
Method Article
Kriyo-Elektron Tomografisi için Çift Işınlı Taramalı Elektron Mikroskobu Kullanarak Vitreus Biyolojik Numunelerinden Lamel Hazırlanması
Bu Makalede
Erratum Notice
Özet
Kriyo-elektron tomografisi için dalma dondurulmuş biyolojik örneklerden vitreus ızgara üzerinde lamel üretmek için odaklanmış iyon ışını frezeleme kullanımı.
Özet
Burada, diğer biyolojik örnekler için kolayca uyarlanabilen Plasmodium falciparum ile enfekte olmuş insan eritrositlerinin dalma dondurulmuş ızgaralarından kriyo-lamel hazırlamak için bir protokol sunulmaktadır. Numune hazırlamak, frezelemek ve lamelleri görüntülemek için temel prensipler tüm cihazlarda ortaktır ve protokol, kriyo-elektron mikroskobu (cryoEM) ve kriyo-elektron tomografisi (cryoET) için ızgara üzerinde kriyo-lamel hazırlığı için genel bir kılavuz olarak takip edilebilir. Hücreleri destekleyen elektron mikroskobu ızgaraları, manuel veya otomatik bir dalma dondurucu kullanılarak sıvı azot soğutmalı sıvı etana daldırılır, daha sonra bir kriyo-aşama ile donatılmış bir ışık mikroskobunda taranır. Dondurulmuş ızgaralar, odaklanmış bir iyon demeti (cryoFIB-SEM) ile donatılmış bir kriyo-taramalı elektron mikroskobuna aktarılır. Izgaralar, frezelemeden önce rutin olarak püskürtme kaplamalıdır, bu da frezeleme sırasında yük birikiminin dağılmasına yardımcı olur. Alternatif olarak, ızgaralara tam kalınlığı daha hassas bir şekilde kontrol edilebilen bir karbon-platin tabakası uygulamak için bir e-beam döner kaplayıcı kullanılabilir. KriyoFIB-SEM'in içine girdikten sonra, bir gaz enjeksiyon sistemi (CBS) aracılığıyla ızgaranın yüzeyine bir organoplatin bileşiğinin ek bir kaplaması uygulanır. Bu tabaka, lamellerin ön kenarını öğütülürken korur, bütünlüğü düzgün ince lameller elde etmek için kritik öneme sahiptir. İlgilenilen bölgeler SEM aracılığıyla tanımlanır ve frezeleme, aşırı ısı oluşumunu önlemek için lamel elektron saydamlığına ulaştığında iyon ışınının akımını azaltarak kademeli bir şekilde gerçekleştirilir. Çoklu lamelleri olan bir ızgara daha sonra eğim serisi edinimi için kriyojenik koşullar altında bir transmisyon elektron mikroskobuna (TEM) aktarılır. Lamel hazırlığı için sağlam ve kontaminasyonsuz bir iş akışı, hücresel kriyoEM, kriyoET ve alt tomogram ortalaması dahil olmak üzere aşağı akış teknikleri için önemli bir adımdır. Özellikle yüksek basınçlı dondurulmuş numunelerin kaldırılması ve frezelenmesi için bu tekniklerin geliştirilmesi, sahada yüksek önceliğe sahiptir.
Giriş
Sadece biyolojik örneklerin hücresel içeriği <500 nm kalınlığında, kriyojenik sıcaklıklarda iletim elektron mikroskobu (TEM) ile etkili bir şekilde görüntülenebilir, bu da örneklerin aralığını virüslere, prokaryotlara, basit tek hücreli organizmalara ve daha büyük ökaryotik hücrelerin daha ince bölgelerine sınırlar1. Şebeke odaklı iyon demeti (FIB) frezeleme, kriyojenik sıcaklıklarda (< -150 ° C) elektron şeffaf lamellerde daha kalın daldırma dondurulmuş biyolojik numunelerin inceltilmesini sağlar. Elde edilen lameller daha sonra görselleştirme ve tomografik veri toplama için bir TEM'e aktarılır ve hücrelerin içindeki hücresel ve moleküler özelliklerin yüksek çözünürlüklü 3D rekonstrüksiyonlarını mümkün kılar (incelemeler için bakınız Rigort ve ark., 20122, Oikonomou ve ark., 20163 ve Wagner ve ark., 20204).
FIB-frezeleme, numunelerin aşağı akış analizine hazırlanmak için rutin olarak inceltildiği malzeme bilimleri alanından ortaya çıkmıştır5. İki optik sütuna sahip bir taramalı elektron mikroskobunda (SEM) gerçekleştirilir: geleneksel taramalı elektron mikroskobu optikleri ve FIB-SEM adı verilen odaklanmış bir iyon ışını (FIB) üretebilen ve hassas bir şekilde kontrol edebilen optikler içeren ikinci bir sütun. Bu, numunenin belirli bir bölgesinin bir galyum kaynağı tarafından üretilen iyonlar tarafından ablatlanmasına, fazla malzemenin uzaklaştırılmasına ve bir lamel6'nın arkasında bırakılmasına izin verir. Frezeleme işlemi, ilgilenilen bölgeleri bulmak ve frezeleme ilerlemesini izlemek için kullanılan numunenin topografyasının SEM görüntülemesi ile yönlendirilir. Biyolojik uygulamalar için, temel kurulum büyük ölçüde aynıdır, ancak frezeleme kriyojenik sıcaklıklarda gerçekleştirilir. Bu, standart FIB-SEM'lerin sabit sıcaklıkları ve düşük yüzey kontaminasyon oranlarını koruyan kriyo-soğutmalı aşamalara ve ayrıca sapma veya kontaminasyon olmadan numune transferini kolaylaştırmak için hava kilitlerine sahip olacak şekilde uyarlanmasını gerektirmiştir. Numune mekikleri ayrıca, TEM ızgaraları, planşetler ve kılcal damarlar dahil olmak üzere kriyoFIB-SEM'in içine bir dizi farklı taşıyıcının monte edilmesine izin verecek şekilde modifiye edilmiştir. Birkaç kilit araştırmacı grubu, bu yöntemlerin geliştirilmesinde ve bu alanda devam eden teknolojik gelişmelerde merkezi olmuştur 7,8,9,10,11,12. Kriyojenik sıcaklıkta biyolojik FIB frezeleme için ticari çözümler artık daha yaygın olarak mevcuttur ve optimize edilmiş bir numune göz önüne alındığında, lamellerin ızgara üzerinde frezelenmesi daha rutin hale gelmektedir.
Bir dizi oda sıcaklığı ve kriyoEM tekniği, mütevazı çözünürlükte tüm çok hücreli organizmalardan hücresel düzeyde karmaşık süreçlerin bağlamını anlamaya ve daha da ayrıntılı olarak yaşamın tüm ölçeklerinde hücresel bilgiyi görselleştirmek için kullanılabilir. in situ moleküler yapılar13,14,15,16,17,18,19. Klasik oda sıcaklığı teknikleri, TEM tarafından hücresel morfolojinin analizi için ultramikrotomi ile sabit ve lekeli, reçine gömülü hücrelerin ve dokuların kesitlendirilmesini içerir (gözden geçirme için bakınız Studer ve ark., 200820). Korunmuş hücrelerin bloklarının yüzeyini görüntülemek için SEM tarafından ikincil elektron saçılmasından yararlanılan alternatif teknikler, malzemeyi bir bıçakla (seri blok yüz görüntüleme) veya odaklanmış bir iyon ışınıyla aşamalı olarak çıkarmadan önce geliştirilmiştir21,22,23. Bu teknik aynı zamanda kriyojenik sıcaklıklarda (kriyo-hacim görüntüleme olarak adlandırılır) vitreus, lekesiz hücre veya doku blokları üzerinde bir kriyoFIB-SEM ile başarıyla uygulanmıştır24. Alternatif olarak, daha kalın lameller (~ 15 μm kalınlığında) öğütülebilir ve STEM görüntüleme ile incelenebilir25. Bu teknikler kullanılarak, popülasyon bilgisi toplamak için birçok hücre içeren bütün bloklar görüntülenebilir veya bütün bir organ / organizma 3D olarak görüntülenebilir ve yeniden yapılandırılabilir. Bununla birlikte, hücrelerden yüksek çözünürlüklü moleküler bilgiye erişmek için, numunelerin neredeyse yerli, dondurulmuş hidratlanmış bir durumda korunması ve bu nedenle kriyojenik koşullar altında hazırlanması gerekir. Vitröz kesitlerin kriyo-elektron mikroskopisi (CEMOVIS), yüksek basınçlı dondurulmuş biyolojik materyal bloklarının kriyojenik koşullar altında bir ultramikrotom ile kesitlendiği bir tekniktir. Bu, kriyo-kesitli şeritler üretir (40-100 nm kalınlığında)26, bir EM ızgarasına bağlanır ve TEM'de görüntülenir. Bununla birlikte, bıçağın vitreus numunesi ile fiziksel etkileşimi, hücresel yapıyı ciddi şekilde bozabilen çatlama ve sıkıştırma artefaktlarına neden olur27,28,29,30. Daha kalın bölümler bu eserlere daha yatkındır, bu da ~ 70 nm'den daha kalın bölümlerin kullanılmasını pratik hale getirmez26. Bu sınırlama, tomogramdaki biyolojik yapının 3B görünümünü büyük ölçüde kısıtlar. Kriyojenik sıcaklıklarda FIB-frezeleme bu sorunları yaşamaz, ancak numunenin parçaları arasında diferansiyel frezeleme oranlarının neden olduğu kendi eserlerine sahiptir ve bu da bir lamel içinde perdeleme adı verilen değişken kalınlıklara yol açar. Bu sorun, frezeleme sırasında lamelin ön kenarını koruyan bir gaz enjeksiyon sistemi (GIS) ile uygulanan bir organoplatin kaplamanın uygulanmasıyla hafifletilir31. Izgara üstü FIB frezeleme için numune kalınlığının üst sınırı, numuneyi vitröz tutarken dondurma kabiliyeti ile tanımlanır32Bununla birlikte, kriyo kaldırma tekniklerinin ve biyolojik numuneler için uyarlanmış numune taşıyıcılarının tanıtılmasıyla birlikte, FIB-frezeleme yüksek basınçlı dondurulmuş numuneleri işlemek için de kullanılabilir31,33,34,35. Ek olarak, daldırma dondurulmuş numuneler çok ince olamaz, çünkü gerekli büyütmede eğim serilerini toplamak için yeterli yüzey alanı sağlayacak makul büyüklükte bir lamel üretmek için yeterli biyolojik malzeme olması gerekir. Bu sorun, bakteri veya maya gibi daha küçük hücrelerin kümelerinin öğütülmesiyle hafifletilebilir. Son lamel kalınlığı (~ 100-300 nm) genellikle numune bütünlüğü ve frezeleme stratejisi tarafından belirlenir. Daha ince lameller, sub-tomogram ortalaması gibi yüksek çözünürlüklü yapısal çalışmalar için daha iyidir, ancak daha kalın lameller, CEMOVIS tarafından elde edilebilenden çok daha büyük hücresel hacimler içerir ve doğal olarak korunmuş bir numunede daha fazla hücresel bağlam sağlar. FIB-frezeleme, elektron kırınım çalışmaları için daldırma-dondurulmuş protein kristallerini inceltmek için de kullanılabilir36.
Vitröz hücrelerin FIB frezelenmesi, bilimsel sorunun in situ olarak yakın doğal örneklerin yüksek çözünürlüklü moleküler detayını gerektirmesi durumunda gerçekleştirilmesi gereken zamana ve çabaya değer. Lamellerin rutin üretimi için daha fazla tesise erişim ile, hız sınırlama adımı genellikle numunenin camsı olduğundan ve sağlam ve homojen ince lameller üretmek için uygun bir kalınlıkta olduğundan emin olmak için zaman alınması gereken frezelemeden önce numunenin optimizasyonudur. Burada tarif edilen, sıtmanın etken maddesi olan Plasmodium falciparum parazitleri ile enfekte olmuş FIB-öğütme dalma-dondurulmuş insan kırmızı kan hücreleri için numune optimizasyonudur, ancak bu yaklaşım herhangi bir örnek için uyarlanabilir.
Protokol
İnsan kanı, İngiltere Ulusal Kan ve Nakil servisi aracılığıyla anonimleştirilmiş bağışçılardan elde edildi ve alındıktan sonraki 2 hafta içinde kullanıldı. Kullanımı için etik onay gerekmez.
1. Plasmodium falciparum ile enfekte kırmızı kan hücrelerinin hazırlanması ve daldırılması
- Olgun şizontları% 70 (v / v) izotonik yoğunluk gradyan ortamı üzerinde santrifüjleme (1.580 x g) ile izole edin (insan eritrositlerinde 3D7 Plasmodium falciparum'un aseksüel kan aşamalarının nasıl kültürleneceğine ilişkin standart prosedürler için bakınız Blackman M.J., 199537).
- 6.7 mM fosfat tamponunda, pH 7.1'de% 10 Giemsa boyası ile boyamadan önce şizontların morfolojik homojenliğini kontrol etmek için hava ile kurutulmuş ince kan filmlerini %100 metanol içeren bir cam slayt üzerine sabitleyin.
NOT: Belirli çıkış noktalarında duran şizontların preparatını zenginleştirmek için, şizontlar bileşik 2 ve E64 inhibitörleri ile daha da senkronize edilebilir (bu inhibitörlerin etkisinin bir açıklaması için Temsili Sonuçlar ve Şekil 1'e bakınız).
DİKKAT: Plasmodium falciparum bir insan patojenidir ve sadece yerel sağlık ve güvenlik kurallarına uygun olarak uygun bir muhafaza tesisinde ele alınmalıdır. - Şizontları (240 x g) pelet haline getirmek için nazikçe santrifüj edin ve RPMI ortamının hücre peletinin 2x hacminde yeniden askıya alın, bu da %50'lik bir hematokrit süspansiyonu ile sonuçlanır.
- Manuel bir daldırma dondurma makinesinde, kızdırma deşarjlı bir yüzeyin karbon tarafına 2,5 μL schizonts uygulayın (60 s, havada 30 mA'lık kızdırma-deşarj ünitesi ayarlarını kullanın, yalnızca ızgaranın karbon tarafını işleyin) 2/4 delikli karbon filmli 200 örgü bakır ızgara ve yırtık kenarlı grade 1 filtre kağıdı kullanarak ızgaranın arkasından ~ 20 s boyunca lekeleyin. Sıvı azot soğutmalı sıvı etan'a daldırın ve ızgaraları depolamaya aktarın (bu çalışmada kullanılan ekipman için Malzeme Tablosuna bakınız).
NOT: Deney burada duraklatılabilir ve ızgaralar süresiz olarak sıvı azot altında saklanabilir.
DİKKAT: Sıvı azot boğucudur ve donmaya neden olur; Oksijen izleme ile uygun bir ortamda dikkatli kullanın.
DİKKAT: Sıvı etan ciddi yanıklara neden olur ve yanıcıdır; ateşleme kaynaklarından uzakta bir duman davlumbazında kullanın.
NOT: Dalma dondurulmuş schizonts artık geçerli değildir. Bu, insan kanının birkaç altın ızgara ile hava çözülmüş dalma donmuş şizontlarla inkübe edilmesi ve donmamış kontrollere kıyasla birkaç gün sonra parazit büyümesinin gözlenmemesiyle belirlendi. Bu nedenle, dondurulmuş şizontların ızgaraları, normal güvenlik ve dekontaminasyon prosedürleri (eldivenler, %>70 etanol ile yüzey/alet sterilizasyonu ve ızgaraların %>70 etanol) kullanılarak muhafaza tesislerinin dışında elleçlenmesi güvenlidir. - Bir ışık mikroskobu için bir kriyo-aşama kullanarak ızgaraları eleyin, ızgara boyunca buz gradyanına özellikle dikkat edin. Buzun daha ince bölgelerinde, hücre kapsamı için ayrı ayrı ızgara karelerini kontrol edin.
NOT: En iyi kareler, ızgara karesinin merkezinde bir hücre kalınlığında olmalıdır (Şekil 2). Bu, bir lamelin karenin merkezi boyunca, kenarlardaki daha kalın buza ızgara çubuklarına çarpmadan öğütülebilmesini sağlar. Deney burada duraklatılabilir ve ızgaralar süresiz olarak sıvı azot altında saklanabilir. Hücreler bir floresan işaretleyici taşıyorsa, ızgaralar, doğrudan frezeleme için kriyoFIB-SEM'deki ızgara konumlarıyla ilişkilendirilebilen X / Y konumlarını bulmak için kriyoCLEM tarafından da taranabilir.
2. Daldırma dondurulmuş hücrelerin ızgara üzerinde FIB frezelenmesi
- Kesme bölümünün merkezini ve jantın karşı tarafını belirtmek için cryoFIB'ye özgü otomatik ızgara jantlarının önünü siyah silinmez bir işaretleyici kalemle işaretleyin (Şekil 3). Kırpma istasyonunu kurun ve ızgaraları karbon tarafı aşağı doğru işaretlenmiş halkalara klipsleyin.
- Izgaraları kriyoFIB-SEM mekiğine (mekiğe bağlı olarak tipik olarak 2 ızgara) karbon tarafına yükleyin ve argon atmosferinde (5 x 10-2 mbar) (60 s için 5 mA - kalınlık değişkendir) veya hücrelerin yüzeyine bir karbon / platin e-ışın döner kaplama (~ 4 nm kalınlığında) bir platin püskürtme kaplaması uygulayın.
NOT: Her iki kaplama türü de SEM görüntüleme sırasında yük dağılımına yardımcı olur. E-beam döner kaplayıcının yararı, kaplamanın tam kalınlığının belirlenebilmesidir. - Mekiği kriyoFIB-SEM'e yükleyin ve her ızgaradaki hücre dağılımını SEM ile 5 kV'da (13 pA veya 25 pA) değerlendirin. Izgaradaki buz gradyanlarına bakmak için düşük büyütmeli bir genel bakış (~ 100x) alın. Ardından, tek tek ızgara karelerine bakmak ve ızgaranın görünür hücresel özelliklere ve öğütülecek düşük yüzey kirliliğine sahip alanlarını tanımlamak için daha yüksek büyütmeli (~ 5.000x) görüntüler alın.
- Gaz enjeksiyon sistemini (GIS) kullanarak her ızgaranın yüzeyine >2 μm organoplatin kaplama uygulayın. Bunu yapmak için, CBS iğnesini ızgaranın üzerindeki odaya yerleştirin ve bir buhar akışı üretmek için organoplatin kaynağını belirli bir süre için (örneğin, 3-10 s için ~ 27 ° C) ayarlanmış bir sıcaklığa ısıtın.
NOT: CBS iğnesi aracılığıyla organoplatin kaplamanın uygulama açısı, sıcaklığı ve zamanlaması, eşit bir kaplamayı en üst düzeye çıkarmak için optimize edilmelidir. Bu, kullanılan spesifik kriyoFIB-SEM'e bağlı olacaktır (daha fazla açıklama için Temsili Sonuçlara bakınız). - Numuneyi, ızgaranın düzlemi iyon ışınının insidans açısından ~ 10 ° olacak şekilde eğin ve hem SEM hem de FIB görüntülerinde görülecek uygun bir ızgara karesinin merkezini hareket ettirin.
- İyon ışınını düşük akımda (nominal olarak 1,5 pA, 30 kV) kullanarak ızgarayı inceleyin ve bir ızgara karesinin merkezindeki hücreleri görselleştirmek için yeterince yüksek büyütmeye (~ 7.000x) gidin. Numuneyi ilk frezeleme akımında (300 pA) odak noktasına getirin ve astigmatizma için düzeltin. Parlaklığı ve kontrastı ayarlayın, ardından biri 3 μm kalınlığında korunan bir bölgenin üstünde ve diğeri altında olmak üzere frezelemek için iki dikdörtgen deseni işaretleyin, bunun merkezi son lamelin istenen yeridir.
NOT: Desenlerin seçilen genişliği, çevreleyen hücre katmanının topografyasına ve hücre boyutuna bağlı olacaktır. 7-20 μm, şizontlar için uygun bir genişliktir, ancak daha geniş lamellerin öğütülmesi daha uzun sürer. İlk frezeleme adımı için desenlerin seçilen yüksekliği, yaklaşık 6 μm'den başlayarak numune kalınlığına bağlıdır; Bunun frezeleme sırasında ayarlanması gerekebilir. Frezeleme, desenlerin dış, üst ve alt kenarlarından lamel yüzüne doğru yönlü olarak gerçekleştirilir. Bu, her iki desenin aynı anda veya sırayla öğütüldüğü paralel olarak yapılabilir, önce lamellerin üstünden ve daha sonra altından malzeme çıkarılır. - Frezelemeye ilk akımda başlayın; canlı ilerlemeyi iyon ışını görünümünde ve aralıklı olarak SEM (5 kV, 13 veya 25 pA) ile izleyin. İyon ışınının, korunan bölgenin üstündeki ve altındaki numuneyi kırıp kırmadığını kontrol edin. Değilse, daha fazla malzeme çıkarmak için dikdörtgen desenlerin yüksekliğini artırın. Lamelin üstündeki ve altındaki yüzey iyon ışını görünümünde tamamen pürüzsüz olduğunda durun.
NOT: İyon ışını, dikdörtgen desenlerin içi odak düzleminde hiçbir özellik içermediğinde, korunan bölgenin üstündeki ve altındaki numuneyi kırmıştır. Izgara çubukları gibi bazı özellikler arka planda odak dışında görünebilir. - Bir sonraki frezeleme akımına geçin (100 pA); Odaklanın ve parlaklığı / kontrastı ayarlayın. İki dikdörtgen desen arasındaki boşluğu 1,5 μm'ye düşürün ve desen yüksekliğini yalnızca öğütülmemiş malzemeyi kaplayacak şekilde azaltın. Lamelin üstündeki ve altındaki yüzey tamamen pürüzsüz olana kadar yeni akımda frezelemeye başlayın.
- 0,3 μm kalınlığa ulaşana kadar bu işlemi adım adım tekrarlayın ve iyon ışını akımını Tablo 1'deki frezeleme şemasına göre her seferinde azaltın.
- Bir veya her iki ızgarada birkaç lamel (miktar, numunenin kalınlığına ve mevcut zamana bağlıdır) 0,3 μm kalınlığa kadar öğütün, her bir lamellerin X / Y / Z konumunu kaydedin ve lamelleri son kalınlıklarına (60-200 nm) parlatmak için oturumun sonunda ~ 1-2 saat ayırın.
- Tüm ızgaranın düşük büyütmeli bir SEM genel bakışını alın ve ızgaranın önündeki lamellerden, ızgaranın arkasındaki lamellere, ızgaranın arkasındaki lamellere, (iyon ışını kaynağından en uzakta) başlayan bir parlatma rotası planlayın (Şekil 5).
NOT: Bu, ablatlanmış malzemenin cilalı lamellerin yüzeyine geri depolanmasını azaltır. - İki dikdörtgen desen arasındaki boşluğu 100-200 nm'ye düşürün ve son parlatma adımına 30 pA'lık bir iyon ışını akımında başlayın. SEM ile ilerlemeyi 2-3 kV'da izleyin (13 pA, bekleme = 300 n, 3072 x 2048, tam kare için ~ 2 s) ve kontrast SEM tarafından lamelde kaybolduğunda veya lamelin organoplatin kaplaması bütünlüğünü kaybetmeye başladığında parlatmayı bırakın.
- Izgaraları çıkarmadan önce, tüm ızgaranın düşük büyütmeli bir SEM görüntüsünü elde edin ve her lamelin görüntülerini kaydedin. Bunları daha sonra TEM'de ızgarayı çapraz kontrol etmek için kullanın. Deneyi burada duraklatın ve ızgaraları sıvı azot altında lamelli saklayın - dikkatli kullanın.
NOT: Izgaralar, kriyoFIB-SEM'den çıkarıldıktan sonra hafifçe püskürtülebilir, bu da TEM'deki sürüklenmeyi ve şarjı yüksek büyütmede sınırlamaya yardımcı olabilir, ancak çok fazla püskürtme kaplaması lamellerin içindeki biyolojik içeriği gizleyebileceğinden bu dikkatli yapılmalıdır. Bu aşamada lamelleri floresan açısından taramak mümkündür; Bununla birlikte, yeterli sinyal elde etmek, etiketli proteinin lamel kalınlığı içindeki bolluğuna bağlı olacaktır. Lamellere verilen hasarı sınırlamak ve yüzey kontaminasyonunu önlemek için ızgaraların işlenmesine büyük özen gösterilmelidir.
3. Tilt serisi edinimi ve veri işlemeye genel bakış
- Frezeleme yönünü sahnenin eğim eksenine dik olarak hizalayarak ızgaraları TEM'e yükleyin.
NOT: Izgaraların hizalanması, otomatik ızgara jantındaki işaretler kullanılarak gözle yapılır. ~ 10 ° 'lik bir hata payı kabul edilebilir, aksi takdirde lamellerin her iki tarafındaki açmanın duvarları, ızgara eğilirken lamelleri gizleyebilir. - Tüm ızgaranın düşük büyütmeli bir haritasını (~ 150x) edinin ve lamelleri bulun; Ardından, her lamelin orta büyütme haritasını (lamel boyutuna bağlı olarak ~ 1.500x) edinin ve ilgi alanlarını bulun.
- Lamel düzlemini (ızgarayı değil) optik eksene dik hale getirmek için ızgarayı ±10 ° önceden eğin
NOT: Ön eğimin yönü, düşük ve orta büyütme haritalarında lamellerin ön kenarının konumuna göre belirlenebilir (organoplatin kaplamanın üzerinde sol kısım aranır). Burada kullanılan TEM için, ızgaralar lamellerin ön kenarları haritalarda yukarı bakıyorsa +10° ön eğim gerektirir ve aşağı bakarlarsa -10° ön eğim gerektirir. Her ızgara, nasıl alındıkları ve otomatik yükleyiciye nasıl takıldıkları nedeniyle farklı olabilir. - İlgilenilen bölge için gereken görüş alanına ve çözünürlüğe izin veren bir piksel boyutunda doz-simetrik eğim serisi38'i (örneğin, 3-5°'lik bir artışla -54° ila +54°) edinin. -2 ile -5 μm arasında bir dizi bulanıklaştırma değeri kullanın. Her artışta 3-10 karelik filmler toplayın ve bunun için piksel boyutuna bağlı olarak bu parametreleri ~ 150 e-/Å2 (300 kV TEM için) toplam doz biriktirecek şekilde ayarlayın.
NOT: Bu bölgeler sürüklenebileceğinden lameldeki çatlaklardan kaçınılmalıdır. Yüzey kontaminasyonundan da kaçınılmalıdır, çünkü yüksek eğimde ilgi alanını veya odak alanını gizleyebilir. - Motion, MotionCor239 gibi bir program kullanarak filmleri düzeltir. Düzeltilmiş görüntülere (birikmiş e-/image)40 bir doz ağırlıklandırma filtresi uygulayın ve CTFFIND441 gibi bir program kullanarak her görüntünün bulanıklaşmasını tahmin edin.
- Eğim serilerindeki görüntülerin hizalamasını ve açısal ilişkisini hesaplamak için etomo (IMOD)42 gibi bir programda güvensiz hizalama (yama izleme) kullanın.
- Hizalama ve döndürme bilgilerini, defokus değerleriyle birlikte üç boyutlu CTF düzeltmesi uygulayabilen bir programa, örneğin NovaCTF43'e girin. Aşağı akış analiziyle ilgili binning faktörlerine sahip çıktı tomogramları elde etmek için düzeltilmiş bir tomogram hesaplayın.
- Rekonstrüksiyonları analiz edin ve filtreleme, segmentasyon veya alt tomogram ortalaması gibi herhangi bir aşağı akış işlemine hazırlanın.
Sonuçlar
P. falciparum schizonts'un dalma dondurma için hazırlanması
Bileşik 2 ve E64 inhibitörleri, çıkışın farklı aşamalarında şizontları durdurmak için kullanılır ve sonraki çalışmalar için zenginleştirilmiş bir schizonts popülasyonu oluşturur. Bu önemlidir, çünkü tamamlayıcı bir korelasyon tekniği olmadan, spesifik hücre altı hedefleri veya hücre tiplerini frezelemek, süreç esasen kör olduğu için zordur. Bileşik 2, vakuol rüptüründen önce çıkışı durduran bir protein kinaz inhibitörüdür. Schizontlar 4 saat boyunca bileşik 2 üzerinde senkronize edilebilir, daha sonra inhibitörü çıkarmak için bileşik 2-free media ile yıkanabilir, bu noktada schizonts yaklaşık 30 dakika sonra olgunlaşır ve çıkar. Alternatif olarak, bileşik 2 senkronize şizontlar, geri dönüşümsüz bir geniş spektrumlu sistein proteaz inhibitörü olan E64'e yıkanabilir ve vakuol rüptürü noktasından sonra, ancak konakçı hücre rüptüründen önce çıkışı durdurmak için yaklaşık 1 saat boyunca inkübe edilebilir. Tedavi edilen şizontların morfolojisi ve homojenliği, dalma donmasından önce Giemsa boyalı kan yaymaları ile kontrol edilmelidir (Şekil 1). Schizontlar, bu yayında açıklanan yöntem kullanılarak bu durumlardan herhangi birinde dondurulmuş olarak daldırılabilir.

Resim 1: Giemsa boyalı kan yaymaları ile bileşik 2 ve E64 duraklamış P. falciparum schizontlarının morfolojisi. (A) Bileşik 2'nin varlığında, parazitoforöz vakuol (PV), tek bir hemozoin kristali kümesi (koyu kahverengi daire) ile merozoitlerle (mor daireler) yoğun bir şekilde paketlenir. PV ile konakçı kırmızı kan hücresindeki (hRBC) (gri bant) çevresindeki hemoglobin arasındaki sınır, hRBC zarının yanı sıra iyi tanımlanmıştır. (B) E64 varlığında, PV membranı yırtılır ve merozoitler hRBC içinde yayılır. Her şizont hala tek bir hemozoin kristali kümesi içerir. Konakçı hücre zarı sızdırmaz ve kısmen çökmüştür, bu nedenle hücre periferisinde hemoglobin görünmez ve hRBC zarının sınırı kolayca görünmez. Ölçek çubuğu, 5 μm. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
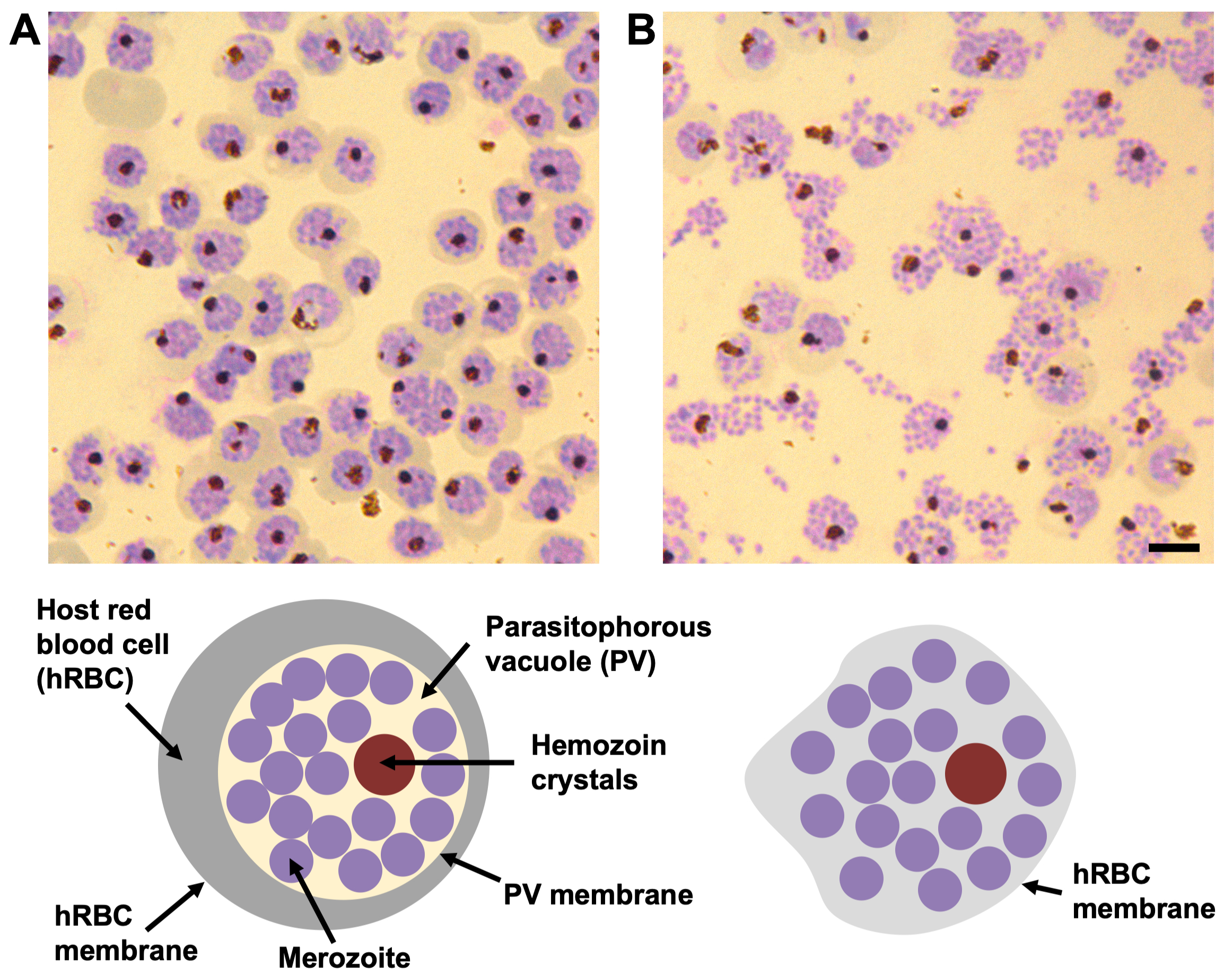{kind=link}
Dalma dondurma optimizasyonu
Altın ve bakır 200 örgü ızgaralar üzerinde 2/2, 3/3, 3.5/1 ve 5/2 (kare) delikli karbon dahil olmak üzere P. falciparum ile enfekte kırmızı kan hücreleri için lekelenme koşullarının optimizasyonu sırasında farklı delik boyutlarına sahip bir dizi ızgara denenmiştir. 2/4 delikli karbon filmli 200 örgülü bakır bulucu ızgaraları, uzun vitreus lamelleri öğütmek için uygun kalınlıkta bir hücre tabakası sağlar. Daha büyük veya daha küçük delikler genellikle sırasıyla çok ince veya kalın hücre katmanlarıyla sonuçlanır (Şekil 2A-C). 2/4 delikli karbon ile, şizontlar ızgarayı arkadan (karbon olmayan taraftan) lekeleyerek 2 μm deliklerden çekilir, bu da hücrelerin karbon filmin üstünde ve altında dışarı çıkmasına neden olur. Delikler arasındaki 4 μm karbon gerilmesi, ortaya çıkan lamellerin çoğunun ortasından geçen ve güç katan bir karbon şeridi ile sonuçlanır. Bulucu ızgaraları korelasyon ve eleme amaçları için en uygun olanıdır44, ancak ağ tasarımındaki sayıların / harflerin frezeleme alanlarını engelleyeceğinden çok büyük olmadığından emin olmak için özen gösterilmelidir.
Manuel bir pistondaki lekelenme süresi yaklaşık 20 sn'dir, ancak lekelenmeyi durdurmak için kesin nokta, ızgaradan çekilen sıvı damlacığı filtre kağıdına yayılmayı bıraktığında gözle değerlendirildi. Lekeleme işlemini başlatmak için damlanın yüzey gerilimini kırmak için yırtık bir kenar gerekliydi. Bu çalışmada otomatik bir daldırma dondurucu kullanılmamıştır, ancak bu örnek için makul bir başlangıç noktası, manuel bir piston için kullanılanla aynı hacimde hücre ve ızgara tipi kullanmak ve ızgaraları yüksek nem (~% 70) ve ortam sıcaklığı (~ 25 ° C) olan koşullarda arkadan lekelediğinizden emin olmak olacaktır. Blotlama süreleri ve koşullarının, kullanılan belirli otomatik sistem için optimize edilmesi gerekir.
P. falciparum schizonts'un dalma dondurulmuş ızgaraları, TEM yerine kriyo-aşama ile donatılmış bir ışık mikroskobu ile tarandı, çünkü numune elektronlara aktarılamadı. Daha ince numuneler için, ızgaralar, ulusal bir tesise erişim için bir ön koşul olabilecek cryoFIB-SEM'e numune transferinden önce TEM (~ 150x büyütmede tüm ızgara atlası) tarafından taranabilir. Buz kalınlığının gradyanına ızgara boyunca ve ayrıca bireysel ızgara kareleri içinde özel dikkat gösterilmelidir. İyi ızgara kareleri bir hücre kalınlığında olmalı veya karbon filme merkezlerinden çarpmalıdır (Şekil 2C). Bu, ızgara karesinin kenarındaki ızgara çubuklarına karşı daha kalın buzda frezelenmeyi önler ve iyon ışınının hücre tabakasının üstünden ve altından geçmesini sağlayarak kama yerine serbest bir lamel üretir. Tekrarlanabilir lekeleme ve daldırma dondurma koşulları optimize edildikten sonra, FIB frezelemeden önce genellikle tarama gerekli değildir.

Şekil 2. Dalma dondurulmuş P. falciparum schizonts ızgaralarındaki hücre dağılımının kriyo-ışık mikroskobu ile analiz edilmesi. (A) Kriyo-ışık mikroskobu ile bir ızgara karesi boyunca çok kalın olan buzun hücreleri ve karbon filmini gizleyen bir örneği. Ölçek çubuğu, 10 μm. (B) Delik boyutunun (5/2 kare delikli karbon filmli 300 örgü bakır ızgara) hücreler için çok büyük olmasına bir örnek, buzsuz boş bölgelerle çevrili çok ince bir biyolojik malzeme tabakasına neden olur. Bunun gibi ızgaralar çok kısa, kararsız lameller üretir. Ölçek çubuğu, 10 μm. (C) 2/4 delikli karbon filmli 200 örgülü bakır ızgarada iyi hücre dağılımına bir örnek. Bu şizontlar E64 inhibitörü ile tedavi edildi. İyi tanımlanmış bir periferiye sahip büyük hücreler (kırmızı kutu, ~ 5 μm çapında), hala sağlam bir vakuol zarına sahip olan enfekte kırmızı kan hücreleridir. Küçük hücre kümeleri (mavi kutu, ~ 1 μm), kısmen çökmüş bir konakçı kırmızı kan hücresinde bulunan bireysel merozoitlerdir. Her şizontun merkezinde, hemozoin kristallerinin konumunu gösteren siyah bir nokta vardır (Ek ayrıntılar için Şekil 1'e bakınız). Hücre morfolojisindeki farkı, kriyoFIB-SEM'in içinde bir kez görmek kadar kolay değildir; Bu nedenle, kriyo-ışık mikroskobu ile ön tarama, tekrarlanabilir lekelenme koşullarına ulaşılana kadar faydalıdır. Izgara çubuklarının (beyaz kesikli alanlar) yanındaki ızgara karesinin kenarlarının etrafındaki hücre kapsamı frezeleme için çok kalındır. Izgara karesinin ortasındaki daha ince bölge (sarı kesikli alan), lamelleri çevreleyen hücre tabakasına genişleterek öğütmek için ideal bir yerdir. Ölçek çubuğu, 6 μm. (D) KriyoFIB'ye özgü bir otomatik ızgara jantının, biri kesme bölümünde (siyah braket) ve diğeri tam tersi olmak üzere saat 12 ve saat 6 konumunda iki siyah işaretli görüntüsü. Siyah ok, frezeleme yönünü temsil eder. (E) Izgaralar otomatik yükleyici kasetine yüklendiğinde, işaretlerin yükleme cımbızının her iki tarafına eşit uzaklıkta olması gerekir, bu da lamellerin sahnenin eğim eksenine dik olarak uzanmasına neden olur. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
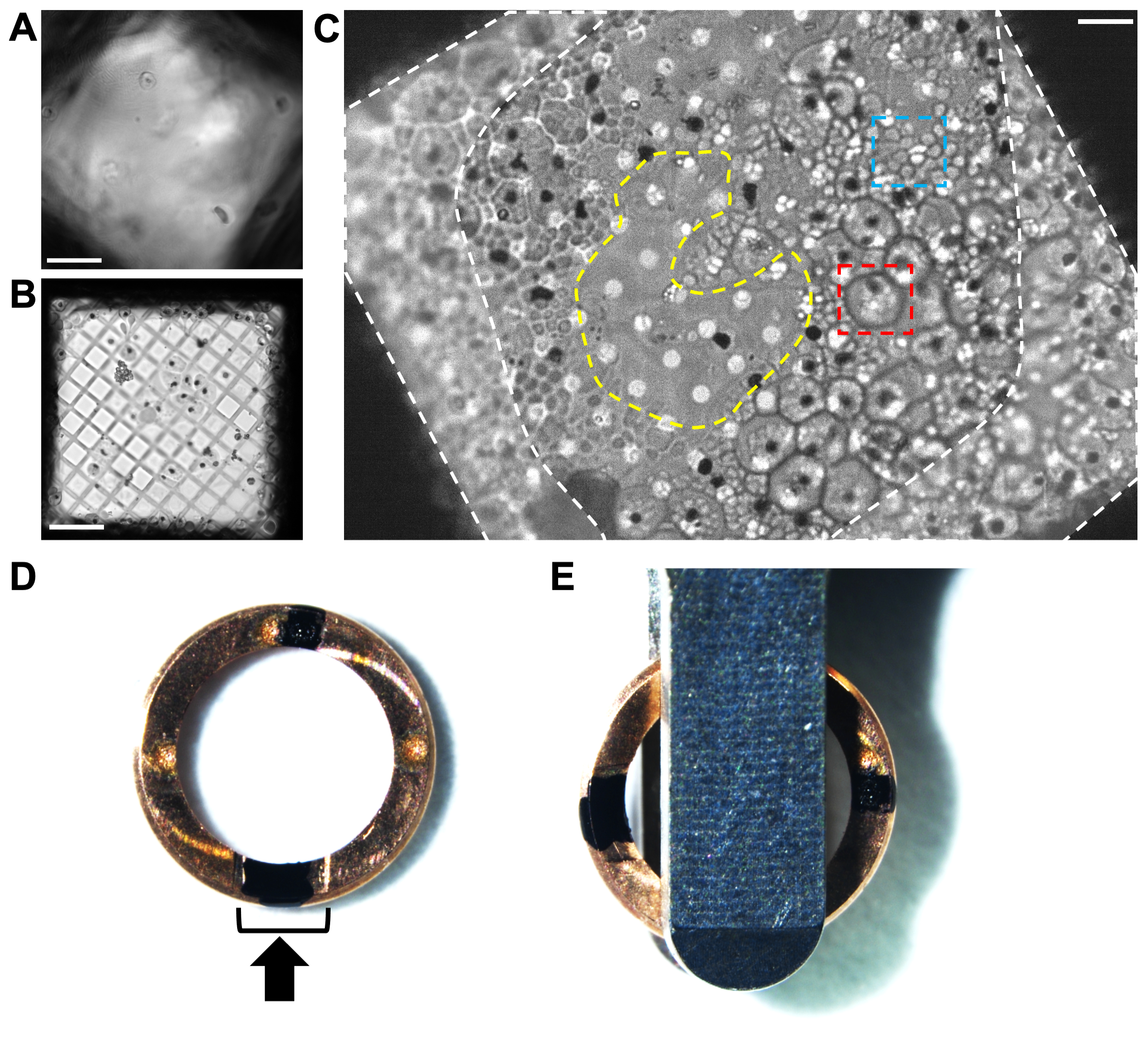{kind=link}
Tomografi için cryoFIB'e özgü otomatik ızgara jantlarının işaretlenmesi
Tipik olarak, ızgaralar, kullanımı kolaylaştırmak ve sertlik sağlamak için frezelemeden önce otomatik ızgara jantlarına kırpılır, bu da sonraki transfer adımları sırasında lamelleri hasardan korur. CryoFIB'e özgü otomatik ızgara jantları, frezeleme sırasında ızgara yüzeyinin daha fazlasına erişmeye yardımcı olmak için kesme özelliği ile tasarlanmıştır. Frezeleme yönünün TEM'in eğim eksenine dik olarak yönlendirilmesi önemlidir, böylece eğim serisi edinimi, lamelleri uzunluğu boyunca döndürerek devam eder. Bu, lamelleri çevreleyen açmanın yüksek duvarlarının, ızgara eğilirken biyolojik bilgiyi gizlememesini sağlar.
Tipik olarak, otomatik ızgara jantı, kriyoFIB-SEM mekiği içinde ve daha sonra TEM'i yüklerken görsel hizalamaya yardımcı olmak için işaretlenir. Bu örnekler için, saat 12 ve saat 6 pozisyonlarında, biri klips halkasının kesik bölümünün ortasında, ikincisi ise tam tersi olmak üzere silinmez bir işaretleyici ile iki işaret uygulanmıştır (Şekil 2D). TEM'e yükleme yaparken (bkz. Malzeme Tablosu), her iki işaret de yükleme cımbızının her iki tarafında görünür olmalı ve cımbızın kenarına 90° hizalanmalıdır (Şekil 2E). İyon ışınına yakın bazı mürekkepler frezelemeye müdahale edebileceğinden, üreticinin otomatik ızgara jantlarındaki oyulmuş noktaları kullanarak ızgaraların hizalanmasını önerdiğine dikkat edilmelidir.
Kırpılmış ızgaralar, bir kriyo-aşama için modifiye edilmiş bir kasetle ışık mikroskobu ile taranabilir, bu da kırpma işleminin ızgaranın karbon filmini tahrip etmediğini kontrol etmek için faydalı olabilir. Numune kasetine bağlı olarak, kırpılmamış ızgaralar da frezelenebilir, ancak lamelleri kıracağı için ızgaranın herhangi bir bükülmesini sınırlamak için cryoFIB-SEM'den TEM'e aktarım sırasında büyük özen gösterilmelidir. Kırpılmamış ızgaralar, yandan girişli kriyo tutucular tarafından TEM'e yüklenebilir, ancak frezelemeden sonra kırpma yapılırsa lamellerin kırılma olasılığı yüksektir.
Organoplatin kaplama
Organoplatin kaplama, kriyoFIB-SEM'e yüklendikten sonra bir veya her iki ızgarada gerçekleştirilir. Gaz enjeksiyon sisteminin (GIS) iğnesi, ızgaranın yüzeyi boyunca belirli bir süre ısıtılmış bir kaynaktan organoplatin buharı akışını yönlendirmek için numunenin üzerindeki odaya yerleştirilir. Buhar soğuk yüzeylerde yoğunlaşır ve katı bir tabaka oluşturur (~ 2 μm kalınlığında). Bu katın bütünlüğü, düzgün ince lamellerin öğütülmesini sağlamak için kritik öneme sahiptir. Organoplatin kaplama için en uygun uygulama koşulları genellikle cihazın üreticisi tarafından önceden belirlenir, ancak yine de bazı optimizasyonlar gerekebilir. Çoğu sistem, CBS iğnesini frezeleme yönüne yakın veya frezeleme yönüne dik olarak hizalar, odadaki portların geometrisine ve sahnenin dönme sınırlarına bağlı olarak. Izgaranın kaplanması, küçük bir bölgenin frezelenmesi, sahnenin eğilmesi ve katın kalınlığının SEM ile ölçülmesi ile farklı ayarlar denenebilir.
KriyoFIB-SEM'in kendisinin kurulumunun yanı sıra, 1) numunenin topografyası, 2) ızgaradaki yüzey kontaminasyonu ve 3) CBS iğnesinden buhar akışının tekrarlanabilirliği dahil olmak üzere organoplatin kaplamanın uygulanmasını bir dizi başka faktör etkileyebilir. CBS akışı yönlü olduğundan, düzensiz topografya, hücrelerin gölgesindeki bölgelerin veya yüzey kontaminasyonunun kaplanmamış olmasına veya daha ince bir kaplamaya sahip olmasına neden olabilir. Bu, parlatma sırasında organoplatin tabakasının çökmesine neden olabilir (Şekil 3). Frezelenecek bir alan seçerken, çevredeki topografyaya dikkat etmek gerekir, örneğin, büyük yüzey kirliliği, hücre kümeleri veya ızgaranın yüzeyinden birkaç ızgara karesine kadar dışarı yansıyan kırık karbon, buhar akışını engelleyebilir ve bir lamelleri zayıflatabilecek daha ince organoplatin gölgesi oluşturabilir. Ek olarak, lamelmanın ön kenarına yakın çok küçük yüzey kirliliği parçacıklarından da kaçınılmalıdır, çünkü frezeleme sırasında patlayabilirler, parlatma sırasında lamelde bir delik oluşmasına neden olabilecek zayıflamış bir çıplak buz yaması bırakırlar. Son olarak, frezeleme başladıysa ve organoplatin kaplama dengesiz görünüyorsa, tekrar daha uzun süre kaplayın veya bir yedek ızgaraya geçin.

Şekil 3: İnce, eşit öğütülmüş lameller elde etmek için kaliteli bir organoplatin kaplama kritik öneme sahiptir . (A) Organoplatin kaplamanın (OP, sarı) çok ince uygulandığı, lamelmanın ön kenarında parlatma (yeşil daire) ve lamelin tüm genişliği boyunca düzensiz frezeleme (çizgiler) sırasında gelişen bir deliğe yol açan bir lamelin ön kenarının mikrografisi. Organoplatin yüzeyi, iyon ışını tarafından kesilmiştir ve kaplamanın ön kenarının arkasında bir malzeme spreyine yol açmıştır (sarı kesikli çizgi). (B) Organoplatin kat (OP) daha kalın bir şekilde uygulanmıştır, bu da daha eşit şekilde inceltilmiş bir lamel ile sonuçlanır. Katın bütünlüğü, lamelin tüm genişliği boyunca korunur ve organoplatin kaplama ile vitrifiye biyolojik materyal arasındaki arayüz iyi tanımlanmıştır (sarı kesikli çizgi). Karbon tabakası, lamelin (turuncu) arkasından geçerken görülebilir. Ölçek çubukları, 1 μm. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}
KriyoFIB-SEM'de ızgara kalitesinin değerlendirilmesi ve frezeleme prosesi
Izgaralar kriyoFIB-SEM'e aktarıldıktan sonra, karbon filmin bütünlüğü ve hücrelerin ızgaralardaki dağılımı SEM ile taranabilir (Şekil 4A-C). Buz gradyanları, ızgara çubuklarının konumları ve ızgara numaralarının bulucu ızgaralarındaki konumları, 30 kV'da düşük büyütmeli SEM ile kontrol edilebilir (Şekil 4B), ancak frezeleme konumlarını yüksek büyütmede konumlandırırken ve yüzey topografyasından kontrastı artırmak için frezeleme işlemini izlerken voltaj 5 kV'a düşürülmelidir.

Şekil 4: Izgara kalitesinin değerlendirilmesi ve kriyoFIB-SEM'de öğütülecek alanların belirlenmesi . (A) SEM aracılığıyla 5 kV'da bir ızgaranın düşük büyütmeli bir genel bakışı. Otomatik ızgara jantının kesit bölümü, görüntünün altında görülebilir. Ölçek çubuğu, 0,5 mm. (B) SEM tarafından 30 kV'da aynı ızgara, daha kalın buz (daha koyu ızgara kareleri) ve daha ince buz (daha açık ızgara kareleri) alanlarını gösterir. İç kısım, 30 kV'de görülebilen bulucu ızgarasındaki numaralandırmayı gösteren oklarla birlikte kutudaki bölgeyi gösterir. Ölçek çubuğu, 0,5 mm. (C) Hücrelerin karbon film üzerindeki dağılımını ve ızgara çubuklarının konumunu değerlendiren iki ızgara karesine orta büyütmeli bir genel bakış. Ölçek çubuğu, 50 μm. (D) Yüksek büyütmede ilk kesim için frezeleme desenlerinin düzenlenmesi (1,5 pA ve 30 kV'de iyon ışını görünümü). Kırmızı bölge (3 μm kalınlığında) korunurken, sarı bölgeler iyon ışını tarafından ablatlanacaktır. Beyaz kesikli çizgi, son lamelin konumunu gösterir. Ölçek çubuğu, 10 μm. (E) Cilalanmış 200 nm kalınlığında (10 μm genişlik x 15 μm uzunluğunda) SEM aracılığıyla 3 kV'da yüksek büyütmeli bir görünüm. Lamel içindeki kontrast kaybı 3 kV'da uygun kalınlığa ulaşıldığını gösterir. Parlak beyaz ön kenar, frezelemeden önce CBS aracılığıyla ızgaraya uygulanan kalan organoplatin tabakadır. Ölçek çubuğu, 5 μm. (F) (E)'den aynı lamel, iyon ışını kullanılarak 30 kV ve 1,5 pA'da görüntülendi. Siyah kare (beyaz ok) boyunca ince beyaz çizgi, lamelin en ön kenarında kalan organoplatin kattır. Ölçek çubuğu, 10 μm. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
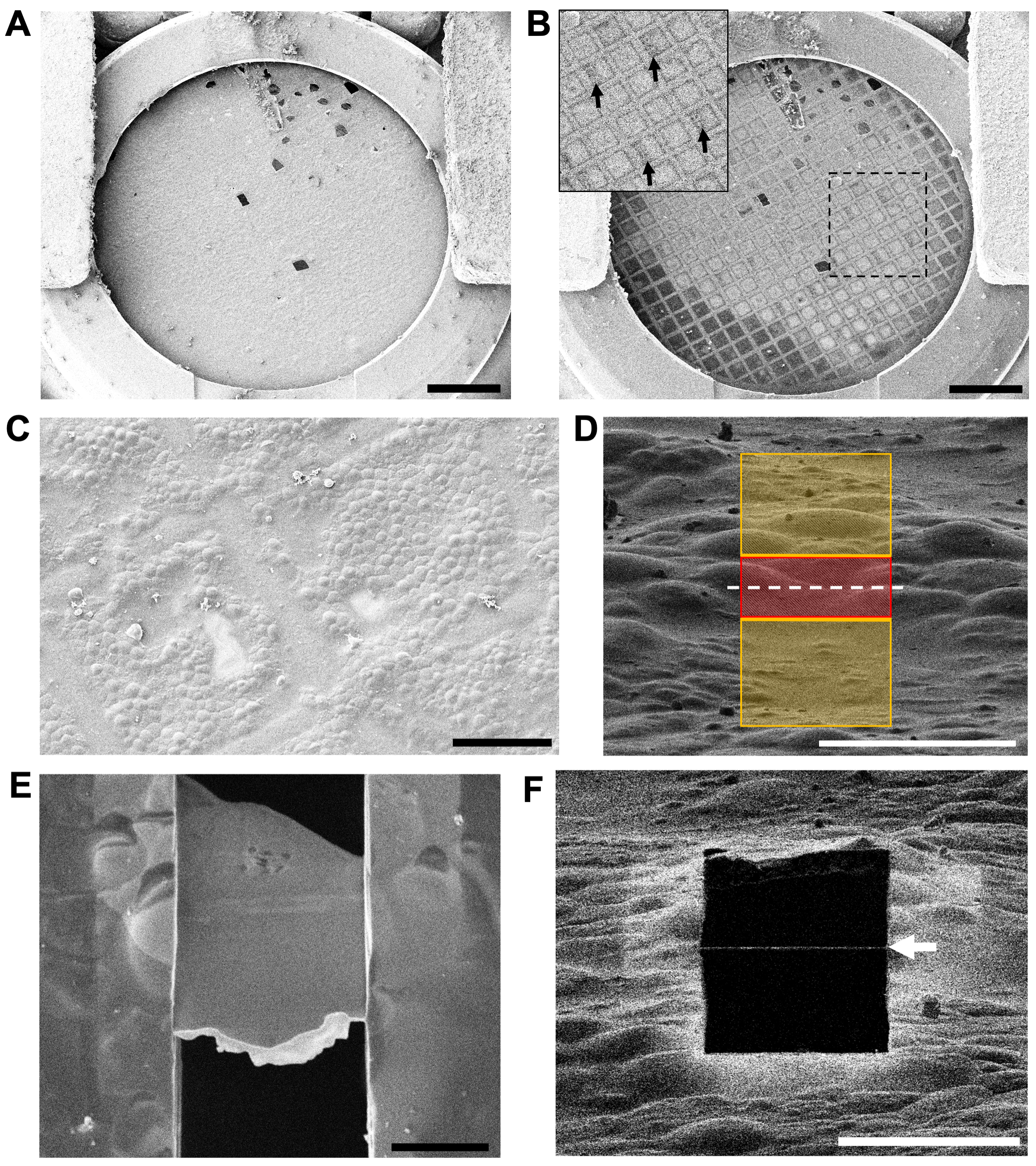{kind=link}
Frezelenecek bir bölge, iyon ışını görünümünde yüksek büyütmede (şizontlar için ~ 7.000x) korunan bir bölgenin her iki tarafına bir çift dikdörtgen desen yerleştirilerek seçilir (Şekil 4D). Frezeleme bölgesine yakın hiçbir yüzey kontaminasyon partikülünün bağlanmaması çok önemlidir, çünkü bunlar koruyucu organoplatin kaplamanın uygulanmasını engellemiş olabilir. Bölgenin topografyasının, nihai kalınlığına ulaşıldıktan sonra lamelin kenarlarını desteklemeye uygun olması da önemlidir.
Sıtma şizontları için (hücre boyutu: ~5 μm çap x 2 μm kalınlığında, disk şeklinde) 7-20 μm genişliğinde lameller öğütülebilir. Hücre tabakası uygun şekilde kalınsa, lameller genellikle ~ 10-15 μm uzunluğunda olacak ve karbon tabakasının üstünde ve altında birden fazla hücre yakalayacaktır (Şekil 4E-F). 8 saatlik bir seansta 5-10 lamellerin öğütülmesi beklenebilir (6-7 saat frezeleme ve 1-2 saat parlatma). Bu, numunenin kalınlığına ve lamellerin genişliğine bağlı olarak değişecektir, daha kalın numuneler ve daha geniş lamellerin öğütülmesi daha uzun sürecektir. Bir dizi lamel oluşturmak için sadece bir avuç iyi ızgara karesi gerektirdiğinden hasarlı ızgaralar bile öğütülebilir (Şekil 5A). Ek olarak, numune beklenenden daha inceyse, örneğin, aşırı lekelenme veya kültürün hematokritindeki varyasyon nedeniyle, daha kısa lameller nispeten hızlı bir şekilde öğütülebilir; ancak, daha kısa uzunlukları TEM'de veri toplama için kullanılabilir alanı sınırlayacaktır (Şekil 5B).

Şekil 5: Frezeleme sırasında nihai lamel kalınlığına ne zaman ulaşıldığının belirlenmesi. (A) Kırpma sırasında karbon hasarını gösteren 5 kV'da SEM tarafından bir ızgaranın düşük büyütmeli genel bakışı. Hasar görmemiş alanlar, aşırı lekelenme nedeniyle çok ince numune içeriyordu; Bununla birlikte, kriyoFIBSEM'de tam günlük bir oturumda bu ızgarada (beyaz kesikli anahat içindeki bölge) altı lamel öğütmek hala mümkündü. Ölçek çubuğu, 0,5 mm. (B) Bu ızgaradan (3 kV'de SEM) üretilen kısa bir lamel (~ 10 μm genişliğinde x 3 μm uzunluğunda, organoplatin tabakası hariç), yine de eğim serilerinin toplanacağı iki bölge sağladı. Ölçek çubuğu, 25 μm. (C) Son parlatma adımı sırasında lamellerin inceltildikçe (soldan sağa hareket ettikçe) kontrastın nasıl kaybolduğunu gösteren bir dizi lamel görüntüsü (3 kV). Tüm görüntülerde lamelin ortasındaki koyu siyah çizgi, ızgaradan bir karbon film şerididir. Bu bölgenin önündeki hücreler karbon filmin üzerinde vitrifiye edildi ve bu bölgenin arkasındaki hücreler karbon filmin altında vitrifiye edildi. Lamelin ön kenarının sol tarafındaki organoplatin kaplama yapısal bütünlüğünü kaybetmek üzereyken frezeleme durduruldu. Bu durma noktası, tüm lamel eşit bir kalınlığa getirilmeden önceydi, bu yüzden lamelin arkasında hala daha yüksek kontrastlı bir malzeme var. (D) (A)'da gösterilen ızgara üzerinde öğütülmüş lamelleri temel alan bir parlatma yolu örneği. Bir parlatma rotası, FIB kaynağına yakın lamellerde başlamalı ve öğütülmüş malzemenin lamellerin yüzeyine yeniden birikmesini sınırlamak için FIB kaynağından uzaklaşmalıdır. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}
Parlatma sırasında, bir lamelin nihai kalınlığı, öğütülen bölgedeki numunenin yapısına, organoplatin kaplamanın bütünlüğüne ve mevcut zamana bağlı olacaktır. İdeal olarak, numune, 3 kV'da SEM tarafından lamelin tüm yüzeyi boyunca kontrast kaybolana kadar inceltilmelidir, bu da eşit elektron saydam ve yaklaşık 150-200 nm kalınlığında olduğunu düşündürmektedir (Şekil 5C). Bununla birlikte, organoplatin tabakası bir delik geliştirirse veya lamel bükülmeye başlarsa, bu noktadan önce frezelemeyi durdurmak gerekebilir. Bu durumda, lamel hala ön tarafta yeterince ince olabilir ve tomografi için yararlı olmaya devam edebilir. Tersine, lamel sabit görünüyorsa, kontrast aşaması kaybını aşmak mümkündür, bu da frezeleme desenlerini birbirine yaklaştırarak (~ 100 nm veya daha az) daha da inceltilir. Bu, aşağı akış iş akışı için gerekirse hangi kalınlığa bağlı olacaktır. İyon ışını kaynağına daha yakın başlayıp uzaklaşan bir parlatma rotası planlamak için düşük büyütmeli bir SEM görüntüsü gereklidir (Şekil 5D). Parlatma işleminin yönü, ablatlanmış malzemenin daha önce bitmiş olan lamel üzerine yeniden birikmesini önlemek için önemlidir.
Eğim serisi verilerinin toplanması ve işlenmesi
TEM'e yüklendikten sonra, düşük büyütmeli bir tam ızgara montajı (~ 150x), frezeleme sonunda çekilen düşük büyütmeli SEM görüntüsüyle ilişkilendirilebilen lamellerin konumlarını tanımlayacaktır. Dalma donmuş şizontlar için, ızgaranın çoğu elektronlara karşı şeffaf değildir, bu nedenle lamel pozisyonları siyah bir arka plan üzerinde beyaz çentikler olarak görünür (Şekil 6A). Lamellerin mikroskobun eğim eksenine göre açısı not edilmelidir, çünkü dikeyden ~ 10 ° 'den daha fazla uzakta olması eğim serisi edinimini zorlaştırabilir. Her bir lamelin bulunduğu yerdeki orta büyütmeli bir montaj (~ 1.500x), biyolojik içeriğe genel bir bakış sunacak ve transfer hasarı, kristalin buz veya aşırı yüzey kirliliğini kontrol edecektir (Şekil 6B-D). Bu, hiçbir yüzey kontaminasyonunun edinme veya odak alanını gizlemediğinden emin olmak için yüksek eğimde de kontrol edilmelidir. Bir edinme bölgesi seçmek sadece biyolojik özelliklere değil, aynı zamanda çevredeki bölgedeki buzun yapısal bütünlüğüne de bağlı olacaktır, örneğin, bu bölgeler sürükleneceği için çatlaklardan kaçınmak veya bir lamelin değişken kalınlığa sahip olacağı aşırı perdelenmeye sahip alanlar. Bir eğim serisi elde etmeden önce, lamellerin düzlemini (ızgarayı değil) optik eksene dik hale getirmek için ızgaraya ±10 ° 'lik bir ön eğim uygulanır. Ön eğimin yönü, orta büyütme montajında lamellerin ön kenarının konumu (organoplatin kaplama üzerinde sol tarafa bakarak) ile belirlenebilir. Burada kullanılan TEM için (300 kV Titan Krios), lamellerin ön kenarları haritada yukarı baktığında, bu +10 ° 'lik bir ön eğim gerektiriyordu ve eğer aşağı işaret ediyorlarsa, bu -10 ° 'lik bir ön eğim gerektiriyordu. Her ızgara, cımbızla alındıkları ve otomatik yükleyiciye yerleştirildikleri yön nedeniyle farklı olabilir (sola veya sağa bakan kesik bölüm, 180° dönüş üretir). Son bir husus piksel boyutudur. Rutin olarak, eğim serileri, ilgilenilen özelliğin boyutu, elde edilen tomografik verilerin hedef çözünürlüğü ve lamelin yüzey alanı dikkate alınarak 2,5-7 ş/ piksel civarında toplanır ve bu da edinme alanının boyutunu sınırlayabilir. Yüksek çözünürlüklü bilgi elde etmek için daha küçük bir piksel boyutu kullanmak mümkündür ve bu örneklerde başarıyla kullandığımız en küçük piksel boyutu 1,4 ş/ pikseldir (veriler gösterilmemiştir). Sürüklenme, daha küçük bir piksel boyutunda daha belirgin olacaktır ve yalnızca çalışmada çözünürlüğü en üst düzeye çıkarmanın önemli olduğu ince (<100 nm) lameller için gerçekten uygundur.

Şekil 6: Eğim serisinin elde edileceği erişilebilir ilgi bölgelerinin seçilmesi (A) Düşük büyütmeli bir TEM haritasının, siyah bir arka plan üzerinde altı beyaz çentik (kırmızı oklar) olarak görünen lameller içeren bir bölgeyi gösteren bir bölümü. Beyaz kareler kırık karbon filmidir. (B) Hasarlı ve saptırılmış bir lamelin orta büyütme haritası. Lamelin (FE) ön kenarında organoplatin kaplama kopmuştur (1). Lamelin merkezinde berrak bir kristal buz parçası vardır (2). İyon ışını, lamelin (BE) arka kenarından geçemedi, çünkü buz ızgara çubuklarının yanında çok kalın. Lamelin sadece küçük bir kısmı BE'deki (3) çevreleyen buzdan arındırılmıştır ve bir kama oluşturur. Kalınlık ayrıca rafların lamellerin (4) üzerinde kesilmesine neden olmuştur, bu da büyük olasılıkla TEM'de yüksek eğimde hücrelerin görünümünü engelleyecektir. (C) TEM'de görülen bütün bir lamelin orta büyütmeli montajı. Lamellerden eğim serileri toplanırken kaçınılması gereken tipik problemler veya bölgeler şunlardır: yüzey kirliliği (SC) ile kaplı, organoplatin kaplama (OP, sarı) ile kaplanmış, çatlakların yakınında (CR ve siyah oklar), biyolojik materyaldeki yoğunluk değişiklikleri nedeniyle perdeleme ile (CU ve mavi braketler içindeki bölge) ve organoplatin kattaki kırılmalarla zayıflamış alanlar (yeşil daire). Eğim serisi alımı için erişilebilen tek bölge, iki siyah kesikli kutunun (1 ve 2 etiketli) içindeki alanlardır. Yüzey kirliliğinin ilgili bölgeyi veya netleme alanını gizlemediğinden emin olmak için görüntü yüksek eğimli olarak kontrol edilmelidir. (D) Çok daha temiz bir lamelin orta büyütmeli montajı, ancak parlatma sırasında organoplatin katın (yeşil daire) incelmesinden kaynaklanan çatlaklara (CR) sahiptir. Burada, ilgilenilen bölge, lamel içinde gözlenen hücre tipi ile vurgulanır; bu durumda, karbon tabakasının (turuncu) arkasında, lamelin arkasına (siyah kesikli kutu) doğru konumlandırılmış bireysel merozoitlerdir. Ölçek çubukları, 3 μm. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 7: FIB ile öğütülmüş lamelden kriyo-ET verilerinin elde edilmesi. (A) Yakın zamanda bir P. falciparum merozoit tarafından istila edilmiş bir kırmızı kan hücresi içeren bir lamel bölgesinin mikrografisi. (B) 'de aynı görüntü, kırmızı kan hücresinin sınırını (kırmızı), bir dizi çift duvarlı hücre içi vezikülü (sırasıyla iç ve dış zarlar için mor ve mavi) ve konakçı hücreden türetilen ikinci bir zarla (sarı) çevrili merozoiti (yeşil) göstermek için ek açıklamalıdır. Kara kutu, bir eğim serisinin edinildiği alanı gösterir. Ölçek çubuğu, 500 nm. (C) XY düzleminde, merozoitin apikal ucunda ve (D) ek açıklamasında elde edilen 8x bağlanmış bir tomogramdan (2.4 ş/ piksel) görüntülenen ortalama 20 merkezi dilim, hücreyi çevreleyen iki zarı (yeşil ve sarı) ve elektron yoğun mantar şeklindeki bir özellik (mor) ile ilişkili olan merozoitin tepesinde (mavi) istiflenmiş dört zarı gösterir. Kırmızı bir ok, tepedeki membran yığınlarının birleşme noktasını ve mantar şeklindeki özelliği gösterir (ayrıca F, i bölümünde de gösterilmiştir). Siyah bir ok, merozoit plazma zarı ile parazit içindeki çok katmanlı veziküllerden biri arasındaki füzyon olayını gösterir (ayrıca bölüm F, ii'de de gösterilmiştir). Konakçı kırmızı kan hücresi zarı (kırmızı) gösterilir ve siyah kesikli bir çizgi, (E) 'de gösterilen XZ düzleminde görüntülenen bir kesitin konumunu gösterir. Enine kesitteki (E) özellikler renklidir ve membranlara işaret eden renkli oklarla (D) parçasındaki ile aynı şekilde etiketlenir. Siyah bir ok, frezelemeden sonra lamele uygulanan bir püskürtme katının konumunu ve belirtilen lamellerin kalınlığını gösterir. Parçalar için (C-E), ölçek çubuğu, 500 nm. (F) (C) bölümündeki kırmızı ve siyah ok başlıkları ile gösterilen, merozoitin (i) tepesindeki membran yığınının lipit çift katmanlarındaki tanımı ve çok katmanlı vezikül ile merozoit plazma zarı arasındaki füzyon olayını gösteren özelliklerin daha ayrıntılı bir görünümü. Ölçek çubuğu, 75 nm. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
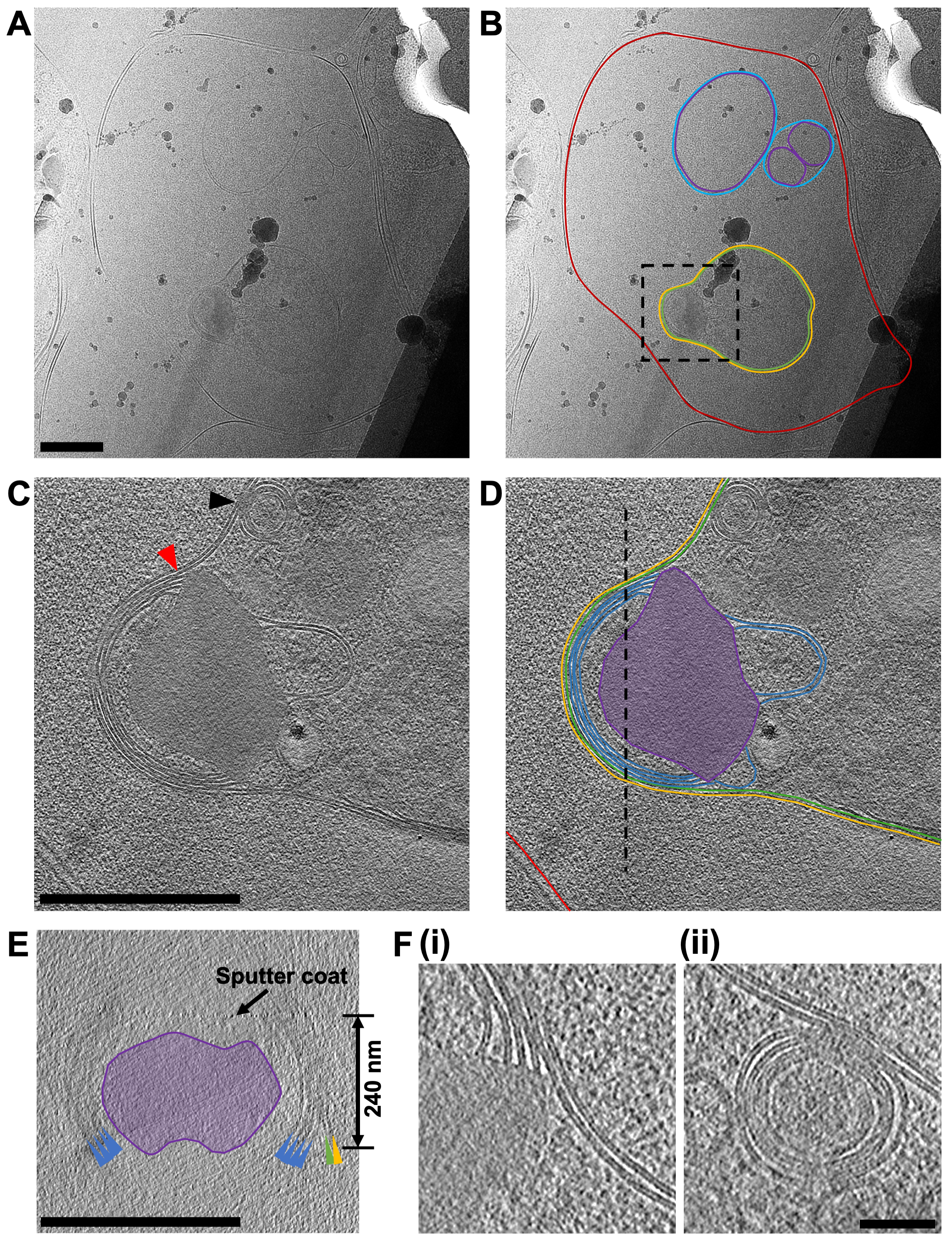{kind=link}
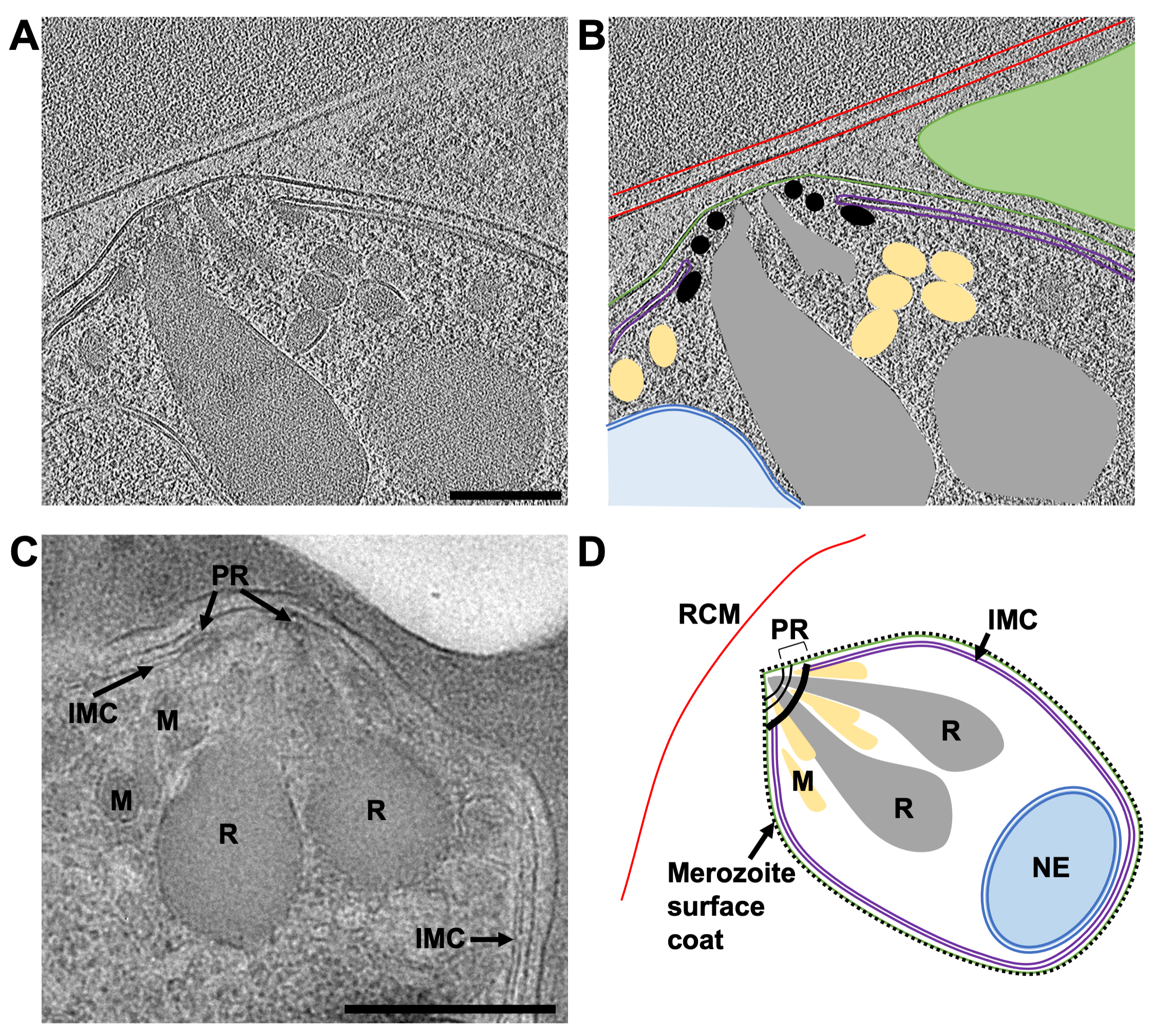
Çalışmanın ana odağı, merozoit çıkış yolunu incelemektir. P. falciparum Ancak, çalışmada kullanılan hücre popülasyonunun hiçbir zaman tamamen homojen olamayacağı göz önüne alındığında, genellikle lameller içindeki hücre gelişiminin diğer aşamaları gözlenmiştir. Burada gösterilen örnek (Şekil 7) yakın zamanda bir tarafından istila edilmiş bir kırmızı kan hücresi içerir. P. falciparum parazit (merozoit). Kullanılan lamel 240 nm kalınlığındaydı ve ızgara, TEM'e yerleştirilmeden önce argon atmosferinde platin ile hafifçe püskürtülmüştü, bu da normalde beklenenden biraz daha az kontrastla sonuçlandı. Kırmızı kan hücresi zarı, hücreyi çevreleyen bütünüyle izlenebilir. Kırmızı kan hücresi içinde üç kapalı zara bağlı yapı, her biri çift zarla çevrili iki vezikül ve ampul şeklinde bir özellik (en geniş kısımlarında 1.2 μm x 0.9 μm) vardı, bu da bir merozoit (Şekil 7A-B). İki vezikülün içeriği, kırmızı kan hücresinin içeriğinin aksine benzer görünüyordu, bu da bu veziküllerin hemoglobin içerebileceğini düşündürüyordu. İnvazyon sonrası konakçı kırmızı kan hücresinde veziküllerin varlığı daha önce gözlenmiştir45. Merozoitteki rhoptries adı verilen salgı organellerinden lipitlerin ve diğer virülans faktörlerinin salgılanmasından kaynaklandığı düşünülmektedir ve bunlar istila sırasında konakçı kırmızı hücreye boşalmaktadır. Merozoy, en içte muhtemelen görünür bir yüzey kaplaması bulunmayan merozoitin doğal plazma zarı olan ve en dışta merozoiti istila ederken saran konakçı kırmızı hücre zarından türetilmesi gereken, yakından ilişkili iki membranla çevrilidir. Merozitin sitoplazması, birçok çok katmanlı vezikül ve tepedeki bir zar yığınına bitişik elektron yoğun mantar şeklinde bir özellik içerir. Bu bölge üzerinden elde edilen bir eğim serisi (2,4 ş/ piksel), mantar şeklindeki özelliğe (Şekil 7C-E). Çarpıcı bir şekilde, bu morfoloji, olgun bir merozoitin apikal ucundaki organellerin ve hücresel yapıların normal düzenlemesinden oldukça farklıdır. Bunu değerlendirmek için, olgun bir merozoit üzerinde apikal uç üzerinde karşılaştırmalı bir eğim serisi (2.74 ş/ piksel), dalma dondurma ve frezelemeden önce E64 ile muamele edilmiş bir şizonttan elde edilmiştir (Şekil 8). Bu, bir merozoitin apikal ucunun, hücrenin apikal ucundaki üç kutupsal halka kümesi içine yerleştirilmiş ve mikronem adı verilen bir dizi daha küçük salgı organeli ile çevrili rhoptries adı verilen iki belirgin kulüp şeklindeki salgı organeli içerdiğini göstermektedir. En içteki polar halka, istilaya neden olan motor proteinleri içeren iç membran kompleksi adı verilen merozoit plazma zarının altında yatan çift membranlı bir yapıya bağlanır. İnvazyon sırasında salgı organellerinin içeriğinin merozoit yüzeyine ve konakçı kırmızı kan hücresine boşaltıldığı, merozoitlerin konakçı kırmızı hücrelere bağlanmasını kolaylaştırdığı ve istilayı tetikleyen motor kompleksini başlattığı gösterilmiştir45. Buradaki veriler, istiladan sonra, merozoitin roptrilere, mikronemelere veya kutupsal halkalara benzeyen gözlemlenebilir yapılar içermediğini göstermektedir, bu da merozoitin apikal ucunun morfolojisinin çarpıcı bir şekilde değiştiğini düşündürmektedir. İstiladan önce rhoptrilerin füzyonu daha önce TEM tarafından sabit, oda sıcaklığında bölümlerin gösterilmesi46, istila sonrası durum merozoitimizde gözlemlediğimiz mantar şeklindeki özelliğin üretimi ile tutarlıdır. Bu özelliğe bağlı membran yığınları daha önce gözlemlenmemiştir ve yeni istila edilen merozoitte mevcut bir IMC belirtisi olmadığından, membran yığınlarının istila tamamlandıktan sonra kalan IMC makinesinin kalıntıları olabileceğini varsayıyoruz, ancak bu doğrulanmaya devam ediyor.

Şekil 8: FIB ile öğütülmüş bir lamelin kriyo-ET'si ve plastik kesitlerin TEM ile olgun merozoitlerin apikal ucu. (A) Olgun bir merozoitin apikal ucunun tipik morfolojisini gösteren 230 nm kalınlığındaki bir lamelden 8x bağlanmış bir tomogramdan (2.74 ş/ piksel) ortalama 20 merkezi dilim. Şizontlar, dalma donmasından önce E64 ile muamele edildi, böylece PV zarı yırtıldı ve merozoitler konakçı kırmızı hücre içinde bulundu. (B), (A) ile aynı şeyi gösterir, ancak konakçı kırmızı kan hücresi zarını (RCM, kırmızı), merozoit plazma zarını (yeşil), iç zar kompleksini (IMC, mor), polar halkaları (PR, siyah), mikronemleri (M, sarı), rhometrileri (R, gri) ve nükleer zarfı (NE, mavi) belirtmek için ek açıklamalarla gösterir. Tomogramın bu bölgesinde düzlem dışı olan komşu bir merozoit yeşil bir üçgenle gösterilir. Ölçek çubuğu, 200 nm. (C) KriyoET tarafından gözlenen hücresel özellikler, plastik bölümlerde korunan şizontların TEM'inden gelenlerle tutarlıdır. Benzer hücresel özellikler, PV membran rüptüründen önce çıkışı durduran, bileşik 2 ile muamele edilmiş bir şizonttan olgun bir merozoitte belirtilir. Ölçek çubuğu, 500 nm. (D) Parçalardaki hücresel özellikleri gösteren bir merozoitin açıklamalı şeması (A-C). Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}
| Dikdörtgen frezeleme desenleri arasındaki korunan bölgenin kalınlığı (μm) | 30 kV'da nomik iyon ışın akımı (pA) |
| 3 | 300 |
| 1.5 | 100 |
| 0.75 | 50 |
| 0.3 | 30 |
| 0.2 - 0.06 (son parlatma) | 30* |
Tablo 1: İnce lamel üretmek için frezeleme stratejisi. Kademeli frezeleme, korunan bölgenin kalınlığına karşılık gelen iyon ışını akımını azaltarak gerçekleştirilir. Bu, numune ısıtmasını sınırlayarak sapmayı önler. * Parlatma, lamellerin her iki tarafından eşit ısı uygulamak için paralel olarak yapılmalı, böylece son kalınlıklarına ulaştıklarında bükülme veya eğilme riski azaltılmalıdır. Diğer adımlar, bir sonraki ışın akımına geçmeden önce deseni lamelin üstünde ve daha sonra lamelin altında frezeleyerek sırayla yapılabilir. Frezeleme konumlarını bulmak için ızgaranın incelenmesi, numune ısıtmasını en aza indirmek için 1,5 pA ışın akımında yapılmalıdır. Bu Tabloyu indirmek için lütfen tıklayınız.
Tartışmalar
KriyoFIB frezeleme daha rutin hale gelirken, frezeleme için en uygun numunelerin hazırlanması; bu nedenle, bu protokoldeki kritik adımların çoğu, numune cryoFIB-SEM'e ulaşmadan önce gerçekleşir. Frezeleme denenmeden önce mümkün olan en iyi numuneyi üretmek için çoklu hücre hazırlama, daldırma dondurma ve ışık ve elektron mikroskobu ile tarama ile numune optimizasyonu gereklidir. Mümkün olan en iyi numuneye sahip olmak sadece başarı şansını arttırmakla kalmaz, aynı zamanda ekipmanın kullanımını da optimize eder. Bu nedenle, çoğu ulusal tesis, makinelerinde zaman vermeden önce yeterli optimizasyonun yapıldığına dair kanıt gerektirir. KriyoFIB-SEM'in içine girdikten sonra, organoplatin katın etkinliğini en üst düzeye çıkarmak, düzgün ince lameller üretmek için kritik öneme sahiptir. Son olarak, sabır ve iyi numune kullanımı, hassas lameller içeren ızgaraları üretmek ve daha sonra aktarmak için birkaç gün boyunca oturması gereken kullanıcının ön koşul becerisidir. Bu, otomatik frezeleme stratejilerinin47,48 tanıtılmasıyla değişebilir, ancak henüz, tüm frezeleme işlemi çoğu tesiste hala büyük ölçüde manueldir.
Çalışma sadece P. falciparum schizonts'a odaklanırken, burada sunulan yöntem, diğer hücre tipleri için ızgara hazırlığını ve frezelemeyi optimize etmek için kolayca değiştirilebilir. Dikkate alınması gereken önemli faktörler, ızgaraya uygulanan hücrelerin yoğunluğu, ızgara tipi (bakır bazı hücreler için toksiktir), karbon filmdeki deliklerin boyutu ve aralığı, lekelenme süresi ve lekelenme yöntemidir (tek / çift taraflı, manuel veya otomatik). Ek olarak, hücreler ızgaralarda yetiştiriliyorsa (genellikle delikli karbon filmli altın ızgaralar), lekelenmeden ve donmadan önce akıcılık ışık mikroskobu ile kontrol edilebilir. Frezeleme stratejisi, hücrelerin ızgarada nasıl biriktirildiğine bağlıdır. Sıtma bulaşmış kırmızı kan hücreleri için, ızgara esas olarak kırılmamış bir hücre tabakası ile kaplanır, bazı bölgelerde daha kalın ve diğer bölgelerde daha incedir. Frezeleme, buz kalınlığının tomografi için uygun boyutta lameller ürettiği bu gradyan boyunca bir bölgede birden fazla noktada gerçekleştirilir. Bu yaklaşım, birden fazla hücreden bir dilim içeren lameller üretebildiğiniz için daha küçük hücreler için iyi çalışır. Daha büyük hücreler veya hücre kümeleri için, bir hücrenin veya hücre kümesinin bir lamel üretebilmesi söz konusu olabilir.
Şebeke işleme ve numune aktarımı, bu iş akışının ana zorluklarından biri olmaya devam etmektedir. Lameller, kırılmalar veya çatlaklar, biyolojiyi gizleyen perdeleme artefaktları, aşırı yüzey kirliliği ve TEM içindeki ızgara oryantasyon problemleri nedeniyle kaybolabilir ve ızgarayı yeniden konumlandırmak için ekstra bir taşıma adımı gerektirir. Kayıpların derecesi günden güne ve şebekeden şebekeye değişir, ancak zaman içinde uygulama ve kullanım deneyimi ile geliştirilir. Bu çalışmada, ızgaraların lamelleri tahrip etmeden otomatik yükleyicide birçok kez dikkatli bir şekilde yeniden yönlendirilebileceği ve bunun bazen yüzey buzunu yıkama yararına sahip olduğu bulunmuştur. Bu iş akışının bir diğer ana sınırlaması, lamel üretmek için gereken süredir. Üretim yavaş olduğundan, frezelemeyi mümkün olduğunca verimli hale getiren uygun şekilde optimize edilmiş bir numuneye sahip olmak çok önemlidir.
Vitröz biyolojik numunelerin FIB frezelenmesine bir dizi adaptasyon yakın zamanda tanıtılmıştır. Bir oyun değiştirici, kriyoFIB-SEM haznesi içinde, yüksek basınçlı dondurulmuş numunelerden daha büyük malzeme bloklarının öğütülmesini sağlayan kriyojenik olarak soğutulmuş kaldırma takımlarının uygulanmasıdır. Bloklar metal bir çubuğa tutturulabilir veya bir tutucuda toplanabilir ve özel olarak modifiye edilmiş bir EM ızgarası içeren ikinci bir numune konumuna taşınabilir. Bloklar daha sonra organoplatin ile kaplanabilir ve lamel üretmek için öğütülebilir. Lamelleri yüksek basınçlı dondurulmuş malzemeden öğütme yeteneği, özellikle bağıntılı floresan mikroskobu12 ile alanları hedefleyen çok daha büyük hücrelerin ve dokuların işlenebileceği anlamına gelir. FIB-frezeleme yöntemine yapılan diğer yeni uyarlamalar arasında, numune16'nın önceden frezelenmesiyle perdeleme artefaktlarının azaltılması, mikroakışkan kriyo-fiksasyon49 ve hücre dağılımını iyileştirmek için elektron mikroskopi ızgaralarının foto-mikrodesenlenmesi50 bulunmaktadır. Ayrıca, bir lamelin her iki tarafındaki mikro genleşme boşluklarının frezelenmesinin, son inceliğine51 ulaştığında çevredeki numune tarafından sıkıştırmayı hafifletebileceği gösterilmiştir. Bu, özellikle son parlatma adımında lamellerin bükülmesinin bazen görüldüğü bu çalışmadaki örnek gibi sürekli hücre katmanlarını frezelerken yararlı olabilir.
Elektron mikroskobunun geleceği muhtemelen sub-tomogram ortalaması ile in situ moleküler yapıların belirlenmesi olacaktır ve FIB frezeleme, bu tür iş akışları için vitreus biyolojik numunelerinin üretimini kolaylaştıracak önemli bir araçtır. FIB-frezeleme biyolojik uygulamalar için henüz emekleme aşamasındayken, hem akademik hem de ulusal tesislerdeki araştırmacıların sıkı çalışmaları ve araştırmayı desteklemek için kriyoFIB-SEM teknolojisinin geliştirilmesine yönelik ticari yatırımlar sayesinde yöntem geliştirme hızlı bir şekilde gerçekleşmektedir.
Açıklamalar
Yazarların açıklayacak hiçbir şeyi yok.
Teşekkürler
Bu araştırma tamamen veya kısmen Wellcome Trust (212916 / Z / 18 / Z) tarafından finanse edilmiştir. Açık erişim amacıyla, yazar bu gönderimden kaynaklanan herhangi bir Yazar Kabul Edilen Makale versiyonuna CC BY kamu telif hakkı lisansı uygulamıştır.
Bu yöntemin geliştirildiği bu proje, Helen R. Saibil, Roland A. Fleck ve Michael J. Blackman'a verilen MR / P010288/1 Tıbbi Araştırma Konseyi hibesi ile finanse edildi. P. falciparum kültürleri, Michael J. Blackman'ın grubunun üyelerinin desteğiyle Francis Crick Enstitüsü'nde yetiştirildi. Yazarlar, ince kan yaymalarında bileşik 2 ve E64 ile tedavi edilen şizontların görüntülerini sağladığı için Dr. Ser Ying (Michele) Tan'a teşekkür eder. Lamellerin çoğu eBIC personelinin desteğiyle üretildi ve araştırma önerisi NT21004 Scios çift ışınlı kriyoFIB-SEM'e erişim için minnettarız. Yazarlar ayrıca, Royal Society Industry Fellowship planının (INF \ R2 \ 202061) CUI içindeki FIB frezeleme tekniğinin geliştirilmesine devam etmedeki desteğini de kabul etmektedir. Yazarlar ayrıca Helen R. Saibil'e bu yöntem makalesi ile ilgili yararlı tartışmalar ve projenin denetlenmesi için teşekkür eder.
Malzemeler
| Name | Company | Catalog Number | Comments |
| c-clips | Thermo Fisher Scientific | 1036171 | |
| Clipping station | Thermo Fisher Scientific | n/a | Direct quote from Thermo |
| Clipping station | Sub-angstrom | CSA-01 | |
| Compound 2 | n/a | n/a | Synthesised by Dr Simon A. Osborne, LifeArc |
| Cryo-FIB specific autogrid rims | Thermo Fisher Scientific | 1205101 | |
| E64 | Sigma | E3132 | |
| Emitech K100X glow discharge unit | Quorum | n/a | |
| Gibco RPMI 1640 media | Thermo Fisher Scientific | 12633012 | Formulation used for culturing is custom made (REF 041-91762 A) and includes Albumax, glutamine, HEPES and hypoxanthine supplements |
| Giemsa Stain | VWR International | 350864X | |
| Glass slides | Thermo Fisher Scientific | 11562203 | |
| Grid boxes | Sub-angstrom | PB | For clipped grids |
| Grid boxes | Thermo Fisher Scientific | n/a | For clipped grids - direct quote from Thermo Fisher Scientific |
| Grid boxes | Agar Scientific | AGG3727 | |
| Home-made manual plunge freezing rig | n/a | n/a | With an insulated ethane pot (high-density foam) and liquid nitrogen bath (polystyrene) on a bench top in a containment facility. |
| Human blood | n/a | n/a | UK National Blood and Transplant service |
| JEOL 4700F Z JSM with a Leica VCT500 stage cooling system | JEOL | n/a | FIB-SEM |
| Leica EM ACE600 with a VCT500 cryostage | Leica | n/a | sputter coater |
| Leica EM ACE900 | Leica | n/a | e-beam rotary coater. In a humidity controlled room. |
| Linkam cryo-stage for light microscope | Linkam | Model No. CMS196 | Cassettes for clipped and un-clipped grids. In a humidity controlled room. |
| Methanol | Sigma | 179957 | |
| Nikon Eclipse E200 light microscope | Nikon | n/a | with Linkam cryo-stage in a humidity controlled room. |
| Percoll | VWR International | 17-0891-01 | Solution for percoll cushion is 35 ml 10x PBS, 150 ml RPMI 1640 media and 315 ml Percoll |
| Quantifoil copper 200 mesh 2/4 holey carbon EM Finder grids | Quantifoil | N1-C17nCuH2-01 | 100 pack |
| Quantifoil copper 200 mesh 2/4 holey carbon EM grids | Quantifoil | N1-C17nCu20-01 | 100 pack |
| Quorum PP3010 prep-chamber | Quorum | n/a | sputter coater |
| Scios dual beam equipped with a Quorum PP3010 transfer stage. | FEI | n/a | FIB-SEM |
| Staedtler Lumocolor fine black permanent marker pen | Viking Direct | ND538522 | |
| TFS Titan Krios 300 kV TEM | Thermo Fisher Scientific | n/a | TEM equipped with a K2 or K3 camera. |
| Whatmann grade 1 filter paper | Sigma | WHA1001150 | |
| Zeiss Axio Scope A1 light microscope | Zeiss | n/a | with Linkam cryo-stage in a humidity controlled room. |
Referanslar
- Baumeister, W. From proteomic inventory to architecture. FEBS Letters. 579 (4), 933-937 (2005).
- Rigort, A., Villa, E., Bäuerlein, F. J. B., Engel, B. D., Plitzko, J. M. Integrative approaches for cellular cryo-electron tomography: Correlative imaging and focused ion beam micromachining. Methods in Cell Biology. 111, 259-281 (2012).
- Oikonomou, C. M., Chang, Y. W., Jensen, G. J. A new view into prokaryotic cell biology from electron cryotomography. Nature Reviews Microbiology. 14, 205-220 (2016).
- Wagner, F. R., et al. Preparing samples from whole cells using focused-ion-beam milling for cryo-electron tomography. Nature Protocols. 15 (6), 2041-2070 (2020).
- Giannuzzi, L. A., Stevie, F. A. A review of focused ion beam milling techniques for TEM specimen preparation. Micron. 30 (3), 197-204 (1999).
- Narayan, K., Subramaniam, S. Focused ion beams in biology. Nature Methods. 12 (11), 1021-1031 (2015).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Acadamy of Science of the United States of America. 109 (12), 4449-4454 (2012).
- Marko, M., Hsieh, C., Schalek, R., Frank, J., Mannella, C. Focused-ion-beam thinning of frozen-hydrated biological specimens for cryo-electron microscopy. Nature Methods. 4 (3), 215-217 (2007).
- Villa, E., Schaffer, M., Plitzko, J. M., Baumeister, W. Opening windows into the cell: Focused-ion-beam milling for cryo-electron tomography. Current Opinion in Structural Biology. 23 (5), 771-777 (2013).
- Wang, K., Strunk, K., Zhao, G., Gray, J. L., Zhang, P. 3D structure determination of native mammalian cells using cryo-FIB and cryo-electron tomography. Journal of Structural Biology. 180 (2), 318-326 (2015).
- de Winter, D. A. M., et al. In-situ integrity control of frozen-hydrated, vitreous lamellas prepared by the cryo-focused ion beam-scanning electron microscope. Journal of Structural Biology. 183 (1), 11-18 (2013).
- Schaffer, M., et al. A cryo-FIB lift-out technique enables molecular-resolution cryo-ET within native Caenorhabditis elegans tissue. Nature Methods. 16 (8), 757-762 (2019).
- Mahamid, J., et al. Visualizing the molecular sociology at the HeLa cell nuclear periphery. Science. 351 (6276), 969-972 (2016).
- Albert, S., et al. Proteasomes tether to two distinct sites at the nuclear pore complex. Proceedings of the National Acadamy of Science of the United States of America. 114 (52), 13726-13731 (2017).
- Guo, Q., et al. In situ structure of neuronal C9orf72 poly-GA aggregates reveals proteasome recruitment. Cell. 172 (4), 696-705 (2018).
- Schaffer, M., et al. Optimized cryo-focused ion beam sample preparation aimed at in situ structural studies of membrane proteins. Journal of Structural Biology. 197 (2), 73-82 (2017).
- Szwedziak, P., Wang, Q., Bharat, T. A. M., Tsim, M., Löwe, J. Architecture of the ring formed by the tubulin homologue FtsZ in bacterial cell division. eLife. 3, 04601 (2014).
- Carlson, L. A., et al. Cryo electron tomography of native HIV-1 budding sites. PLOS Pathogens. 6 (11), 1001173 (2010).
- Klein, S., et al. SARS-CoV-2 structure and replication characterized by in situ cryo-electron tomography. Nature Communications. 11 (1), 5885 (2020).
- Studer, D., Humbel, B. M., Chiquet, M. Electron microscopy of high pressure frozen samples: Bridging the gap between cellular ultrastructure and atomic resolution. Histochemistry and Cell Biology. 130 (5), 877-889 (2008).
- Schertel, A., et al. Cryo FIB-SEM: Volume imaging of cellular ultrastructure in native frozen specimens. Journal of Structural Biology. 184 (2), 355-360 (2013).
- Murphy, G. E., et al. Correlative 3D imaging of whole mammalian cells with light and electron microscopy. Journal of Structural Biology. 176 (3), 268-278 (2012).
- Heymann, J. A. W., et al. Site-specific 3D imaging of cells and tissues with a dual beam microscope. Journal of Structural Biology. 155 (1), 63-73 (2012).
- Spehner, D., et al. Cryo-FIB-SEM as a promising tool for localizing proteins in 3D. Journal of Structural Biology. 211 (1), 107528 (2020).
- Kamino, T., Yaguchi, T., Ohnishi, T., Ishitani, T., Osumi, M. Application of a FIB-STEM system for 3D observation of a resin-embedded yeast cell. Journal of Electron Microscopy. 53 (5), 563-566 (2004).
- Han, H., Zuber, B., Dubochet, J. Compression and crevasses in vitreous sections under different cutting conditions. Journal of Microscopy. 230, 167-171 (2008).
- Matias, V. R. F., Al-amoudi, A., Dubochet, J., Beveridge, T. J. Cryo-transmission electron microscopy of frozen-hydrated sections of Escherichia coli and Pseudomonas aeruginosa. Journal of Bacteriology. 185 (20), 6112-6118 (2003).
- Al-Amoudi, A., Studer, D., Dubochet, J. Cutting artefacts and cutting process in vitreous sections for cryo-electron microscopy. Journal of Structural Biology. 150 (1), 109-121 (2005).
- Al-Amoudi, A., et al. Cryo-electron microscopy of vitreous sections. The EMBO Journal. 23 (18), 3583-3588 (2004).
- Bouchet-Marquis, C., Dubochet, J., Fakan, S. Cryoelectron microscopy of vitrified sections: A new challenge for the analysis of functional nuclear architecture. Histochemistry and Cell Biology. 125 (1-2), 43-51 (2006).
- Hayles, M. F., et al. The making of frozen-hydrated, vitreous lamellas from cells for cryo-electron microscopy. Journal of Structural Biology. 172 (2), 180-190 (2010).
- Dubochet, J., Adrian, M., Chang, J. -. J., Lepault, J., McDowall, A. W. Cryoelectron microscopy of vitrified specimens. Cryotechniques in Biological Electron Microscopy. , 114-131 (1987).
- Hsieh, C., Schmelzer, T., Kishchenko, G., Wagenknecht, T., Marko, M. Practical workflow for cryo focused-ion-beam milling of tissues and cells for cryo-TEM tomography. Journal of Structural Biology. 185 (1), 32-41 (2014).
- Harapin, J., et al. Structural analysis of multicellular organisms with cryo-electron tomography. Nature Methods. 12 (7), 634-636 (2015).
- Rubino, S., et al. A site-specific focused-ion-beam lift-out method for cryo Transmission Electron Microscopy. Journal of Structural Biology. 180 (3), 572-576 (2012).
- Duyvesteyn, H. M. E., et al. Machining protein microcrystals for structure determination by electron diffraction. Proceedings of the National Acadamy of Science of the United States of America. 115 (38), 9569-9573 (2018).
- Blackman, M. J. Purification of Plasmodium falciparum merozoites for analysis of the processing of merozoite surface protein-1. Microbes as Tools for Cell Biology. 45, 213-220 (1995).
- Hagen, W. J. H., Wan, W., Briggs, J. A. G. Implementation of a cryo-electron tomography tilt-scheme optimized for high resolution subtomogram averaging. Journal of Structural Biology. 197 (2), 191-198 (2017).
- Zheng, S. Q., et al. MotionCor2 - anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nature Methods. 14 (4), 331-332 (2017).
- Grant, T., Grigorieff, N. Measuring the optimal exposure for single particle cryo-EM using a 2.6 Å reconstruction of rotavirus VP6. eLife. 4, 06980 (2015).
- Rohou, A., Grigorieff, N. CTFFIND4: Fast and accurate defocus estimation from electron micrographs. Journal of Structural Biology. 192 (2), 216-221 (2015).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Turoňová, B., Schur, F. K. M., Wan, W., Briggs, J. A. G. Efficient 3D-CTF correction for cryo-electron tomography using NovaCTF improves subtomogram averaging resolution to 3.4 Å. Journal of Structural Biology. 199 (3), 187-195 (2017).
- Arnold, J., et al. Site-specific cryo-focused ion beam sample preparation guided by 3d correlative microscopy. Biophysical Journal. 110 (4), 860-869 (2016).
- Riglar, D. T., et al. Super-resolution dissection of coordinated events during malaria parasite invasion of the human erythrocyte. Cell Host & Microbe. 9 (1), 9-20 (2011).
- Hanssen, E., et al. Electron tomography of Plasmodium falciparum merozoites reveals core cellular events that underpin erythrocyte invasion. Cellular Microbiology. 15 (9), 1457-1472 (2013).
- Buckley, G., et al. Automated cryo-lamella preparation for high-throughput in-situ structural biology. Journal of Structural Biology. 210 (2), 107488 (2020).
- Zachs, T., et al. Fully automated, sequential focused ion beam milling for cryo-electron tomography. eLife. 9, 52286 (2020).
- Fuest, M., et al. In situ microfluidic cryofixation for cryo focused ion beam milling and cryo electron tomography. Scientific Reports. 9 (1), 19133 (2019).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Wolff, G., et al. Mind the gap: Micro-expansion joints drastically decrease the bending of FIB-milled cryo-lamellae. Journal of Structural Biology. 208 (3), 107389 (2019).
Erratum
Formal Correction: Erratum: Preparing Lamellae from Vitreous Biological Samples using a Dual-Beam Scanning Electron Microscope for Cryo-Electron Tomography
Posted by JoVE Editors on 9/11/2023. Citeable Link.
An erratum was issued for: Preparing Lamellae from Vitreous Biological Samples using a Dual-Beam Scanning Electron Microscope for Cryo-Electron Tomography. The Authors section was updated from:
Claudine Bisson1,2
Corey W. Hecksel3,4
James B. Gilchrist3
Roland A. Fleck1
1Centre for Ultrastructural Imaging, New Hunt’s House, Guy’s Campus, King’s College London
2Department of Biological Science, Birkbeck College, University of London
3Electron Bio-Imaging Centre, Diamond Light Source, Harwell Science and Innovation Campus
4SLAC National Accelerator Laboratory, Stanford University
to:
Claudine Bisson1,2
Corey W. Hecksel3,4
James B. Gilchrist3
M. Alejandra Carbajal1
Roland A. Fleck1
1Centre for Ultrastructural Imaging, New Hunt’s House, Guy’s Campus, King’s College London
2Department of Biological Science, Birkbeck College, University of London
3Electron Bio-Imaging Centre, Diamond Light Source, Harwell Science and Innovation Campus
4SLAC National Accelerator Laboratory, Stanford University
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır