas preguntas biológicas, que con frecuencia requieren un enfoque personalizado. El flujo de trabajo de AT propuesto está diseñado para minimizar el tiempo necesario para el procesamiento de muestras, la adquisición de datos, la evaluación y el almacenamiento. Además, el cuchillo modificado proporciona una herramienta útil para simplificar la adquisición de matrices. El diseño compacto de las secciones en las obleas es conveniente, tanto para la observación como para el almacenamiento posterior de las muestras. Esta disposición permite el "cribado lateral" de muestras moviéndose horizontalmente de cinta a cinta y escaneando solo una sección en cada una, lo que reduce significativamente el tiempo requerido para localizar un ROI. Los escenarios de adquisición de datos propuestos facilitan la focalización de áreas pequeñas y distribuidas aleatoriamente. Una vez encontradas, AT/SEM restringe las imágenes de alta resolución precisamente al volumen de interés, ya sea que se realicen manualmente o con la ayuda de la función automática. AT para volúmenes limitados se puede completar manualmente, con el operador navegando a través de la muestra y definiendo las regiones de imagen una por una. El módulo automatizado del software proporciona una estrategia flexible de adquisición de imágenes para la obtención de imágenes de áreas pequeñas en secciones grandes. La automatización en este software permite grabar grandes imágenes de alta resolución en cientos de secciones, logrando volúmenes similares a los de SBFI. La grabación de imágenes generales de todas las secciones simplifica la localización del ROI y reduce el tiempo que se pasa en el microscopio. Dado que las secciones no se dañan durante la grabación de resúmenes y vistas previas de mayor resolución, AT / SEM permite reutilizar la muestra para recopilar más datos de otros ROI o a una resolución más alta.
El tiempo de adquisición de imágenes es uno de los aspectos más importantes (y más caros) de 3D EM y, por lo tanto, debe considerarse en el diseño del experimento. Si bien no es sorprendente que la obtención de imágenes de áreas grandes tome más tiempo que la obtención de imágenes de áreas pequeñas, es fácil subestimar el impacto: dependiendo de los parámetros de imagen seleccionados, el tiempo de adquisición en cada sección puede variar de segundos a horas. Los parámetros críticos de imagen incluyen el tamaño del campo de visión, la resolución y el tiempo de permanencia. Suponiendo una resolución objetivo de 10 nm por píxel y un tiempo de permanencia de 1 μs, la obtención de imágenes de un campo de 20 μm x 20 μm, 100 μm x100 μm o 500 μm x500 μm tarda 4 segundos, 100 segundos o 2.500 s en grabarse. Podemos multiplicar estos tiempos de imagen por sección por el número de secciones para estimar el tiempo requerido para el trabajo completo de imágenes. Los tiempos largos de obtención de imágenes por sección pueden ser aceptables si el número de secciones es pequeño o si el tiempo de la herramienta del microscopio no es motivo de preocupación.
Sin embargo, es necesario limitar el tiempo de grabación a un trabajo nocturno o un trabajo de fin de semana en la mayoría de los casos. Un aspecto igualmente crítico de 3D EM que debe considerarse es la cantidad y estructura de los datos de imagen resultantes. El registro de los campos de imagen mencionados anteriormente en 100 secciones genera 400 mb, 10 gb o 250 gb de datos de imagen, respectivamente; las imágenes de 500 μm x 500 μm plantean el problema adicional de ser mayores de 2 gb cada una. Muchos de los programas de software utilizados para la evaluación de datos no pueden abrir imágenes de este tamaño.
Para reducir el tiempo de obtención de imágenes, es importante elegir el tiempo de permanencia de los píxeles para cumplir con los requisitos de relación señal-ruido para la evaluación de datos posterior (por ejemplo, reconstrucción, rastreo) y limitar las grabaciones a ROI definidos. La extensión AT del software facilita la adquisición de imágenes en áreas pequeñas en secciones en serie. El software admite flujos de trabajo manuales y automáticos y muchas variantes semiautomáticas: las áreas de imagen se pueden colocar y enfocar manualmente en cada sección, o el usuario puede usar el buscador automático de secciones y las funciones de alineación de posición. Dependiendo del nivel de automatización elegido y respaldado por el tipo de muestra o los objetivos de imagen, el tiempo requerido para configurar la adquisición de imágenes en cientos de secciones puede tomar un día de trabajo completo (hecho manualmente) o solo unos minutos. En principio, la tomografía de matriz hace que sea más difícil que otros métodos 3D EM adquirir pequeños ROI; la colocación imprecisa de la región en secciones consecutivas debe compensarse mediante la adquisición de áreas más grandes. Por ejemplo, si el ROI es de 20 μm x 20 μm de tamaño y la variabilidad de posición de sección a sección de los campos de imagen es de 10 μm, uno necesita adquirir imágenes de 40 μm x 40 μm para asegurarse de que el ROI se capture completamente en cada imagen, en cada sección. La variabilidad de la posición de la imagen en el mundo real varía de 100 μm a <10 μm dependiendo de la disponibilidad o calidad de las funciones del software para la alineación de la posición o la paciencia del usuario. Con este software, se pueden lograr 10 μm sin demasiada intervención manual en la mayoría de las muestras.
Como cualquier técnica, el AT tiene varios puntos débiles que pueden influir en la adquisición exitosa de datos, y muchos son similares a otros métodos basados en seccionamiento. La falta de distribución homogénea de resina vacía versus tejido puede resultar en matrices curvas o rotas. En casos extremos, las secciones pueden separarse del soporte (Figura 6A). La compresión o estiramiento variable durante el proceso de corte puede crear pliegues que pueden interrumpir la muestra en regiones variables en secciones posteriores(Figura 6B). Las marcas de cuchillo pueden aparecer en la superficie de las secciones recolectadas usando un cuchillo dañado (Figura 6C). Las diferencias en las condiciones de seccionamiento pueden inducir diferencias ocasionales de compresión y grosor de la sección. Las partículas de polvo o suciedad pueden aterrizar en una sección y oscurecer parcialmente la zona de interés(Figura 6D). La adquisición de imágenes puede fallar debido a funciones imperfectas de contraste automático, enfoque automático y estigmatizador automático. El posicionamiento de las áreas de imagen creadas automáticamente puede ser variable y puede no capturar el ROI en todas las secciones.
Pueden surgir varios problemas en la etapa de costura y alineación. La costura automática de las adquisiciones de azulejos de mosaico puede fallar, por ejemplo, debido al gran espacio vacío dentro de la muestra. Debido a un cambio drástico en la forma en 3D, las pilas de imágenes pueden ser difíciles de registrar. Los programas especialmente desarrollados (por ejemplo, IMOD, Fiji, TrackEM2, MIB o MAPS-AT) pueden facilitar la alineación semiautomática32,38,39,40. Las secciones más desafiantes se pueden alinear manualmente utilizando el software de edición de fotos. Desafortunadamente, algunos conjuntos de datos pueden ser imposibles de alinear correctamente.
Las muestras grandes son un desafío para las secciones serie TEM que se ajustan a las rejillas; por otro lado, muchos proyectos no justifican la adquisición automática prolongada utilizando FIB/SEM o SBF-SEM. El AT es una alternativa sencilla a un tem de sección serie tediosa donde la recolección y manipulación de secciones en serie en una oblea son más sencillas que con las rejillas de ranura. Se desarrollaron varias estrategias para facilitar la recopilación de las matrices, y compartimos nuestro método para expandir el conjunto de herramientas existente. En los casos en que la identificación del ROI es un desafío, AT-SEM proporciona una ventaja fundamental, con el cribado eficiente de las muestras donde se requiere una resolución a escala de orgánulos en 50 a 500 secciones. Para volúmenes más grandes, las estrategias de AT de recolección automática se pueden recopilar de manera eficiente si se requieren más secciones. Las muestras de AT se pueden volver a visualizar varias veces, lo que facilita la obtención de imágenes específicas de áreas de alta resolución basadas en imágenes generales previamente adquiridas. Creemos que el análisis dirigido y la reducción del sobremuestreo por at/SEM propuestos aquí disminuyen los requisitos de mano de obra y almacenamiento de datos. En última instancia, las bibliotecas de secciones se pueden recopilar y mantener para su posterior reutilización y consulta. Para la adquisición de volumen, los enfoques FIB o SBF-SEM ofrecen una excelente solución siempre que el ROI sea fácil de identificar en la cara del bloque o si se necesitan grandes volúmenes 3D para el análisis. Sin embargo, FIB / SBF-SEM son menos eficientes cuando la imagen de pila de alta resolución debe recopilarse de un ROI definido de manera específica. Para concluir, los métodos propuestos para el cribado de muestras de AT y el uso de imágenes generales de resolución media permiten limitar la adquisición de imágenes a las partes relevantes de la matriz de secciones. La orientación precisa de las regiones de imágenes acelera el tiempo de obtención de datos y simplifica la evaluación de los datos.
En resumen, aunque el concepto de AT/SEM no es novedoso, su uso todavía no está tan extendido como sus méritos sugieren. En general, proporciona un procedimiento complementario a otros métodos EM existentes. AT/SEM es compatible con la más amplia gama de protocolos de preparación de muestras y flujos de trabajo de imágenes y se puede realizar en cualquier microscopio FIB/SEM o SBF-SEM como técnica de acompañamiento. En este artículo, nos hemos concentrado en AT para registrar datos ultraestructurales de muestras que tienen menos probabilidades de ser abordadas con éxito por otros métodos. Esperamos que el procedimiento descrito para la recopilación conveniente de secciones y las estrategias de adquisición considerablemente automatizadas ayuden en los primeros intentos para aquellos que nunca han encontrado el método y ayuden a perfeccionarlo para aquellos que ya tienen algo de experiencia.
Divulgaciones
El autor Tilman Franke es un empleado de Thermo Fisher que fabrica microscopios electrónicos y programas de pilotaje que se utilizan en el artículo.
Agradecimientos
Queremos agradecer a los miembros de la instalación EM de la Universidad de Lausana por su apoyo durante este desarrollo de diferentes pasos del procedimiento AT. Nos gustaría agradecer a Gareth Griffiths, Marta Rodrigues, Urska Repnik, Christel Genoud, Helmut Gnaegi, Einat Zelinger, Paola Moreno-Roman, Lucy O'Brien y Lindsay Lewellyn por las discusiones durante la preparación del manuscrito y la lectura crítica. Queremos reconocer a los grupos que contribuyeron con las muestras utilizadas para demostrar los diferentes escenarios: Matthias Lutolf, Michail Nikolaev, Devanjali Dutta, Till Matzat y Fanny Langlet.
Materiales
| Name | Company | Catalog Number | Comments |
| Cutting | |||
| AT sectioning knife | Diatome | DUATS3530 | Diatome Jumbo knife |
| Diamond knife for trimming 90° | Diatome | DTB90 | Diatome trimming 20° Glass knife |
| Pattex contact adhesive | Pattex | PCL3C | |
| Silicon wafer | Ted Pella | 16015 | Resistance: 1-30 Ohms Type P: (Boron) (1 primary flat) Roughness: 2 nm No SiO2 top coating TTV: = <20 µm Wafer is polished on one side |
| Ultramicrotome | Leica UC6 | Alternative: Leica UC7 | |
| Wafer cleaving kit | EMS | 7642 | EMF, Small Sample Cleaver, CatNo. 7652 |
| Image acquisition | |||
| FESEM | Thermo Fischer Helios | 1072419 | Alternatives: Zeiss, Jeol, Hitachi, TESCAN |
| Maps 3 for SEM with Correlative Workflow & Array Tomography | Thermo Fisher Scientific | 1135932 | Maps provides automation of SEM imaging workflows and allows importing of 3rd party data for CLEM and navigation. |
| Image analysis | |||
| Amira x.y | Thermo Fisher Scientific | 1131599 | Amira is a 3D data visualization and analysis software with several practical functions for Array Tomography data reconstruction. |
| Image processing | Open source | Fiji (http://fiji.sc/#download) | IMOD, MIB (See text for refferences) |
Referencias
- Luckner, M., Wanner, G. From light microscopy to analytical scanning electron microscopy (SEM) and focused ion beam (FIB)/SEM in biology: Fixed coordinates, flat embedding, absolute references. Microscopy and Microanalysis. 24 (5), 526-544 (2018).
- White, J. G., Southgate, E., Thomson, J. N., Brenner, S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philosophical Transaction of Royal Society of London B Biological Sciences. 314 (1165), 1 (1986).
- Zheng, Z., et al. A complete electron microscopy volume of the brain of adult Drosophila melanogaster. Cell. 174 (3), 730-743 (2018).
- Mulcahy, B., et al. A pipeline for volume electron microscopy of the caenorhabditis elegans nervous system. Frontiers in Neural Circuits. 12, 94 (2018).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Baumeister, W., Grimm, R., Walz, J. Electron tomography of molecules and cells. Trends in Cell Biology. 9 (2), 81-85 (1999).
- Hoog, J. L., et al. Organization of interphase microtubules in fission yeast analyzed by electron tomography. Developmental Cell. 12 (3), 349-361 (2007).
- Weber, M. S., Wojtynek, M., Medalia, O. Cellular and structural studies of eukaryotic cells by cryo-electron tomography. Cells. 8 (1), 57 (2019).
- Micheva, K. D., Smith, S. J. Array tomography: a new tool for imaging the molecular architecture and ultrastructure of neural circuits. Neuron. 55 (1), 25-36 (2007).
- Horstmann, H., Korber, C., Satzler, K., Aydin, D., Kuner, T. Serial section scanning electron microscopy (S3EM) on silicon wafers for ultra-structural volume imaging of cells and tissues. PLoS One. 7 (4), 35172 (2012).
- Hayworth, K. J., et al. Imaging ATUM ultrathin section libraries with WaferMapper: a multi-scale approach to EM reconstruction of neural circuits. Frontiers in Neural Circuits. 8, 68 (2014).
- Kubota, Y., et al. A carbon nanotube tape for serial-section electron microscopy of brain ultrastructure. Nature Communication. 9 (1), 437 (2018).
- Wacker, I. U., et al. Multimodal Hierarchical Imaging of Serial Sections for Finding Specific Cellular Targets within Large Volumes. Journal of Visualized Experiments. (133), e57059 (2018).
- Burel, A., et al. A targeted 3D EM and correlative microscopy method using SEM array tomography. Development. 145 (12), (2018).
- Templier, T. MagC, magnetic collection of ultrathin sections for volumetric correlative light and electron microscopy. Elife. 8, 45696 (2019).
- Denk, W., Horstmann, H. Serial block-face scanning electron microscopy to reconstruct three-dimensional tissue nanostructure. PLoS Biology. 2 (11), 329 (2004).
- Knott, G., Marchman, H., Wall, D., Lich, B. Serial section scanning electron microscopy of adult brain tissue using focused ion beam milling. Journal of Neuroscience. 28 (12), 2959-2964 (2008).
- Wanner, A. A., Genoud, C., Masudi, T., Siksou, L., Friedrich, R. W. Dense EM-based reconstruction of the interglomerular projectome in the zebrafish olfactory bulb. Nature Neuroscience. 19 (6), 816-825 (2016).
- Smith, D., Starborg, T. Serial block face scanning electron microscopy in cell biology: Applications and technology. Tissue Cell. 57, 111-122 (2019).
- Kizilyaprak, C., Bittermann, A. G., Daraspe, J., Humbel, B. M. FIB-SEM tomography in biology. Methods in Molecular Biology. 1117, 541-558 (2014).
- Xu, C. S., et al. Enhanced FIB-SEM systems for large-volume 3D imaging. Elife. 6, 25916 (2017).
- Hayworth, K. J., et al. Gas cluster ion beam SEM for imaging of large tissue samples with 10 nm isotropic resolution. Nature Methods. 17 (1), 68-71 (2020).
- Maco, B., et al. Correlative in vivo 2 photon and focused ion beam scanning electron microscopy of cortical neurons. PLoS One. 8 (2), 57405 (2013).
- Russell, M. R., et al. 3D correlative light and electron microscopy of cultured cells using serial blockface scanning electron microscopy. Journal of Cell Science. 130 (1), 278-291 (2017).
- Lucas, M. S., Gunthert, M., Gasser, P., Lucas, F., Wepf, R. Bridging microscopes: 3D correlative light and scanning electron microscopy of complex biological structures. Methods in Cell Biology. 111, 325-356 (2012).
- Koga, D., Kusumi, S., Bochimoto, H., Watanabe, T., Ushiki, T. Correlative light and scanning electron microscopy for observing the three-dimensional ultrastructure of membranous cell organelles in relation to their molecular components. Journal of Histochemistry and Cytochemistry. 63 (12), 968-979 (2015).
- Peddie, C. J., et al. Correlative and integrated light and electron microscopy of in-resin GFP fluorescence, used to localise diacylglycerol in mammalian cells. Ultramicroscopy. 143, 3-14 (2014).
- Markert, S. M., et al. 3D subcellular localization with superresolution array tomography on ultrathin sections of various species. Methods in Cell Biology. 140, 21-47 (2017).
- Kolotuev, I., Schwab, Y., Labouesse, M. A precise and rapid mapping protocol for correlative light and electron microscopy of small invertebrate organisms. Biology of the Cell. 102 (2), 121-132 (2009).
- Kolotuev, I. Positional correlative anatomy of invertebrate model organisms increases efficiency of TEM data production. Microscopy and Microanalysis. 20 (5), 1392-1403 (2014).
- Kato, M., Kolotuev, I., Cunha, A., Gharib, S., Sternberg, P. W. Extrasynaptic acetylcholine signaling through a muscarinic receptor regulates cell migration. Proceedings of the National Academy of Sciences. 118 (1), e1904338118 (2021).
- Schindelin, J., et al. Fiji: An open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Miguel-Aliaga, I., Jasper, H., Lemaitre, B. Anatomy and physiology of the digestive tract of Drosophila melanogaster. Genetics. 210 (2), 357-396 (2018).
- Nikolaev, M., et al. Homeostatic mini-intestines through scaffold-guided organoid morphogenesis. Nature. 585 (7826), 574-578 (2020).
- Knoblich, J. A. Mechanisms of asymmetric stem cell division. Cell. 132 (4), 583-597 (2008).
- Daniel, E., et al. Coordination of septate junctions assembly and completion of cytokinesis in proliferative epithelial tissues. Current Biology. 28 (9), 1380-1391 (2018).
- Pasquettaz, R., et al. Peculiar protrusions along tanycyte processes face diverse neural and non-neural cell types in the hypothalamic parenchyma. Journal of Comparative Neurology. , (2020).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Cardona, A., et al. TrakEM2 software for neural circuit reconstruction. PLoS One. 7 (6), 38011 (2012).
- Belevich, I., Joensuu, M., Kumar, D., Vihinen, H., Jokitalo, E. Microscopy image browser: A platform for segmentation and analysis of multidimensional datasets. PLoS Biology. 14 (1), 1002340 (2016).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados