
Method Article
Preparación de laminillas a partir de muestras biológicas vítreas utilizando un microscopio electrónico de barrido de doble haz para la criotomografía electrónica
En este artículo
Erratum Notice
Resumen
Uso de molienda de haz de iones enfocado para producir láminas vítreas en la rejilla a partir de muestras biológicas congeladas por inmersión para tomografía crioelectrónica.
Resumen
Aquí se presenta un protocolo para preparar criolaminillas a partir de rejillas congeladas por inmersión de eritrocitos humanos infectados con Plasmodium falciparum, que podrían adaptarse fácilmente para otras muestras biológicas. Los principios básicos para preparar muestras, fresar y ver laminillas son comunes a todos los instrumentos y el protocolo se puede seguir como una guía general para la preparación de criolaminillas en red para microscopía crioelectrónica (cryoEM) y tomografía crioelectrónica (cryoET). Las rejillas de microscopía electrónica que soportan las células se congelan por inmersión en etano líquido refrigerado por nitrógeno líquido utilizando un congelador de inmersión manual o automatizado, luego se criban en un microscopio óptico equipado con una etapa criogénica. Las rejillas congeladas se transfieren a un microscopio electrónico de criobarrido equipado con un haz de iones enfocado (cryoFIB-SEM). Las rejillas se recubren rutinariamente por pulverización catódica antes del fresado, lo que ayuda a la dispersión de la acumulación de carga durante el fresado. Alternativamente, se puede usar una recubridora rotativa de viga electrónica para aplicar una capa de carbono-platino a las rejillas, cuyo grosor exacto se puede controlar con mayor precisión. Una vez dentro del crioFIB-SEM, se aplica un recubrimiento adicional de un compuesto organoplatino a la superficie de la rejilla a través de un sistema de inyección de gas (GIS). Esta capa protege el borde frontal de la lámina a medida que se fresa, cuya integridad es fundamental para lograr laminillas uniformemente delgadas. Las regiones de interés se identifican a través de SEM y el fresado se lleva a cabo de manera gradual, reduciendo la corriente del haz de iones a medida que la lámina alcanza la transparencia de electrones, para evitar la generación excesiva de calor. Una rejilla con múltiples laminillas se transfiere a un microscopio electrónico de transmisión (TEM) bajo condiciones criogénicas para la adquisición en serie de inclinación. Un flujo de trabajo robusto y libre de contaminación para la preparación de láminas es un paso esencial para las técnicas posteriores, incluida la crioEM celular, la crioET y el promedio de subtomograma. El desarrollo de estas técnicas, especialmente para la extracción y molienda de muestras congeladas a alta presión, es de alta prioridad en el campo.
Introducción
Solo el contenido celular de muestras biológicas de <500 nm de espesor puede ser fotografiado de manera efectiva por microscopía electrónica de transmisión (TEM) a temperaturas criogénicas, limitando el rango de especímenes a virus, procariotas, organismos unicelulares simples y regiones más delgadas de células eucariotas más grandes1. El molienda por haz de iones enfocado en la red (FIB) permite que las muestras biológicas congeladas por inmersión más gruesas se adelgazen a láminas transparentes con electrones a temperaturas criogénicas (< -150 °C). Las laminillas resultantes se transfieren a un TEM para la visualización y la recopilación de datos tomográficos, lo que permite reconstrucciones 3D de alta resolución de las características celulares y moleculares dentro de las células (para revisiones, consulte Rigort et al., 20122, Oikonomou et al., 20163, y Wagner et al., 20204).
El fresado FIB surgió del campo de las ciencias de los materiales, donde las muestras se diluyen rutinariamente para prepararlas para el análisis posterior5. Se lleva a cabo en un microscopio electrónico de barrido (SEM), que tiene dos columnas ópticas: óptica de microscopio electrónico de barrido convencional y una segunda columna que contiene óptica capaz de generar y controlar finamente un haz de iones enfocado (FIB), llamado FIB-SEM. Esto permite que una región específica de la muestra sea ablacionada por iones generados por una fuente de galio, eliminando el exceso de material y dejando una lámina6. El proceso de fresado se guía por imágenes SEM de la topografía de la muestra, que se utiliza para localizar regiones de interés y monitorear el progreso del fresado. Para aplicaciones biológicas, la configuración básica es en gran medida la misma, pero la molienda se lleva a cabo a temperaturas criogénicas. Esto ha requerido que los FIB-SEM estándar se adapten para tener etapas crioenfriadas que mantengan temperaturas constantes y bajas tasas de contaminación superficial, así como esclusas de aire para facilitar la transferencia de muestras sin desvitrificación o contaminación. Los transbordadores de muestra también se han modificado para permitir que se monte una gama de portadores diferentes dentro de la crioFIB-SEM, incluidas rejillas TEM, planchetas y capilares. Varios grupos clave de investigadores han sido fundamentales para el desarrollo de estos métodos y los continuos avances tecnológicos en esta área 7,8,9,10,11,12. Las soluciones comerciales están ahora más ampliamente disponibles para el fresado biológico FIB a temperatura criogénica y el fresado en la red de laminillas se está volviendo más rutinario, dada una muestra optimizada.
Se puede utilizar una gama de técnicas de temperatura ambiente y crioEM para visualizar información celular en todas las escalas de la vida, desde organismos multicelulares completos a una resolución modesta hasta la comprensión del contexto de procesos complejos a nivel celular y con mayor detalle, hasta la determinación de in situ Estructuras moleculares13,14,15,16,17,18,19. Las técnicas clásicas a temperatura ambiente incluyen la seccionación fija y teñida, de células y tejidos incrustados en resina por ultramicrotomía para el análisis de la morfología celular por TEM (para revisión, véase Studer et al., 200820). Se han desarrollado técnicas alternativas que explotan la dispersión secundaria de electrones mediante SEM para obtener imágenes de la superficie de bloques de células preservadas, antes de eliminar progresivamente el material, ya sea con un cuchillo (imagen de cara de bloque en serie) o un haz de iones enfocado.21,22,23. Esta técnica también se ha implementado con éxito a temperaturas criogénicas (denominadas imágenes de criovolumen) con un crioFIB-SEM en bloques vítreos de células o tejidos sin teñir.24. Alternativamente, las laminillas más gruesas (~ 15 μm de espesor) se pueden fresar y estudiar mediante imágenes STEM.25. Usando estas técnicas, se pueden obtener imágenes de bloques enteros que contienen muchas células para recopilar información de la población o se puede obtener una imagen de un órgano / organismo completo y reconstruirlo en 3D. Sin embargo, para acceder a la información molecular de alta resolución de las células, las muestras deben conservarse en un estado casi nativo, congelado e hidratado y, por lo tanto, deben prepararse en condiciones criogénicas. La criomicroscopía electrónica de secciones vítreas (CEMOVIS) es una técnica mediante la cual se seccionan bloques congelados de material biológico a alta presión en condiciones criogénicas con un ultramicrótomo. Esto produce cintas de criosecciones (40-100 nm de espesor)26, que se adjuntan a una cuadrícula EM y se visualizan en el TEM. Sin embargo, la interacción física del cuchillo con la muestra vítrea causa grietas y artefactos de compresión que pueden distorsionar gravemente la estructura celular.27,28,29,30. Las secciones más gruesas son más propensas a estos artefactos, por lo que no es práctico usar secciones más gruesas que ~ 70 nm26. Esta limitación restringe en gran medida la vista 3D de la estructura biológica en el tomograma. La molienda FIB a temperaturas criogénicas no experimenta estos problemas, pero tiene sus propios artefactos causados por las tasas de molienda diferenciales en partes de la muestra, lo que lleva a espesores variables dentro de una lámina, llamada cortina. Este problema se mitiga mediante la aplicación de una capa organoplatino aplicada a través de un sistema de inyección de gas (GIS), que protege el borde frontal de la lámina durante la molienda31. El límite superior del espesor de la muestra para el fresado FIB en la red se define por la capacidad de congelar la muestra mientras se mantiene vítrea.32, aunque, con la introducción de técnicas de elevación criogénica y portadores de muestras adaptados para muestras biológicas, la molienda FIB también se puede utilizar para procesar muestras congeladas a alta presión31,33,34,35. Además, las muestras congeladas por inmersión no pueden ser demasiado delgadas, ya que debe haber suficiente material biológico para generar una lámina de tamaño razonable que proporcione suficiente área de superficie para recolectar series de inclinación con el aumento requerido. Este problema puede aliviarse moliendo grupos de células más pequeñas, como bacterias o levaduras. El espesor final de la lámina (~ 100-300 nm) generalmente está dictado por la integridad de la muestra y la estrategia de fresado. Las laminillas más delgadas son mejores para trabajos estructurales de alta resolución, como el promedio de subtomograma, pero las laminillas más gruesas contienen volúmenes celulares mucho más grandes que los que se pueden lograr con CEMOVIS, proporcionando más contexto celular en una muestra casi conservada de forma nativa. La molienda FIB también se puede utilizar para diluir cristales de proteína congelados por inmersión para estudios de difracción de electrones36.
La molienda FIB de células vítreas vale la pena el tiempo y el esfuerzo para llevar a cabo si la cuestión científica requiere detalles moleculares de alta resolución de especímenes casi nativos in situ. Con acceso a más instalaciones para la producción rutinaria de laminillas, el paso limitante de la velocidad es a menudo la optimización de la muestra antes de la molienda, donde se debe tomar tiempo para garantizar que la muestra sea vítrea y un grosor adecuado para producir laminillas robustas y uniformemente delgadas. Aquí se describe la optimización de la muestra para los glóbulos rojos humanos congelados por inmersión de molienda FIB infectados con parásitos Plasmodium falciparum , el agente causante de la malaria, pero este enfoque podría adaptarse para cualquier muestra dada.
Protocolo
La sangre humana se obtuvo de donantes anónimos a través del Servicio Nacional de Sangre y Trasplante del Reino Unido y se utiliza dentro de las 2 semanas posteriores a la recepción. No se requiere aprobación ética para su uso.
1. Preparación y congelación por inmersión de glóbulos rojos infectados con Plasmodium falciparum
- Aislar esquizontes maduros por centrifugación (1.580 x g) sobre un medio de gradiente de densidad isotónica al 70% (v/v) (para procedimientos estándar sobre cómo cultivar estadios sanguíneos asexuales de 3D7 Plasmodium falciparum en eritrocitos humanos ver Blackman M.J., 199537).
- Fijar las películas de sangre fina secadas al aire en un portaobjetos de vidrio con metanol al 100% para comprobar la homogeneidad morfológica de los esquizontes antes de teñir con tinción de Giemsa al 10% en tampón fosfato de 6,7 mM, pH 7,1.
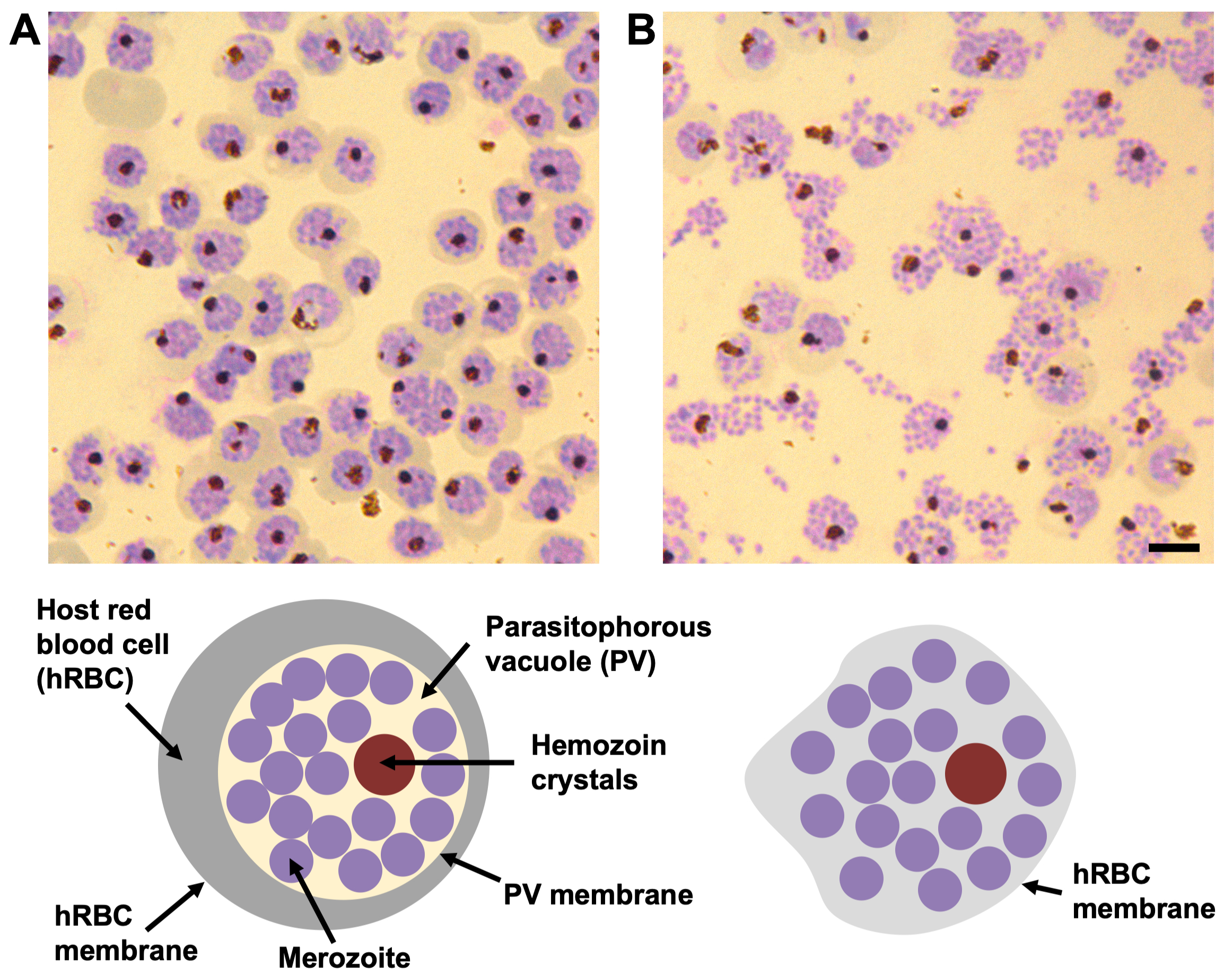
NOTA: Para enriquecer la preparación para esquizontes estancados en puntos específicos de salida, los esquizontes pueden sincronizarse aún más con los inhibidores compuestos 2 y E64 (ver Resultados representativos y Figura 1 para una explicación del efecto de estos inhibidores).
PRECAUCIÓN: Plasmodium falciparum es un patógeno humano y solo debe manipularse en una instalación de contención adecuada siguiendo las pautas locales de salud y seguridad. - Centrifugar suavemente los esquizontes (240 x g) para granularlos y resuspender en 2x volumen el pellet celular del medio RPMI, lo que resulta en una suspensión de hematocrito al 50%.
- En una plataforma de congelación por inmersión manual, aplique 2.5 μL de esquizontes al lado de carbono de una rejilla de cobre de malla incandescente (use configuraciones de unidad de descarga incandescente de 60 s, 30 mA en el aire, tratando solo el lado de carbono de la rejilla) Rejilla de cobre de malla 200 con una película de carbono de 2/4 agujereada y borre para ~ 20 s desde la parte posterior de la rejilla usando papel de filtro de grado 1 con un borde rasgado. Sumérjase en el etano líquido refrigerado por nitrógeno líquido y transfiera las rejillas al almacenamiento (consulte la Tabla de materiales para conocer el equipo utilizado en este estudio).
NOTA: El experimento se puede pausar aquí, y las rejillas se pueden almacenar bajo nitrógeno líquido indefinidamente.
PRECAUCIÓN: El nitrógeno líquido es un asfixiante y causa congelación; Manejar con cuidado en un ambiente adecuado con monitoreo de oxígeno.
PRECAUCIÓN: El etano líquido causa quemaduras graves y es inflamable; Úselo en una campana extractora lejos de fuentes de ignición.
NOTA: Los esquizontes congelados de inmersión ya no son viables. Esto se determinó incubando sangre humana con varias rejillas doradas de esquizontes congelados por inmersión descongelados por aire y no observando el crecimiento de parásitos después de varios días, en comparación con los controles no congelados. Por lo tanto, las rejillas congeladas de esquizontes son seguras de manejar fuera de las instalaciones de contención utilizando procedimientos normales de seguridad y descontaminación (guantes, esterilización de superficies/herramientas con etanol >70% y eliminación de rejillas en etanol >70%). - Rejillas de cribado utilizando una etapa criogénica para un microscopio óptico, prestando especial atención al gradiente de hielo a través de la rejilla. En las áreas más delgadas de hielo, verifique los cuadrados de cuadrícula individuales para la cobertura de la celda.
NOTA: Los mejores cuadrados deben tener una celda de espesor en el centro del cuadrado de cuadrícula (Figura 2). Esto asegura que una lámina se pueda fresar a través del centro del cuadrado sin golpear el hielo más grueso en los bordes contra las barras de rejilla. El experimento se puede pausar aquí, y las rejillas se pueden almacenar bajo nitrógeno líquido indefinidamente. Si las células llevan un marcador fluorescente, las rejillas también pueden ser examinadas por cryoCLEM para localizar posiciones de interés X / Y, que se pueden correlacionar con ubicaciones de rejilla en el crioFIB-SEM para dirigir el fresado.
2. Fresado FIB en red de células congeladas por inmersión
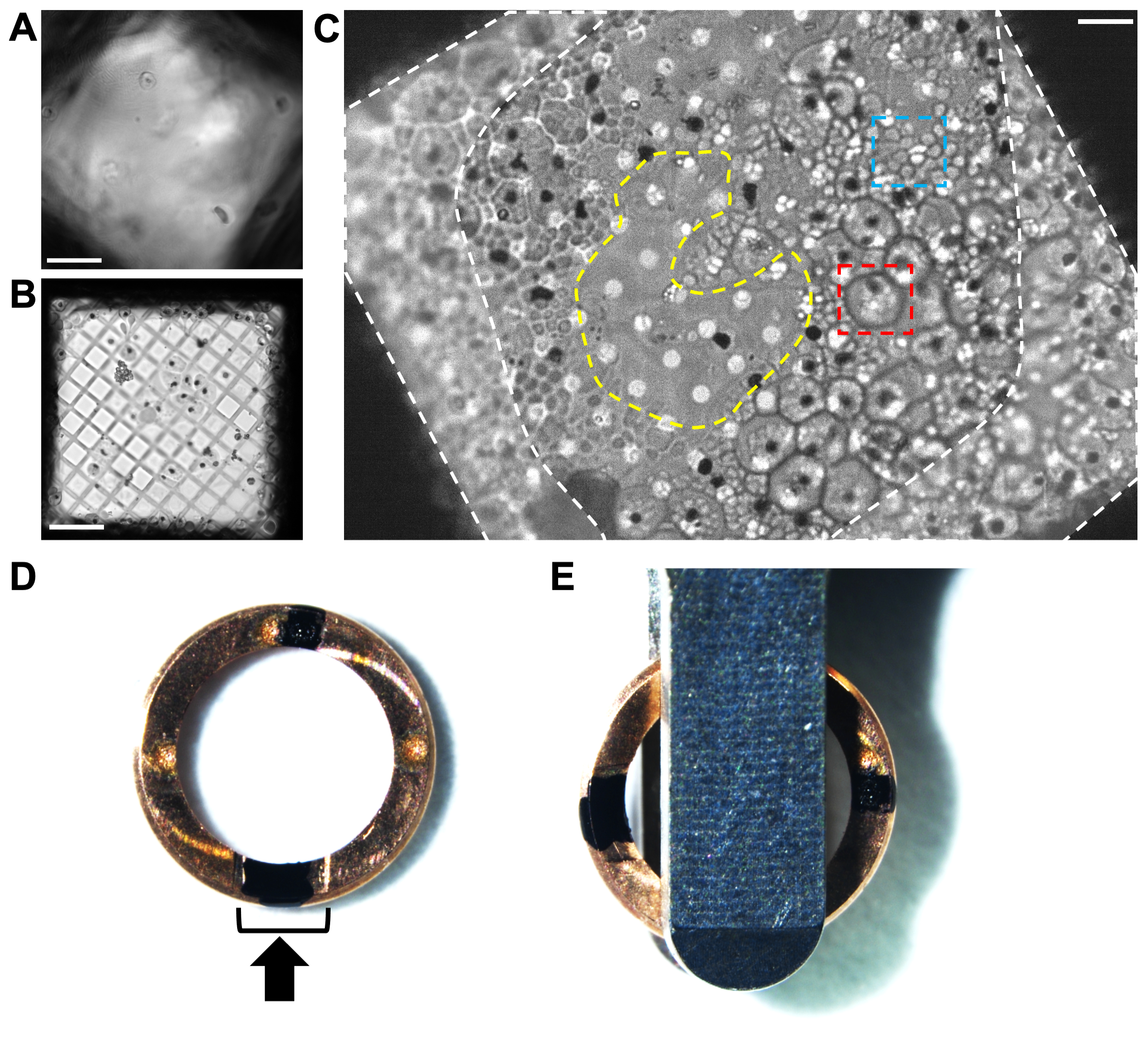
- Marque la parte frontal de las llantas de autorejilla específicas de crioFIB con un rotulador indeleble negro para indicar el centro de la sección de corte y el lado opuesto de la llanta (Figura 3). Configure la estación de recorte y las rejillas de clip en los anillos marcados con el lado de carbono hacia abajo.
- Cargue las rejillas en la lanzadera cryoFIB-SEM (típicamente 2 rejillas, dependiendo de la lanzadera) con el lado del carbono hacia arriba y aplique una capa de pulverización catódica de platino en una atmósfera de argón (5 x 10-2 mbar) (5 mA durante 60 s - el espesor es variable) o una capa rotativa de haz electrónico de carbono / platino (~ 4 nm de espesor) a la superficie de las celdas.
NOTA: Ambos tipos de recubrimientos ayudan a la dispersión de la carga durante las imágenes SEM. El beneficio del recubridor rotativo de viga electrónica es que se puede especificar el grosor exacto del recubrimiento. - Cargue la lanzadera en el crioFIB-SEM y evalúe la distribución celular en cada red mediante SEM a 5 kV (13 pA o 25 pA). Tome una visión general de bajo aumento (~ 100x) para ver los gradientes de hielo en la cuadrícula. Luego, tome imágenes de mayor aumento (~ 5,000x) para mirar los cuadrados de la cuadrícula individuales e identificar las áreas de la cuadrícula con características celulares visibles y baja contaminación superficial para moler.
- Aplique una capa organoplatino de >2 μm a la superficie de cada rejilla utilizando el sistema de inyección de gas (GIS). Para hacer esto, inserte la aguja GIS en la cámara sobre la rejilla y caliente la fuente de organoplatino a una temperatura establecida durante un tiempo establecido (por ejemplo, ~ 27 ° C durante 3-10 s) para producir un flujo de vapor.
NOTA: El ángulo de aplicación, la temperatura y el tiempo de la capa organoplatino a través de la aguja GIS deben optimizarse para maximizar un recubrimiento uniforme. Esto dependerá de la crioFIB-SEM específica utilizada (ver Resultados representativos para una explicación más detallada). - Incline la muestra de modo que el plano de la cuadrícula sea ~10° desde el ángulo de incidencia del haz de iones y mueva el centro de un cuadrícula cuadrado adecuado para ser visto en las imágenes SEM y FIB.
- Examine la cuadrícula utilizando el haz de iones a baja corriente (nominalmente 1.5 pA, 30 kV) y vaya a un aumento lo suficientemente alto (~ 7,000x) para visualizar las celdas en el centro de un cuadrado de cuadrícula. Enfocar la muestra a la primera corriente de fresado (300 pA) y corregir el astigmatismo. Ajuste el brillo y el contraste, luego marque dos patrones rectangulares para fresar, uno por encima y otro por debajo de una región protegida de 3 μm de espesor, cuyo centro es la ubicación deseada de la lámina final.
NOTA: El ancho elegido de los patrones dependerá de la topografía de la capa de celda circundante y del tamaño de la celda. 7-20 μm es un ancho adecuado para esquizontes, pero las laminillas más anchas tardan más en fresarse. La altura elegida de los patrones para el primer paso de fresado depende del grosor de la muestra, comenzando alrededor de 6 μm; Esto puede necesitar ser ajustado durante el fresado. El fresado se lleva a cabo direccionalmente desde los bordes superior e inferior exterior de los patrones hacia la cara de la lámina. Esto se puede hacer en paralelo, donde ambos patrones se muelen concurrente o secuencialmente, primero eliminando material por encima de la lámina y luego por debajo. - Comience a fresar a la primera corriente; monitorizar el progreso en directo en la vista del haz de iones y de forma intermitente mediante SEM (5 kV, 13 o 25 pA). Compruebe que el haz de iones ha atravesado la muestra por encima y por debajo de la región protegida. Si no, aumente la altura de los patrones rectangulares para eliminar más material. Deténgase cuando la superficie por encima y por debajo de la lámina esté completamente lisa en la vista del haz de iones.
NOTA: El haz de iones ha atravesado la muestra por encima y por debajo de la región protegida cuando el interior de los patrones rectangulares no contiene características en el plano focal. Algunas entidades, como las barras de cuadrícula, pueden ser visibles fuera de foco en segundo plano. - Cambio a la siguiente corriente de fresado (100 pA); Enfoca y ajusta el brillo/contraste. Disminuya el espacio entre los dos patrones rectangulares a 1,5 μm y disminuya la altura del patrón para cubrir solo el material sin fresar. Comience a fresar a la nueva corriente hasta que la superficie por encima y por debajo de la lámina esté completamente lisa.
- Repita este proceso paso a paso hasta alcanzar un espesor de 0,3 μm, reduciendo la corriente del haz de iones cada vez de acuerdo con el esquema de fresado de la Tabla 1.
- Moler varias laminillas (la cantidad depende del espesor de la muestra y del tiempo disponible) en una o ambas rejillas hasta un espesor de 0,3 μm, registrar la posición X/Y/Z de cada lámina y reservar ~1-2 h al final de la sesión para pulir las laminillas hasta su espesor final (60-200 nm).
- Tome una visión general SEM de bajo aumento de toda la cuadrícula y planifique una ruta de pulido comenzando desde las laminillas en la parte frontal de la cuadrícula (más cerca de la fuente del haz de iones) hasta las laminillas en la parte posterior de la rejilla (más alejadas de la fuente del haz de iones) (Figura 5).
NOTA: Esto reduce la redeposición del material ablacionado de nuevo en la superficie de las laminillas pulidas. - Reduzca el espacio entre los dos patrones rectangulares a 100-200 nm y comience el paso final de pulido a una corriente de haz de iones de 30 pA. Monitoree el progreso por SEM a 2-3 kV (13 pA, permanencia = 300 n, 3072 x 2048, ~ 2 s para un fotograma completo) y detenga el pulido cuando el contraste se pierde en la lámina por SEM o cuando la capa organoplatino de la lámina comienza a perder integridad.
- Antes de quitar las rejillas, adquiera una imagen SEM de bajo aumento de toda la cuadrícula y guarde las imágenes de cada lámina. Utilícelos para verificar la cuadrícula más adelante en el TEM. Haga una pausa en el experimento aquí y almacene las rejillas con laminillas bajo nitrógeno líquido, manéjelas con cuidado.
NOTA: Las rejillas pueden estar ligeramente recubiertas por pulverización catódica al retirarlas del crioFIB-SEM, lo que puede ayudar a limitar la deriva y la carga en el TEM a gran aumento, pero esto debe hacerse con precaución ya que demasiado recubrimiento por pulverización catódica puede oscurecer el contenido biológico dentro de las láminas. Es posible examinar las laminillas para detectar fluorescencia en esta etapa; Sin embargo, obtener suficiente señal dependerá de la abundancia de la proteína marcada dentro del grosor de la lámina. Se debe tener mucho cuidado al manipular las rejillas para limitar el daño a las láminas y evitar la contaminación de la superficie.
3. Adquisición de la serie Tilt y visión general del procesamiento de datos
- Cargue las rejillas en el TEM, alineando la dirección de fresado perpendicular al eje de inclinación del plato.
NOTA: La alineación de las rejillas se realiza a simple vista utilizando las marcas en el borde de la rejilla automática. Un margen de error de ~ 10 ° es aceptable, de lo contrario, las paredes de la zanja a ambos lados de la lámina pueden oscurecer la lámina a medida que la rejilla se inclina. - Adquiera un mapa de bajo aumento (~ 150x) de toda la cuadrícula y localice las laminillas; Luego, adquiera un mapa de aumento medio (~ 1,500x, dependiendo del tamaño de la lámina) de cada lámina y localice las áreas de interés.
- Incline previamente la rejilla ±10° para que el plano de la lámina (no la rejilla) sea perpendicular al eje óptico.
NOTA: La dirección de la pre-inclinación se puede determinar por la posición del borde frontal de las láminas (buscando la capa de organoplatino sobrante) en los mapas de aumento bajo y medio. Para el TEM utilizado aquí, las cuadrículas requieren una preinclinación de +10° si los bordes frontales de las laminillas apuntan hacia arriba en los mapas y requieren una preinclinación de -10° si apuntan hacia abajo. Cada rejilla puede ser diferente debido a cómo fueron recogidas e insertadas en el cargador automático. - Adquiera la serie de inclinación simétrica de dosis38 (por ejemplo, -54° a +54° con un incremento de 3-5°) a un tamaño de píxel que permita tanto el campo de visión como la resolución requeridos para la región de interés. Utilice un rango de valores de desenfoque entre -2 y -5 μm. Recopile películas con 3-10 fotogramas en cada incremento y, para ello, ajuste estos parámetros en función del tamaño del píxel para acumular una dosis total de ~150 e-/Å2 (para un TEM de 300 kV).
NOTA: Se deben evitar las grietas en la lámina, ya que estas regiones pueden derivarse. También debe evitarse la contaminación de la superficie, ya que puede oscurecer la región de interés o el área de enfoque en alta inclinación. - Motion corrige las películas usando un programa como MotionCor239. Aplique un filtro de ponderación de dosis a las imágenes corregidas (e-/imagen acumulada)40 y estime el desenfoque de cada imagen utilizando un programa como CTFFIND441.
- Utilice la alineación sin fiducial (seguimiento de parches) en un programa como etomo (IMOD)42 para calcular la alineación y la relación angular de las imágenes en la serie de inclinación.
- Ingrese la información de alineación y rotación, junto con los valores de desenfoque en un programa que pueda aplicar corrección CTF tridimensional, por ejemplo, NovaCTF43. Calcule una tomografía corregida para obtener tomogramas de salida con factores de agrupación relevantes para el análisis posterior.
- Analice las reconstrucciones y prepárese para cualquier procesamiento posterior, por ejemplo, filtrado, segmentación o promedio de subtogramas.
Resultados
Preparación de esquizontes de P. falciparum para la congelación por inmersión
Los inhibidores compuestos 2 y E64 se utilizan para detener los esquizontes en diferentes etapas de salida, generando una población enriquecida de esquizontes para su posterior estudio. Esto es importante porque sin una técnica correlativa complementaria, fresar objetivos subcelulares específicos o tipos de células es un desafío, ya que el proceso es esencialmente ciego. El compuesto 2 es un inhibidor de la proteína quinasa que detiene la salida antes de la ruptura de la vacuola. Los esquizontes pueden sincronizarse en el compuesto 2 durante 4 h, luego lavarse con medios libres de compuesto 2 para eliminar el inhibidor, momento en el que los esquizontes madurarán y saldrán después de aproximadamente 30 minutos. Alternativamente, los esquizontes sincronizados del compuesto 2 pueden lavarse en E64, un inhibidor irreversible de la cisteína proteasa de amplio espectro, e incubarse durante aproximadamente 1 h para detener la salida después del punto de ruptura de la vacuola, pero antes de la ruptura de la célula huésped. La morfología y homogeneidad de los esquizontes tratados deben verificarse mediante frotis de sangre teñidos con Giemsa antes de la congelación por inmersión (Figura 1). Los esquizontes se pueden congelar por inmersión en cualquiera de estos estados utilizando el método descrito en esta publicación.

Figura 1: La morfología de los esquizontes P. falciparum del compuesto 2 y E64 por frotis de sangre teñidos con Giemsa. (A) En presencia del compuesto 2, la vacuola parasitófora (PV) está densamente empaquetada con merozoítos (círculos púrpuras) con un solo grupo de cristales de hemozoína (círculo marrón oscuro). El límite entre la PV y la hemoglobina circundante en el glóbulo rojo del huésped (hRBC) (banda gris) está bien definido, así como la membrana hRBC. (B) En presencia de E64, la membrana fotovoltaica se rompe y los merozoítos se extienden dentro del hRBC. Cada schizont todavía contiene un solo grupo de cristales de hemozoína. La membrana de la célula huésped tiene fugas y está parcialmente colapsada, por lo tanto, no hay hemoglobina visible dentro de la periferia celular y el límite de la membrana hRBC no es fácilmente visible. Barra de escala, 5 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Optimización de la congelación por inmersión
Se probó una gama de rejillas con diferentes tamaños de orificios durante la optimización de las condiciones de transferencia para los glóbulos rojos infectados con P. falciparum, incluidos 2/2, 3/3, 3.5/1 y 5/2 (cuadrado) de carbono agujereado en rejillas de malla 200 de oro y cobre. Las rejillas de búsqueda de cobre de malla 200 con película de carbono agujereada de 2/4 proporcionan una capa de células adecuadamente gruesa para fresar láminas vítreas largas. Los agujeros más grandes o más pequeños generalmente resultaron en capas celulares que eran demasiado delgadas o gruesas, respectivamente (Figura 2A-C). Con 2/4 de carbono agujereado, los esquizontes se atraen a través de los orificios de 2 μm borrando la rejilla desde la parte posterior (lado sin carbono), lo que hace que las células sobresalgan por encima y por debajo de la película de carbono. El tramo de carbono de 4 μm entre los agujeros da como resultado una tira de carbono que atraviesa el centro de la mayoría de las laminillas resultantes, agregando resistencia. Las rejillas del buscador son las más adecuadas para fines correlativos y de cribado44, pero se debe tener cuidado para asegurarse de que los números / letras en el diseño de malla no sean demasiado grandes, ya que esto bloqueará las áreas para el fresado.
El tiempo de secado en un émbolo manual es de aproximadamente 20 s, pero el punto exacto en el que detener la secado se juzgó a simple vista cuando la gota de líquido que se extraía de la rejilla dejó de extenderse sobre el papel de filtro. Se requirió un borde rasgado para romper la tensión superficial de la gota para iniciar el proceso de secado. En este estudio no se utilizó un congelador automático, pero un punto de partida razonable para esta muestra sería utilizar el mismo volumen de celdas y tipo de rejilla que se utiliza para un émbolo manual, asegurándose de secar las rejillas desde la parte posterior en condiciones con alta humedad (~ 70%) y temperatura ambiente (~ 25 ° C). Los tiempos y condiciones de transferencia tendrían que optimizarse para el sistema automatizado particular utilizado.
Las rejillas congeladas de schizontes de P. falciparum se examinaron con un microscopio óptico equipado con una etapa criogénica en lugar de TEM, porque la muestra no era transmisible a los electrones. Para muestras más delgadas, las cuadrículas pueden ser examinadas por TEM (atlas de cuadrícula completa con un aumento de ~ 150x) antes de la transferencia de muestras al crioFIB-SEM, que puede ser un requisito previo para acceder a una instalación nacional. Se debe prestar especial atención al gradiente del espesor del hielo a través de la cuadrícula y también dentro de los cuadrados de la cuadrícula individuales. Los buenos cuadrados de cuadrícula deben tener una celda de espesor o golpear la película de carbono en su centro (Figura 2C). Esto evita el fresado en el hielo más grueso contra las barras de rejilla alrededor del borde del cuadrado de la rejilla y asegurará que el haz de iones se rompa por encima y por debajo de la capa de celda, produciendo una lámina libre en lugar de una cuña. Una vez que se han optimizado las condiciones reproducibles de secado y congelación por inmersión, el cribado ya no suele ser necesario antes del fresado FIB.

Figura 2. Análisis de la distribución celular en rejillas de escischontes congelados de P. falciparum por microscopía crio-óptica. (A) Un ejemplo de hielo demasiado grueso a través de un cuadrado de cuadrícula por microscopía de luz criogénica, oscureciendo las células y la película de carbono. Barra de escala, 10 μm. (B) Un ejemplo del tamaño del agujero (rejilla de cobre de malla 300 con película de carbono de 5/2 agujeros cuadrados) que es demasiado grande para las células, lo que resulta en una capa muy delgada de material biológico rodeada de regiones vacías sin hielo. Rejillas como esta producen laminillas muy cortas e inestables. Barra de escala, 10 μm. (C) Un ejemplo de buena distribución celular en una rejilla de cobre de malla 200 con película de carbono agujereada de 2/4. Estos esquizontes fueron tratados con el inhibidor de E64. Las células grandes (caja roja, ~ 5 μm de diámetro) con una periferia bien definida son glóbulos rojos infectados que todavía tienen una membrana de vacuola intacta. Los grupos de células pequeñas (caja azul, ~ 1 μm) son los merozoítos individuales contenidos dentro de un glóbulo rojo huésped parcialmente colapsado. Cada esquizonte tiene una mancha negra en su centro, que indica la posición de los cristales de hemozoína (Ver Figura 1 para detalles adicionales). La diferencia en la morfología celular no es tan fácil de ver una vez dentro del crioFIB-SEM; Por lo tanto, el precribado mediante microscopía crio-óptica es beneficioso hasta que se hayan alcanzado condiciones de transferencia reproducibles. La cobertura de celdas alrededor de los bordes del cuadrado de cuadrícula junto a las barras de cuadrícula (áreas discontinuas blancas) es demasiado gruesa para el fresado. La región más delgada en el centro del cuadrado de la cuadrícula (área discontinua amarilla) es un lugar ideal para fresar, extendiendo la lámina hacia la capa circundante de células. Barra de escala, 6 μm. (D) Imagen de una llanta de autorrejilla específica de crioFIB con dos marcas negras, una dentro de la sección de corte (soporte negro) y la otra opuesta, a las 12 en punto y a las 6 en punto. La flecha negra representa la dirección de fresado. (E) Cuando las rejillas se cargan en el casete del cargador automático, las marcas deben estar equidistantes a ambos lados de las pinzas de carga, lo que hace que las laminillas se encuentren perpendiculares al eje de inclinación de la plata. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Marcado de llantas de autorejilla específicas de crioFIB para tomografía
Por lo general, las rejillas se enganchan en las llantas de la rejilla automática antes del fresado para facilitar el manejo y proporcionar rigidez, lo que protege las láminas de daños durante los pasos de transferencia posteriores. Las llantas de autogrid específicas de CryoFIB se han diseñado con una función de corte para ayudar a acceder a más de la cara de la rejilla durante el fresado. Es importante orientar la dirección de fresado perpendicular al eje de inclinación del TEM para que la adquisición en serie de inclinación proceda girando la lámina a lo largo de su longitud. Esto asegura que las altas paredes de la zanja que rodea la lámina no oscurezcan la información biológica a medida que la rejilla está inclinada.
Por lo general, el borde de la rejilla automática está marcado para ayudar con la alineación visual dentro de la lanzadera cryoFIB-SEM y más tarde al cargar el TEM. Para estas muestras, se aplicaron dos marcas a las 12 en punto y a las 6 en punto con un marcador indeleble (ver Tabla de materiales), una dentro del centro de la sección de corte del anillo de clip y la segunda directamente opuesta (Figura 2D). Al cargar en el TEM (consulte la Tabla de materiales), ambas marcas deben ser visibles a ambos lados de las pinzas de carga y alineadas 90° con el borde de las pinzas (Figura 2E). Cabe señalar que el fabricante recomienda alinear las rejillas utilizando los puntos grabados en las llantas de la red automática, ya que ciertas tintas cercanas al haz de iones pueden interferir con el fresado.
Las rejillas recortadas se pueden cribar mediante microscopía óptica con un casete modificado para una etapa criogénica, lo que puede ser beneficioso para verificar que el proceso de recorte no haya destruido la película de carbono de la rejilla. Dependiendo del casete de muestra, también se pueden fresar rejillas sin recortar, pero se debe tener mucho cuidado durante la transferencia del crioFIB-SEM al TEM para limitar cualquier flexión de la rejilla, ya que esto romperá las laminillas. Las rejillas sin recortar pueden cargarse en el TEM mediante crioportadores de entrada lateral, pero existe una alta probabilidad de que las láminas se rompan si el recorte se lleva a cabo después del fresado.
Recubrimiento organoplatino
El recubrimiento organoplatino se realiza en una o ambas rejillas una vez que se han cargado en el crioFIB-SEM. La aguja del sistema de inyección de gas (GIS) se inserta en la cámara sobre la muestra para dirigir un flujo de vapor organoplatino a través de la superficie de la rejilla desde una fuente calentada durante un tiempo determinado. El vapor se condensa en superficies frías y forma una capa sólida (~2 μm de espesor). La integridad de esta capa es fundamental para permitir que las laminillas uniformemente delgadas sean fresadas. Las condiciones óptimas de aplicación para la capa de organoplatino generalmente están predeterminadas por el fabricante del instrumento, pero aún puede ser necesaria cierta optimización. La mayoría de los sistemas alinean la aguja SIG cerca de la dirección de fresado o perpendicular a la dirección de fresado, dependiendo de la geometría de los puertos en la cámara y los límites de rotación de la etapa. Se pueden probar diferentes configuraciones recubriendo la rejilla, fresando una pequeña región, inclinando el escenario y midiendo el grosor de la capa mediante SEM.
Además de la configuración de la crioFIB-SEM en sí, una serie de otros factores pueden afectar la aplicación de la capa organoplatino, incluyendo 1) la topografía de la muestra, 2) la contaminación superficial en la rejilla y 3) la reproducibilidad del flujo de vapor de la aguja GIS. Como el flujo SIG es direccional, la topografía desigual puede causar que las regiones en la sombra de las células o la contaminación de la superficie no estén recubiertas o tengan una capa más delgada. Esto puede conducir a un colapso de la capa organoplatino durante el pulido (Figura 3). Al seleccionar un área para fresar, es necesario tener en cuenta la topografía circundante, por ejemplo, la contaminación superficial grande, los grupos de células o el carbono roto que se proyectan desde la superficie de la rejilla hasta un par de cuadrados de rejilla, pueden bloquear el flujo de vapor, creando una sombra de organoplatino más delgado que puede debilitar una lámina. Además, también deben evitarse las partículas muy pequeñas de contaminación superficial cerca del borde frontal de la lámina, ya que pueden desprenderse durante el fresado, dejando un parche debilitado de hielo desnudo que puede provocar el desarrollo de un agujero en la lámina durante el pulido. Finalmente, si la molienda ha comenzado y la capa de organoplatino se ve inestable, cubra nuevamente, por más tiempo, o cambie a una rejilla de respaldo.

Figura 3: Un recubrimiento organoplatino de buena calidad es fundamental para obtener laminillas delgadas y uniformemente molidas. (A) Una micrografía del borde frontal de una lámina donde el recubrimiento organoplatino (OP, amarillo) se ha aplicado demasiado fino, lo que lleva a un orificio en el borde frontal de la lámina que se desarrolló durante el pulido (círculo verde) y un fresado desigual en todo el ancho de la lámina (rayas). La superficie organoplatino ha sido cortada por el haz de iones, lo que lleva a una pulverización de material detrás del borde frontal del recubrimiento (línea discontinua amarilla). (B) La capa organoplatino (OP) se ha aplicado más gruesamente, lo que resulta en una lámina más uniformemente adelgazada. La integridad de la capa se conserva en todo el ancho de la lámina y la interfaz entre la capa organoplatino y el material biológico vitrificado está bien definida (línea discontinua amarilla). La capa de carbono se puede ver corriendo a través de la parte posterior de la lámina (naranja). Barras de escala, 1 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Evaluación de la calidad de la rejilla en el crioFIB-SEM y el proceso de molienda
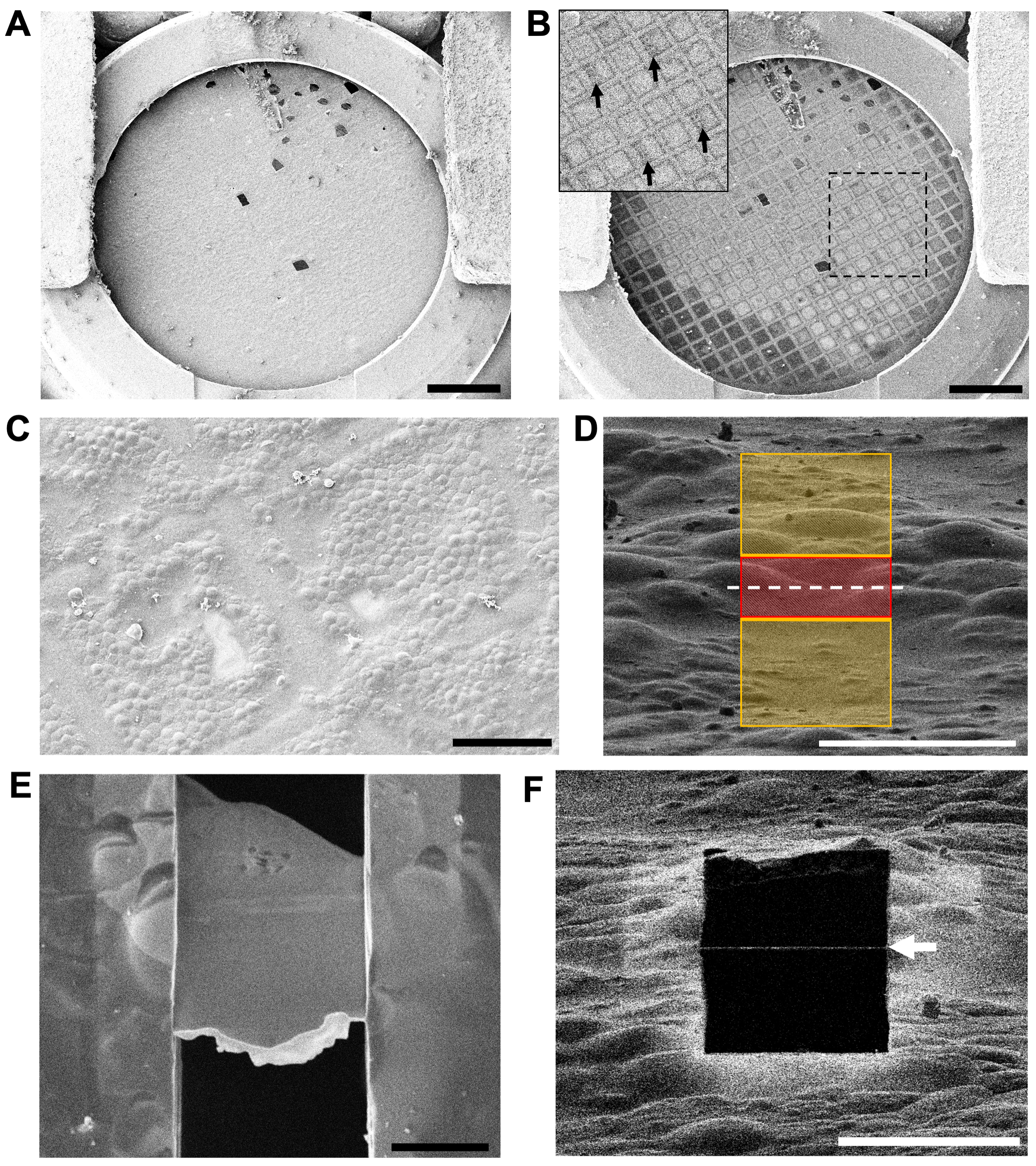
Una vez que las rejillas se han transferido a la crioFIB-SEM, la integridad de la película de carbono y la distribución de las células en las rejillas pueden ser examinadas por SEM (Figura 4A-C). Los gradientes de hielo, las posiciones de las barras de rejilla y la ubicación de los números de cuadrícula en las rejillas de búsqueda se pueden verificar mediante SEM de bajo aumento a 30 kV (Figura 4B), pero el voltaje debe reducirse a 5 kV mientras se ubican las posiciones de fresado a gran aumento y se monitorea el proceso de fresado para aumentar el contraste de la topografía de la superficie.

Figura 4: Evaluación de la calidad de la red y localización de áreas para fresar en el crioFIB-SEM . (A) Una visión general de bajo aumento de una red a 5 kV a través del SEM. La sección de corte del borde de la rejilla automática es visible en la parte inferior de la imagen. Barra de escala, 0,5 mm. (B) La misma cuadrícula a 30 kV por SEM, que muestra áreas de hielo más grueso (cuadrados de cuadrícula más oscuros) y hielo más delgado (cuadrados de cuadrícula más claros). El recuadro muestra la región en el cuadro, con flechas que indican la numeración en la cuadrícula del buscador, que es visible a 30 kV. Barra de escala, 0,5 mm. (C) Una visión general de aumento medio de dos cuadrados de cuadrícula que evalúa la distribución de las celdas en la película de carbono y la ubicación de las barras de cuadrícula. Barra de escala, 50 μm. (D) La disposición de los patrones de fresado para el primer corte a gran aumento (vista del haz de iones a 1,5 pA y 30 kV). La región roja (3 μm de espesor) está protegida, mientras que las regiones amarillas serán ablacionadas por el haz de iones. La línea discontinua blanca indica la posición de la lámina final. Barra de escala, 10 μm. (E) Una vista de gran aumento a 3 kV a través del SEM de una lámina pulida de 200 nm de espesor (10 μm de ancho x 15 μm de largo). La pérdida de contraste dentro de la lámina a 3 kV indica que se ha alcanzado un espesor adecuado. El borde frontal blanco brillante es la capa organoplatino restante que se aplica a la rejilla a través del SIG antes del fresado. Barra de escala, 5 μm. (F) La misma lámina de (E) vista usando el haz de iones a 30 kV y 1.5 pA. La delgada línea blanca a través del cuadrado negro (flecha blanca) es la capa de organoplatino restante en el borde frontal de la lámina. Barra de escala, 10 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Una región a fresar se selecciona colocando un par de patrones rectangulares a cada lado de una región protegida a gran aumento (~7,000x para esquizontes) en la vista de haz de iones (Figura 4D). Es fundamental que no se adhieran partículas de contaminación superficial cerca de la región de molienda, ya que pueden haber oscurecido la aplicación de la capa protectora de organoplatino. También es importante que la topografía de la región sea adecuada para soportar los lados de la lámina una vez que se haya alcanzado su espesor final.
Para esquizontes palúdicos (tamaño celular: ~ 5 μm de diámetro x 2 μm de espesor, forma de disco) se pueden fresar láminas de entre 7 y 20 μm de ancho. Si la capa celular es adecuadamente gruesa, las laminillas generalmente terminarán ~ 10-15 μm de longitud, capturando múltiples células por encima y por debajo de la capa de carbono (Figura 4E-F). Se puede esperar fresar 5-10 laminillas en una sesión de 8 h (6-7 h de fresado y 1-2 h de pulido). Esto variará dependiendo del grosor de la muestra y el ancho de las laminillas, con muestras más gruesas y laminillas más anchas que tardarán más en moler. Incluso las rejillas dañadas se pueden fresar, ya que solo se requiere un puñado de buenos cuadrados de cuadrícula para generar un conjunto de laminillas (Figura 5A). Además, si la muestra es más delgada de lo esperado, por ejemplo, debido a un exceso de transferencia o variación en el hematocrito del cultivo, las laminillas más cortas se pueden moler con relativa rapidez; sin embargo, su menor longitud limitará el área disponible para la recopilación de datos en el TEM (Figura 5B).

Figura 5: Determinación de cuándo se ha alcanzado el espesor final de la lámina durante el fresado. (A) Descripción general de bajo aumento de una cuadrícula por SEM a 5 kV que muestra el daño de carbono durante el recorte. Las áreas no dañadas contenían una muestra muy delgada debido al exceso de borrado; sin embargo, todavía era posible fresar seis laminillas en esta cuadrícula (la región dentro del contorno de guiones blancos) en una sesión de día completo en el crioFIBSEM. Barra de escala, 0,5 mm. (B) Una lámina corta (~10 μm de ancho x 3 μm de largo, sin incluir la capa organoplatino) producida a partir de esta rejilla (SEM a 3 kV), que todavía proporcionaba dos regiones de las cuales recolectar series de inclinación. Barra de escala, 25 μm. (C) Una serie de imágenes SEM (3 kV) de una lámina durante el paso final de pulido que muestra cómo se pierde contraste en la lámina a medida que se adelgaza (moviéndose de izquierda a derecha). La línea negra oscura a través del centro de la lámina en todas las imágenes es una tira de película de carbono de la cuadrícula. Las células delante de esta región fueron vitrificadas por encima de la película de carbono y las células detrás de esta región fueron vitrificadas por debajo de la película de carbono. El fresado se detuvo cuando la capa de organoplatino en el lado izquierdo del borde frontal de la lámina estaba a punto de perder integridad estructural. Este punto de parada fue antes de que toda la lámina se llevara a un grosor uniforme, por lo que todavía hay un material de mayor contraste en la parte posterior de la lámina. (D) Un ejemplo de una ruta de pulido basada en las laminillas fresadas en la rejilla que se muestra en (A). Una ruta de pulido debe comenzar en las laminillas más cercanas a la fuente FIB, alejándose de la fuente FIB para limitar la redeposición de material fresado en la superficie de las laminillas. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Durante el pulido, el espesor final de una lámina dependerá de la estructura de la muestra en la región que se está fresando, la integridad de la capa de organoplatino y el tiempo disponible. Idealmente, la muestra debe adelgazarse hasta que SEM pierda contraste en toda la superficie de la lámina a 3 kV, lo que sugiere que es uniformemente transparente a los electrones y de alrededor de 150-200 nm de espesor (Figura 5C). Sin embargo, puede ser necesario detener la molienda antes de este punto si la capa organoplatino desarrolla un orificio o la lámina comienza a doblarse. En este caso, la lámina aún puede ser lo suficientemente delgada en la parte delantera y seguir siendo útil para la tomografía. Por el contrario, es posible adelgazar más allá de la etapa de pérdida de contraste si la lámina se ve estable, haciéndola aún más delgada al acercar los patrones de fresado (~ 100 nm o menos). Esto dependerá del grosor que se requiera para el flujo de trabajo posterior. Se requiere una imagen SEM de bajo aumento para planificar una ruta de pulido, comenzando más cerca de la fuente del haz de iones y trabajando lejos (Figura 5D). La dirección de la ruta de pulido es importante para evitar la redeposición de material ablacionado en láminas que ya se han terminado.
Recopilación y procesamiento de datos de la serie basculante
Una vez cargado en el TEM, un montaje de cuadrícula completa de bajo aumento (~ 150x) identificará las posiciones de las laminillas, que se pueden correlacionar con la imagen SEM de bajo aumento tomada al final del fresado. Para los esquizontes congelados por inmersión, la mayor parte de la rejilla no es transparente a los electrones, por lo que las posiciones de las láminas aparecen como muescas blancas sobre un fondo negro (Figura 6A). Se debe tener en cuenta el ángulo de las laminillas con respecto al eje de inclinación del microscopio, ya que más de ~ 10 ° de distancia de la perpendicular podría dificultar la adquisición de la serie de inclinación. Un montaje de aumento medio (~ 1,500x) en la ubicación de cada lámina dará una visión general del contenido biológico y verificará si hay daños por transferencia, hielo cristalino o contaminación excesiva de la superficie (Figura 6B-D). Esto también debe verificarse a gran inclinación para garantizar que ninguna contaminación superficial oscurezca el área de adquisición o enfoque. La elección de una región de adquisición dependerá no solo de las características biológicas, sino también de la integridad estructural del hielo en la región circundante, por ejemplo, evitando grietas, ya que estas regiones se desplazarán, o áreas con cortinas excesivas, donde una lámina tendrá un grosor variable. Antes de adquirir una serie de inclinación, se aplica una preinclinación de ±10° a la rejilla para hacer que el plano de las laminillas (no la rejilla) sea perpendicular al eje óptico. La dirección de la pre-inclinación se puede determinar por la posición del borde frontal de las laminillas (buscando la capa de organoplatino sobrante) en el montaje de aumento medio. Para el TEM utilizado aquí (300 kV Titan Krios), si los bordes frontales de las laminillas apuntaban hacia arriba en el mapa, esto requería una pre-inclinación de +10° y si apuntaban hacia abajo, esto requería una pre-inclinación de -10°. Cada rejilla puede ser diferente debido a la orientación en la que fueron recogidas en las pinzas e insertadas en el cargador automático (sección cortada hacia la izquierda o hacia la derecha, produce una rotación de 180°). Una consideración final es el tamaño de píxel. Rutinariamente, las series de inclinación se recogen alrededor de 2,5-7 Å/píxel, teniendo en cuenta el tamaño de la característica de interés, la resolución objetivo de los datos tomográficos resultantes y el área de superficie de la lámina, lo que puede limitar el tamaño del área de adquisición. Es posible utilizar un tamaño de píxel más pequeño para obtener información de alta resolución y el más pequeño que hemos utilizado con éxito en estas muestras es de 1,4 Å/píxel (datos no mostrados). La deriva será más evidente en un tamaño de píxel más pequeño y solo es realmente adecuada para láminas delgadas (<100 nm) donde maximizar la resolución es importante en el estudio.

Figura 6: Elección de regiones accesibles de interés de las que adquirir series de inclinación (A) Una sección de un mapa TEM de bajo aumento que muestra una región que contiene laminillas, que aparecen como seis muescas blancas sobre un fondo negro (flechas rojas). Los cuadrados blancos son película de carbono rota. (B) Un mapa de aumento medio de una lámina dañada y desvitrificada. En el borde frontal de la lámina (FE), la capa organoplatino se ha desprendido (1). Hay un parche transparente de hielo cristalino en el centro de la lámina (2). El haz de iones no ha podido abrirse paso en el borde posterior de la lámina (BE) porque el hielo es demasiado grueso junto a las barras de rejilla. Solo una pequeña porción de la lámina está libre del hielo circundante en el BE (3), creando una cuña. El grosor también ha causado que los estantes se corten por encima de la lámina (4), lo que probablemente bloqueará la vista de las células en alta inclinación en el TEM. (C) Montaje de aumento medio de una lámina completa vista en el TEM. Los problemas típicos o regiones que deben evitarse al recolectar series de inclinación de láminas son: cubiertas con contaminación superficial (SC), cubiertas por la capa organoplatino (OP, amarillo), cerca de grietas (CR y flechas negras), con cortinas debido a cambios de densidad en el material biológico (CU y región dentro de los corchetes azules) y áreas debilitadas por roturas en la capa organoplatino (círculo verde). Las únicas regiones accesibles para la adquisición de series de inclinación son las áreas dentro de las dos cajas de guiones negros (etiquetadas 1 y 2). La vista debe verificarse a gran inclinación para asegurarse de que la contaminación de la superficie no oscurezca la región de interés o el área de enfoque. (D) Un montaje de aumento medio de una lámina mucho más limpia, pero que todavía tiene grietas (CR) causadas por el adelgazamiento de la capa organoplatino (círculo verde) durante el pulido. Aquí, la región de interés se destaca por el tipo de célula observada dentro de la lámina, que en este caso son los merozoítos individuales, colocados detrás de la capa de carbono (naranja) hacia la parte posterior de la lámina (caja discontinua negra). Barras de escala, 3 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

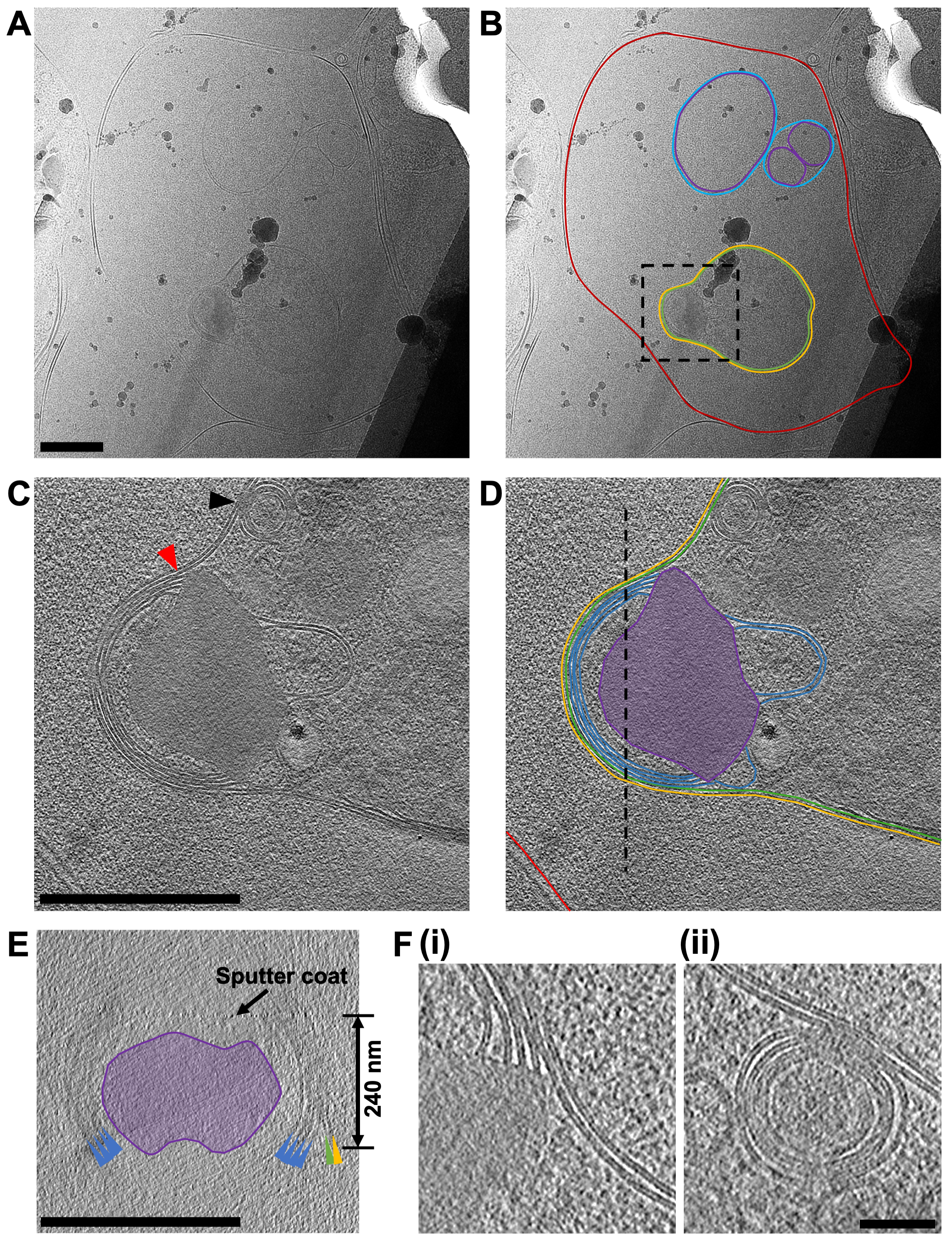
Figura 7: Adquisición de datos crio-ET de láminas fresadas por FIB. (A) Una micrografía de una región de una lámina que contiene un glóbulo rojo que ha sido invadido recientemente por un merozoíto de P. falciparum. En (B) la misma imagen está anotada para mostrar el límite del glóbulo rojo (rojo), una serie de vesículas intracelulares de doble pared (púrpura y azul para las membranas interna y externa, respectivamente) y el merozoíto (verde) rodeado por una segunda membrana derivada de la célula huésped (amarillo). Una caja negra muestra el área donde se adquirió una serie de inclinación. Barra de escala, 500 nm. (C) Un promedio de 20 cortes centrales vistos en el plano XY de un tomograma agrupado 8x (2.4 Å / píxel) adquirido en el extremo apical del merozoíto y en (D) su anotación, que muestra las dos membranas que rodean la célula (verde y amarillo) y cuatro membranas apiladas en el vértice del merozoíto (azul) que están asociadas con una característica densa en forma de hongo de electrones (púrpura). Una flecha roja indica la unión de las pilas de membrana en el ápice y la característica en forma de hongo (también se muestra en la parte F, i). Una flecha negra indica un evento de fusión entre la membrana plasmática del merozoíto y una de las vesículas multicapa dentro del parásito (también se muestra en la parte F, ii). La membrana de los glóbulos rojos del huésped se muestra (rojo) y una línea discontinua negra muestra la posición de una sección transversal vista en el plano XZ, que se muestra en (E). Las características en la sección transversal (E) están coloreadas y etiquetadas igual que en la parte (D), con flechas de colores apuntando a las membranas. Una flecha negra indica la posición de una capa de pulverización catódica aplicada a la lámina después del fresado y el grosor de la lámina en indicado. Para piezas (C-E), barra de escala, 500 nm. (F) Una vista más detallada de las características indicadas por las puntas de flecha rojas y negras en la parte (C), que muestra la definición en las bicapas lipídicas de la pila de membranas en el vértice del merozoíto (i) y el evento de fusión entre la vesícula multicapa con la membrana plasmática del merozoíto. Barra de escala, 75 nm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
El objetivo principal del trabajo es diseccionar la vía de salida de merozoíto en P. falciparum Pero dado que la población de células utilizadas en el estudio nunca puede ser completamente homogénea, a menudo se observaron otras etapas del desarrollo celular dentro de las laminillas. El ejemplo que se muestra aquí (Figura 7) contiene un glóbulo rojo que ha sido invadido recientemente por un P. falciparum Parásito (merozoíto). La lámina utilizada tenía un espesor de 240 nm y la rejilla había sido ligeramente pulverizada con platino en una atmósfera de argón antes de la inserción en el TEM, lo que resultó en un contraste ligeramente menor de lo que normalmente se esperaría. La membrana de los glóbulos rojos podría rastrearse en su totalidad alrededor de la célula. Dentro del glóbulo rojo había tres estructuras cerradas unidas a la membrana, dos vesículas, cada una rodeada por una membrana doble y una característica en forma de bombilla (1,2 μm x 0,9 μm en sus partes más anchas), lo que es consistente con que sea un merozoíto (Figura 7A-B). El contenido de las dos vesículas parecía ser similar en contraste con el contenido de los glóbulos rojos, lo que sugiere que estas vesículas pueden contener hemoglobina. La presencia de vesículas dentro del glóbulo rojo del huésped después de la invasión se ha observado previamente45. Se cree que surgen de la secreción de lípidos y otros factores de virulencia de orgánulos secretores llamados rovroteros en el merozoíto, que se descargan en el glóbulo rojo del huésped durante la invasión. El merozoíto está rodeado por dos membranas estrechamente asociadas, la más interna de las cuales es presumiblemente la membrana plasmática nativa del merozoíto, en la que no hay una capa superficial visible, y la más externa debe derivar de la membrana de glóbulos rojos del huésped que envuelve al merozoíto a medida que invade. El citoplasma del merozoíto contiene muchas vesículas de múltiples capas y una característica densa en forma de hongo de electrones adyacente a una pila de membranas en el ápice. Una serie de inclinación adquirida sobre esta región (2,4 Å/píxel) muestra que contiene una pila de cuatro membranas, que parecen estar conectadas a la característica en forma de hongo (Figura 7C-E). Sorprendentemente, esta morfología es bastante diferente a la disposición normal de orgánulos y estructuras celulares en el extremo apical de un merozoíto maduro. Para evaluar esto, se obtuvo una serie de inclinación comparativa (2,74 Å/píxel) sobre el extremo apical en un merozoíto maduro de un esquizont que había sido tratado con E64 antes de la congelación por inmersión y molienda (Figura 8). Esto muestra que el extremo apical de un merozoíto contiene dos orgánulos secretores prominentes en forma de maza llamados rhoptries ubicados dentro de un conjunto de tres anillos polares en la punta apical de la célula y rodeados por una serie de orgánulos secretores más pequeños llamados micronemas. El anillo polar más interno está unido a una estructura de membrana doble que subyace a la membrana plasmática del merozoíto, llamada complejo de membrana interna, que contiene proteínas motoras que impulsan la invasión. Se ha demostrado que durante la invasión el contenido de los orgánulos secretores se descarga sobre la superficie del merozoíto y en el glóbulo rojo del huésped, facilitando la unión de los merozoítos a los glóbulos rojos del huésped e iniciando el complejo motor que impulsa la invasión.45. Los datos aquí muestran que después de la invasión, el merozoíto no contiene estructuras observables que se asemejen a rhoptries, micronemes o anillos polares, lo que sugiere que la morfología del extremo apical del merozoíto cambia dramáticamente. La fusión de rovétricos antes de la invasión se ha demostrado previamente mediante TEM de secciones fijas a temperatura ambiente.46, que es consistente con la producción de la característica en forma de hongo que observamos en nuestro merozoíto posterior a la invasión. Las pilas de membranas conectadas a esta característica no se han observado previamente y, dado que no hay indicios de un IMC presente en el merozoíto recién invadido, planteamos la hipótesis de que las pilas de membranas pueden ser los restos de la maquinaria IMC que queda después de que se complete la invasión, pero esto aún no se ha confirmado.

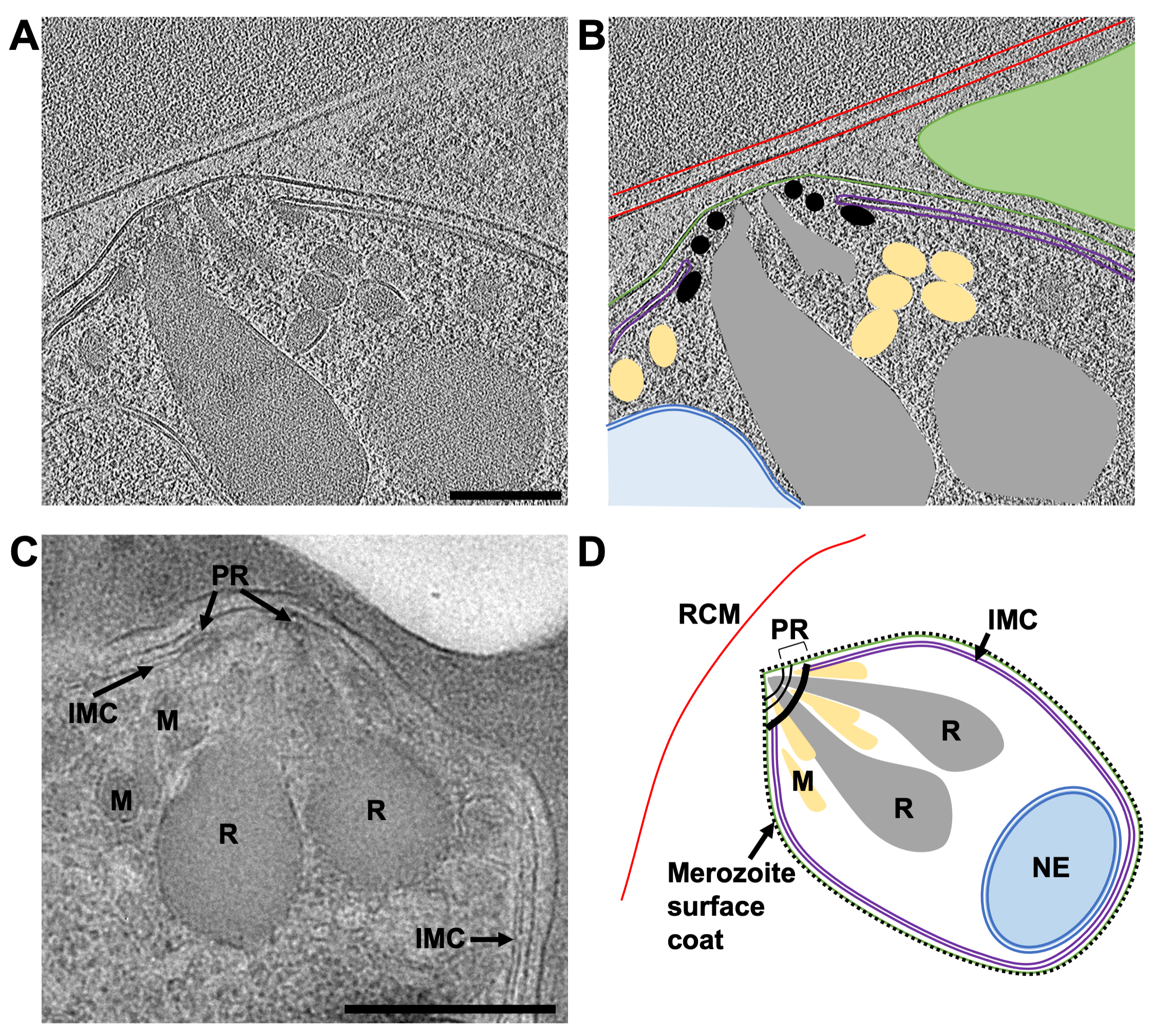
Figura 8: El extremo apical de merozoítos maduros por crio-ET de una lámina molida FIB y TEM de secciones plásticas. (A) Un promedio de 20 cortes centrales de un tomograma de 8x binned (2.74 Å / píxel) de una lámina de 230 nm de espesor que muestra la morfología típica del extremo apical de un merozoíto maduro. Los esquizontes fueron tratados con E64 antes de la congelación por inmersión, por lo que la membrana fotovoltaica se ha roto y los merozoítos están contenidos dentro del glóbulo rojo del huésped. (B) muestra lo mismo que en (A), pero con anotaciones para indicar la membrana de glóbulos rojos del huésped (RCM, rojo), la membrana plasmática de merozoíto (verde), el complejo de membrana interna (IMC, púrpura), anillos polares (PR, negro), micronemes (M, amarillo), ropterías (R, gris) y la envoltura nuclear (NE, azul). Un merozoíto vecino, que está fuera del plano en esta región del tomograma, está indicado por un triángulo verde. Barra de escala, 200 nm. (C) Las características celulares observadas por cryoET son consistentes con las de TEM de esquizontes conservados en secciones de plástico. Características celulares similares se indican en un merozoíto maduro de un esquizonte que ha sido tratado con el compuesto 2, deteniendo la salida antes de la ruptura de la membrana fotovoltaica. Barra de escala, 500 nm. (D) Un esquema anotado de un merozoíto que muestra las características celulares en partes (A-C). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
| Espesor de la región protegida entre los patrones de fresado rectangulares (μm) | Corriente de haz de iones (pA) nominal a 30 kV |
| 3 | 300 |
| 1.5 | 100 |
| 0.75 | 50 |
| 0.3 | 30 |
| 0.2 - 0.06 (pulido final) | 30* |
Tabla 1: Estrategia de molienda para producir laminillas finas. El fresado gradual se lleva a cabo reduciendo la corriente del haz de iones correspondiente al espesor de la región protegida. Esto limita el calentamiento de la muestra, evitando la desvitrificación. *El pulido debe realizarse en paralelo para aplicar calor uniformemente a las láminas desde ambos lados, reduciendo el riesgo de que se doblen o arqueen a medida que alcanzan su espesor final. Otros pasos se pueden hacer secuencialmente, fresando el patrón por encima de la lámina y luego por debajo de la lámina, antes de pasar a la siguiente corriente de haz. La inspección de la rejilla para localizar las posiciones de fresado debe llevarse a cabo a una corriente de haz de 1,5 pA para minimizar el calentamiento de la muestra. Haga clic aquí para descargar esta tabla.
Discusión
Mientras que la molienda crioFIB se está volviendo más rutinaria, la preparación de muestras óptimas para la molienda no lo ha hecho; por lo tanto, la mayoría de los pasos críticos en este protocolo ocurren antes de que la muestra llegue al crioFIB-SEM. Se requiere la optimización de la muestra mediante múltiples rondas de preparación celular, congelación por inmersión y cribado por microscopía óptica y electrónica para producir la mejor muestra posible antes de intentar la molienda. Tener la mejor muestra posible no solo aumenta las posibilidades de éxito sino que también optimiza el uso del equipo. Por esta razón, la mayoría de las instalaciones nacionales requieren evidencia de que se ha llevado a cabo una optimización suficiente antes de otorgar tiempo en sus máquinas. Una vez dentro de la crioFIB-SEM, maximizar la efectividad de la capa organoplatino es fundamental para producir laminillas uniformemente delgadas. Finalmente, la paciencia y el buen manejo de la muestra es una habilidad previa del usuario, que deberá sentarse durante varios días produciendo y luego transfiriendo rejillas que contengan delicadas laminillas. Esto puede cambiar con la introducción de estrategias de fresado automatizado47,48, pero hasta ahora, todo el proceso de fresado sigue siendo en gran medida manual en la mayoría de las instalaciones.
Si bien el trabajo se centró únicamente en los esquizontes de P. falciparum , el método presentado aquí podría modificarse fácilmente para optimizar la preparación de la rejilla y el fresado para otros tipos de células. Los factores importantes a considerar son la densidad de las celdas que se aplican a la rejilla, el tipo de rejilla (el cobre es tóxico para algunas celdas), el tamaño y el espaciado de los orificios en la película de carbono, el tiempo de transferencia y el método de transferencia (de una o dos caras, manual o automatizado). Además, si las células se cultivan en las rejillas (generalmente rejillas doradas con una película de carbono agujereada), la confluencia se puede verificar mediante microscopía óptica antes de secarse y congelarse. La estrategia de fresado depende de cómo se depositan las celdas en la rejilla. Para los glóbulos rojos infectados con malaria, la rejilla está esencialmente recubierta por una capa ininterrumpida de células, más gruesas en algunas áreas y más delgadas en otras. El fresado se lleva a cabo en múltiples puntos en una región a lo largo de este gradiente donde el espesor del hielo genera laminillas que son de un tamaño adecuado para la tomografía. Este enfoque funciona bien para células más pequeñas, ya que puede producir laminillas que contienen un corte a través de múltiples células. Para células más grandes o grupos de células, podría darse el caso de que una célula o grupo de células pueda producir una lámina.
El manejo de la red y la transferencia de muestras siguen siendo uno de los principales desafíos de este flujo de trabajo. Las laminillas pueden perderse debido a roturas o grietas, artefactos de cortina que oscurecen la biología, contaminación excesiva de la superficie, así como problemas de orientación de la rejilla dentro del TEM, lo que requiere un paso de manejo adicional para reposicionar la rejilla. El grado de pérdidas varía de un día a otro y de una red a otra, pero se mejora a través de la práctica y la experiencia de manejo a lo largo del tiempo. En este estudio, se encontró que las rejillas se pueden reorientar cuidadosamente en el cargador automático varias veces sin destruir las laminillas y esto a veces tiene el beneficio de lavar el hielo de la superficie. Otra limitación principal de este flujo de trabajo es el tiempo que lleva producir laminillas. Como la producción es lenta, es fundamental tener una muestra correctamente optimizada, haciendo que la molienda sea lo más eficiente posible.
Recientemente se han introducido varias adaptaciones a la molienda FIB de muestras biológicas vítreas. Un cambio de juego es la implementación de herramientas de elevación enfriadas criogénicamente dentro de la cámara crioFIB-SEM que permiten fresar bloques más grandes de material a partir de muestras congeladas a alta presión. Los bloques pueden fijarse a una varilla de metal o recogerse en una pinza y moverse a una segunda posición de muestra, que contiene una rejilla EM especialmente modificada. Los bloques pueden ser recubiertos con organoplatino y molidos para generar laminillas. La capacidad de fresar laminillas a partir de material congelado a alta presión significa que se pueden procesar células y tejidos mucho más grandes, específicamente dirigidos a áreas mediante microscopía de fluorescencia correlativa12. Otras adaptaciones recientes al método de fresado FIB incluyen la reducción de artefactos de cortina mediante el prefresado en cuña de la muestra16, la criofijación microfluídica49 y el fotomicropatrón de rejillas de microscopía electrónica para mejorar la distribución celular50. Además, se ha demostrado que los huecos de microexpansión de molienda a ambos lados de una lámina pueden aliviar la compresión de la muestra circundante a medida que alcanza su delgadez final51. Esto puede ser particularmente útil cuando se fresan capas celulares continuas, como la muestra en este estudio, donde a veces se observa flexión de laminillas durante el paso final de pulido.
El futuro de la microscopía electrónica probablemente será la determinación de estructuras moleculares in situ mediante el promedio de subtomogramas y el fresado FIB es una herramienta importante que facilitará la producción de muestras biológicas vítreas para este tipo de flujos de trabajo. Si bien la molienda FIB todavía está en su infancia para aplicaciones biológicas, el desarrollo de métodos está ocurriendo a un ritmo rápido gracias al arduo trabajo de los investigadores, tanto académicos como en instalaciones nacionales, además de la inversión comercial en el desarrollo de tecnología crioFIB-SEM para apoyar la investigación.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Esta investigación fue financiada en su totalidad, o en parte, por el Wellcome Trust (212916/Z/18/Z). Con el propósito de acceso abierto, el autor ha aplicado una licencia de derechos de autor pública CC BY a cualquier versión del manuscrito aceptado por el autor que surja de esta presentación.
Este proyecto para el cual se desarrolló este método fue financiado por una subvención del Consejo de Investigación Médica MR / P010288 / 1 otorgada a Helen R. Saibil, Roland A. Fleck y Michael J. Blackman. Los cultivos de P. falciparum se cultivaron en el Instituto Francis Crick, con el apoyo de miembros del grupo de Michael J. Blackman. Los autores desean agradecer al Dr. Ser Ying (Michele) Tan por proporcionar las imágenes de los esquizontes tratados con compuestos 2 y E64 en frotis de sangre delgada. La mayoría de las laminillas se produjeron con el apoyo del personal de eBIC y estamos agradecidos por el acceso a la crioFIB-SEM de doble haz Scios en la propuesta de investigación NT21004. Los autores también reconocen el apoyo del programa Royal Society Industry Fellowship (INF \ R2 \ 202061) para continuar el desarrollo de la técnica de fresado FIB dentro del CUI. Los autores también desean agradecer a Helen R. Saibil por las útiles discusiones en relación con este documento de métodos y la supervisión del proyecto.
Materiales
| Name | Company | Catalog Number | Comments |
| c-clips | Thermo Fisher Scientific | 1036171 | |
| Clipping station | Thermo Fisher Scientific | n/a | Direct quote from Thermo |
| Clipping station | Sub-angstrom | CSA-01 | |
| Compound 2 | n/a | n/a | Synthesised by Dr Simon A. Osborne, LifeArc |
| Cryo-FIB specific autogrid rims | Thermo Fisher Scientific | 1205101 | |
| E64 | Sigma | E3132 | |
| Emitech K100X glow discharge unit | Quorum | n/a | |
| Gibco RPMI 1640 media | Thermo Fisher Scientific | 12633012 | Formulation used for culturing is custom made (REF 041-91762 A) and includes Albumax, glutamine, HEPES and hypoxanthine supplements |
| Giemsa Stain | VWR International | 350864X | |
| Glass slides | Thermo Fisher Scientific | 11562203 | |
| Grid boxes | Sub-angstrom | PB | For clipped grids |
| Grid boxes | Thermo Fisher Scientific | n/a | For clipped grids - direct quote from Thermo Fisher Scientific |
| Grid boxes | Agar Scientific | AGG3727 | |
| Home-made manual plunge freezing rig | n/a | n/a | With an insulated ethane pot (high-density foam) and liquid nitrogen bath (polystyrene) on a bench top in a containment facility. |
| Human blood | n/a | n/a | UK National Blood and Transplant service |
| JEOL 4700F Z JSM with a Leica VCT500 stage cooling system | JEOL | n/a | FIB-SEM |
| Leica EM ACE600 with a VCT500 cryostage | Leica | n/a | sputter coater |
| Leica EM ACE900 | Leica | n/a | e-beam rotary coater. In a humidity controlled room. |
| Linkam cryo-stage for light microscope | Linkam | Model No. CMS196 | Cassettes for clipped and un-clipped grids. In a humidity controlled room. |
| Methanol | Sigma | 179957 | |
| Nikon Eclipse E200 light microscope | Nikon | n/a | with Linkam cryo-stage in a humidity controlled room. |
| Percoll | VWR International | 17-0891-01 | Solution for percoll cushion is 35 ml 10x PBS, 150 ml RPMI 1640 media and 315 ml Percoll |
| Quantifoil copper 200 mesh 2/4 holey carbon EM Finder grids | Quantifoil | N1-C17nCuH2-01 | 100 pack |
| Quantifoil copper 200 mesh 2/4 holey carbon EM grids | Quantifoil | N1-C17nCu20-01 | 100 pack |
| Quorum PP3010 prep-chamber | Quorum | n/a | sputter coater |
| Scios dual beam equipped with a Quorum PP3010 transfer stage. | FEI | n/a | FIB-SEM |
| Staedtler Lumocolor fine black permanent marker pen | Viking Direct | ND538522 | |
| TFS Titan Krios 300 kV TEM | Thermo Fisher Scientific | n/a | TEM equipped with a K2 or K3 camera. |
| Whatmann grade 1 filter paper | Sigma | WHA1001150 | |
| Zeiss Axio Scope A1 light microscope | Zeiss | n/a | with Linkam cryo-stage in a humidity controlled room. |
Referencias
- Baumeister, W. From proteomic inventory to architecture. FEBS Letters. 579 (4), 933-937 (2005).
- Rigort, A., Villa, E., Bäuerlein, F. J. B., Engel, B. D., Plitzko, J. M. Integrative approaches for cellular cryo-electron tomography: Correlative imaging and focused ion beam micromachining. Methods in Cell Biology. 111, 259-281 (2012).
- Oikonomou, C. M., Chang, Y. W., Jensen, G. J. A new view into prokaryotic cell biology from electron cryotomography. Nature Reviews Microbiology. 14, 205-220 (2016).
- Wagner, F. R., et al. Preparing samples from whole cells using focused-ion-beam milling for cryo-electron tomography. Nature Protocols. 15 (6), 2041-2070 (2020).
- Giannuzzi, L. A., Stevie, F. A. A review of focused ion beam milling techniques for TEM specimen preparation. Micron. 30 (3), 197-204 (1999).
- Narayan, K., Subramaniam, S. Focused ion beams in biology. Nature Methods. 12 (11), 1021-1031 (2015).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Acadamy of Science of the United States of America. 109 (12), 4449-4454 (2012).
- Marko, M., Hsieh, C., Schalek, R., Frank, J., Mannella, C. Focused-ion-beam thinning of frozen-hydrated biological specimens for cryo-electron microscopy. Nature Methods. 4 (3), 215-217 (2007).
- Villa, E., Schaffer, M., Plitzko, J. M., Baumeister, W. Opening windows into the cell: Focused-ion-beam milling for cryo-electron tomography. Current Opinion in Structural Biology. 23 (5), 771-777 (2013).
- Wang, K., Strunk, K., Zhao, G., Gray, J. L., Zhang, P. 3D structure determination of native mammalian cells using cryo-FIB and cryo-electron tomography. Journal of Structural Biology. 180 (2), 318-326 (2015).
- de Winter, D. A. M., et al. In-situ integrity control of frozen-hydrated, vitreous lamellas prepared by the cryo-focused ion beam-scanning electron microscope. Journal of Structural Biology. 183 (1), 11-18 (2013).
- Schaffer, M., et al. A cryo-FIB lift-out technique enables molecular-resolution cryo-ET within native Caenorhabditis elegans tissue. Nature Methods. 16 (8), 757-762 (2019).
- Mahamid, J., et al. Visualizing the molecular sociology at the HeLa cell nuclear periphery. Science. 351 (6276), 969-972 (2016).
- Albert, S., et al. Proteasomes tether to two distinct sites at the nuclear pore complex. Proceedings of the National Acadamy of Science of the United States of America. 114 (52), 13726-13731 (2017).
- Guo, Q., et al. In situ structure of neuronal C9orf72 poly-GA aggregates reveals proteasome recruitment. Cell. 172 (4), 696-705 (2018).
- Schaffer, M., et al. Optimized cryo-focused ion beam sample preparation aimed at in situ structural studies of membrane proteins. Journal of Structural Biology. 197 (2), 73-82 (2017).
- Szwedziak, P., Wang, Q., Bharat, T. A. M., Tsim, M., Löwe, J. Architecture of the ring formed by the tubulin homologue FtsZ in bacterial cell division. eLife. 3, 04601 (2014).
- Carlson, L. A., et al. Cryo electron tomography of native HIV-1 budding sites. PLOS Pathogens. 6 (11), 1001173 (2010).
- Klein, S., et al. SARS-CoV-2 structure and replication characterized by in situ cryo-electron tomography. Nature Communications. 11 (1), 5885 (2020).
- Studer, D., Humbel, B. M., Chiquet, M. Electron microscopy of high pressure frozen samples: Bridging the gap between cellular ultrastructure and atomic resolution. Histochemistry and Cell Biology. 130 (5), 877-889 (2008).
- Schertel, A., et al. Cryo FIB-SEM: Volume imaging of cellular ultrastructure in native frozen specimens. Journal of Structural Biology. 184 (2), 355-360 (2013).
- Murphy, G. E., et al. Correlative 3D imaging of whole mammalian cells with light and electron microscopy. Journal of Structural Biology. 176 (3), 268-278 (2012).
- Heymann, J. A. W., et al. Site-specific 3D imaging of cells and tissues with a dual beam microscope. Journal of Structural Biology. 155 (1), 63-73 (2012).
- Spehner, D., et al. Cryo-FIB-SEM as a promising tool for localizing proteins in 3D. Journal of Structural Biology. 211 (1), 107528 (2020).
- Kamino, T., Yaguchi, T., Ohnishi, T., Ishitani, T., Osumi, M. Application of a FIB-STEM system for 3D observation of a resin-embedded yeast cell. Journal of Electron Microscopy. 53 (5), 563-566 (2004).
- Han, H., Zuber, B., Dubochet, J. Compression and crevasses in vitreous sections under different cutting conditions. Journal of Microscopy. 230, 167-171 (2008).
- Matias, V. R. F., Al-amoudi, A., Dubochet, J., Beveridge, T. J. Cryo-transmission electron microscopy of frozen-hydrated sections of Escherichia coli and Pseudomonas aeruginosa. Journal of Bacteriology. 185 (20), 6112-6118 (2003).
- Al-Amoudi, A., Studer, D., Dubochet, J. Cutting artefacts and cutting process in vitreous sections for cryo-electron microscopy. Journal of Structural Biology. 150 (1), 109-121 (2005).
- Al-Amoudi, A., et al. Cryo-electron microscopy of vitreous sections. The EMBO Journal. 23 (18), 3583-3588 (2004).
- Bouchet-Marquis, C., Dubochet, J., Fakan, S. Cryoelectron microscopy of vitrified sections: A new challenge for the analysis of functional nuclear architecture. Histochemistry and Cell Biology. 125 (1-2), 43-51 (2006).
- Hayles, M. F., et al. The making of frozen-hydrated, vitreous lamellas from cells for cryo-electron microscopy. Journal of Structural Biology. 172 (2), 180-190 (2010).
- Dubochet, J., Adrian, M., Chang, J. -. J., Lepault, J., McDowall, A. W. Cryoelectron microscopy of vitrified specimens. Cryotechniques in Biological Electron Microscopy. , 114-131 (1987).
- Hsieh, C., Schmelzer, T., Kishchenko, G., Wagenknecht, T., Marko, M. Practical workflow for cryo focused-ion-beam milling of tissues and cells for cryo-TEM tomography. Journal of Structural Biology. 185 (1), 32-41 (2014).
- Harapin, J., et al. Structural analysis of multicellular organisms with cryo-electron tomography. Nature Methods. 12 (7), 634-636 (2015).
- Rubino, S., et al. A site-specific focused-ion-beam lift-out method for cryo Transmission Electron Microscopy. Journal of Structural Biology. 180 (3), 572-576 (2012).
- Duyvesteyn, H. M. E., et al. Machining protein microcrystals for structure determination by electron diffraction. Proceedings of the National Acadamy of Science of the United States of America. 115 (38), 9569-9573 (2018).
- Blackman, M. J. Purification of Plasmodium falciparum merozoites for analysis of the processing of merozoite surface protein-1. Microbes as Tools for Cell Biology. 45, 213-220 (1995).
- Hagen, W. J. H., Wan, W., Briggs, J. A. G. Implementation of a cryo-electron tomography tilt-scheme optimized for high resolution subtomogram averaging. Journal of Structural Biology. 197 (2), 191-198 (2017).
- Zheng, S. Q., et al. MotionCor2 - anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nature Methods. 14 (4), 331-332 (2017).
- Grant, T., Grigorieff, N. Measuring the optimal exposure for single particle cryo-EM using a 2.6 Å reconstruction of rotavirus VP6. eLife. 4, 06980 (2015).
- Rohou, A., Grigorieff, N. CTFFIND4: Fast and accurate defocus estimation from electron micrographs. Journal of Structural Biology. 192 (2), 216-221 (2015).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Turoňová, B., Schur, F. K. M., Wan, W., Briggs, J. A. G. Efficient 3D-CTF correction for cryo-electron tomography using NovaCTF improves subtomogram averaging resolution to 3.4 Å. Journal of Structural Biology. 199 (3), 187-195 (2017).
- Arnold, J., et al. Site-specific cryo-focused ion beam sample preparation guided by 3d correlative microscopy. Biophysical Journal. 110 (4), 860-869 (2016).
- Riglar, D. T., et al. Super-resolution dissection of coordinated events during malaria parasite invasion of the human erythrocyte. Cell Host & Microbe. 9 (1), 9-20 (2011).
- Hanssen, E., et al. Electron tomography of Plasmodium falciparum merozoites reveals core cellular events that underpin erythrocyte invasion. Cellular Microbiology. 15 (9), 1457-1472 (2013).
- Buckley, G., et al. Automated cryo-lamella preparation for high-throughput in-situ structural biology. Journal of Structural Biology. 210 (2), 107488 (2020).
- Zachs, T., et al. Fully automated, sequential focused ion beam milling for cryo-electron tomography. eLife. 9, 52286 (2020).
- Fuest, M., et al. In situ microfluidic cryofixation for cryo focused ion beam milling and cryo electron tomography. Scientific Reports. 9 (1), 19133 (2019).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Wolff, G., et al. Mind the gap: Micro-expansion joints drastically decrease the bending of FIB-milled cryo-lamellae. Journal of Structural Biology. 208 (3), 107389 (2019).
Erratum
Formal Correction: Erratum: Preparing Lamellae from Vitreous Biological Samples using a Dual-Beam Scanning Electron Microscope for Cryo-Electron Tomography
Posted by JoVE Editors on 9/11/2023. Citeable Link.
An erratum was issued for: Preparing Lamellae from Vitreous Biological Samples using a Dual-Beam Scanning Electron Microscope for Cryo-Electron Tomography. The Authors section was updated from:
Claudine Bisson1,2
Corey W. Hecksel3,4
James B. Gilchrist3
Roland A. Fleck1
1Centre for Ultrastructural Imaging, New Hunt’s House, Guy’s Campus, King’s College London
2Department of Biological Science, Birkbeck College, University of London
3Electron Bio-Imaging Centre, Diamond Light Source, Harwell Science and Innovation Campus
4SLAC National Accelerator Laboratory, Stanford University
to:
Claudine Bisson1,2
Corey W. Hecksel3,4
James B. Gilchrist3
M. Alejandra Carbajal1
Roland A. Fleck1
1Centre for Ultrastructural Imaging, New Hunt’s House, Guy’s Campus, King’s College London
2Department of Biological Science, Birkbeck College, University of London
3Electron Bio-Imaging Centre, Diamond Light Source, Harwell Science and Innovation Campus
4SLAC National Accelerator Laboratory, Stanford University
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados