
Method Article
Protocolo para blastóides humanos modelando desenvolvimento e implantação de blastocisto
* Estes autores contribuíram igualmente
Neste Artigo
Resumo
Um protocolo delineando a formação de blastóides humanos que, eficientemente, oportuna e sequencialmente geram células semelhantes a blastocistos.
Resumo
Um modelo do blastocisto humano formado a partir de células-tronco (blastoid) apoiaria avanços científicos e médicos. No entanto, seu poder preditivo dependerá de sua capacidade de recapitular eficiente, oportuna e fielmente as sequências de desenvolvimento de blastocisto (morfogênese, especificação, padronização) e formar células que refletem o estágio blastocisto. Aqui mostramos que células-tronco pluripotentes humanas ingênuas cultivadas em condições PXGL e, em seguida, inibidas triplamente para o Hipopótamo, transformando o fator de crescimento- β e caminhos de quinase extracelular regulados por sinais sofrem eficientemente de morfogênese para formar blastóides (>70%). Combinando com o tempo de desenvolvimento (~4 dias), os blastóides desenrolam a sequência blastocisto de especificação produzindo análogos do trophoblasto e do epiblasto, seguidos pela formação de análogos do endoderme primitivo e dos trophoblastos polares. Isso resulta na formação de células transcricionalmente semelhantes ao blastocisto (>96%) e uma minoria de análogos pós-implantação. Os blastóides padronizam eficientemente formando o eixo embrionário-abembrínico marcado pela maturação da região polar (NR2F2+), que adquire o potencial específico para se ligar direcionalmente às células endometrial estimuladas hormonalmente, como no útero. Tal blastóide humano é um modelo escalável, versátil e ético para estudar o desenvolvimento humano e a implantação in vitro.
Introdução
A falta de modelos experimentais limitou a compreensão da embriogênese humana primitiva. O conhecimento atual dos aspectos específicos humanos do desenvolvimento embrionário é derivado de embriões de fertilização in vitro (FIV) excedentes doados para pesquisa. No entanto, a disponibilidade limitada, as dificuldades das manipulações experimentais e a qualidade variável dos embriões dificultam as investigações científicas. Pelo contrário, um modelo in vitro fiel de embriões humanos permitiria manipulações experimentais complexas, proporcionando assim uma oportunidade ética para complementar a pesquisa sobre embriões humanos 1,2,3,4. Um modelo previamente desenvolvido de blastocistos de camundongos combinou células-tronco embrionárias de camundongos e células-tronco trophoblastos5. Neste protocolo detalhado, um método para gerar um modelo do blastocisto humano a partir de células-tronco pluripotentes ingênuos que são fiéis aos critérios elementares de blastocisto é descrito6.
Quatro critérios para blastóides humanos. Aqui, na tentativa de estabelecer uma definição padronizada de blastoids humanos, propomos quatro critérios mínimos. Embora não exaustivos, esses critérios podem servir de base para avaliar os parâmetros que permitem a formação de blastóides humanos (Figura 1A). (1) Os blastóides devem formar-se eficientemente em termos de morfologia e de geração dos análogos das três linhagens, ou seja, epiblasto (Epi), trophectoderme (TE) e endoderme primitivo (PrE). É provável que a ineficiência aponte para um estado celular inicial inadequado ou/e condição de cultura (por exemplo, meio blastóide). (2) Os blastóides devem gerar análogos das três linhagens de acordo com a sequência de desenvolvimento (Epi/TE primeiro, PrE/polarTE passado)7,8 e tempo (indução ~ 3 dias; dias embrionários 5-7)7,9. (3) Os blastóides devem formar análogos do estágio blastocisto, mas não de estágios pós-implantação (por exemplo, epiblasto pós-implantação, trophoblasto ou células de amnion). (4) Finalmente, os blastóides devem ser capazes de recapitular características funcionais de implantação e desenvolvimento de blastocisto. Usando este protocolo, os blastóides humanos formam-se eficientemente usando múltiplas linhas celulares (>70%), são capazes de gerar os analógicos celulares blastocistos sequencialmente e dentro de 4 dias, e os analógicos são transcriçãomente semelhantes ao estágio blastocisto (>96% com base em várias análises)6,10,11. Finalmente, os blastóides geram robustamente o eixo embrionário-abembrínico, que lhes permite interagir com células endometrial hormonalmente estimuladas através da região polar, e expandir robustamente as linhagens sobre a cultura estendida (tempo equivalente: dia embrionário 13).
A importância do estado celular inicial. As células-tronco pluripotentes humanas (hPSCs) podem ser estabilizadas em diferentes estados que tentam capturar estágios precisos de desenvolvimento. Esses estados são sustentados por condições culturais que, embora ainda subótimas, restringem as células em um epiblasto pré-implantação (~ dias embrionários 5-7) ou pós-implantação (~ dias embrionários 8-14) estágio12. A análise transcriômica mostrou que os hPSCs cultivados em PD0325901, XAV939, Gö6983 e fator inibidor da leucemia (LIF; denominados PXGL ingênuos hPSCs)13,14 são mais semelhantes ao epiblasto blastocisto em comparação com hPSCs cultivados em fator de crescimento do fibroblasto (FGF) 2 e activina15 (denominados hPSCs12) e às células-tronco pluripotentes estendidas humanas (hEPSCs)16 (ver análise em referências17, 18,19). Assim, o transcriptome de hPSCs preparados combina melhor com um epiblasto de macaco cinomolgus pós-implantação/pré-gastrulação20. Critérios moleculares adicionais, como expressão transposon, metilação de DNA e estado cromossomo X, confirmaram que variações do estado ingênuo se assemelham mais ao epiblasto do blastocisto em comparação com o estadoprimed 17,21. Finalmente, as linhas de hPSCs ingênuos foram derivadas diretamente de blastocistos usando as condições de cultura PXGL22.
As células de blastocisto humanos ainda não foram comprometidas. A especificação de linhagem murina ocorre a partir do estágio de morula que precede o estágio blastocisto23. Pelo contrário, experimentos de dissociação e reagregação mostraram que as células de trophectoderm humanas de blastocistos precoces ainda não estão comprometidas24. Assim, a análise das células dos blastocistos humanos por sequenciamento de RNA unicelular (scRNAseq) mostrou que a primeira especificação de linhagem (trophoblast/epiblasto) ocorre após a formação da cavidade blastocisto7. Esta especificação humana diferida se correlaciona com observações de que os hPSCs são potentes para formar trophoblastos 25,26,27 quando os PSCs do rato estão em grande parte comprometidos com a linhagem do epiblasto. Essas observações combinadas levaram à possibilidade de que os hPSCs ingênuos refletissem um estágio blastocisto e mantivessem o potencial de formar as três linhagens blastocistas. Ultimamente, a potência dos hPSCs para especificar análogos extraembrínicos tem sido proposta para mudar de trophectoderm para amnion durante a progressão de estado ingênuo para primed27. Assim, os hPSCs ingênuos são mais semelhantes ao estágio de pré-implantação 17,18,21 e têm uma capacidade aprimorada de formar trophoblasts em comparação com os hPSCs27, hEPSCs16 ou estados reprogramados intermediários28, que são propensos a formar análogos pós-implantação 10 (Figura 1B ). O estado celular inicial é, portanto, crucial para formar os análogos extraembrínicos apropriados. Embora uma análise completa lado a lado dos análogos de trophectoderme convertidos ainda esteja a ser feita, um estado ingênuo PXGL refletindo o blastocisto inicial parece importante para formar blastóides de alta fidelidade.
Solicitando especificação e morfogênese sinalizando inibição de caminhos. A inibição da via de sinalização do Hipopótamo é um mecanismo conservado que conduz a especificação do trophoblasto em camundongos, vacas e humanos 9,29,30. Além disso, desde 2013, sabe-se que a inibição do NODAL (A83-01) e da quinase extracelular regulada por sinal (ERK; PD0325901 ou equivalente) e a ativação das vias de sinalização da proteína morfogenética óssea (BMP) desencadeiam hPSCs preparados para ativar a rede transcricional associada à linhagem trophoblast 25,31,32,33,34. Além disso, recentemente vários relatórios também confirmaram que a inibição tanto da via NODAL quanto da ERK e da ativação do BMP facilitam a diferenciação do trophoblasto dos ingênues hPSCs 25,31,32,33,34. Finalmente, se a especificação do trophoblasto é desencadeada a partir de um estado ingênuo, as células recapitulam aspectos da progressão do desenvolvimento do trophectoderme26. No entanto, linhas auto-renovadores que refletem o trophectoderm blastocisto não foram estabilizadas in vitro. Após a especificação do trophoblasto, a indução do fator de crescimento epidérmico (EGF) e as vias de sinalização WNT, juntamente com a inibição do HDAC, podem facilitar a progressão do desenvolvimento do trophoblasto34,35 e estabilizar as células em linhas de células-tronco trofoblastos humanos (hTSCs) refletindo citotrofoblastos pós-implantação18,35. Tais linhas podem ser derivadas tanto de blastocistos quanto de tecidos placentários35.
A segunda linhagem extraembriônica, denominada PrE, é especificada após trophoblastos e se origina do epiblasto 7,9. Ao contrário da Murine PrE36, acredita-se que a contraparte humana seja independente da sinalização FGF 37,38. Linhas que refletem o endodermo extraembriônico (denominado nEnd) foram estabelecidas a partir de hPSCs ingênuos por indução de vias de sinalização usando ativação A, Wnt e LIF39. Inconsistente com os experimentos de inibição de embriões, a inibição do ERK tem sido demonstrada para evitar a formação de tais células nEND in vitro39. Até agora, tais linhas não foram derivadas diretamente de blastocistos.
Ultimamente, os modelos do embrião inicial têm sido formados pela combinação de variações das mídias anteriormente desenvolvidas para as células35 e nEND39, utilizando assim ativadores do fator de crescimento transformador- β (TGF-β), EGF e vias de sinalização Wnt28,40. Esses modelos de embrião formam-se em baixa eficiência (10%-20%) e formam células semelhantes ao estágiopós-implantação 10, incluindo análogos do epiblasto pós-implantação, trophoblasto, amnion, gastrula, tecidos mesodérmicos (~ dia embrionário 14) e citotrofoblastos10. Pelo contrário, uma inibição tripla das vias Hippo, ERK e TGF-β guia eficientemente a formação de blastoids que compreendem células blastocistos41. Juntamente com o estado celular inicial, propomos que a inibição de vias triplas (Hipopótamo, ERK, TGF-β) é o segundo parâmetro essencial para formar blastóides de alta fidelidade (Figura 1B).
Avaliação do estado celular e estágio refletido usando scRNAseq. Os estados das células que compõem blastóides podem ser avaliados através da análise scRNAseq. Sua semelhança transcricional com estágios embrionários específicos pode ser medida apenas usando células blastoid e em comparação com hPSCs ou hTSCs que refletem os estágios pós-implantação20,35. A realização de análises de cluster usando diferentes níveis de definição revela como as subpopulações se fundem progressivamente quando a definição diminui, revelando assim as semelhanças dos clusters. Embora a idealidade no número de clusters possa ser medida42, o clustering de alta resolução também informa sobre a eventual presença de pequenas subpopulações anormais, por exemplo, refletindo os estágios pós-implantação10. Os genes expressos diferencialmente entre clusters podem fornecer informações sobre seus análogos no processo de desenvolvimento, avaliando os níveis de expressão de conjuntos genéticos de referência que definem linhagens específicas do estágio. Isso permite medir o enriquecimento de subpopulações blastóides através de mapas de distância não supervisionados (por exemplo, usando genes enriquecidos superiores) ou por análise de enriquecimento de conjunto genético (GSEA)43. Usando este protocolo blastoid, apenas três clusters principais formam que refletem transcrição as três linhagens blastocisto. Um cluster inclui tanto os hPSCs ingênuos iniciais quanto o epiblasto analógico dos blastóides. A análise de células em diferentes pontos de tempo mostrou a natureza sequencial da especificação de linhagens (os trophoblastos começam a especificar dentro de 24 h, e células primitivas de endoderme dentro de 60 h). Um agrupamento de alta resolução capturou uma subpopulação de células (3,2%) expressando genes específicos para embriões de estágio gastrulação (possivelmente mesoderme ou amnion). Note-se que os hPSCs in ingênuos iniciais também consistiram em 5% das células pós-implantação, como descrito anteriormente44. Em uma segunda análise, as células blastóides podem ser fundidas em silico com células de referência isoladas do concepti em diferentes estágios 45,46,47, a fim de inferir a equivalência do estágio. Aqui, foram utilizadas células isoladas do conceito pré-implantação45,46, dos blastocistos in vitro cultivados 45 e dos embriõesembriões em estágio gastrulatório 47. Usando este protocolo, foi quantificado que as células blastóides incompatíveis reveladas por clustering de alta resolução de fato agrupam-se com mesoderme pós-implantação e amnion. Em etapas futuras, o benchmarking do transcriptome deve ser complementado com a análise da expressão transposon, da metilação do DNA e do status do cromossomo X que também fornecem marcos dos estágios de desenvolvimento21.
Avaliação da formação do eixo e outras funcionalidades dos blastóides humanos. Um blastocisto maduro é caracterizado pela formação dos trophoblastos de padronização do eixo embrionário-abembrínico para implantação. Usando este protocolo blastoid, um eixo se forma robustamente exemplificado por um amadurecimento dos trofoblastos proximais (por exemplo, NR2F2+/CDX2-) que adquirem a capacidade de anexar às células organoides endometrial apenas quando são estimulados hormonalmente48,49. A comparação com troforesferas que não formam o epiblasto mostra que essas células internas induzem os trophoblastos de abutting a amadurecer de modo a mediar o apego inicial ao endométrio. Quando cultivadas em um meio de cultura estendida projetado para blastocistos de macaco cinomolgus50, todas as três linhagens do blastoid se expandem consistentemente por seis dias adicionais (tempo equivalente ao dia 13), embora sua organização não reflita esse estágio de desenvolvimento.
A implicação de alta eficiência e alta fidelidade de blastóides humanos. A conservação de princípios de desenvolvimento descobertos em organismos modelo é inerentemente difícil de testar no conceito humano devido ao acesso restrito e às dificuldades técnicas em manipulá-lo geneticamente e fisicamente. Um modelo blastóide de alta eficiência e alta fidelidade permitiria telas genéticas e medicamentos de alto rendimento, que estão na base de descobertas científicas e biomédicas. Além disso, a incorporação de complexas modificações genéticas para alterar e registrar processos biológicos complementaria tais estudos. No geral, propomos que a inibição tripla (Hippo, TGF-β, ERK) dos ingênuos HPSCs PXGL seja condutiva para a formação eficiente de blastóides humanos de alta fidelidade que cumprem os quatro critérios mínimos. A natureza escalável e versátil deste protocolo torna-o adequado para gerar hipóteses direcionadas que podem então ser validadas usando blastocistos humanos. Como tal, os blastóides humanos não substituirão o uso do conceito humano para pesquisas in vitro , mas podem atuar como uma maneira poderosa de canalizar a pesquisa através de abordagens experimentais anteriormente inacessíveis no centro do processo de descoberta científica e biomédica. O protocolo mostra como formar blastóides humanos e também como analisar as células que estão contidas no blastoid.
Protocolo
As Diretrizes para Pesquisa de Células-Tronco e Tradução Clínica da Sociedade Internacional para Pesquisa de Células-Tronco (ISSCR) recomendam que a pesquisa sobre blastóides humanos só seja permitida após revisão e aprovação por meio de um processo especializado de revisão científica e ética 3,4. Todos os procedimentos experimentais foram conduzidos seguindo as diretrizes do comitê de ética em pesquisa humana do Instituto de Biotecnologia Molecular da Academia Austríaca de Ciências (IMBA) sob a aprovação Rivron_Stellungnahme_2020-04-22. O cumprimento dessas diretrizes é necessário para a publicação de resultados de pesquisas em revistas científicas.
1. Cultura de célula-tronco embrionária ingênua humana em condição PXGL
NOTA: Os hPSCs ingênuos podem ser obtidos em laboratórios relevantes. As linhas utilizadas aqui foram obtidas nos laboratórios de Yasuhiro Takashima (atualmente na CiRA, Kyoto, Japão) e de Austin Smith (atualmente no Living Systems Institute, Exeter, Reino Unido). Alternativamente, os hPSCs ingênuos podem ser redefinidos em casa a partir de linhas de hPSCs primed, como descrito anteriormente13,14. Os hPSCs ingênuos parecem estáveis para múltiplas passagens (> 15), mas a qualidade da cultura pode variar com o tempo. Se a qualidade do hPSC ingênuo diminuir, descongele um novo frasco de células ou gere hPSCs ingênuos de novo a partir de PSCs preparados. Para todas as composições de mídia, consulte a Tabela Suplementar 1.
- Preparação da camada alimentador embrionário de camundongos irradiados (MEFs)
- Um dia antes da aprovação de hPSCs ingênuos, prepare uma placa de cultura celular de 6 poços com camadas de MEF irradiadas seguindo os passos descritos abaixo.
- Cubra uma placa de cultura celular de 6 poços com 1 mL de gelatina de 0,1% em PBS por poço. Incubar a placa a 37 °C por 30 min. Remova a solução de gelatina.
- Prepare o meio MEF a 37 °C.
- Descongele mefs em um banho de água a 37 °C até que apenas uma pequena moita de gelo seja deixada. Dissolva o volume do frasco com 1 mL de meio MEF preparado usando uma pipeta P1000.
- Transfira a suspensão da célula para um tubo de 15 mL. Abaixe a suspensão em 200 x g por 4 min. Aspire o supernasce e resuspenda a pelota MEF adicionando meio MEF fresco (suficiente para 1,5 mL por poço).
- Conte as células usando slides de contagem de células e adicione 300.000 células por poço e transfira a placa para uma incubadora normoxy a 37 °C.
NOTA: Se os MEFs se desprendem ao longo do tempo, mefs frescos podem ser adicionados ao meio PXGL durante a mudança de mídia de rotina.
- Passagem de células-tronco pluripotentes humanas
- Antes de passar hPSCs, verifique sua morfologia sob o microscópio. Colônias normalmente têm uma morfologia em forma de cúpula com bordas brilhantes e definidas. Se as colônias individuais apresentarem uma morfologia lisonjeira ou se colônias diferenciadas começarem a emergir, siga as instruções do passo 1.2.8.
- Aspire as células médias e lave com PBS uma vez. Adicione 500 μL de solução de descolamento celular por poço de uma placa de 6 poços.
- Incubar as células por 5 min a 37 °C. Use uma pipeta P1000 e pipeta as células várias vezes para dissociar as colônias em células únicas.
- Colete as células e transfira-as para um tubo de 15 mL contendo tampão de lavagem (1 mL por poço de uma placa de 6 poços). Gire as células a 200 x g por 4 min.
- Aspire o supernatante e resuspenda a pelota em meio PXGL fresco (suficiente para 1,5 mL por poço). Considere uma relação de divisão de 1:3-1:6 para a passagem rotineira.
NOTA: Após cada 3-4 passagens ou se a qualidade da cultura celular diminuir com base na morfologia celular (por exemplo, o surgimento de colônias planas na população), adicione 10 μM Y-27632 e o extrato de membrana do porão do fator de crescimento (5 μL / bem) ao meio durante as primeiras 24 horas após a passagem. - Antes de replacar os hPSCs, prepare as placas com MEFs frescos aspirando o meio MEF e lavando as células uma vez com PBS. Em seguida, use uma pipeta P1000 para transferir 1,5 mL de suspensão celular hPSCs por poço de uma placa de 6 poços contendo os MEFs.
NOTA: Certifique-se de que a pipetação leva a uma semeadura homogênea das células em toda a área do poço. Isso garantirá o crescimento de colônias com tamanhos homogêneos e sincronia relativa das células. - A cultura hPSCs em condições hipoxilicas a 37 °C em um ambiente umidificado. Após 24 horas, os hPSCs devem ser anexados. Um alto número de células não aderentes (ou flutuantes) refletem um problema de viabilidade ou de apego na passagem.
- Troque o meio com 1,5 mL de meio PXGL por bem diário. Passar hPSCs a cada 3-4 dias ou usá-los para experimentos de formação de blastoid.
NOTA: Após descongelar hPSCs, passe-os por um mínimo de três passagens antes de iniciar um experimento blastoid.
2. Formação de Blastoids
- Formação de agregados ingênuos do PSC
- Prepare e pré-aqueça a mídia PXGL, mídia basal N2B27, buffer de lavagem, PBS e mídia de agregação antes de iniciar o experimento. Exclua os MEFs da suspensão dos hPSCs para formar blastoids seguindo as etapas descritas abaixo.
- Para exclusão de MEF, prepare uma placa revestida de gelatina adicionando 1 mL de gelatina de 0,1% no poço de uma placa de 6 poços e incubando a 37 °C por 30-90 min.
- Para colher as células, aspire o meio e lave as células com 1 mL de PBS.
- Adicione 500 μL de solução de descolamento celular (por poço de uma placa de 6 poços) e incubar a 37 °C por 5 min.
- Verifique as células sob o microscópio para seguir a dissociação das colônias em células únicas (alguns aglomerados multicelulares podem ser dissociados mais tarde por tubulação suave).
- Diluir a solução de descolamento celular com 1 mL de tampão de lavagem. Recolher as células da placa tubo suavemente de 5 a 10 vezes. Transfira a suspensão da célula para um tubo de 15 mL. Gire as células a 200 x g por 4 min.
- Aspire o supernasciente, resuspenque as células em 1,5 mL de meio PXGL (por poço de uma placa de 6 poços) e semente as células nas placas revestidas de gelatina para exclusão de MEF e incubar a 37 °C por 60-90 min.
- Uma vez que as células ingênuas sejam semeadas para exclusão de MEF, remova o PBS de microwells e equilibre os poços com 200 μL de meio basal N2B27 (por 1 chip de microwell) e incubar por 60 min a 37 °C.
- Colete o supernante contendo as células ingênuas não-sectadas, transfira-o para um tubo de 15 mL e gire as células a 200 x g por 4 min.
- Aspire a mídia e resuspense as células em 1 mL de mídia basal N2B27. Conte as células usando slides de contagem de células. Gire as células a 200 x g por 4 min.
- Aspire o meio e adicione uma quantidade apropriada de 10 μM Y-27632 contida em mídia N2B27 para obter uma densidade celular de 30.000 células por 50 μL.
NOTA: O número ideal de célula de semeadura inicial pode variar entre as diferentes linhas celulares. Por exemplo, para semear 50-60 células/microwell, 30.000 células (incluindo o excedente considerando que algumas células caem fora do poço) são semeadas em 1 poço de 96 placas de poço que contém 430 microwells. Um número de célula inicial inadequado pode resultar na pequena formação agregada sem cavidade ou formação de estrutura cavitada que atinge mais de 250 μm. - Aspire o meio a partir de matrizes de microwell equilibradas e adicione 25 μL de mídia N2B27 com 10 μM Y-27632. Adicione 50 μL de suspensão celular e incubar a 37 °C por 15 min (até que as células caiam no fundo do poço). Em seguida, adicione 125 μL de N2B27 médio suplementado com 10 μM Y-27632.
- Desenvolvimento de blastóides
- Dentro de 24h, podem ser observados agregados de hPSCs ingênuos (dia 0) no chip de microwell. Para iniciar a formação blastóide, prepare o meio PALLY e siga os passos descritos abaixo.
- Pré-aqueça o meio PALLY a 37 °C por 30 min.
NOTA: 1-Oleoyl Ácido lisofosfóctiico (LPA) e 10 μM Y-27632 precisam ser adicionados pouco antes do uso. A concentração ideal de LPA varia entre 0,5-5 μM. Isso deve ser titulado para linhas hPSC individuais usadas para blastoids. - Aspire o meio de agregação. Adicione 200 μL de meio PALLY pré-aquecido aos microwells. Coloque a placa de cultura celular de volta em uma incubadora hipoxica a 37 °C. Repita a mudança de mídia no primeiro dia.
- No dia 2, remova o meio PALLY e adicione 200 μL de n2B27 médio suplementado com LPA e 10 μM Y-27632.
NOTA: No segundo dia, a maioria dos agregados continua a crescer. No entanto, alguns agregados formam pequenas cavidades. A cultura continuamente blastóides em PALLY até o dia 4 ou no meio da cultura in vitro (IVC1) a partir do dia 2. No entanto, após esta mudança de mídia melhora a formação de PrE em blastoids maduros. - Repita a mudança de mídia no dia 3. A formação completa de blastoid ocorre até o dia 4.
NOTA: Os blastóides são considerados totalmente desenvolvidos quando foram submetidos à morfogênese completa com base na morfometria do dia 7 blastocistos humanos (por exemplo, uma faixa de diâmetro de 150 -250 μm; um aglomerado interno cercado por um epitélio de células semelhantes ao trophectoderm) e formaram células semelhantes ao trophectoderm polar (NR2F2+/CDX2-) e células semelhantes a PrE (GATA4+). Isso pode ser avaliado usando a coloração da imunofluorescência ou a triagem celular ativada por fluorescência (FACS).
3. Formação de blastóides em microplacões de 96 poços de ultra baixo fixação
- Prepare a suspensão celular ingênua de hPSCs para formação de blastoid seguindo as etapas descritas acima de 2.1.1 - 2.1.11.
- Aspire o meio e adicione uma quantidade apropriada de mídia N2B27 contendo 10 μM Y-27632 para obter a densidade celular de 70 células por 100 μL do meio.
NOTA: O número ideal de célula de semeadura inicial pode variar entre diferentes linhas celulares. Por exemplo, 70 células/bem pode ser o número de celular ideal para a maioria das linhas celulares. - Centrifugar a placa a 200 x g por 2 min à temperatura ambiente para agrupar células na parte inferior dos poços.
- Incubar a placa em uma incubadora a 37 °C em condições de cultura hipóxica. Dentro de 24 h, podem ser observados agregados de hPSCs ingênuos (dia 0) nos poços.
- Prepare o meio 2x PALLY. Adicione 100 μL de meio PALLY 2x pré-armado aos poços.
- Coloque a placa de cultura celular de volta em uma incubadora hipoxica a 37 °C. Após 24 horas, aspire metade da mídia (100 μL) e substitua-a por 100 μL de meio PALLY pré-armado. Repita o passo até o dia 4. Certifique-se de não aspirar os agregados.
NOTA: No segundo dia, a maioria dos agregados continua a crescer. No entanto, alguns agregados têm pequenas cavidades cheias de líquido. No dia 4, a maioria das estruturas cavitadas sofrem morfogênese completa para formar estruturas semelhantes a blastocistos.
4. Formação de troforos
- Para a formação da troforosfera, siga o protocolo de formação de blastóides da etapa 2.1.1 (exclusão mef) até a etapa 2.1.12 (última etapa do protocolo de semeadura).
- Uma vez que os agregados de hPSCs ingênuos tenham se formado após 24 horas, troque o meio de agregação com PALY (sem LIF) complementado com 3 μM SC-144 para a formação de troforoses representando trophectoderme precoce e PALLY complementado com 2 μM XMU-MP-1 para a formação de troforosferas representando trophiftoderm maduro.
- Atualize o meio diariamente. A formação completa da troforosfera ocorre até o dia 4.
5. Análise do estado das células blastóides e seu estágio refletido usando scRNAseq
- Para pegar blastóides e realizar a dissociação, aqueça a incubadora de agitação a 37 °C e coloque-a a 100 rpm.
- Colete blastoids da placa inicial de 96 poços e transfira-os para vários poços de uma placa de 96 poços usando um tubo bucal equipado com um capilar de vidro.
NOTA: Os blastóides (> 70%) devem ser selecionados com base nos critérios morfométricos (tamanho = 150-250 μm com um cluster interno único) a fim de evitar contaminação com estruturas não-blastóides (< 30%). - Lave uma vez com 200 μL de PBS usando um P200 visualizando sob um estereómico. Transfira para um poço contendo 50 μL de colagem e incubar por 30 minutos na incubadora de agitação.
- Transfira os blastóides para um poço com 100 μL de 10x trypsin-EDTA e misture bem. Incubar por 20 min na incubadora de agitação.
- Dissociar os blastóides em célula única usando pipeta P200. Transfira as células para um tubo de 15 mL com tampão FACS (1% FBS em PBS).
- Para capturar proporções específicas dos análogos das três linhagens, colora os análogos TE e PrE com anticorpos TROP2 e PDGFRa, respectivamente.
NOTA: O número de células PrE em blastocistos humanos é menor em comparação com os blastocistos do rato, o que pode refletir defeitos de desenvolvimento de blastocistos formados através de fertilização in vitro (FIV) ou uma diferença de espécie. Nos blastóides, os análogos pre são menos abundantes do que os analógicos TE e EPI e representam 7,4% das células mediante a contagem de células GATA4+ por imagens de imunofluorescência. Além disso, o processo de dissociação pode induzir vieses nas proporções dos diferentes tipos de células como análogos de PrE para representar 1%-2% das células após dissociação blastóide, marcação PDGFRa e, análise FACS. - Células de tipo FACS dos três analógicos de linhagem em 384 placas contendo um tampão de lise para análise smart-seq2. Exclua células mortas marcadas pela coloração DAPI (realizada de acordo com as instruções do fabricante).
- Para avaliar os estados celulares (tipo celular e estágio de desenvolvimento), compare os dados transcriptômicos do blastoid com os controles apropriados.
6. Cultura estendida para avaliar a progressão do desenvolvimento de blastóides
- Cultura blastóides humanos em placas revestidas de matriz de membrana do porão (fundo de vidro).
- Cubra a placa com matriz de membrana do porão.
- Inspecione visualmente os blastóides para avaliar e registrar morfologia.
NOTA: Apenas blastoids que exibem a clássica morfologia blastocisto de bola oca com ICM compacto têm o potencial de crescer e desenvolver ainda mais. - Adicione 100 μL de CMRL médio-1 por um poço da placa de 96 poços, e coloque a placa na incubadora pelo menos 2 h antes que os blastóides transfiram para o equilíbrio.
- Usando um estereótipo, identifique visualmente os blastóides com boa morfologia, transfira os blastóides selecionados em um poço de placa de 96 poços contendo 100 μl de CMRL médio-1 para remover os traços de mídia blastoid.
- Transfira os blastóides para o poço contendo cmrl media-1 pré-equilibrado. Coloque a placa na incubadora e incubar a 37°C durante a noite.
NOTA: Até 5 blastoids podem ser cultivados em um poço de 96 placas de poço. Ter muitos blastóides em um único poço pode levar à formação de agregados de múltiplos blastóides. - No dia seguinte, inspecione visualmente os blastóides sob um microscópio. Se os blastóides estiverem ligados, adicione 100 μL de cmrl media-1 pré-equilibrado suplementado com matriz de membrana de 5% do porão. Coloque a placa na incubadora e incubar a 37 °C durante a noite.
- No dia seguinte, monitore os blastóides sob um microscópio. Remova metade da mídia (100 μL) e substitua-a por 100 μL de cmrl media-2 pré-equilibrada complementada com matriz de membrana de 5% do porão.
- Nos dias seguintes, substitua metade da mídia (100 μL) por mídia CMRL pré-equilibrada-3 complementada com matriz de membrana de 5% do porão. Coloque a placa na incubadora e incubar a 37 °C durante a noite. Repita todos os dias por até 4-6 dias de cultura in vitro .
NOTA: Temos blastóides cultivados por até 6 dias em condições de cultura estendida que corresponde ao tempo equivalente ao dia 13 de embriões humanos in vitro cultivados.
7. Blastóides imunossustaining
- Aspire o meio. Lave amostras 3x com PBS por 5 min.
- Adicione 200 μL de paraformaldeído frio de 4% (PFA) em PBS e fixe amostras por 30 minutos a temperatura ambiente. Remova a solução PFA e lave amostras 3x com PBS por 10 minutos.
NOTA: Se os blastóides foram cultivados em chips de microwell, transfira os blastóides do chip para placas de fundo U de 96 poços para as seguintes etapas. - Permeabilize e bloqueie os blastóides em 100 μL de solução de bloqueio por poço (PBS contendo 0,3% Triton-X 100 e 10% soro de burro normal) por 60 min.
NOTA: Dependendo das espécies hospedeiras dos anticorpos, adapte o soro em conformidade. - Remova a solução de bloqueio. Adicione 100 anticorpos primários diluídos em solução de bloqueio fresco e incubar amostras durante a noite a 4°C.
NOTA: As concentrações dos anticorpos primários devem ser determinadas com base nas instruções do fabricante. - Lave amostras 3x com 0,1% Triton-X 100 em PBS (solução de lavagem) por 10 min. Adicione 100 μL de anticorpos secundários na solução de bloqueio juntamente com 20 μg/mL Hoechst mancha nuclear e incubar amostras por 1 h à temperatura ambiente. Proteja amostras contra a luz.
NOTA: As concentrações de anticorpos secundários devem ser determinadas com base nas instruções do fabricante. - Lave amostras 3x com solução de lavagem por 10 minutos. Para imagens, transfira as amostras para o fundo do vidro μ-slide em PBS.
NOTA: O meio de montagem deve ser selecionado com base no objetivo utilizado para a imagem. Por exemplo, 80% de glicerol na PBS é possível usar para a montagem das amostras enquanto usa objetivos de óleo.
Resultados
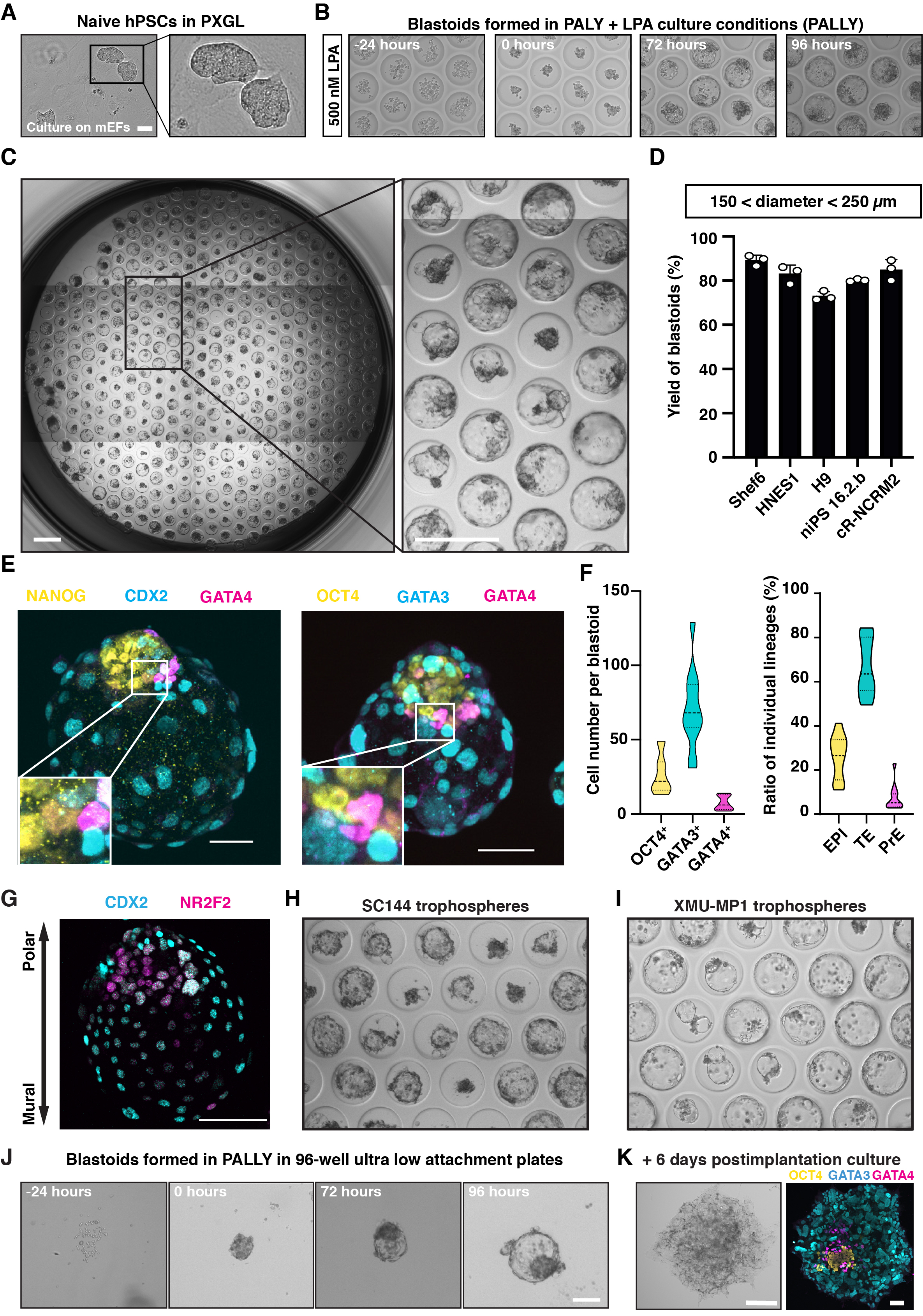
Tipicamente, hPSCs ingênuos cultivados em PXGL (Figura 2A) são estruturas agregadas e cavitadas que emergem entre 48 e 72 h após a indução pally e atingem um diâmetro de 150-250 μm dentro de 96 h (Figura 2B). Usando números de células de semeadura ideais (1) (2) duração da agregação pré-cultura com N2B27 (0 a 24 h), (3) concentração de componentes químicos individuais (especialmente LPA) e (4) duração do tratamento PALLY, a eficiência de indução atinge 70%-80% conforme definido com base em parâmetros morfométricos (tamanho total de 150-250 μm, cavidade regular única, cavidade celular única interna; Figura 2C,D) e a presença das três linhagens. Um estado celular inicial subótimo e/ou condições de indução resultarão em formação menos eficiente ou não de blastoid. Para garantir a máxima eficiência e apenas formar análogos pré-implantação, é crucial usar uma cultura de alta qualidade de hPSCs PXGL ingênuos. Isso pode ser avaliado medindo pelo FACS o percentual de células positivas para os marcadores superficiais SUSD2 (estado ingênuo) e CD24 (estado preparado). Marcadores de superfície adicionais específicos para as linhagens extraembryônicas fora do alvo (por exemplo, amnion, mesoderme extraembryônico) também seriam úteis, mas, pelo que sabemos, não estão disponíveis no momento. Se a eficiência de formação obtida for menor do que os resultados relatados, é importante verificar cuidadosamente todos os componentes do meio blastóide, especialmente lPA que é reconstituído em PBS e que, como ligante GPCR, pode ser mais instável em comparação com moléculas sintéticas reconstituídas em DMSO. Na maioria dos casos, mesmo que o rendimento não seja máximo, as estruturas cavitadas ainda são compostas das três linhagens blastocisto. O surgimento de três linhagens blastocisto e formação de eixo embrionário-abembriônico pode ser confirmado pela mancha de imunofluorescência de marcadores (EPI: NANOG, OCT4, TE: GATA3, Polar-TE NR2F2, Mural-TE: CDX2, PrE: GATA4; Figura 2E,G). As troforosferas, que são compostas apenas de TE, ajudam a dissecar ainda mais o papel da comunicação intercelular. As troforos podem formar-se a 50%-60 % de eficiência dentro de 96 h de indução (Figura 2H,I). A formação de blastoid pode ser realizada não apenas em matrizes de microwell caseiros, mas também em acessórios ultra-baixos disponíveis comercialmente 96 placas de bem com otimização de condições de indução (ver Protocolo e Figura 2J). Os blastóides também têm a capacidade de se desenvolver ainda mais por mais 6 dias, o que equivale ao tempo equivalente ao 13º dia de embrião, com protocolo de diferenciação in vitro (Figura 2K).
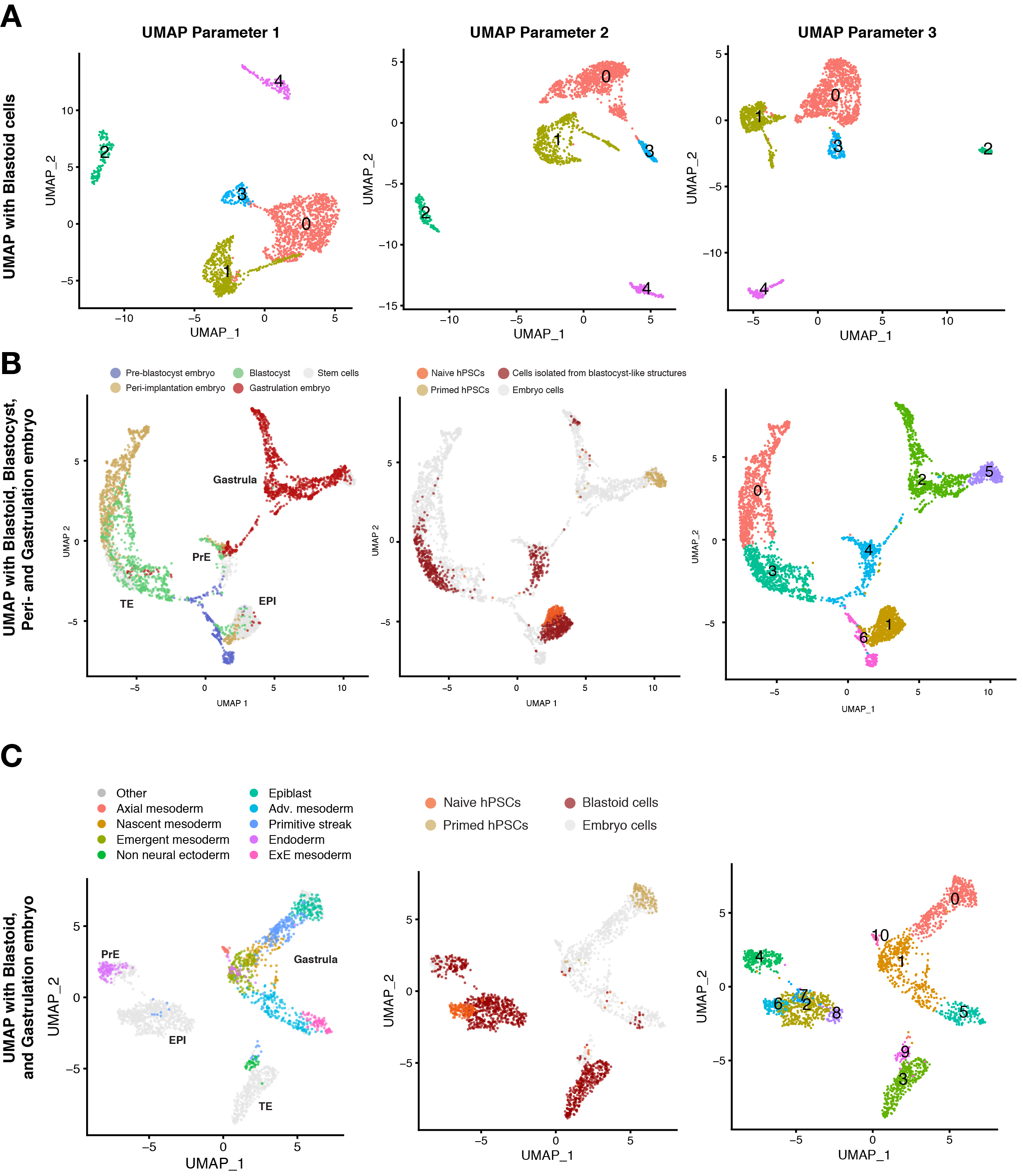
Para caracterizar ainda mais o estado celular das células blastoid, deve-se utilizar a tecnologia de sequenciamento de RNA unicelular. A UMAP é comumente aplicada para visualizar uma distribuição de estados celulares e a análise de agrupamento não supervisionada é realizada nele para avaliar a proximidade de estados células individuais. Diferentes parâmetros na análise de dados unicelulares podem afetar a forma como as células são exibidas nas UMAPs, levando assim a aglomerados com diferentes posições e formas espaciais e relativas (Figura 3A). No entanto, nesta análise, as células exibem perfis de agrupamento notavelmente distintas, independentemente dos parâmetros utilizados para realizar o clustering e a visualização dos dados, o que permite distinguir com alta confiança as três linhagens blastocistos. Usamos células de embriões colhidos em diferentes estágios de desenvolvimento como referência. A fusão desses conjuntos de dados mostra que a maioria do análogo do trophectoderm de blastoid agrupado com trophectoderm pré-implantação, mas não com trophoblastos pós-implantação (Figura 3B). Esses resultados também foram confirmados por um consórcio independente10.
Quando os embriões gastrulating do estágio 7 (CS7) do Carnegie são introduzidos no mapa de referência, uma pequena população de células blastoid (3%) agrupadas com as linhagens de mesoderme e amnion desses embriões (Figura 3C). Quando células semelhantes a amnion são introduzidas no mapa de referência, uma pequena população de células blastoid (< 2%) agrupadas com células semelhantes a amônio.
No geral, apenas as estruturas que compreendem uma única cavidade regular, um único aglomerado de células internas, um tamanho global que varia de 150-250 μm, compreendendo análogos transcriômicos das três linhagens blastocisto, e em grande parte desprovidos de outras linhagens (por exemplo, amnion, mesoderme, mesoderme extraembryônico) são considerados como blastoids humanos.

Figura 1: Quatro recursos e duas abordagens para gerar blastoids de alta fidelidade. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Blastoid e troforos derivados de agregados de Ingïve hPSC. (A) Imagens de contraste de fase que mostram hPSCs ingênuos cultivados em meio PXGL co-cultivados com MEF irradiado. Barra de escala: 50 μm. (B) Imagens de contraste de fase mostrando a mudança morfológica de hPSCs ingênuos agregam cultivados em uma matriz de microwell de hidrogel não aderente com 500 nM LPA (meio PALLY). Barra de escala: 200 μm. (C) Blastóides humanos formados em uma matriz de microwell após 96 h. Barras de escala: 400 μm. (D) Quantificação da porcentagem de microwells contendo um blastóide humano induzido pela condição de cultura PALLY com concentração de LPA otimizada de diferentes linhas ingênuas hPSC (n= 3 matrizes de microwell). (E) Coloração de imunofluorescência de blastóides humanos com marcadores epiblasto (EPI) (amarelo) NANOG e OCT4; os marcadores TE (ciano) CDX2 e GATA3; e o marcador primitivo de endoderme (magenta) SOX17 e GATA4. Barra de escala: 100 μm. (H-I) Quantificação do número celular (esquerda) e porcentagem de células (direita) pertencentes a cada linhagem em blastóide (96 h) com base na coloração da imunofluorescência de OCT4, GATA3 e GATA4. (G) Mancha de imunofluorescência de blastóides humanos para CDX2 (ciano) e NR2F2 (magenta). (F) Imagens de contraste de fase de troforferas de estágio inicial e tardio na matriz de microwell induzidas pela adição de 3 μM SC144 (H) ou 2 μM XMU-MP-1 (I), respectivamente. (J) Imagens de contraste de fase mostrando a mudança morfológica de hPSCs ingênuos agregam cultivados em placa de fixação ultra-baixa 96 com 500 nM LPA (meio PALLY). (K) Imagem de contraste de fase (esquerda) e coloração de imunofluorescência (direita) para OCT4 (amarelo), GATA3 (ciano) e GATA4 (magenta) em blastoid cultivado em condição de cultura pós-implantação por 6 dias. Barra de escala: 100 μm. Este número é adaptado a partir de 6,10. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Caracterização da composição de blastóides por sequenciamento de célula única. (A) Análise de clustering não supervisionada com diferentes parâmetros em UMP do transcriptome de células únicas derivadas dos diferentes pontos de tempo do blastóide (24 h, 60 h, 96 h), hPSCs ingênuos, hPSCs primed e hTSC (representam o citotrofoblasto pós-implantação). (B) UMAP do transcriptome de células derivadas de blastoid (96 h), hPSCs ingênuos e hPSCs primed integrados com conjuntos de dados publicados de embriões humanos de pré-implantação, peri-implantação (blastocistos cultivados in vitro ) e gastrulação (estágio Carnegie 7, ou seja, entre estágios E16-19). As células individuais são coloridas com base em sua origem em embriões humanos (esquerda), células derivadas de blastóides ou células-tronco (meio), e o resultado de análise de agrupamento não supervisionada (à direita). (C) UMAPs do transcriptome de células derivadas de blastóides, hPSCs ingênuos, hPSCs preparados e integrados com conjunto de dados publicados a partir de gastrulação (Estágio Carnegie 7, ou seja, entre E16-19) estágio embrião. As células individuais são coloridas com base em sua origem em embriões humanos (esquerda), células derivadas de blastóides ou células-tronco (meio), e o resultado de análise de agrupamento não supervisionada (à direita). Este número é adaptado a partir de6. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Tabela Suplementar 1: Toda a composição de mídia utilizada neste estudo. Clique aqui para baixar esta Tabela.
Discussão
No presente estudo, mostramos, passo a passo, como estabelecer blastóides humanos com alta eficiência usando um protocolo simples e robusto. Após a agregação de hPSCs PXGL ingênuos e sua inibição tripla, os blastóides formam-se eficientemente (> 70%) e geram sequencialmente os 3 análogos blastocistos dentro de 4 dias. Limitações na eficiência e qualidade dos blastóides (por exemplo, presença de células fora do alvo) podem ocorrer se o estado inicial for sub-ideal. Note-se que medimos que os HPSCs PXGL contêm cerca de 5% das células que refletem os estágios pós-implantação. Essas células podem limitar a formação de blastóides de alta qualidade. Além do estado inicial ingênuo do PXGL que reflete o epiblasto do blastocisto, outro fator crucial é o meio usado para a formação de blastoid. A fim de formar rapidamente células semelhantes a blastocistos e impedir a formação de células fora do alvo, semelhantes à pós-implantação, propomos que a inibição de vias triplas (Hipopótamo, ERK, TGF-β) seja essencial. Enquanto diferentes linhas celulares dão diferentes rendimentos de blastoid em inibição de ERK/TGF-β (geralmente em torno de 10%-20%), a exposição ao LPA resulta na formação de um rendimento de blastoid igualmente alto em todas as linhas celulares, ao mesmo tempo em que usa rigorosos critérios de especificação morfométrica e de linhagem. LPA possivelmente age na inibição da via hipopótamo, que desempenha um papel crítico na primeira segregação de linhagem entre linhagens de epiblasto e trophectoderme em camundongos ehumanos 8,51. A melhoria significativa da eficiência blastoid por LPA sugere que os mecanismos de especificação de células internas mediados pelo hipopótamo em jogo no blastocisto são cooptados durante a formação de blastoid. Uma limitação atual reside no fato de que, devido a uma sub-idealidade dos protocolos usados para cultivar blastocisto humano ou blastóides no dia 7-13 (após a formação de blastocisto/blastoid), não somos capazes de avaliar até que ponto podemos modelar adequadamente o desenvolvimento pós-implantação.
Analisar o estado transcriômico das células blastóides pode ser facilmente alcançado usando scRNAseq, mapas de referência adequados e métodos bioinforáticos. Anteriormente, a análise transcriômica mostrou que os hPSCs cultivados em PXGL são mais semelhantes ao epiblasto blastocisto em comparação com o estado primed. Limitações na análise dos dados podem ocorrer se o mapa de referência for composto apenas por células em estágio de blastocisto. O mapa de referência deve incluir células originárias de embriões pós-implantação, a fim de avaliar a presença de células potencialmente fora do alvo. No futuro, a fim de marcar células blastoid, um mapa de referência incluindo todos os tecidos do conceito humano pré e pós-implantação seria extremamente valioso. Além disso, mapas de referência celular único multi-omics, por exemplo, incluindo transcriptome, acessibilidade de cromatina e metilação de DNA, ajudariam ainda mais. Finalmente, métodos bioinforáticos padronizados para avaliar quantitativamente as semelhanças entre células de modelos de embriões e conceitos de referência, e identificar positivamente células fora do alvo ajudariam ainda mais a analisar e comparar resultados imparcialmente.
Ao todo, os blastóides formados pela inibição tripla das vias Hipopótamo, TGF-β e ERK possuem as quatro características de 1) morfogênese altamente eficiente, 2) sequência correta de especificação de linhagem, 3) alta pureza de células semelhantes a blastocistos no nível do transcriptome, 4) capacidade para modelar o desenvolvimento da peri-implantação. Essas características dos blastóides facilitarão a construção de hipóteses sobre o desenvolvimento e implantação de blastocistos, no entanto, eles não recapitulam estágios anteriores de desenvolvimento embrionário. Em contraste com a acessibilidade limitada e versatilidade do blastocisto humano, os blastóides são favoráveis às telas genéticas e medicamentosas para as investigações funcionais de desenvolvimento e implantação de blastocistos. No futuro, esse conhecimento básico poderia contribuir para melhorar a formulação da mídia de FIV, desenvolver contraceptivos pós-fertilização e gerenciar melhor a gravidez precoce.
Divulgações
O Instituto de Biotecnologia Molecular da Academia Austríaca de Ciências registrou um pedido de patente EP21151455.9 descrevendo os protocolos para formação de blastóides humanos e o ensaio de interação blastoid-endométrio. HK, AJ, HHK e NR são os inventores desta patente. Todos os outros autores não declaram interesses concorrentes.
Agradecimentos
Este projeto recebeu financiamento do Conselho Europeu de Pesquisa (ERC) no âmbito do programa de pesquisa e inovação Horizon 2020 da União Europeia (contrato de subvenção da ERC-Co nº 101002317 'BLASTOID: uma plataforma de descoberta para embriogênese humana precoce'). H.H.K. é apoiado pelo Fundo Austríaco de Ciência (FWF), Programa Lise Meitner M3131-B. Agradecemos a Yasuhiro Takashima por compartilhar as linhas celulares H9 e H9-GFP, e Austin Smith, Peter Andrews e Ge Guo por compartilharem as linhas celulares HNES1, Shef6, niPSC 16.2b e cR-NCRM2. Agradecemos a Hossein Baharvand por compartilhar os organoides endometrial. Agradecemos a Joshua M. Brickman por compartilhar o RNA isolado de células diferenciadas pre e células nEND. Agradecemos a Shankar Srinivas por compartilhar os dados de sequenciamento de RNA unicelular do embrião peri-gastrulation. Agradecemos a Aleksand Bykov e Luisa Cochella pela assistência técnica para a preparação da biblioteca SMARTSeq2. Agradecemos às instalações ngs, biopóticas e células-tronco da IMBA por assistência crítica.
Materiais
| Name | Company | Catalog Number | Comments |
| Neurobasal media | in house | ||
| DMEM/F12 | in house | ||
| 100X N2 supplemen | Gibco | 17502048 | |
| 50X B27 supplement | Gibco | 17504044 | |
| 100X Glutamax | Gibco | 35050038 | |
| 100 mM Sodium Pyruvate | Gibco | 11360039 | |
| MEM-Non-essential amino acids | Gibco | 11140050 | |
| 1 M Hepes | in house | ||
| 50 mM 2-Mercaptoethanol | Thermofisher | 31350010 | |
| 100X Penicillin-Streptomycin | Sigma-Aldrich | P0781 | |
| Bovine Serum Albumin solution | Sigma-Aldrich | A7979 | |
| PD0325901 | Medchem express | HY-10254 | |
| XAV-939 | Medchem express | HY-15147 | |
| Gö 6983 | Medchem express | HY-13689 | |
| Human recombinant Leukemia Inhibitory Factor | in house | ||
| A83-01 | Medchem express | HY-10432 | |
| 1-Oleoyl Lysophosphatidic acid (LPA) | Peprotech | 2256236 | |
| Y-27632 | Medchem express | HY-10583 | |
| CMRL medium | Gibco | 21530027 | |
| Fetal Bovine Serum (FBS) | Sigma-Aldrich | F7524 | |
| KnockOut Serum Replacement (KSR) | Thermofisher | 10-828-028 | |
| Accutase | Biozym | B423201 | cell detachment solution |
| Geltrex | Thermofisher | A1413302 | growth factor basement membrane extract |
| TROP2 antibody | R&D systems | MAB650 | |
| PDGFRα antibody | R&D systems | AF307 | |
| SC-144 | Axon | 2324 | |
| XMU-MP-1 | Med Chem Express | HY-100526 | |
| Matrigel | basement membrane matrix | ||
| Countess cell counting chamber slides | Thermo fisher | cell counting slides | |
| DAPI Staining Solution | Miltenyi Biotec | 130-111-570 |
Referências
- Rivron, N., et al. Debate ethics of embryo models from stem cells. Nature. 564 (7735), 183-185 (2018).
- Hyun, I., Munsie, M., Pera, M. F., Rivron, N. C., Rossant, J. Toward Guidelines for Research on Human Embryo Models Formed from Stem Cells. Stem Cell Reports. 14 (2), 169-174 (2020).
- Clark, A. T., et al. Human embryo research, stem cell-derived embryo models and in vitro gametogenesis: Considerations leading to the revised ISSCR guidelines. Stem Cell Reports. 16 (6), 1416-1424 (2021).
- Lovell-Badge, R., et al. ISSCR Guidelines for Stem Cell Research and Clinical Translation: The 2021 update. Stem Cell Reports. 16 (6), 1398-1408 (2021).
- Rivron, N. C., et al. Blastocyst-like structures generated solely from stem cells. Nature. 557 (7703), 106-111 (2018).
- Kagawa, H., et al. Human blastoids model blastocyst development and implantation. Nature. , 04267-04268 (2021).
- Meistermann, D., et al. Integrated pseudotime analysis of human pre-implantation embryo single-cell transcriptomes reveals the dynamics of lineage specification. Cell Stem Cell. 28 (9), 1625-1640 (2021).
- Gerri, C., et al. Initiation of a conserved trophectoderm program in human, cow and mouse embryos. Nature. 587 (7834), 443-447 (2020).
- Gerri, C., Menchero, S., Mahadevaiah, S. K., Turner, J. M. A., Niakan, K. K. Human Embryogenesis: A Comparative Perspective. Annual Review of Cell and Developmental Biology. 36, 411-440 (2020).
- Zhao, C., et al. Reprogrammed iBlastoids contain amnion-like cells but not trophectoderm. bioRxiv. , 2021.05.07.442980 (2021).
- Zijlmans, D. W. Integrated multi-omics reveal polycomb repressive complex 2 restricts human trophoblast induction. Nat. Cell Biol. 24, 858-871 (2022).
- Nichols, J., Smith, A. Naive and primed pluripotent states. Cell Stem Cell. 4 (6), 487-492 (2009).
- Guo, G., et al. Epigenetic resetting of human pluripotency. Development. 144 (15), 2748-2763 (2017).
- Takashima, Y., et al. Resetting transcription factor control circuitry toward ground-state pluripotency in human. Cell. 158 (6), 1254-1269 (2014).
- Thomson, J. A. Embryonic stem cell lines derived from human blastocysts. Science. 282 (5391), 1145-1147 (1998).
- Yang, Y., et al. Derivation of Pluripotent Stem Cells with In Vivo Embryonic and Extraembryonic Potency. Cell. 169 (2), 243-257 (2017).
- Stirparo, G. G., et al. Integrated analysis of single-cell embryo data yields a unified transcriptome signature for the human pre-implantation epiblast. Development. 145 (3), 158501 (2018).
- Castel, G., et al. Induction of Human Trophoblast Stem Cells from Somatic Cells and Pluripotent Stem Cells. Cell Reports. 33 (8), 108419 (2020).
- Posfai, E., et al. Evaluating totipotency using criteria of increasing stringency. Nature Cell Biology. 23 (1), 49-60 (2021).
- Nakamura, T., et al. A developmental coordinate of pluripotency among mice, monkeys and humans. Nature. 537 (7618), 57-62 (2016).
- Theunissen, T. W., et al. Molecular Criteria for Defining the Naive Human Pluripotent State. Cell Stem Cell. 19 (4), 502-515 (2016).
- Guo, G., et al. Naive Pluripotent Stem Cells Derived Directly from Isolated Cells of the Human Inner Cell Mass. Stem Cell Reports. 6 (4), 437-446 (2016).
- Rossant, J. Genetic Control of Early Cell Lineages in the Mammalian Embryo. Annual Review of Genetics. 52, 185-201 (2018).
- De Paepe, C., et al. Human trophectoderm cells are not yet committed. Human reproduction. 28 (3), 740-749 (2013).
- Amita, M., et al. Complete and unidirectional conversion of human embryonic stem cells to trophoblast by BMP4. Proceedings of the National Academy of Sciences of the United States of America. 110 (13), 1212-1221 (2013).
- Io, S., et al. Capturing human trophoblast development with naive pluripotent stem cells in vitro. Cell Stem Cell. 28 (6), 1023-1039 (2021).
- Guo, G., et al. Human naive epiblast cells possess unrestricted lineage potential. Cell Stem Cell. 28 (6), 1040-1056 (2021).
- Liu, X., et al. Modelling human blastocysts by reprogramming fibroblasts into iBlastoids. Nature. 591 (7851), 627-632 (2021).
- Hirate, Y., et al. Polarity-dependent distribution of angiomotin localizes Hippo signaling in preimplantation embryos. Current biology: CB. 23 (13), 1181-1194 (2013).
- Cockburn, K., Biechele, S., Garner, J., Rossant, J. The Hippo pathway member Nf2 is required for inner cell mass specification. Current Biology: CB. 23 (13), 1195-1201 (2013).
- Xu, R. H., et al. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nature Biotechnology. 20 (12), 1261-1264 (2002).
- Krendl, C., et al. GATA2/3-TFAP2A/C transcription factor network couples human pluripotent stem cell differentiation to trophectoderm with repression of pluripotency. Proceedings of the National Academy of Sciences of the United States of America. 114 (45), 9579-9588 (2017).
- Xu, R. H., et al. Basic FGF and suppression of BMP signaling sustain undifferentiated proliferation of human ES cells. Nature Methods. 2 (3), 185-190 (2005).
- Horii, M., Bui, T., Touma, O., Cho, H. Y., Parast, M. M. An Improved Two-Step Protocol for Trophoblast Differentiation of Human Pluripotent Stem Cells. Current Protocols in Stem Cell Biology. 50 (1), 96 (2019).
- Okae, H., et al. Derivation of Human Trophoblast Stem Cells. Cell Stem Cell. 22 (1), 50-63 (2018).
- Yamanaka, Y., Lanner, F., Rossant, J. FGF signal-dependent segregation of primitive endoderm and epiblast in the mouse blastocyst. Development. 137 (5), 715-724 (2010).
- Kuijk, E. W., et al. The roles of FGF and MAP kinase signaling in the segregation of the epiblast and hypoblast cell lineages in bovine and human embryos. Development. 139 (5), 871-882 (2012).
- Roode, M., et al. Human hypoblast formation is not dependent on FGF signalling. Developmental Biology. 361 (2), 358-363 (2012).
- Linneberg-Agerholm, M., et al. Naïve human pluripotent stem cells respond to Wnt, Nodal and LIF signalling to produce expandable naïve extra-embryonic endoderm. Development. 146 (24), (2019).
- Yu, L., et al. Blastocyst-like structures generated from human pluripotent stem cells. Nature. 591 (7851), 620-626 (2021).
- Yanagida, A., et al. Naive stem cell blastocyst model captures human embryo lineage segregation. Cell Stem Cell. 28 (6), 1016-1022 (2021).
- Zappia, L., Oshlack, A. Clustering trees: a visualization for evaluating clusterings at multiple resolutions. GigaScience. 7 (7), (2018).
- Subramanian, A., et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proceedings of the National Academy of Sciences of the United States of America. 102 (43), 15545-15550 (2005).
- Messmer, T., et al. Transcriptional Heterogeneity in Naive and Primed Human Pluripotent Stem Cells at Single-Cell Resolution. Cell Reports. 26 (4), 815-824 (2019).
- Zhou, F., et al. Reconstituting the transcriptome and DNA methylome landscapes of human implantation. Nature. 572 (7771), 660-664 (2019).
- Petropoulos, S., et al. Single-Cell RNA-Seq Reveals Lineage and X Chromosome Dynamics in Human Preimplantation Embryos. Cell. 167 (1), 285 (2016).
- Tyser, R. C. V., et al. A spatially resolved single cell atlas of human gastrulation. bioRxiv. , (2020).
- Turco, M. Y., et al. hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nature Cell Biology. 19 (5), 568-577 (2017).
- Boretto, M., et al. Development of organoids from mouse and human endometrium showing endometrial epithelium physiology and long-term expandability. Development. 144 (10), 1775-1786 (2017).
- Ma, H., et al. In vitro culture of cynomolgus monkey embryos beyond early gastrulation. Science. 366 (6467), (2019).
- Nishioka, N., et al. The Hippo signaling pathway components Lats and Yap pattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass. Developmental Cell. 16 (3), 398-410 (2009).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados