
Method Article
Protocolo para el modelado de blastoides humanos Desarrollo e implantación de blastocistos
* Estos autores han contribuido por igual
En este artículo
Resumen
Un protocolo que describe la formación de blastoides humanos que generan de manera eficiente, oportuna y secuencial células similares a los blastocistos.
Resumen
Un modelo del blastocisto humano formado a partir de células madre (blastoide) apoyaría los avances científicos y médicos. Sin embargo, su poder predictivo dependerá de su capacidad para recapitular de manera eficiente, oportuna y fiel las secuencias de desarrollo de blastocistos (morfogénesis, especificación, patrones), y para formar células que reflejen la etapa de blastocisto. Aquí mostramos que las células madre pluripotentes humanas ingenuas cultivadas en condiciones PXGL y luego triplemente inhibidas para el hipopótamo, transformando el factor de crecimiento β y las vías de quinasa reguladas por señales extracelulares se someten eficientemente a la morfogénesis para formar blastoides (>70%). Coincidiendo con el tiempo de desarrollo (~ 4 días), los blastoides desenrollan la secuencia de especificación del blastocisto mediante la producción de análogos del trofoblasto y el epiblasto, seguidos de la formación de análogos del endodermo primitivo y los trofoblastos polares. Esto resulta en la formación de células transcripcionalmente similares al blastocisto (>96%) y una minoría de análogos post-implantación. Los blastoides modelan eficientemente formando el eje embrionario-abembrionario marcado por la maduración de la región polar (NR2F2+), que adquiere el potencial específico para unirse direccionalmente a las células endometriales estimuladas hormonalmente, como en el útero. Tal blastoide humano es un modelo escalable, versátil y ético para estudiar el desarrollo humano y la implantación in vitro.
Introducción
La falta de modelos experimentales ha limitado la comprensión de la embriogénesis humana temprana. El conocimiento actual de los aspectos humanos específicos del desarrollo embrionario se deriva de los embriones excedentes de fertilización in vitro (FIV) donados para la investigación. Sin embargo, la limitada disponibilidad, las dificultades de las manipulaciones experimentales y la calidad variable de los embriones dificultan las investigaciones científicas. Por el contrario, un modelo in vitro fiel de embriones humanos permitiría manipulaciones experimentales complejas, proporcionando así una oportunidad ética para complementar la investigación en embriones humanos 1,2,3,4. Un modelo previamente desarrollado de blastocistos de ratón combinó células madre embrionarias de ratón y células madre trofoblásticas5. En este protocolo detallado, se describe un método para generar un modelo del blastocisto humano a partir de células madre pluripotentes naïve que es fiel a los criterios de blastocisto elemental6.
Cuatro criterios para los blastoides humanos. Aquí, en un intento de establecer una definición estandarizada de blastoides humanos, proponemos cuatro criterios mínimos. Aunque no son exhaustivos, estos criterios podrían servir de base para evaluar los parámetros que permiten la formación de blastoides humanos (Figura 1A). (1) Los blastoides deben formarse eficientemente en términos de morfología y de generación de los análogos de los tres linajes, a saber, epiblasto (Epi), trofoectodermo (TE) y endodermo primitivo (PrE). Es probable que la ineficiencia apunte a un estado celular inicial inadecuado o/o a una condición de cultivo (por ejemplo, medio blastoideo). (2) Los blastoides deben generar análogos de los tres linajes de acuerdo con la secuencia de desarrollo (Epi/TE primero, PrE/polarTE último)7,8 y el tiempo (inducción ~ 3 días; días embrionarios 5-7)7,9. (3) Los blastoides deben formar análogos de la etapa de blastocisto, pero no de las etapas posteriores a la implantación (por ejemplo, epiblasto postimplantación, trofoblasto o células de amnios). (4) Finalmente, los blastoides deben ser capaces de recapitular las características funcionales de la implantación y el desarrollo de blastocistos. Usando este protocolo, los blastoides humanos se forman de manera eficiente utilizando múltiples líneas celulares (>70%), son capaces de generar los análogos celulares de blastocistos secuencialmente y dentro de los 4 días, y los análogos son transcripcionalmente similares a la etapa de blastocisto (>96% basado en varios análisis)6,10,11. Finalmente, los blastoides generan robustamente el eje embrio-abembrionario, lo que les permite interactuar con las células endometriales estimuladas hormonalmente a través de la región polar, y expandir robustamente los linajes en el cultivo extendido (equivalente en el tiempo: día embrionario 13).
La importancia del estado inicial de la célula. Las células madre pluripotentes humanas (hPSC) se pueden estabilizar en diferentes estados que intentan capturar etapas precisas del desarrollo. Estos estados son sostenidos por condiciones de cultivo que, aunque todavía subóptimas, restringen las células en una etapa 12 de epiblasto preimplantacional (~ embrionario días 5-7) o postimplantacional (~ días embrionarios8-14). El análisis transcriptómico mostró que las hPSC cultivadas en PD0325901, XAV939, Gö6983 y el factor inhibidor de la leucemia (LIF; denominadas hPSC naïve PXGL)13,14 son más similares al epiblasto blastocisto en comparación con las hPSC cultivadas en factor de crecimiento de fibroblastos (FGF) 2 y activina15 (denominadas hPSCs12 cebadas) y a células madre pluripotentes extendidas humanas (hEPSC)16 (ver análisis en las referencias17, 18,19). En consecuencia, el transcriptoma de las hPSC cebadas coincide mejor con un epiblasto de mono cynomolgus post-implantación/pre-gastrulación20. Criterios moleculares adicionales, como la expresión del transposón, la metilación del ADN y el estado del cromosoma X, confirmaron que las variaciones del estado ingenuo se asemejan más al epiblasto blastocisto en comparación con el estado cebado17,21. Finalmente, las líneas de hPSC naïve se han derivado con éxito directamente de blastocistos utilizando condiciones de cultivo PXGL22.
Las células de blastocisto temprano en humanos aún no están comprometidas. La especificación del linaje murino se produce a partir de la etapa de la mórula que precede a la etapa de blastocisto23. Por el contrario, los experimentos de disociación y reagregación han demostrado que las células del trofoectodermo humano de los blastocistos tempranos aún no están comprometidas24. En consecuencia, el análisis de las células de blastocistos humanos mediante secuenciación de ARN unicelular (scRNAseq) ha demostrado que la primera especificación de linaje (trofoblasto/epiblasto) se produce después de la formación de la cavidad del blastocisto7. Esta especificación humana diferida se correlaciona con las observaciones de que las hPSC son potentes para formar trofoblastos 25,26,27 cuando las PSC de ratón están en gran medida comprometidas con el linaje de epiblastos. Estas observaciones combinadas llevaron a la posibilidad de que las hPSC ingenuas reflejen una etapa de blastocisto y conserven el potencial de formar los tres linajes de blastocistos. Últimamente, se ha propuesto que la potencia de las hPSC para especificar análogos extraembrionarios cambia de trophectoderm a amnion durante la progresión de estado ingenuo a estado cebado27. Por lo tanto, las hPSC naïve son más similares a la etapa preimplantacional 17,18,21 y tienen una mayor capacidad para formar trofoblastos en comparación con las hPSCs27, hEPSC 16 o estados reprogramados intermedios28, que son propensos a formar análogos postimplantación10 (Figura 1B ). Por lo tanto, el estado celular inicial es crucial para formar los análogos extraembrionarios apropiados. Aunque aún queda por hacer un análisis exhaustivo lado a lado de los análogos del trofoectodermo convertidos, un estado ingenuo de PXGL que refleje el blastocisto temprano parece importante para formar blastoides de alta fidelidad.
Provoque la especificación y la morfogénesis mediante la inhibición de las vías de señalización. La inhibición de la vía de señalización de Hippo es un mecanismo conservado que impulsa la especificación de trofoblastos en ratones, vacas y humanos 9,29,30. Asimismo, desde 2013, se sabe que la inhibición del NODAL (A83-01) y la quinasa regulada por señal extracelular (ERK; PD0325901 o equivalente) y la activación de las vías de señalización de la proteína morfogenética ósea (BMP) desencadenan hPSCs preparadas para activar la red transcripcional asociada con el linaje trofoblasto 25,31,32,33,34. Además, recientemente varios informes también confirmaron que la inhibición de la vía NODAL y ERK y la activación de BMP facilitan la diferenciación trofoblástica de las hPSC naïve 25,31,32,33,34. Finalmente, si la especificación del trofoblasto se desencadena a partir de un estado ingenuo, las células recapitulan aspectos de la progresión del desarrollo del trofoectodermo26. Sin embargo, las líneas autorrenovadoras que reflejan el trofoectodermo del blastocisto no se han estabilizado in vitro. Siguiendo la especificación del trofoblasto, la inducción del factor de crecimiento epidérmico (EGF) y las vías de señalización Wnt junto con la inhibición de HDAC podría facilitar la progresión del desarrollo del trofoblasto 34,35 y estabilizar las células en líneas de células madre trofoblásticas humanas (hTSC) que reflejan citotrofofoblastos post-implantación 18,35. Tales líneas pueden derivarse tanto de blastocistos como de tejidos placentarios35.
El segundo linaje extraembrionario, denominado PrE, se especifica después de los trofoblastos y se origina en el epiblasto 7,9. Contrariamente a la PrE36 murina, se cree que la contraparte humana es independiente de la señalización FGF37,38. Las líneas que reflejan el endodermo extraembrionario (denominado nEnd) se establecieron a partir de hPSC naïve mediante la inducción de vías de señalización utilizando activina A, Wnt y LIF39. Inconsistente con los experimentos de inhibición embrionaria, se ha demostrado que la inhibición de ERK previene la formación de tales células nEND in vitro39. Hasta ahora, tales líneas no se han derivado directamente de blastocistos.
Últimamente, se han formado modelos del embrión temprano combinando variaciones de los medios previamente desarrollados para las células hTSCs35 y nEND39, utilizando así activadores del factor de crecimiento transformador β (TGF-β), EGF y las vías de señalización Wnt28,40. Estos modelos embrionarios se forman a baja eficiencia (10% -20%) y forman células que se asemejan a la etapa post-en lugar de pre-implantación10, incluyendo análogos del epiblasto post-implantación, trofoblasto, amnios, gastrula, tejidos mesodérmicos (~ embrionario día 14) y citotrofoblastos10. Por el contrario, una triple inhibición de las vías Hippo, ERK y TGF-β guía eficientemente la formación de blastoides que comprenden células similares a los blastocistos41. Junto con el estado celular inicial, proponemos que la inhibición de vías triples (Hippo, ERK, TGF-β) es el segundo parámetro esencial para formar blastoides de alta fidelidad (Figura 1B).
Evaluación del estado celular y la etapa reflejada utilizando scRNAseq. Los estados de las células que componen blastoides se pueden evaluar a través del análisis scRNAseq. Su similitud transcripcional con etapas embrionarias específicas se puede medir utilizando células blastoides solas y en comparación con hPSCs o hTSCs cebadas que reflejan las etapas post-implantación20,35. La realización de análisis de conglomerados utilizando diferentes niveles de definición revela cómo las subpoblaciones se fusionan progresivamente cuando la definición disminuye, revelando así las similitudes de los grupos. Aunque la optimalidad en el número de clusters se puede medir42, el clustering de alta resolución también informa sobre la eventual presencia de pequeñas subpoblaciones anormales, por ejemplo reflejando las etapas post-implantación10. Los genes expresados diferencialmente entre grupos pueden proporcionar información sobre sus análogos en el proceso de desarrollo mediante la evaluación de los niveles de expresión de los conjuntos de genes de referencia que definen los linajes específicos de la etapa. Esto permite medir el enriquecimiento de las subpoblaciones blastoides ya sea a través de mapas de distancia no supervisados (por ejemplo, utilizando genes enriquecidos superiores) o mediante análisis de enriquecimiento de conjuntos de genes (GSEA)43. Usando este protocolo blastoide, solo se forman tres grupos principales que reflejan transcripcionalmente los tres linajes de blastocistos. Un grupo incluye tanto las hPSC iniciales ingenuas como el análogo de epiblastos de los blastoides. El análisis de células en diferentes puntos de tiempo mostró la naturaleza secuencial de la especificación de linajes (los trofoblastos comienzan a especificar dentro de las 24 h y las células endodermos primitivas dentro de las 60 h). Una agrupación de alta resolución capturó una subpoblación de células (3,2%) que expresan genes específicos de embriones en etapa de gastrulación (posiblemente mesodermo o amnios). Cabe destacar que las hPSC iniciales ingenuas también comprendieron el 5% de las células similares a la postimplantación, como se describió anteriormente44. En un segundo análisis, las células blastoides se pueden fusionar in silico con células de referencia aisladas de concepti en diferentes etapas 45,46,47 para inferir la equivalencia de etapas. Aquí, se utilizaron como puntos de referencia células aisladas de concepti preimplantación 45,46, blastocistos cultivados in vitro 45 y embriones en estadio de gastrulación47. Usando este protocolo, se cuantificó que las células blastoides no coincidentes reveladas por agrupamiento de alta resolución de hecho se agrupan con mesodermo y amnio posterior a la implantación. En pasos futuros, la evaluación comparativa del transcriptoma debe complementarse con el análisis de la expresión del transposón, la metilación del ADN y el estado del cromosoma X, que también proporcionan puntos de referencia de las etapas del desarrollo21.
Evaluación de la formación de ejes y otras funcionalidades de blastoides humanos. Un blastocisto maduro se caracteriza por la formación del eje embrionario-abembrionario que modela trofoblastos para la implantación. Utilizando este protocolo blastoideo, se forma un eje robusto ejemplificado por una maduración de los trofoblastos proximales (por ejemplo, NR2F2+/CDX2-) que adquieren la capacidad de adherirse a las células organoides endometriales solo cuando son estimuladas hormonalmente48,49. La comparación con las trofoesferas que no forman el epiblasto muestra que estas células internas inducen a los trofoblastos colindantes a madurar para mediar la unión inicial al endometrio. Cuando se cultivan en un medio de cultivo extendido diseñado para blastocistos de mono cynomolgus50, los tres linajes del blastoide se expanden constantemente durante seis días adicionales (equivalente en el tiempo del día 13), aunque su organización no refleja esa etapa de desarrollo.
La implicación de los blastoides humanos de alta eficiencia y alta fidelidad. La conservación de los principios de desarrollo que se descubrieron en los organismos modelo es inherentemente difícil de probar en el conceptus humano debido al acceso restringido y a las dificultades técnicas para manipularlo genética y físicamente. Un modelo blastoide de alta eficiencia y alta fidelidad permitiría cribas genéticas y farmacológicas de alto rendimiento, que están en la base de los descubrimientos científicos y biomédicos. Además, la incorporación de modificaciones genéticas complejas para alterar y registrar los procesos biológicos complementaría dichos estudios. En general, proponemos que la triple inhibición (Hippo, TGF-β, ERK) de las hPSC PXGL ingenuas es conductora para la formación eficiente de blastoides humanos de alta fidelidad que cumplen con los cuatro criterios mínimos. La naturaleza escalable y versátil de este protocolo lo hace adecuado para generar hipótesis específicas que luego pueden validarse utilizando blastocistos humanos. Como tal, los blastoides humanos no reemplazarán el uso de conceptus humanos para la investigación in vitro , pero podrían actuar como una forma poderosa de canalizar la investigación a través de enfoques experimentales previamente inaccesibles en el corazón del proceso de descubrimiento científico y biomédico. El protocolo muestra cómo formar blastoides humanos y también cómo analizar las células que están contenidas dentro del blastoide.
Protocolo
Las Directrices para la Investigación con Células Madre y la Traducción Clínica de la Sociedad Internacional para la Investigación con Células Madre (ISSCR) recomiendan que la investigación sobre blastoides humanos sea permisible solo después de la revisión y aprobación a través de un proceso de revisión científica y ética especializada 3,4. Todos los procedimientos experimentales se llevaron a cabo siguiendo las directrices del comité de ética de investigación humana del Instituto de Biotecnología Molecular de la Academia Austriaca de Ciencias (IMBA) bajo la aprobación Rivron_Stellungnahme_2020-04-22. El cumplimiento de estas directrices es necesario para publicar los resultados de la investigación en revistas científicas.
1. Cultivo de células madre embrionarias humanas ingenuas en condición PXGL
NOTA: Las hPSC ingenuas se pueden obtener de los laboratorios pertinentes. Las líneas utilizadas aquí se obtuvieron de los laboratorios de Yasuhiro Takashima (actualmente en CiRA, Kyoto, Japón) y de Austin Smith (actualmente en Living Systems Institute, Exeter, Reino Unido). Alternativamente, las hPSC ingenuas se pueden restablecer internamente desde líneas de hPSC cebadas como se describió anteriormente13,14. Las hPSC ingenuas parecen estables para múltiples pasajes (> 15), pero la calidad de la cultura puede variar con el tiempo. Si la calidad de las hPSC ingenuas disminuye, descongele un nuevo vial de células o genere hPSC ingenuas de novo a partir de PSC cebadas. Para todas las composiciones de medios, véase el cuadro complementario 1.
- Preparación de la capa de alimentación embrionaria de ratón irradiada (MEFs)
- El día antes de la transmisión de hPSC ingenuas, prepare una placa de cultivo celular de 6 pocillos con capas de MEF irradiadas siguiendo los pasos que se describen a continuación.
- Cubra una placa de cultivo celular de 6 pocillos con 1 ml de gelatina al 0,1% en PBS por pocillo. Incubar la placa a 37 °C durante 30 min. Retire la solución de gelatina.
- Preparar el medio MEF a 37 °C.
- Descongele los MEF en un baño de agua a 37 °C hasta que solo quede un pequeño grupo de hielo. Disolver el volumen del vial con 1 ml de medio MEF preparado utilizando una pipeta P1000.
- Transfiera la suspensión celular a un tubo de 15 ml. Gire hacia abajo la suspensión a 200 x g durante 4 min. Aspire el sobrenadante y resuspenda el pellet MEF agregando medio MEF fresco (suficiente para 1.5 mL por pocillo).
- Cuente las células usando diapositivas de conteo de células y agregue 300,000 células por pozo y transfiera la placa a una incubadora normoxy a 37 ° C.
NOTA: Si los MEF se separan con el tiempo, se pueden agregar MEFs nuevos al medio PXGL durante el cambio de medios de rutina.
- Paso a través de células madre pluripotentes naïve humanas
- Antes de pasar las hPSC, verifique su morfología bajo el microscopio. Las colonias suelen tener una morfología en forma de cúpula con bordes brillantes y definidos. Si las colonias individuales muestran una morfología más plana o si comienzan a surgir colonias diferenciadas, siga las instrucciones del paso 1.2.8.
- Aspire el medio y lave las células con PBS una vez. Agregue 500 μL de solución de desprendimiento celular por pocillo de una placa de 6 pocillos.
- Incubar las células durante 5 min a 37 °C. Use una pipeta P1000 y pipetee las células varias veces para disociar las colonias en celdas individuales.
- Recoja las células y transfiéralas a un tubo de 15 ml que contenga tampón de lavado (1 ml por pocillo de una placa de 6 pocillos). Girar hacia abajo las células a 200 x g durante 4 min.
- Aspire el sobrenadante y resuspenda el pellet en medio PXGL fresco (suficiente para 1,5 ml por pocillo). Considere una proporción de división de 1: 3-1: 6 para el pasaje rutinario.
NOTA: Después de cada 3-4 pasajes o si la calidad del cultivo celular disminuye en función de la morfología celular (por ejemplo, la aparición de colonias planas en la población), agregue 10 μM Y-27632 y el extracto de membrana basal del factor de crecimiento (5 μL / pozo) al medio durante las primeras 24 h después del pasaje. - Antes de volver a colocar las hPSC, prepare las placas con MEFs frescos aspirando el medio MEF y lavando las células una vez con PBS. Luego, use una pipeta P1000 para transferir 1.5 ml de suspensión celular de hPSCs por pocillo de una placa de 6 pocillos que contiene los MEF.
NOTA: Asegúrese de que el pipeteo conduzca a una siembra homogénea de las células en toda el área del pozo. Esto asegurará el crecimiento de colonias con tamaños homogéneos y sincronía relativa de las células. - Cultivo de hPSCs en condiciones hipóxicas a 37 °C en ambiente humidificado. Después de 24 h, se deben conectar hPSC. Un alto número de células no adherentes (o flotantes) reflejan un problema de viabilidad o de unión al pasar.
- Cambie el medio con 1.5 ml de medio PXGL por pozo diariamente. Pasa hPSCs cada 3-4 días o úsalos para experimentos de formación de blastoides.
NOTA: Después de descongelar las hPSC, páselas por un mínimo de tres pasajes antes de comenzar un experimento de blastoides.
2. Formación de blastoides
- Formación de agregados PSC ingenuos
- Prepare y precaliente el medio PXGL, el medio basal N2B27, el tampón de lavado, el PBS y los medios de agregación antes de comenzar el experimento. Excluya los MEF de la suspensión de hPSCs para formar blastoides siguiendo los pasos que se describen a continuación.
- Para la exclusión de MEF, prepare una placa recubierta de gelatina agregando 1 ml de gelatina al 0,1% en el pozo de una placa de 6 pocillos e incubando a 37 ° C durante 30-90 min.
- Para cosechar las células, aspire el medio y lave las células con 1 ml de PBS.
- Añadir 500 μL de solución de desprendimiento celular (por pocillo de una placa de 6 pocillos) e incubar a 37 °C durante 5 min.
- Revise las células bajo el microscopio para seguir la disociación de las colonias en células individuales (algunos grupos multicelulares pueden disociarse más tarde mediante un pipeteo suave).
- Diluya la solución de desprendimiento celular con 1 ml de tampón de lavado. Recoja las células de la placa pipeteando suavemente de 5 a 10 veces. Transfiera la suspensión celular a un tubo de 15 ml. Girar hacia abajo las células a 200 x g durante 4 min.
- Aspirar el sobrenadante, resuspendir las células en 1,5 ml de medio PXGL (por pocillo de una placa de 6 pocillos) y sembrar las células en las placas recubiertas de gelatina para la exclusión de MEF e incubar a 37 ° C durante 60-90 min.
- Una vez que las células naïve se siembran para la exclusión de MEF, retire el PBS de los micropocillos y equilibre los pozos con 200 μL de medio basal N2B27 (por 1 chip de micropocillo) e incube durante 60 min a 37 °C.
- Recoja el sobrenadante que contiene las células naïve no unidas, transfiéralo a un tubo de 15 ml y gire hacia abajo las células a 200 x g durante 4 minutos.
- Aspirar los medios y resuspendir las células en 1 mL de medios basales N2B27. Cuente las celdas mediante diapositivas de recuento de celdas. Girar hacia abajo las células a 200 x g durante 4 min.
- Aspirar el medio y añadir una cantidad adecuada de 10 μM Y-27632 contenidos en el medio N2B27 para obtener una densidad celular de 30.000 células por 50 μL.
NOTA: El número óptimo de células de siembra inicial puede variar entre las diferentes líneas celulares. Por ejemplo, para sembrar 50-60 células / micropozo, 30,000 células (incluido el excedente considerando que algunas células caen fuera del micropozo) se siembran en 1 pozo de la placa de 96 pocillos que contiene 430 micropocillos. Un número de células de partida inadecuado puede dar lugar a la formación de pequeños agregados sin cavidad o formación de estructura cavitada que alcanza más de 250 μm. - Aspire el medio a partir de matrices de micropocillos equilibrados y agregue 25 μL de medios N2B27 con 10 μM Y-27632. Añadir 50 μL de suspensión celular e incubar a 37 °C durante 15 min (hasta que las células caigan en el fondo del micropocillo). Luego, agregue 125 μL de medio N2B27 suplementado con 10 μM Y-27632.
- Desarrollo de blastoides
- Dentro de las 24 h, se pueden observar agregados de hPSC naïve (día 0) en el chip del micropocillo. Para iniciar la formación de blastoides, prepare el medio PALLY y siga los pasos que se detallan a continuación.
- Medio PALLY precalentado a 37 °C durante 30 min.
NOTA: El ácido 1-oleoil lisofosfatídico (LPA) y 10 μM Y-27632 deben agregarse justo antes de su uso. La concentración óptima de LPA varía entre 0,5-5 μM. Esto debe ajustarse para las líneas hPSC individuales utilizadas para blastoides. - Aspirar el medio de agregación. Añadir 200 μL de medio PALLY precalentado a los micropocillos. Vuelva a colocar la placa de cultivo celular en una incubadora hipóxica a 37 °C. Repita el cambio de medios el día 1.
- El día 2, retire el medio PALLY y agregue 200 μL de medio N2B27 suplementado con LPA y 10 μM Y-27632.
NOTA: En el día 2 la mayoría de los agregados continúan creciendo. Sin embargo, algunos agregados forman pequeñas cavidades. Cultivo continuo de blastoides en PALLY hasta el día 4 o en medio de cultivo in vitro (IVC1) a partir del día 2. Sin embargo, después de este cambio de medios mejora la formación de PrE en blastoides maduros. - Repita el cambio de medios el día 3. La formación completa de blastoides ocurre en el día 4.
NOTA: Se considera que los blastoides se han desarrollado completamente cuando se han sometido a una morfogénesis completa basada en la morfometría de los blastocistos humanos del día 7 (por ejemplo, un rango de diámetro de 150 -250 μm; un grupo interno rodeado por un epitelio de células similares al trofoectodermo) y han formado células polares similares al trofoectodermo (NR2F2 + / CDX2-) y células similares a PrE (GATA4 +). Esto se puede evaluar mediante tinción por inmunofluorescencia o clasificación celular activada por fluorescencia (FACS).
3. Formación de blastoides en microplacas de fijación ultra baja de 96 pocillos
- Preparar la suspensión celular ingenua de hPSCs para la formación de blastoides siguiendo los pasos descritos anteriormente de 2.1.1 a 2.1.11.
- Aspirar el medio y añadir una cantidad adecuada de medios N2B27 que contengan 10 μM Y-27632 para obtener la densidad celular de 70 células por 100 μL del medio.
NOTA: El número óptimo de células de siembra inicial puede variar entre las diferentes líneas celulares. Por ejemplo, 70 celdas/pozo puede ser el número de celda óptimo para la mayoría de las líneas celulares. - Centrifugar la placa a 200 x g durante 2 min a temperatura ambiente para agrupar las células en el fondo de los pozos.
- Incubar la placa en una incubadora a 37 °C en condiciones de cultivo hipóxico. Dentro de las 24 h se pueden observar agregados de hPSC naïve (día 0) en los pozos.
- Preparar 2x PALLY medio. Agregue 100 μL de medio 2x PALLY precalentado a los pozos.
- Vuelva a colocar la placa de cultivo celular en una incubadora hipóxica a 37 °C. Después de 24 h, aspire la mitad del medio (100 μL) y reemplácelo por 100 μL de medio PALLY precalentado. Repita el paso hasta el día 4. Asegúrese de no aspirar los agregados.
NOTA: El día 2, la mayoría de los agregados continúan creciendo. Sin embargo, algunos agregados tienen pequeñas cavidades llenas de líquido. En el día 4, la mayoría de las estructuras cavitadas se someten a una morfogénesis completa para formar estructuras similares a blastocistos.
4. Formación de trofosferas
- Para la formación de trofoesferas, siga el protocolo de formación de blastoides desde el paso 2.1.1 (exclusión MEF) hasta el paso 2.1.12 (último paso del protocolo de siembra).
- Una vez que se hayan formado agregados de hPSCs naïve después de 24 h, intercambie el medio de agregación con PALY (sin LIF) suplementado con 3 μM SC-144 para la formación de trofoesferas que representan el trofoectodermo temprano y PALLY suplementado con 2 μM XMU-MP-1 para la formación de trofoesferas que representan el trofoectodermo maduro.
- Refresque el medio diariamente. La formación completa de la trofosfera ocurre en el día 4.
5. Análisis del estado de las células blastoides y su etapa reflejada utilizando scRNAseq
- Para recoger blastoides y realizar la disociación, caliente la incubadora de agitación a 37 ° C y configúrela a 100 rpm.
- Recolecte blastoides de la placa inicial de 96 pocillos y transfiéralos a múltiples pozos de una placa de 96 pocillos con fondo en U utilizando una pipeta bucal equipada con un capilar de vidrio.
NOTA: Los blastoides (> 70%) deben seleccionarse en función de los criterios morfométricos (tamaño = 150-250 μm con un grupo interno único) para evitar la contaminación con estructuras no blastoides (< 30%). - Lavar una vez con 200 μL de PBS usando un P200 mediante la visualización bajo un estereomicroscopio. Transferir a un pozo que contenga 50 μL de colagenasa e incubar durante 30 min en la incubadora de agitación.
- Transfiera los blastoides a un pozo con 100 μL de 10x tripsina-EDTA y mezcle bien. Incubar durante 20 min en la incubadora de agitación.
- Disociar los blastoides en una sola celda mediante el uso de pipeta P200. Transfiera las células a un tubo de 15 ml con tampón FACS (1% FBS en PBS).
- Para capturar proporciones específicas de los análogos de los tres linajes, tiñe los análogos TE y PrE con anticuerpos TROP2 y PDGFRa respectivamente.
NOTA: El número de células PrE en blastocistos humanos es menor en comparación con los blastocistos de ratón, lo que podría reflejar defectos de desarrollo de blastocistos formados a través de la fertilización in vitro (FIV) o una diferencia de especie. En los blastoides, los análogos de PrE son menos abundantes que los análogos de TE y EPI y representan el 7,4% de las células al contar las células GATA4+ por imágenes de inmunofluorescencia. Además, el proceso de disociación podría inducir sesgos en las proporciones de los diferentes tipos de células como análogos de PrE para representar el 1% -2% de las células tras la disociación blastoidea, el etiquetado PDGFRa y el análisis FACS. - Células clasificadas por FACS de los tres análogos de linaje en placas 384 que contienen un tampón de lisis para el análisis smart-seq2. Excluya las células muertas marcadas por tinción DAPI (realizada de acuerdo con las instrucciones del fabricante).
- Para evaluar los estados celulares (tipo de célula y etapa de desarrollo), compare los datos transcriptómicos de blastoide con los controles apropiados.
6. Cultivo extendido para evaluar la progresión del desarrollo de los blastoides
- Cultivo de blastoides humanos en placas recubiertas de matriz de membrana basal (fondo de vidrio).
- Cubra la placa con matriz de membrana basal.
- Inspeccione visualmente los blastoides para evaluar y registrar la morfología.
NOTA: Solo los blastoides que muestran la morfología clásica de blastocisto de bola hueca con ICM compacto tienen el potencial de crecer y desarrollarse aún más. - Agregue 100 μL de CMRL medio-1 por un pocillo de la placa de 96 pocillos, y coloque la placa en la incubadora al menos 2 h antes de la transferencia de blastoides para el equilibrio.
- Usando un estereomicroscopio, identifique visualmente los blastoides con buena morfología, transfiera los blastoides seleccionados en un pozo de placa de 96 pocillos que contenga 100 μl de medio CMRL-1 para eliminar los rastros de medios blastoides.
- Transfiera los blastoides al pozo que contiene medios CMRL preequilibrados-1. Coloque la placa en la incubadora e incube a 37 ° C durante la noche.
NOTA: Se pueden cultivar hasta 5 blastoides en un pozo de 96 placas de pozo. Tener demasiados blastoides en un solo pozo puede conducir a la formación de agregados de múltiples blastoides. - Al día siguiente, inspeccione visualmente los blastoides bajo un microscopio. Si los blastoides están unidos, agregue 100 μL de medios CMRL preequilibrados-1 suplementados con matriz de membrana basal al 5%. Coloque la placa en la incubadora e incube a 37 °C durante la noche.
- Al día siguiente, monitoree los blastoides bajo un microscopio. Retire la mitad del medio (100 μL) y reemplácelo con 100 μL de medios CMRL-2 preequilibrados suplementados con matriz de membrana basal al 5%.
- En los días siguientes, reemplace la mitad del medio (100 μL) con medios CMRL-3 preequilibrados suplementados con matriz de membrana basal al 5%. Coloque la placa en la incubadora e incube a 37 °C durante la noche. Repetir todos los días durante un máximo de 4-6 días de cultivo in vitro .
NOTA: Hemos cultivado blastoides durante un máximo de 6 días en condiciones de cultivo extendido, lo que corresponde al tiempo equivalente al día 13 de embriones humanos cultivados in vitro .
7. Blastoides inmunotintivos
- Aspirar el medio. Lave las muestras 3 veces con PBS durante 5 min.
- Agregue 200 μL de paraformaldehído (PFA) al 4% en frío en PBS y fije las muestras durante 30 min a temperatura ambiente. Retire la solución de PFA y lave las muestras 3 veces con PBS durante 10 min.
NOTA: Si los blastoides se cultivaron en chips de micropocillos, transfiera los blastoides del chip a placas de fondo en U de 96 pocillos para los siguientes pasos. - Permeabilizar y bloquear los blastoides en 100 μL de solución bloqueadora por pocillo (PBS que contiene 0,3% de Tritón-X 100 y 10% de suero de burro normal) durante 60 min.
NOTA: Dependiendo de la especie huésped de los anticuerpos, adapte el suero en consecuencia. - Quite la solución de bloqueo. Añadir anticuerpos primarios de 100 μL diluidos en solución de bloqueo fresca e incubar muestras durante la noche a 4°C.
NOTA: Las concentraciones de anticuerpos primarios deben determinarse en función de las instrucciones del fabricante. - Lave las muestras 3x con Triton-X 100 al 0,1% en PBS (solución de lavado) durante 10 min. Añadir 100 μL de anticuerpos secundarios en solución bloqueante junto con 20 μg/ml de tinción nuclear de Hoechst e incubar muestras durante 1 h a temperatura ambiente. Proteja las muestras de la luz.
NOTA: Las concentraciones de anticuerpos secundarios deben determinarse en función de las instrucciones del fabricante. - Lavar las muestras 3x con solución de lavado durante 10 min. Para obtener imágenes, transfiera las muestras al fondo de vidrio μ en PBS.
NOTA: El medio de montaje debe seleccionarse en función del objetivo utilizado para la obtención de imágenes. Por ejemplo, el 80% de glicerol en PBS es posible de usar para montar las muestras mientras se usan objetivos de aceite.
Resultados
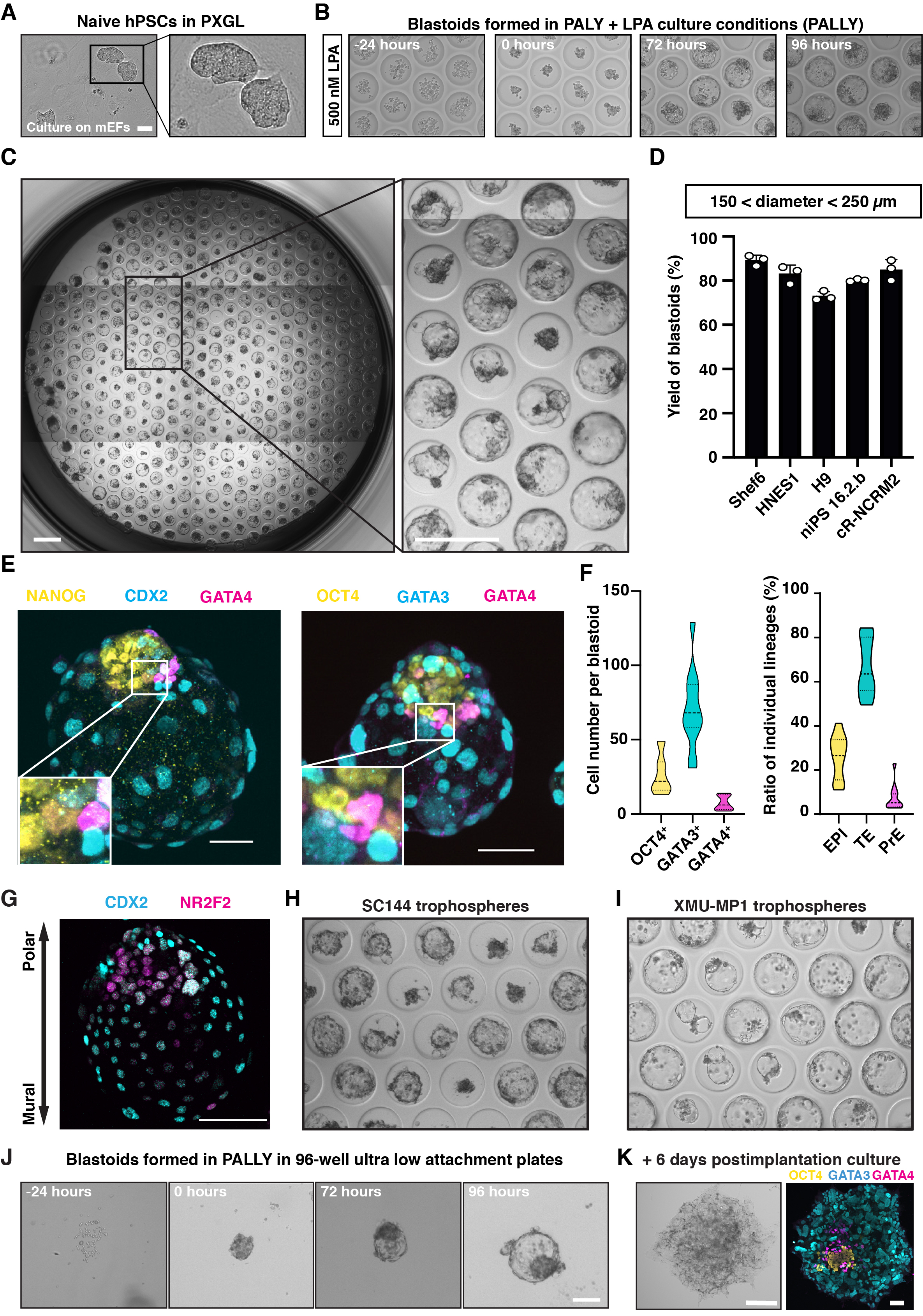
Típicamente, las hPSC naïve cultivadas en PXGL (Figura 2A) son estructuras agregadas y cavitadas que emergen entre 48 y 72 h después de la inducción PALLY y alcanzan un diámetro de 150-250 μm dentro de las 96 h (Figura 2B). Utilizando un número óptimo (1) de células de siembra, (2) la duración de la agregación previa al cultivo con N2B27 (0 a 24 h), (3) la concentración de componentes químicos individuales (especialmente LPA) y (4) la duración del tratamiento con PALLY, la eficiencia de inducción alcanza el 70% -80% según se define en función de los parámetros morfométricos (tamaño total de 150-250 μm, cavidad regular única, grupo de células internas únicas; Figura 2C,D) y la presencia de los tres linajes. Un estado celular inicial subóptimo y / o condiciones de inducción darán como resultado una formación de blastoides menos eficiente o nula. Para garantizar la máxima eficiencia y formar solo análogos preimplantacionales, es crucial utilizar un cultivo de alta calidad de hPSC PXGL ingenuas. Esto se puede evaluar midiendo por FACS el porcentaje de células positivas para los marcadores de superficie SUSD2 (estado ingenuo) y CD24 (estado cebado). Los marcadores de superficie adicionales específicos para los linajes extraembrionarios fuera del objetivo (por ejemplo, amnios, mesodermo extraembrionario) también serían útiles, pero, hasta donde sabemos, actualmente no están disponibles. Si la eficiencia de formación obtenida es inferior a los resultados reportados, es importante revisar cuidadosamente todos los componentes del medio blastoide, especialmente el LPA que se reconstituye en PBS y que, como ligando GPCR, puede ser más inestable en comparación con las moléculas sintéticas reconstituidas en DMSO. En la mayoría de los casos, incluso si el rendimiento no es máximo, las estructuras cavitadas todavía están compuestas por los tres linajes de blastocistos. La aparición de tres linajes de blastocistos y la formación del eje embrionario-abembrionario pueden confirmarse por la tinción de inmunofluorescencia de marcadores (EPI: NANOG, OCT4, TE: GATA3, Polar-TE NR2F2, Mural-TE: CDX2, PrE: GATA4; Figura 2E,G). Las trofosferas, que solo están compuestas de TE, ayudan a diseccionar aún más el papel de la comunicación intercelular. Las trofosferas pueden formarse con una eficiencia del 50% -60% dentro de las 96 h de la inducción (Figura 2H, I). La formación de blastoides se puede realizar no solo en matrices de micropocillos caseras, sino también en placas de pozo 96 de fijación ultra baja disponibles comercialmente con optimización de las condiciones de inducción (consulte el Protocolo y la Figura 2J). Los blastoides también tienen la capacidad de desarrollarse aún más durante 6 días adicionales, que es equivalente en el tiempo del embrión del día 13, con protocolo de diferenciación in vitro (Figura 2K).
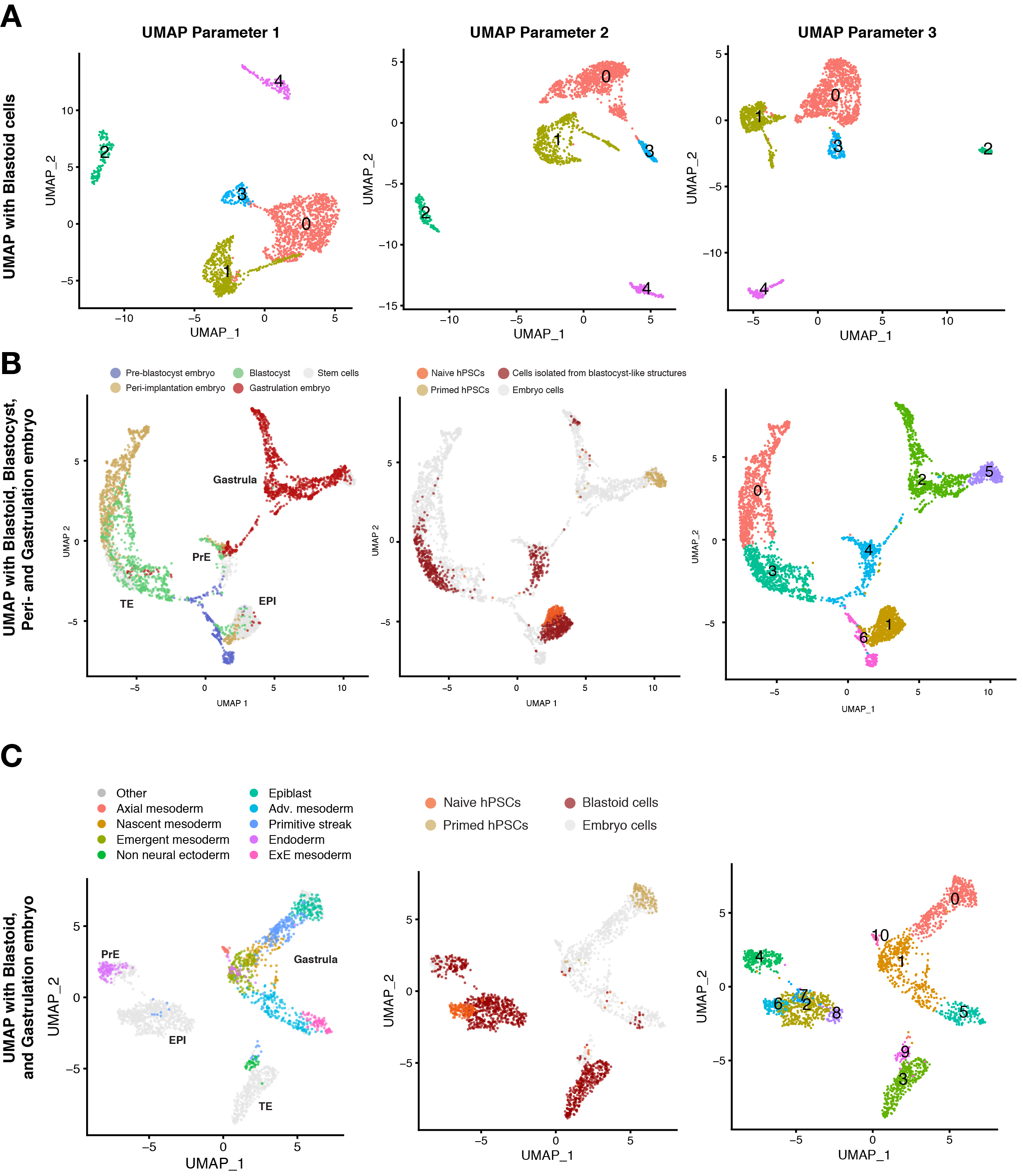
Para caracterizar aún más el estado celular de las células blastoides, se debe utilizar la tecnología de secuenciación de ARN unicelular. UMAP se aplica comúnmente para visualizar una distribución de estados celulares y se realiza un análisis de agrupamiento no supervisado para evaluar la proximidad de estados celulares individuales. Los diferentes parámetros en el análisis de datos de una sola celda pueden afectar la forma en que se muestran las celdas en los UMAP, lo que lleva a clústeres con diferentes posiciones y formas espaciales y relativas (Figura 3A). Sin embargo, en este análisis, las células muestran perfiles de clustering marcadamente reproduciblemente distintos independientemente de los parámetros utilizados para realizar el clustering y la visualización de los datos, lo que permite distinguir con alta confianza los tres linajes de blastocistos. Utilizamos como referencia células de embriones cosechados en diferentes etapas de desarrollo. La fusión de estos conjuntos de datos muestra que la mayoría de los análogos del trofoectodermo de blastoides se agruparon con el trofoectodermo preimplantacional, pero no con los trofoblastos postimplantacionales (Figura 3B). Estos resultados también fueron confirmados por un consorcio independiente10.
Cuando se introducen embriones gastrulatantes en estadio Carnegie 7 (CS7) en el mapa de referencia, una pequeña población de células blastoides (3%) se agrupa con los linajes mesodermo y amnios de estos embriones (Figura 3C). Cuando se introducen células similares a los amnios en el mapa de referencia, una pequeña población de células blastoides (< 2%) se agrupa con tales células similares a los amnios.
En general, solo las estructuras que comprenden una sola cavidad regular, un solo grupo de células internas, un tamaño total que oscila entre 150-250 μm, que comprende análogos transcriptómicos de los tres linajes de blastocistos, y en gran medida desprovistos de otros linajes (por ejemplo, amnios, mesodermo, mesodermo extraembrionario) se consideran blastoides humanos.

Figura 1: Cuatro características y dos enfoques para generar blastoides de alta fidelidad. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Blastoides y trofosferas derivadas de agregados de hPSC ingenuo. (A) Imágenes de contraste de fase que muestran hPSC naïve cultivadas en medio PXGL cocultivadas con MEF irradiado. Barra de escala: 50 μm. (B) Imágenes de contraste de fase que muestran el cambio morfológico de agregados de hPSCs naïve cultivados en una matriz de micropocillos de hidrogel no adherente con LPA de 500 nM (medio PALLY). Barra de escala: 200 μm. (C) Blastoides humanos formados en una matriz de micropocillos después de 96 h. Barras de escala: 400 μm. (D) Cuantificación del porcentaje de micropocillos que contienen un blastoide humano inducido por la condición de cultivo PALLY con concentración optimizada de LPA a partir de diferentes líneas hPSC ingenuas (n = 3 matrices de micropocillos). (E) Tinción de inmunofluorescencia de blastoides humanos con marcadores de epiblasto (EPI) (amarillo) NANOG y OCT4; los marcadores TE (cian) CDX2 y GATA3; y el marcador primitivo de endodermo (magenta) SOX17 y GATA4. Barra de escala: 100 μm. (H-I) Cuantificación del número celular (izquierda) y porcentaje de células (derecha) pertenecientes a cada linaje en blastoide (96 h) en base a tinción de inmunofluorescencia de OCT4, GATA3 y GATA4. (G) Tinción por inmunofluorescencia de blastoides humanos para CDX2 (cian) y NR2F2 (magenta). (F) Imágenes de contraste de fase de trofoes en etapa temprana y tardía en matriz de micropocillos inducidas por adición de 3 μM SC144 (H) o 2 μM XMU-MP-1 (I), respectivamente. (J) Imágenes de contraste de fase que muestran el cambio morfológico de agregados hPSCs ingenuos cultivados en placa de pocillo 96 de fijación ultra baja con 500 nM LPA (medio PALLY). (K) Imagen de contraste de fase (izquierda) y tinción de inmunofluorescencia (derecha) para OCT4 (amarillo), GATA3 (cian) y GATA4 (magenta) en blastoide cultivado en condición de cultivo postimplantacional durante 6 días. Barra de escala: 100 μm. Esta cifra se adapta de 6,10. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Caracterización de la composición de blastoides mediante secuenciación unicelular. (A) Análisis de agrupamiento no supervisado con diferentes parámetros en UMAP del transcriptoma de células individuales derivados de los diferentes puntos de tiempo de blastoide (24 h, 60 h, 96 h), hPSC naïve, hPSCs cebadas y hTSC (representan el citotrofoblasto postimplantación). (B) UMAP del transcriptoma de células derivadas de blastoides (96 h), hPSC naïve y hPSC cebadas integradas con conjuntos de datos publicados de embriones humanos de etapas preimplantacionales, periimplantacionales (blastocistos cultivados in vitro ) y gastrulación (etapa 7 de Carnegie, es decir, entre E16-19). Las células individuales se colorean en función de su origen en embriones humanos (izquierda), células derivadas de blastoides o células madre (centro) y el resultado del análisis de agrupamiento no supervisado (derecha). (C) UMAP del transcriptoma de células derivadas de blastoides, hPSC naïve, hPSCs cebadas, e integrados con el conjunto de datos publicados del embrión en etapa de gastrulación (etapa Carnegie 7, es decir, entre E16-19). Las células individuales se colorean en función de su origen en embriones humanos (izquierda), células derivadas de blastoides o células madre (centro) y el resultado del análisis de agrupamiento no supervisado (derecha). Esta cifra está adaptada de6. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Tabla suplementaria 1: Toda la composición de los medios utilizada en este estudio. Haga clic aquí para descargar esta tabla.
Discusión
En el presente estudio, mostramos, paso a paso, cómo establecer blastoides humanos con altas eficiencias utilizando un protocolo simple y robusto. Tras la agregación de hPSC PXGL naïve y su triple inhibición, los blastoides se forman de manera eficiente (> 70%) y generan secuencialmente los 3 análogos de blastocistos en 4 días. Las limitaciones en la eficiencia y la calidad de los blastoides (por ejemplo, la presencia de células fuera del objetivo) pueden ocurrir si el estado inicial es subóptimo. Cabe destacar que hemos medido que las hPSC PXGL contienen aproximadamente el 5% de las células que reflejan las etapas posteriores a la implantación. Estas células podrían limitar la formación de blastoides de alta calidad. Más allá del estado inicial ingenuo de PXGL que refleja el epiblasto blastocisto, otro factor fundamental es el medio utilizado para la formación de blastoides. Con el fin de formar rápidamente células similares a blastocistos y prevenir la formación de células similares a las postimplantaciones fuera del objetivo, proponemos que la inhibición de las vías triples (Hippo, ERK, TGF-β) es esencial. Mientras que las diferentes líneas celulares dan diferentes rendimientos de blastoideo en la inhibición de ERK / TGF-β (generalmente alrededor del 10% -20%), la exposición a LPA da como resultado la formación de un rendimiento de blastoides igualmente alto en todas las líneas celulares, mientras se utilizan estrictos criterios morfométricos y de especificación de linaje. LpA posiblemente actúa sobre la inhibición de la vía del hipopótamo, que juega un papel crítico en la primera segregación del linaje entre los linajes epiblasto y trofetodermo enratones y humanos 8,51. La mejora significativa de la eficiencia de los blastoides por LPA sugiere que los mecanismos de especificación de células internas-externas mediadas por la vía de Hippo en juego en el blastocisto son cooptados durante la formación de blastoides. Una limitación actual radica en el hecho de que, debido a una suboptimidad de los protocolos utilizados para cultivar blastocistos humanos o blastoides en el día 7-13 equivalente a un tiempo (después de la formación de blastocistos/blastoides), no somos capaces de evaluar hasta qué punto podemos modelar adecuadamente el desarrollo post-implantación.
El análisis del estado transcriptómico de las células blastoides se puede lograr fácilmente utilizando scRNAseq, mapas de referencia adecuados y métodos bioinformáticos. Anteriormente, el análisis transcriptómico mostró que las hPSC cultivadas en PXGL son más similares al epiblasto blastocisto en comparación con el estado cebado. Las limitaciones en el análisis de los datos pueden ocurrir si el mapa de referencia solo comprende células en etapa de blastocisto. El mapa de referencia debe incluir células procedentes de embriones postimplantados con el fin de evaluar la presencia de posibles células fuera del objetivo. En el futuro, con el fin de comparar las células blastoides, un mapa de referencia que incluya todos los tejidos del conceptus humano pre y postimplantación sería extremadamente valioso. Además, los mapas de referencia multiómicos de una sola célula, por ejemplo, incluyendo el transcriptoma, la accesibilidad a la cromatina y la metilación del ADN, ayudarían aún más. Finalmente, los métodos bioinformáticos estandarizados para evaluar cuantitativamente las similitudes entre las células de los modelos embrionarios y los conceptos de referencia, y para identificar positivamente las células fuera del objetivo ayudarían aún más a analizar y comparar los resultados de manera imparcial.
En conjunto, los blastoides formados por la triple inhibición de las vías Hippo, TGF-β y ERK poseen las cuatro características de 1) morfogénesis altamente eficiente, 2) secuencia correcta de especificación de linaje, 3) alta pureza de células similares a blastocistos a nivel de transcriptoma, 4) capacidad para modelar el desarrollo periimplantacional. Estas características de los blastoides facilitarán la construcción de hipótesis sobre el desarrollo e implantación de blastocistos, sin embargo, no recapitulan las primeras etapas del desarrollo embrionario. En contraste con la limitada accesibilidad y versatilidad del blastocisto humano, los blastoides son susceptibles de exámenes genéticos y farmacológicos para las investigaciones funcionales del desarrollo e implantación de blastocistos. En el futuro, estos conocimientos básicos podrían contribuir a mejorar la formulación de medios de FIV, desarrollar anticonceptivos posteriores a la fertilización y gestionar mejor el embarazo temprano.
Divulgaciones
El Instituto de Biotecnología Molecular de la Academia de Ciencias de Austria ha presentado una solicitud de patente EP21151455.9 que describe los protocolos para la formación de blastoides humanos y el ensayo de interacción blastoide-endometrio. HK, AJ, HHK y NR son los inventores de esta patente. Todos los demás autores no declaran intereses contrapuestos.
Agradecimientos
Este proyecto ha recibido financiación del Consejo Europeo de Investigación (ERC) en el marco del programa de investigación e innovación Horizonte 2020 de la Unión Europea (acuerdo de subvención ERC-Co n.º 101002317 'BLASTOID: una plataforma de descubrimiento para la embriogénesis humana temprana'). H.H.K. cuenta con el apoyo del Fondo Austriaco para la Ciencia (FWF), Lise Meitner Programme M3131-B. Agradecemos a Yasuhiro Takashima por compartir las líneas celulares H9 y H9-GFP, y a Austin Smith, Peter Andrews y Ge Guo por compartir las líneas celulares HNES1, Shef6, niPSC 16.2b y cR-NCRM2. Agradecemos a Hossein Baharvand por compartir los organoides endometriales. Agradecemos a Joshua M. Brickman por compartir el ARN aislado de células diferenciadas de PrE y células nEND. Agradecemos a Shankar Srinivas por compartir los datos de secuenciación de ARN unicelular del embrión de perigastrulación. Agradecemos a Aleksand Bykov y Luisa Cochella por la asistencia técnica para la preparación de la biblioteca SMARTSeq2. Agradecemos a las instalaciones de NGS, Biooptic y Stem Cell en IMBA por su asistencia crítica.
Materiales
| Name | Company | Catalog Number | Comments |
| Neurobasal media | in house | ||
| DMEM/F12 | in house | ||
| 100X N2 supplemen | Gibco | 17502048 | |
| 50X B27 supplement | Gibco | 17504044 | |
| 100X Glutamax | Gibco | 35050038 | |
| 100 mM Sodium Pyruvate | Gibco | 11360039 | |
| MEM-Non-essential amino acids | Gibco | 11140050 | |
| 1 M Hepes | in house | ||
| 50 mM 2-Mercaptoethanol | Thermofisher | 31350010 | |
| 100X Penicillin-Streptomycin | Sigma-Aldrich | P0781 | |
| Bovine Serum Albumin solution | Sigma-Aldrich | A7979 | |
| PD0325901 | Medchem express | HY-10254 | |
| XAV-939 | Medchem express | HY-15147 | |
| Gö 6983 | Medchem express | HY-13689 | |
| Human recombinant Leukemia Inhibitory Factor | in house | ||
| A83-01 | Medchem express | HY-10432 | |
| 1-Oleoyl Lysophosphatidic acid (LPA) | Peprotech | 2256236 | |
| Y-27632 | Medchem express | HY-10583 | |
| CMRL medium | Gibco | 21530027 | |
| Fetal Bovine Serum (FBS) | Sigma-Aldrich | F7524 | |
| KnockOut Serum Replacement (KSR) | Thermofisher | 10-828-028 | |
| Accutase | Biozym | B423201 | cell detachment solution |
| Geltrex | Thermofisher | A1413302 | growth factor basement membrane extract |
| TROP2 antibody | R&D systems | MAB650 | |
| PDGFRα antibody | R&D systems | AF307 | |
| SC-144 | Axon | 2324 | |
| XMU-MP-1 | Med Chem Express | HY-100526 | |
| Matrigel | basement membrane matrix | ||
| Countess cell counting chamber slides | Thermo fisher | cell counting slides | |
| DAPI Staining Solution | Miltenyi Biotec | 130-111-570 |
Referencias
- Rivron, N., et al. Debate ethics of embryo models from stem cells. Nature. 564 (7735), 183-185 (2018).
- Hyun, I., Munsie, M., Pera, M. F., Rivron, N. C., Rossant, J. Toward Guidelines for Research on Human Embryo Models Formed from Stem Cells. Stem Cell Reports. 14 (2), 169-174 (2020).
- Clark, A. T., et al. Human embryo research, stem cell-derived embryo models and in vitro gametogenesis: Considerations leading to the revised ISSCR guidelines. Stem Cell Reports. 16 (6), 1416-1424 (2021).
- Lovell-Badge, R., et al. ISSCR Guidelines for Stem Cell Research and Clinical Translation: The 2021 update. Stem Cell Reports. 16 (6), 1398-1408 (2021).
- Rivron, N. C., et al. Blastocyst-like structures generated solely from stem cells. Nature. 557 (7703), 106-111 (2018).
- Kagawa, H., et al. Human blastoids model blastocyst development and implantation. Nature. , 04267-04268 (2021).
- Meistermann, D., et al. Integrated pseudotime analysis of human pre-implantation embryo single-cell transcriptomes reveals the dynamics of lineage specification. Cell Stem Cell. 28 (9), 1625-1640 (2021).
- Gerri, C., et al. Initiation of a conserved trophectoderm program in human, cow and mouse embryos. Nature. 587 (7834), 443-447 (2020).
- Gerri, C., Menchero, S., Mahadevaiah, S. K., Turner, J. M. A., Niakan, K. K. Human Embryogenesis: A Comparative Perspective. Annual Review of Cell and Developmental Biology. 36, 411-440 (2020).
- Zhao, C., et al. Reprogrammed iBlastoids contain amnion-like cells but not trophectoderm. bioRxiv. , 2021.05.07.442980(2021).
- Zijlmans, D. W. Integrated multi-omics reveal polycomb repressive complex 2 restricts human trophoblast induction. Nat. Cell Biol. 24, 858-871 (2022).
- Nichols, J., Smith, A. Naive and primed pluripotent states. Cell Stem Cell. 4 (6), 487-492 (2009).
- Guo, G., et al. Epigenetic resetting of human pluripotency. Development. 144 (15), Cambridge, England. 2748-2763 (2017).
- Takashima, Y., et al. Resetting transcription factor control circuitry toward ground-state pluripotency in human. Cell. 158 (6), 1254-1269 (2014).
- Thomson, J. A. Embryonic stem cell lines derived from human blastocysts. Science. 282 (5391), New York, N.Y. 1145-1147 (1998).
- Yang, Y., et al. Derivation of Pluripotent Stem Cells with In Vivo Embryonic and Extraembryonic Potency. Cell. 169 (2), 243-257 (2017).
- Stirparo, G. G., et al. Integrated analysis of single-cell embryo data yields a unified transcriptome signature for the human pre-implantation epiblast. Development. 145 (3), Cambridge, England. 158501(2018).
- Castel, G., et al. Induction of Human Trophoblast Stem Cells from Somatic Cells and Pluripotent Stem Cells. Cell Reports. 33 (8), 108419(2020).
- Posfai, E., et al. Evaluating totipotency using criteria of increasing stringency. Nature Cell Biology. 23 (1), 49-60 (2021).
- Nakamura, T., et al. A developmental coordinate of pluripotency among mice, monkeys and humans. Nature. 537 (7618), 57-62 (2016).
- Theunissen, T. W., et al. Molecular Criteria for Defining the Naive Human Pluripotent State. Cell Stem Cell. 19 (4), 502-515 (2016).
- Guo, G., et al. Naive Pluripotent Stem Cells Derived Directly from Isolated Cells of the Human Inner Cell Mass. Stem Cell Reports. 6 (4), 437-446 (2016).
- Rossant, J. Genetic Control of Early Cell Lineages in the Mammalian Embryo. Annual Review of Genetics. 52, 185-201 (2018).
- De Paepe, C., et al. Human trophectoderm cells are not yet committed. Human reproduction. 28 (3), 740-749 (2013).
- Amita, M., et al. Complete and unidirectional conversion of human embryonic stem cells to trophoblast by BMP4. Proceedings of the National Academy of Sciences of the United States of America. 110 (13), 1212-1221 (2013).
- Io, S., et al. Capturing human trophoblast development with naive pluripotent stem cells in vitro. Cell Stem Cell. 28 (6), 1023-1039 (2021).
- Guo, G., et al. Human naive epiblast cells possess unrestricted lineage potential. Cell Stem Cell. 28 (6), 1040-1056 (2021).
- Liu, X., et al. Modelling human blastocysts by reprogramming fibroblasts into iBlastoids. Nature. 591 (7851), 627-632 (2021).
- Hirate, Y., et al. Polarity-dependent distribution of angiomotin localizes Hippo signaling in preimplantation embryos. Current biology: CB. 23 (13), 1181-1194 (2013).
- Cockburn, K., Biechele, S., Garner, J., Rossant, J. The Hippo pathway member Nf2 is required for inner cell mass specification. Current Biology: CB. 23 (13), 1195-1201 (2013).
- Xu, R. H., et al. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nature Biotechnology. 20 (12), 1261-1264 (2002).
- Krendl, C., et al. GATA2/3-TFAP2A/C transcription factor network couples human pluripotent stem cell differentiation to trophectoderm with repression of pluripotency. Proceedings of the National Academy of Sciences of the United States of America. 114 (45), 9579-9588 (2017).
- Xu, R. H., et al. Basic FGF and suppression of BMP signaling sustain undifferentiated proliferation of human ES cells. Nature Methods. 2 (3), 185-190 (2005).
- Horii, M., Bui, T., Touma, O., Cho, H. Y., Parast, M. M. An Improved Two-Step Protocol for Trophoblast Differentiation of Human Pluripotent Stem Cells. Current Protocols in Stem Cell Biology. 50 (1), 96(2019).
- Okae, H., et al. Derivation of Human Trophoblast Stem Cells. Cell Stem Cell. 22 (1), 50-63 (2018).
- Yamanaka, Y., Lanner, F., Rossant, J. FGF signal-dependent segregation of primitive endoderm and epiblast in the mouse blastocyst. Development. 137 (5), 715-724 (2010).
- Kuijk, E. W., et al. The roles of FGF and MAP kinase signaling in the segregation of the epiblast and hypoblast cell lineages in bovine and human embryos. Development. 139 (5), Cambridge, England. 871-882 (2012).
- Roode, M., et al. Human hypoblast formation is not dependent on FGF signalling. Developmental Biology. 361 (2), 358-363 (2012).
- Linneberg-Agerholm, M., et al. Naïve human pluripotent stem cells respond to Wnt, Nodal and LIF signalling to produce expandable naïve extra-embryonic endoderm. Development. 146 (24), Cambridge, England. (2019).
- Yu, L., et al. Blastocyst-like structures generated from human pluripotent stem cells. Nature. 591 (7851), 620-626 (2021).
- Yanagida, A., et al. Naive stem cell blastocyst model captures human embryo lineage segregation. Cell Stem Cell. 28 (6), 1016-1022 (2021).
- Zappia, L., Oshlack, A. Clustering trees: a visualization for evaluating clusterings at multiple resolutions. GigaScience. 7 (7), (2018).
- Subramanian, A., et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proceedings of the National Academy of Sciences of the United States of America. 102 (43), 15545-15550 (2005).
- Messmer, T., et al. Transcriptional Heterogeneity in Naive and Primed Human Pluripotent Stem Cells at Single-Cell Resolution. Cell Reports. 26 (4), 815-824 (2019).
- Zhou, F., et al. Reconstituting the transcriptome and DNA methylome landscapes of human implantation. Nature. 572 (7771), 660-664 (2019).
- Petropoulos, S., et al. Single-Cell RNA-Seq Reveals Lineage and X Chromosome Dynamics in Human Preimplantation Embryos. Cell. 167 (1), 285(2016).
- Tyser, R. C. V., et al. A spatially resolved single cell atlas of human gastrulation. bioRxiv. , (2020).
- Turco, M. Y., et al. hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nature Cell Biology. 19 (5), 568-577 (2017).
- Boretto, M., et al. Development of organoids from mouse and human endometrium showing endometrial epithelium physiology and long-term expandability. Development. 144 (10), Cambridge, England. 1775-1786 (2017).
- Ma, H., et al. In vitro culture of cynomolgus monkey embryos beyond early gastrulation. Science. 366 (6467), New York, N.Y. (2019).
- Nishioka, N., et al. The Hippo signaling pathway components Lats and Yap pattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass. Developmental Cell. 16 (3), 398-410 (2009).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoExplorar más artículos
This article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados