
Method Article
Imagerie time-lapse de la souris Macrophage Chemotaxis
Dans cet article
Résumé
Ici, nous décrivons des méthodes utilisant time-lapse, microscopie phase-contraste à l’image de la souris résident macrophages péritonéal dans un complément chemotactic C5a gradient. Les protocoles peuvent être étendus à d’autres cellules immunitaires.
Résumé
La chimiotaxis est une guidage par les récepteurs des cellules le long d’un gradient chimique, tandis que la chimiokinesis est la stimulation de la motilité cellulaire aléatoire par un produit chimique. La chimiokinesis et la chimiotaxis sont fondamentales pour la mobilisation et le déploiement des cellules immunitaires. Par exemple, les chimiothérapeutiques (cytokines chemotactic) peuvent rapidement recruter des neutrophiles et des monocytes circulants vers des sites extravasculaires de l’inflammation. Les récepteurs chimiotérants appartiennent à la grande famille des récepteurs couplés aux protéines G. Comment la chimiorattractant (c.-à-d., ligand) gradients direct migration cellulaire par l’intermédiaire de G protéine couplée signalage récepteur n’est pas encore entièrement compris. Dans le domaine de l’immunologie, les neutrophiles sont des cellules modèles populaires pour étudier la chimiotaxie in vitro. Ici, nous décrivons un essai en chimiotaxis bidimensionnel en temps réel (2D) adapté aux macrophages résidents de souris, qui ont traditionnellement été plus difficiles à étudier. Les macrophages se déplacent à un rythme lent de 1 m/min sur une surface 2D et sont moins bien adaptés aux essais de migration à source ponctuelle (p. ex., migration vers la pointe d’une micropipette remplie de chimiottractant) que les neutrophiles ou le discoideum de dictyostelium, qui déplacent un ordre de grandeur plus rapidement. Les essais Transwell largement utilisés sont utiles pour étudier l’activité chimioctictique de différentes substances, mais ne fournissent pas d’informations sur la morphologie cellulaire, la vitesse ou la navigation chimotatique. Ici, nous décrivons un essai de chimiotaxis de chimiotaxis de microscopie en accéléré qui permet la quantification de la vitesse cellulaire et de l’efficacité chimiomoctictique et fournit une plate-forme pour délimiter les transducteurs, les voies de signalisation et les effecteurs de la chimiotaxis.
Introduction
Les cellules immunitaires migrent généralement individuellement sur une surface 2D d’une manière amoeboid1,2, qui implique des cycles répétés de saillie de l’avant, l’adhérence cellulaire integrine-négociée, et la rétraction de l’arrière. Une étape préalable est la polarisation cellulaire, dans laquelle les cellules forment les extrémités avant et arrière3. La chimiotaxis commence par la détection de chimioattractants par des récepteurs couplés à la protéine G et d’un réseau complexe de signalisation médié par des protéines heterotrimeric G ancrées par membrane et de petites protéines monomérites G, ainsi que par des facteurs d’échange de nucléotides de guanine à taux phospholipid (GEF)4,5. L’activation des Rho GTPases des sous-familles Cdc42 et Rac induisent des saillies à l’avant6 et les membres de la sous-famille Rho, en particulier RhoA, activent la contraction de l’arrière5,7. Dans un environnement tridimensionnel (3D), les intégraux sont largement redondants pour la migration de leucocytes et RhoA devient plus important pour presser les cellules par des passages étroits8, tandis que l’activation Arp2/3 induite par cdc42 ou Rac reste importante pour la direction chimotatique9,10.
Les cellules immunitaires peuvent faire face à des chimiorattractants différents, en particulier dans les arrangements des dommages de tissu, invasion d’agents pathogènes, et inflammation. Les chimiorattractants endogènes exprimés sur les phagocytes complètent C3a et C5a, sont rapidement générés par l’activation de la cascade de complément, et sont reconnus par les récepteurs C3a et C5a de complément. De même, les cellules nécrotiques recrutent des phagocytes via des récepteurs de peptide formyl, qui reconnaissent les peptides formyl dérivés des mitochondries ainsi que les peptides formyl dérivés de bactéries11. Les cellules immunitaires expriment également des récepteurs couplés à la protéine G pour les chimione, une grande famille de peptides chimioattractants impliqués dans la régulation du trafic de cellules immunitaires pendant l’homéostasie et l’inflammation. Les chemokines sont classés en quatre groupes en fonction de l’espacement des deux premiers résidus de cystéine (C) : C, CC, CXC, et CX3C cytokines, où X est un acide aminé. Ainsi, les cellules immunitaires in vivo doivent répondre de manière appropriée aux signaux spatiaux et temporels très complexes, ce qui rend l’étude de la chimiotaxe une tâche intimidante. Ci-dessous nous fournissons une brève histoire de la chimiotaxis, qui a commencé avec des approches d’imagerie intravitale.
L’étude de la chimiotaxis leucocyte remonte à 188812, lorsque l’ophtalmologiste Theodor Karl Gustav Leber a clairement décrit la migration dirigée des leucocytes à, et l’accumulation à des sites d’inflammation dans un modèle de mycotique (fongicide) kératite. Leber a souligné que l’attrait de l’excès de leucocytes par des substances d’origine pathogène est important pour l’élimination des micro-organismes nocifs par phagocytose, qui avait été décrit par Metchnikoff (également connu sous le nom Metschnikoff) plus tôt dans la même décennie13. Des expériences in vivo ont également été réalisées dans les années 1920 par Clark et Clark14,15, qui ont profité de la transparence des têtards et ont montré que l’inflammation stérile induite par l’huile de croton14 ou d’autres irritants15 a causé des leucocytes à adhérer aux vaisseaux sanguins, suivie par la diapédèse (migration transendéliheliale) et la migration rapide à travers les espaces tissulaires vers l’irritant. Des expériences in vitro utilisant la méthode de microcinematographie développée par Jean Comandon16 ont montré que les leucocytes migrent vers une source de chimiottractant particulaire comme les bactéries17. À cette époque, les identités moléculaires des facteurs chemotatiques étaient inconnues. Dans les années 1960, Stephen Boyden18 a reconnu que les techniques d’étude de l’activité chimiotique des substances solubles manquaient. Il a conçu une chambre, plus tard connue sous le nom de chambre Boyden, avec deux compartiments séparés par une membrane de papier filtre. Une suspension cellulaire est ajoutée au compartiment supérieur et la substance d’essai est ajoutée aux deux compartiments ou uniquement au compartiment inférieur. Après une période d’incubation, la membrane du filtre est enlevée, et les cellules sont fixes et tachées. En comparant le nombre de cellules migrent à travers la membrane du filtre vers le puits inférieur avec la substance d’essai dans les deux compartiments, dans aucun des compartiments, ou seulement dans le compartiment inférieur, l’activité chimiotique peut être déterminée. Les essais transwell sont encore populaires aujourd’hui et ont été modifiés de diverses manières, y compris l’utilisation de différentes membranes en polycarbonate avec des tailles et des densités définies de pores19,20. Un inconvénient majeur des essais De Transwell est qu’il n’est pas pratique de visualiser directement les cellules migrer et le chemin de migration à travers la membrane ne dépasse généralement pas le diamètre d’une cellule immunitaire.
Sally H. Zigmond a développé une chambre21 chimiotaxis qui a permis la visualisation de la formation de gradient et de la morphologie cellulaire utilisant des colorants fluorescents. La chambre se compose d’une glissière en plexiglas (acrylique) avec deux puits linéaires parallèles, chacun avec un volume de 100 ll, séparé par un pont de 1 mm de large de 3 à 10 m sous le plan supérieur de la glissière. Un coverlip ensemencé avec des cellules est inversé et placé sur la glissière de telle sorte qu’il s’étende sur les deux puits. Après l’ajout d’un chimioattractant à l’un des puits, un gradient chimioattractant raide se forme à travers le pont, généralement dans les 30-90 min. Les leucocytes polymorphonucléaires humains (granulocytes) dans la chambre Zigmond sont observés s’orientant vers le chimiottractant. Des variantes de la chambre Zigmond ont été signalées, y compris les chambres Dunn22 et Insall23, qui utilisent toutes deux un coverlip ensemencé avec des cellules placées sur deux puits séparés par un pont de 1 mm de large. La chambre Dunn se compose de puits concentriques séparés par un pont circulaire, tandis que la chambre Insall est plus étroitement liée à la chambre Zigmond, mais fournit des ponts de deux largeurs différentes, 0,5 mm et 1 mm. Une nouvelle chambre de chimiotaxis, appelée Chimiotaxis de gligéque et fabriquée par moulage d’injection en plastique, a été décrite par Zemgel et coll.24. La chambre de chimiotaxis se compose de deux réservoirs de 40 L séparés par un canal de 1 mm de large d’une longueur de 2 mm et d’une hauteur de 70 m. Le fond de la chambre est formé par une feuille de plastique perméable et mince avec la même épaisseur et les propriétés optiques d’un couvre-verre no 1.524. Ici, nous décrivons un essai de chimiotaxis utilisant la chambre de Chimiotaxis de glidre pour visualiser la migration des macrophages péritonéaux de résident de souris pour jusqu’à 14 h dans un gradient chemotactic (complément C5a).
Protocole
Les protocoles suivent les lignes directrices de notre comité local d’éthique de la recherche, ainsi que les lignes directrices sur les soins aux animaux.
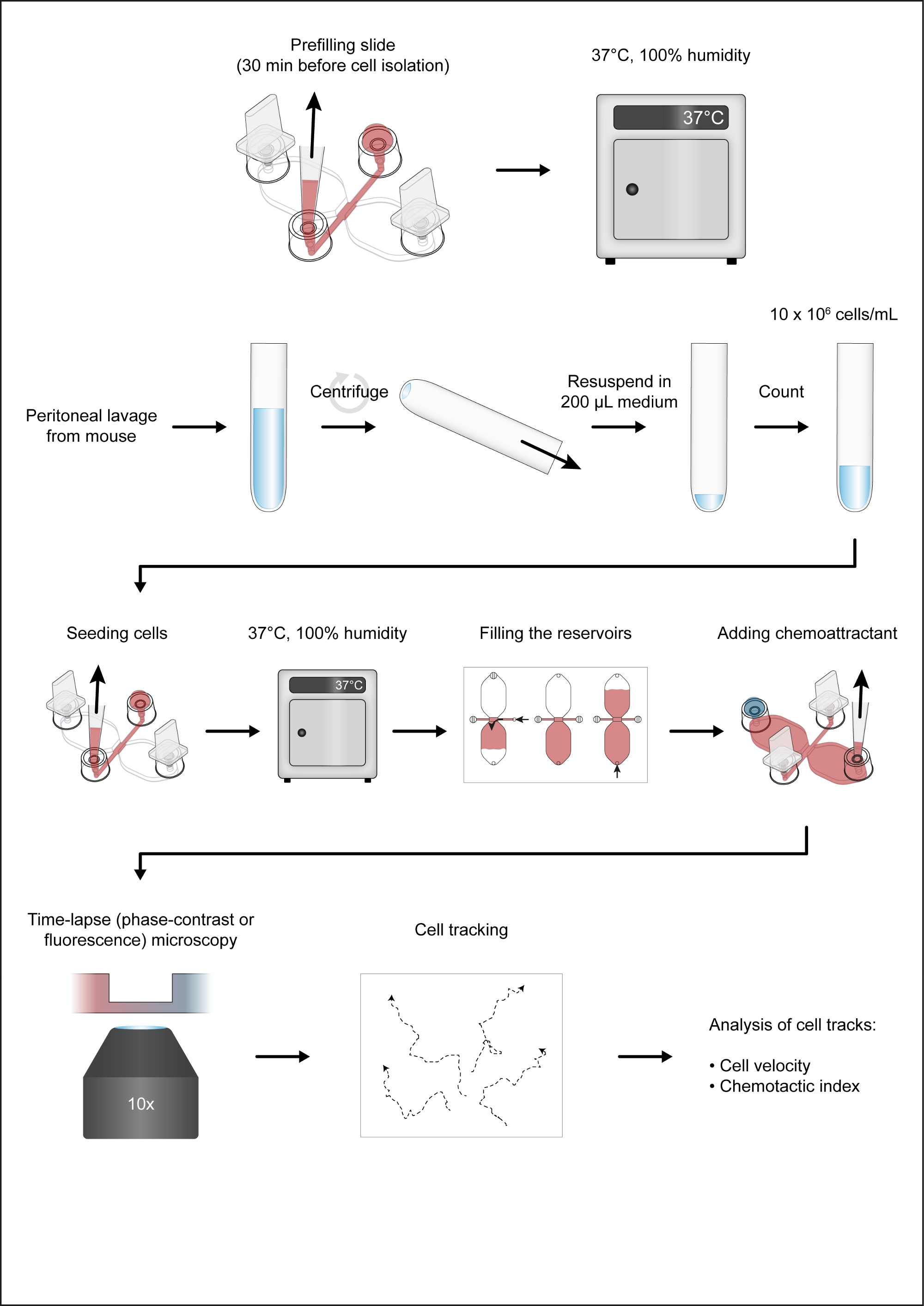
REMARQUE : La figure 1 montre un flux de travail de l’essai de chimiotaxis.
1. Préfiltrer les diapositives Chemotaxis
-
Préfillez les canaux de raccordement de 1 mm de large et de 2 mm de long d’une ou deux diapositives de chimiotaxis à l’aide du milieu RPMI 1640 HEPES modifié, composé de RPMI 1640 sans bicarbonate contenant 20 mM 4-(hydroxéthyle)-1-piperazinezineethane l’acide bosïton (HEPES), le sérum bovin fœtal (FBS) à 10 % inactivé par la chaleur et les antibiotiques tels que la pénicilline (100 U/mL) et la streptomycine (100 g/mL), préparés en diluant la pénicilline/streptomycine 100x, et 1 g/mL lipopolysaccharide (d’E. coli),et un récepteur 4 ligand de toll utilisé pour activer les cellules.
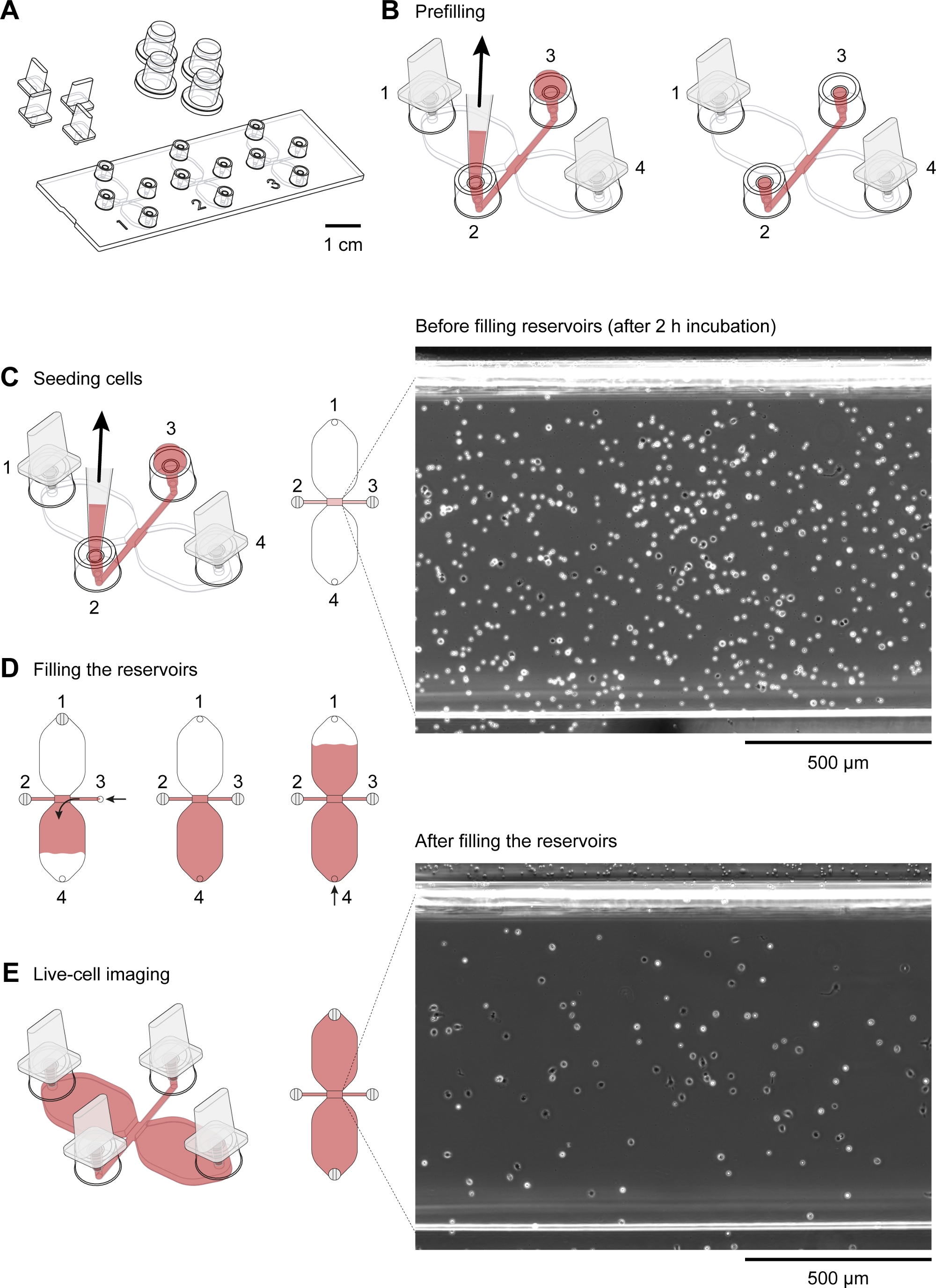
- Placer une glissière de chimiotaxis(figure 2A) dans un plat rond de culture cellulaire (10 cm de diamètre), à la fois préchauffé à 37 oC, et placer le plat sur un bloc d’aluminium chauffé (37 oC). Insérer des bouchons dans les ports 1 et 4(figure 2B).
REMARQUE: Un bloc d’aluminium maintenu à 37 oC et placé à l’intérieur du capot de flux lamineur est utile pour préparer les chambres de chimiotaxis. Idéalement, le bloc chauffé devrait fournir une zone de travail plate et des puits pour divers tubes, tels que des tubes de 50 ml et des tubes de microcentrifuge de 2 ml. - À l’aide d’une pointe de pipette de 10 à 200 l avec une pointe biseautée, déposez 15 L de milieu RPMI 1640 HEPES modifié dans le remplissage du port 3(figure 2B). Ensuite, avec le volume encore réglé à 15 l et le bouton de commande de la pipette (2-20 L) déprimée, insérer la pointe de la pipette dans le port 2 et aspirer 15 ll à un rythme modérément rapide(figure 2B). Cela préfulera le canal de raccordement de 1 mm x 2 mm (zone d’observation), ainsi que les deux canaux d’approvisionnement en flanque (entre la zone d’observation centrale et les ports 2 et 3, respectivement). Couvrir les ports de remplissage 2 et 3 avec des bouchons.
- Après le préremvrage, placez les chimiotaxis glisser sur un support conservé dans une chambre d’humidité fermée dans un incubateur autrement sec et sans CO2à 37 oC.
REMARQUE: Il est important d’utiliser la pointe de pipette correcte pour remplir la glissière de chimiotaxis. Une pointe de pipette biseautée s’insurce les coins dans le haut du port de remplissage, tandis que les pointes de pipette pointues couramment utilisées peuvent être insérées plus profondément dans le port de remplissage et peuvent augmenter considérablement la résistance à l’écoulement des fluides.
- Placer une glissière de chimiotaxis(figure 2A) dans un plat rond de culture cellulaire (10 cm de diamètre), à la fois préchauffé à 37 oC, et placer le plat sur un bloc d’aluminium chauffé (37 oC). Insérer des bouchons dans les ports 1 et 4(figure 2B).
2. Isolation des macrophages péritonéaux résidents de souris
- Sacrifiez une souris de 3 à 4 mois à l’aide d’une forte concentration de l’isoflurane anesthésique volatil (5 % dans l’air) ou de dioxyde de carbone25, suivi d’une dislocation cervicale. La perte du réflexe de redressement chez les rongeurs est corrélée avec la perte de conscience chez l’homme26. Nettoyer l’abdomen de la souris avec 80% d’éthanol dans l’eau, puis faire une incision de la peau moyenne de 1 à 2 cm à l’aide de ciseaux chirurgicaux avec des pointes émoussées. Peler la peau pour exposer la paroi abdominale sous-jacente.
- Insérez un cathéter en plastique de 24 G dans la cavité péritonéale. À l’aide d’une seringue en plastique de 5 ml, lavez la cavité à l’aide de la solution de sel tampon de Hank(HBSS) de 2 x 4,5 ml de glace froide, sans Ca2 et Mg2. Laissez environ 0,5 ml de HBSS résiduel dans la seringue afin que les tissus aspirés par inadvertance sur la pointe du cathéter puissent être expulsés.
- Transférer le milieu lavé, généralement de 8 à 8,5 ml au total, dans un tube de fond rond en polypropylène de 14 mL. Centrifuge le tube à 300 x g pendant 6,5 minutes à température ambiante.
REMARQUE: Le tube de fond rond permet au supernatant d’être entièrement décanté et réduit l’agglutination cellulaire. - Jetez le supernatant et réutilisez les cellules péritonéales (généralement 4 x 106 cellules par souris) dans 200 L du milieu RPMI 1640 HEPES modifié. Diluer un échantillon de la suspension cellulaire 1:20 et utiliser un dispositif de comptage, comme une chambre de comptage améliorée Neubauer, pour compter les cellules. Ensuite, diluer la suspension cellulaire à une concentration finale de 10 x 106 cellules/mL et maintenir les cellules dans un tube rond de microcentrifuge en polypropylène de fond de 2 ml à 37 oC à l’aide d’un bloc d’aluminium chauffé (voir NOTE à l’étape 1.1.1).
3. Ensemencement des cellules péritonéales dans les diapositives de chimiotaxis
- Après avoir conduit la suspension cellulaire de haut en bas de 5x avec le volume de pipette fixé à 100 L (ou à la moitié du volume de suspension) pour réduire l’agglutination, déposer délicatement 10 L de la suspension cellulaire dans le port 3 d’une chambre de chimiotaxis(figure 2C). Placez la pointe de la pipette dans le port 2 et dessinez lentement la suspension cellulaire dans le canal de raccordement(figure 2C). Dès que la suspension de la cellule a été introduite, retirez les bouchons aux ports 1 et 4, ce qui aidera à arrêter le flux de la suspension cellulaire. Placez des bouchons sur les quatre ports de remplissage.
- Répétez l’étape 3.1 pour toutes les chambres de chimiotaxis. À l’aide d’un petit microscope inversé et d’une lentille objective de phase-contraste 10x, inspectez les diapositives de chimiotaxis pour les bulles d’air indésirables.
- Placer les diapositives de chimiotaxis ensemencées avec des cellules péritonéales dans une chambre d’humidité à 37 oC pendant 2 à 3 h.
4. Remplir les réservoirs et ajouter le chemoattractant
- Inspectez la zone d’observation (canal reliant les deux réservoirs de 40 L) à l’aide d’un microscope inversé.
REMARQUE: À ce stade, la densité cellulaire sera plus élevée qu’après le remplissage des réservoirs, car les cellules faiblement adhérentes, principalement lescellules CD19 (cellules B1), seront lavées de la zone d’observation pendant la procédure de remplissage(figure 2C-E). - Placez les bouchons dans les ports de remplissage 1 et 2(figure 2D). Assurez-vous que le port de remplissage 3 est rempli au sommet avec moyen et exempt de bulles d’air. Utilisez une aiguille stérile de seringue 27 G pour déloger les bulles d’air indésirables, si nécessaire.
- À l’aide d’une pipette mécanique de volume de 10 à 10 l, aspirez à 60 lL de milieu HEPES RPMI 1640 modifié et placez la pointe de la pipette dans le port de remplissage 3. Utilisez l’anneau de réglage du volume de la pipette pour injecter lentement et régulièrement le milieu dans le réservoir de telle sorte que le milieu atteint le sommet du port de remplissage 4 après 1-2 min (figure 2D).
- Remplissez le deuxième réservoir. Déplacez la prise du port 1 et insérez-la lentement dans le port 3(figure 2D). Ensuite, aspirez à 50 lil de milieu HEPES RPMI 1640 modifié et placez la pointe de la pipette dans le port de remplissage 4. Utilisez l’anneau de réglage du volume de la pipette de volume de 10 à 100 l pour injecter lentement et régulièrement le milieu dans le deuxième réservoir de telle sorte que le milieu atteint le sommet du port de remplissage 1 après 1 à 2 min (figure 2D).
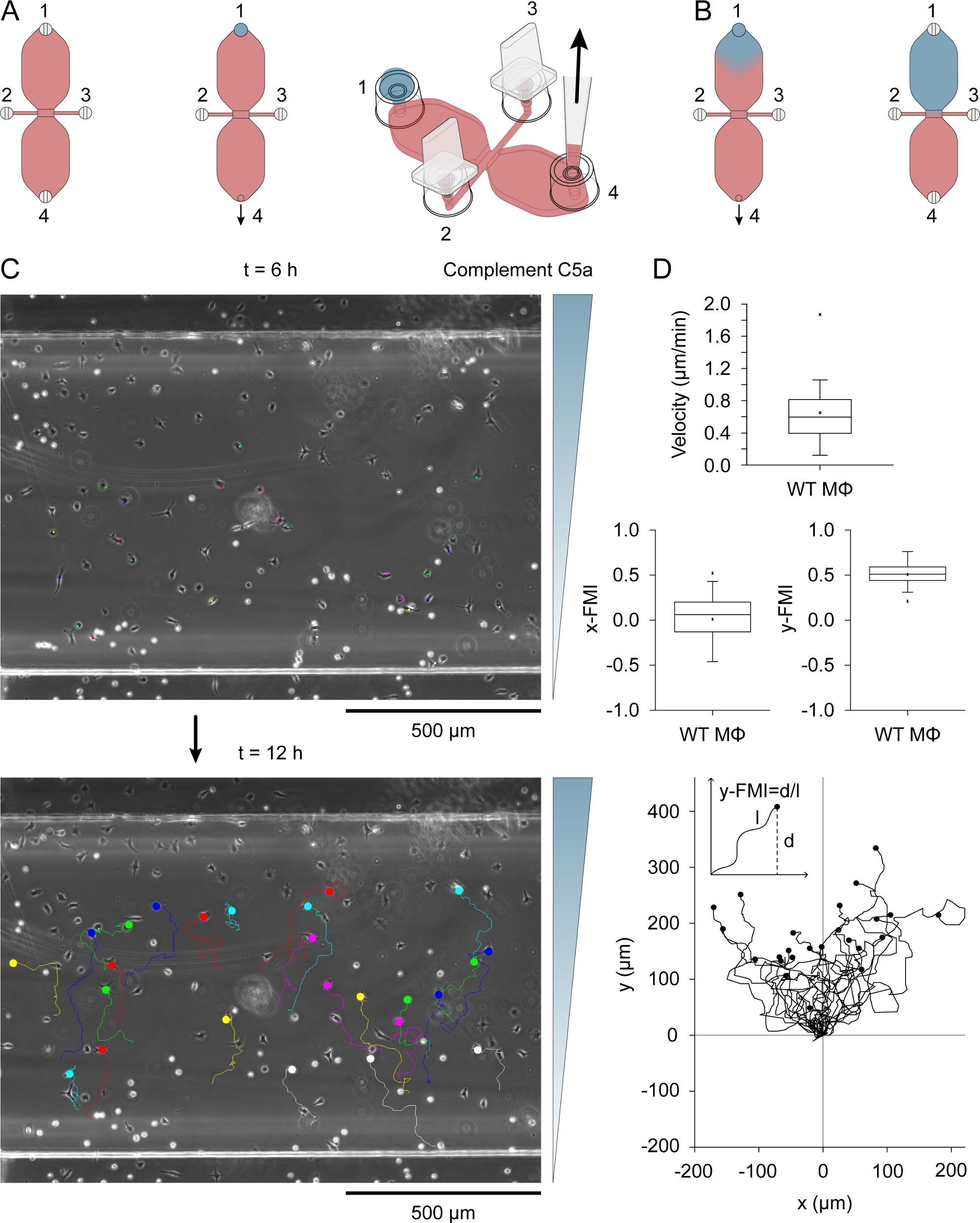
- Placez 495 L de RPMI 1640 HEPES modifié dans un tube de microcentrifuge de 2 ml (en bas rond) et ajoutez 5 L de Patent Blue V (solution de stock : 10 mg/mL en saline tamponnée par phosphate [PBS]), un colorant bleu utilisé comme indicateur visuel de la formation de gradient de concentration. Mélanger par un bref vortexing. Ajouter 5,4 L de complément de souris recombinante C5a (solution stock: 50 g/mL dans PBS avec 0,1% albumin de sérum bovin) et mélanger par vortexing bref.
- Déposez 15 L de bleu, complétez le milieu contenant du C5a dans le port de remplissage 1(figure 3A), après s’être assuré que la dépression peu profonde au sommet du port est moyenne libre (sinon la chute peut se déverser).
- Insérez une pointe de pipette de 10 à 200 L dans le port de remplissage 4 et faites pivoter lentement et régulièrement l’anneau de réglage du volume de la pipette de volume de 10 à 100 L pour dessiner la goutte de milieu bleu contenant du C5a dans le réservoir opposé(figure 3B). L’air commencera à entrer dans la courte colonne verticale du port de remplissage 1. Tirez de l’air jusqu’à ce que l’interface fluide-air soit à mi-chemin dans la colonne verticale, puis insérez lentement un bouchon dans le port.
- Soulevez doucement la pipette du port 4, en utilisant l’autre main pour s’assurer que la glissière reste fixée en place. Enfin, branchez lentement le port 4(figure 3B).
- Inspectez la glissière de chimiotaxis sur un microscope inversé.
REMARQUE: Les cellules adhérentes restantes dans la zone d’observation devraient être principalement des macrophages. Ceci peut être confirmé à l’aide d’anticorps anti-F4/80 marqués fluorescents (F4/80 est un marqueur spécifique pour les macrophages de souris). Les cellules B peuvent être identifiées à l’aide d’anticorps anti-CD19 marqués fluorescents et les cellules F4/80-/CD19- peuvent être détectées à l’aide d’une tache d’acide nucléique fluorescent bleu(figure 4).
5. Migration de macrophage d’imagerie par Time-lapse, Microscopy de phase-contraste
- Placez une glissière de chimiotaxis sur la scène d’un microscope inversé équipé d’un incubateur de scène. Maintenir la température à 37 oC.
- Imagez la zone d’observation de 1 mm x 2 mm à l’aide d’une lentille objective de phase-contraste 10x et concentrez-vous sur le lamellipodia macrophage : fines, des saillies de membrane. Capturez des images pendant 14 h à raison de 1 image toutes les 2 minutes.
6. Analyse des images Time-lapse
- Analyser le time-lapse, les images de phase-contraste à l’aide d’un logiciel automatisé d’analyse d’images ou le plugin Manual Tracking, produit par Fabrice P. Cordelières, pour ImageJ.
REMARQUE: Les programmes de suivi automatisé peuvent être utilisés pour analyser les cellules images par une microscopie en time-lapse, en contraste de phase ou en fluorescence. Par exemple, le logiciel iTrack4U basé à Java produit par Cordelières et coll.27 peut être utilisé pour le suivi et l’analyse automatisés des cellules à l’aide d’images en time-lapse, en contraste de phase ou en fluorescence comme entrée. Le suivi manuel prend plus de temps, mais les pistes générées par le suivi manuel du plugin ImageJ peuvent être directement importées et analysées automatiquement par le plugin ImageJ Chemotaxis et l’outil de migration28,29.
Résultats
Un diagramme schématique de la diapositive de chimiotaxis utilisé pour la microscopie vidéo en time-lapse des macrophages péritonéaux de souris migrent dans un gradient chemotactic est montré dans la figure 2A. La diapositive contient trois chambres de chimiotaxis, chacune d’entre elles a quatre ports de remplissage. Les ports peuvent être fermés individuellement à l’aide des prises indiquées au-dessus de la glissière. Alternativement, un bouchon de non-étanchéité peut être placé au-dessus d’un port débranché pour maintenir la stérilité. Après avoir colmaté les ports 1 et 4, la zone d’observation (1 mm de large x 2 mm de long x 70 m de haut canal reliant les deux réservoirs) entre les ports 2 et 3 peut être préremitée avec moyen en plaçant une chute de 15 l dans le port 3 et en aspirant avec une pipette de volume de 2 à 20 L au port 2(figure 2B). Une suspension des cellules péritonéales résidentes de souris (10 x 106 cellules/mL) a été ensemencée dans la zone d’observation en plaçant une goutte de suspension de 10 L dans le port 3 et en aspirant lentement au port 2(figure 2C). Une image typique des cellules semées dans la zone d’observation prise par microscopie de contraste de phase utilisant une lentille objective 10x est montrée dans la figure 2C. Après avoir cocu pendant 2 à 3 h, la glissière de chimiotaxis a été lentement remplie de milieu(figure 2D). Après avoir branché les ports 1 et 2, le milieu a été lentement injecté par port 3 jusqu’à ce qu’il sorte du port 4. Ensuite, la prise a été commutée du port 1 au port 3, puis le deuxième réservoir a été rempli en injectant lentement le milieu par port 4 jusqu’à ce qu’il émerge au port 1. À ce stade, les cellules de la zone d’observation ont été réinspectées à l’aide d’un microscope inversé(figure 2E). En comparant des images peu de temps avant(figure 2C) et après (figure 2E) le remplissage des réservoirs, jusqu’à deux tiers des cellules avaient été lavées de la zone d’observation. En général, les cellules CD19et B1 faiblement adhérentes (cellules B1) ont été lavées et les cellules restantes étaient principalement des cellules F4/80 (macrophages).+ Ceci a été démontré par la microscopie de fluorescence après l’étiquetage de chaque type de cellule avec des anticorps spécifiques étiquetés fluorescents(figure 4). Dans la figure 4A, les cellules péritonéales résidentes de souris fraîchement isolées ont été étiquetées avec des anticorps anti-F4/80 anti-F4/80 fluorophores fluorescents rouges et des anticorps anti-CD19, et les noyaux des cellules ont été étiquetés avec une tache d’acide nucléique fluorescent bleu. F4/80 est un marqueur spécifique pour les macrophages de souris30, tandis que CD19 est un marqueur de cellules B. La figure 4B montre des cellules F4/80- illustrées par microscopie confocale de disque tournant dans la zone d’observation d’une chambre de chimiotaxis.+ Les cellules ont été étiquetées après un essai de chimiotaxis de nuit enregistré par time-lapse, microscopie de phase-contraste.
Le complément C5a (chimioattractant) a été introduit dans l’un des deux réservoirs en plaçant une goutte de 15 L de milieu contenant 0,54 g/mL (souris recombinante) complète C5a et 10 g/mL Patent Blue V dans le port de remplissage 1(figure 3A) après avoir branché les ports 2 et 3. Le milieu chimioattractant a été lentement entraîné dans le réservoir par aspiration lente avec une pipette via le port 4. La figure 3B montre la diffusion du colorant bleu après avoir dessiné la chute de 15 L dans un réservoir. Patent Blue V a été utilisé comme indicateur visuel indirect de la diffusion chimiotractante. Les molécules de Complément C5a sont considérablement plus grandes que celles de Patent Blue V (9,0 kDa contre 0,57 kDa) et diffusent plus lentement. Après diffusion du complément C5a dans le réservoir, sa concentration était de 0,2 g/mL (15 L/40 L [volume de réservoir] x 0,54 g/mL, soit 0,2 g/mL), soit l’équivalent de 22,5 nM. Un gradient légèrement raide s’est formé à travers la zone d’observation après 3 h et a continué à augmenter, atteignant un maximum à environ 12 h31. La figure 3C montre les voies de migration des macrophages migrent dans un gradient C5a de complément, entre 6-12 h après l’ajout du chimioattractant. Vitesse cellulaire et efficacité chimiocticale, indexées comme y-FMI (indice de migration à terme; gamme: -1 à 1) et x-FMI, des macrophages individuels ont été calculés à partir des parcelles de migration(figure 3D). La figure 3D montre également une parcelle de migration produite après avoir normalisé le point de départ de chaque piste de migration à X 0 et Y 0 en dessous des parcelles de boîte. L’encart dans la parcelle de migration montre comment le y-FMI a été calculé pour chaque voie de migration.

Figure 1 : Le flux de travail de l’essai de chimiotaxis. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 2 : Manipulation des diapositives de chimiotaxis. (A) Une vue 3D d’une diapositive de chimiotaxis avec quatre prises et quatre bouchons. La glissière contient trois chambres de chimiotaxis, chacune se compose de deux réservoirs de 40 L reliés par un canal de 1 mm x 2 mm, qui est de 70 m de haut, appelé zone d’observation. (B) Le canal de connexion s’étend aux deux extrémités pour remplir les ports 2 et 3. Après avoir inséré des bouchons dans les ports de remplissage 1 et 4, la zone d’observation a été préremlayée par une goutte de moyenne au port 3 et en aspirant au port 2 avec une pointe de pipette de 10 à 200 l. Par la suite, des plafonds ont été appliqués aux ports 2 et 3 avant d’incuber la glissière à 37 oC et de préparer la suspension cellulaire. (C) La zone d’observation, où le gradient chimioattractant s’est formé, a été ensemencée de macrophages en appliquant une goutte de 10 L de cellules péritonéales résidentes de souris au port 3 et en aspirant lentement au port 2. La glissière a ensuite été incubée dans une chambre d’humidité à 37 oC pendant 2 à 3 h. L’image de contraste de phase montrée sur la droite, obtenue par une lentille objective 10x, montre des cellules péritonéales après l’ensemencement et l’incubation à 37 oC pour 2 h. Barre d’échelle de 500 m. (D) les chambres Chemotaxis ont été remplies de milieu en branchant les ports 1 et 2, puis en injectant lentement le milieu par le port 3 jusqu’à ce qu’il émerge au port 4. Un remplissage lent et régulier peut être réalisé en tournant l’anneau de réglage du volume d’une pipette de volume de 20 à 100 l. Après avoir rempli le premier réservoir, le deuxième réservoir peut être rempli en branchant les ports 2 et 3, puis en injectant lentement le milieu au port 4 jusqu’à ce qu’il émerge au port 1. (E) Image de contraste de phase de la même zone d’observation indiquée ci-dessus (C) après le remplissage des deux réservoirs. Barre d’échelle de 500 m. Éléments graphiques fournis par Elias Horn. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 3 : Chimiotaxis assay. (A) Chemoattractant a été introduit à l’un des deux réservoirs d’une chambre de chimiotaxis en appliquant une goutte de 15 L de milieu contenant 0,54 g/mL complément C5a et 10 g/mL Patent Blue V au port de remplissage 1, suivie d’une aspiration lente au port 4. (B) D’abord après avoir été entraîné dans le réservoir, le milieu bleu contenant des chimiorattractants avait une forme de chute à peu près inversée, puis lentement diffusé dans tout le réservoir. (C) Les pistes migratoires des macrophages migrent dans un gradient chimioattractant (complément C5a) entre 6-12 h après l’introduction de la chimioattractante à l’un des réservoirs. La direction du gradient est indiquée à droite. La fin de chaque voie de migration est indiquée par un cercle rempli. (D) Blox traces de vitesse, x-FMI (indice de migration x-forward) et y-FMI (indice de migration à terme), un indice de l’efficacité chimioctic qui varie de -1 à 1. Les données ont été obtenues par l’analyse de 25 voies de migration macrophage. Des macrophages dans la moitié inférieure de la zone d’observation et montrant le déplacement d’au moins une largeur de cellule au-dessus de 6 h ont été choisis au hasard pour l’analyse. Voici une parcelle de pistes de migration après avoir normalisé le point de départ à X 0 et Y 0. L’indice de chimiotaxis (y-FMI) a été calculé en divisant le déplacement net le long de l’axe Y (d) par la longueur accumulée (l) de la trajectoire migratoire, comme le montre schématiquement. Éléments graphiques dans les panneaux A et B fournis par Elias Horn. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 4 : Images fluorescentes de cellules péritonéales résidentes vivantes de souris obtenues en tournant la microscopie confocale de disque. (A) Image de mise au point étendue (plus brillante fusion de tous les Z-planes) de cellules péritonéales souris fraîchement isolées étiquetées avec des anticorps fluorescents verts anti-F4/80 (marqueur macrophage), anticorps fluorescents rouges anti-CD19 (marqueur cellulaire B) et une tache d’acide nucléique fluorescent bleu. Barre d’échelle de 10 m (B) Instantané (mono-plan Z) de F4/80- cellules (macrophages) dans la zone d’observation d’une chambre de chimiotaxis prise après un essai de chimiotaxis de nuit. Les cellules ont été étiquetées avec des anticorps fluorescents verts anti-F4/80 et une tache bleue d’acide nucléique fluorescent. Les gradients C5a et Patent Blue V ont été éliminés par la procédure d’étiquetage cellulaire, ce qui explique pourquoi le réservoir supérieur dans le diagramme schématique de la chambre de chimiotaxis n’est pas bleu. Barre d’échelle de 10 m. Élément graphique fourni par Elias Horn. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}
Discussion
L’imagerie intravitale remonte au 19ème siècle et fournit un moyen d’étudier le comportement des cellules immunitaires vivantes dans leur environnement naturel. Cependant, même avec la microscopie avancée d’aujourd’hui et les techniques génétiques, il est difficile d’étudier la réponse des cellules à des chimiorattractants spécifiques in vivo. Pour contourner ce problème, Boyden18 a développé des essais Transwell dans les années 1960, mais ces essais de point final n’ont pas fourni la visualisation de la façon dont les cellules effectivement migré vers les chimiorattractants, ce qui rend difficile de distinguer la chimiokinesis, stimulé la migration aléatoire par un indice chimique32, et chimiotaxis, la migration vers des concentrations plus élevées de stimuli chimiques les uns des autres33. Ce problème a été résolu en concevant diverses chambres ouvertes avec un pont, généralement de 1 mm de large, situé entre deux réservoirs et accessible par une lentille objective21,22,23. L’application d’un glissement de couverture inversé, ensemencé avec des cellules adhérentes, ferme les chambres et le chimiorattractant ajouté à l’un des réservoirs diffuse à travers le pont au réservoir opposé, créant un gradient de concentration. Ici, nous décrivons un essai de chimiotaxis en utilisant le même principe, mais en utilisant une chambre fermée avec quatre ports de remplissage. À l’aide de ce système et de la microscopie de phase-contraste, nous avons développé un essai à l’image de la souris résident macrophages péritonéal migrer dans un complément chemotactic C5a gradient31,34,35,36. Cet essai, combiné avec des modèles de souris knock-out, s’est avéré instrumental dans l’étude des rôles de divers Rho GTPases et protéines motrices dans la morphologie macrophage, la motilité, et la chimiotaxis31,34,35,36,37. Nous avons également utilisé cette approche pour image des monocytes sanguins périphériques humains migrent sur une surface 2D ou dans une matrice de type I de collagène 3D38. En outre, l’essai convient aux macrophages dérivés de la moelle osseuse de souris ou aux macrophages dérivés des cellules précurseurs myéloïdes immortalisées sous condition39,40. Nous avons déjà utilisé des sacs en polytétrafluoroéthylène (PTFE) avec des adaptateurs luer aux cellules de moelle osseuse de culture et obtenir des macrophages34. L’avantage des sacs PTFE est que les cellules peuvent être facilement résutilisées et prêtes à l’emploi après avoir placé le sac sur la glace pendant 20-30 min. Notez que nous préfillons la zone d’observation de la diapositive chimiotaxis avant d’introduire les cellules. Cette approche a l’avantage que les bulles d’air indésirables peuvent être ensuite rincés (avec un succès variable) et la zone d’observation présoaked permet l’introduction lente d’une suspension cellulaire par tuyauterie. Le remplissage, cependant, augmente la probabilité que le milieu s’écoule partiellement dans l’un ou les deux réservoirs de flanc, ce qui favorisera l’ensemencement des cellules au-delà de la zone d’observation. Alternativement, la suspension cellulaire peut être directement pipetted dans une zone d’observation sèche, mais les bulles d’air indésirables ne peuvent pas être expulsées par la suite.
La cavité péritonéale de la souris contient deux populations principales de cellules : F4/80, macrophages et (plus petites) cellules CD19et B, à un rapport d’environ 1:2 (figure 4A). Ces deux populations cellulaires représentent plus de 95 % des cellules de cavité péritonéales, tandis que les cellules F4/80restantes-/CD19- peuvent généralement être identifiées comme cd11c- cellules (cellules dendritiques) ou cellules CD3et T. Les cellules B faiblement adhérentes sont lavées de la zone d’observation pendant le remplissage des réservoirs avec le milieu(figure 2). Après avoir ajouté le chimiorattractant à l’un des deux réservoirs, la microscopie en time-lapse et en contraste de phase peut être utilisée pour imager les cellules restantes (macrophages) migrer dans un gradient chimioattractant en évolution. La formation du gradient C5a complémentaire dans la zone d’observation, par diffusion d’un réservoir à l’autre, peut être simulée à l’aide d’un colorant fluorescent avec un poids moléculaire similaire. Un bon substitut pour le complément de souris recombinante C5a (poids moléculaire prévu, 9.0 kDa) est fluorescent étiqueté dextran (10 kDa)31. À l’aide de la microscopie confocienne, le gradient de fluorescence dans le chenal étroit (zone d’observation) reliant les deux réservoirs de la glissière de chimiotaxis peut être mesuré à intervalles fixes et les profils de concentration aux différents points de temps peuvent être tracés24,31. Nous ajoutons régulièrement un colorant bleu nonfluorescent (Patent Blue V) au milieu chimioattractant pour fournir un indicateur visuel pratique de la diffusion et de la formation de gradient. Dans un rayon de 1 h après l’introduction d’un milieu bleu contenant des chimiorattractants dans un réservoir, le réservoir apparaît uniformément bleu et, selon les lois de diffusion de Fick, un gradient se formera à travers la zone d’observation étroite reliant les réservoirs(figure 3B). Plusieurs jours sont nécessaires pour que le soluté (colorant bleu ou chimiorattractant) soit uniformément distribué.
La microscopie de fluorescence peut être remplacée par une microscopie de contraste de phase, qui offre des avantages pour le suivi cellulaire automatisé, parce que les cellules étiquetées fluorescentes peuvent être facilement distinguées de l’arrière-plan. Un autre avantage est que des populations spécifiques de cellules immunitaires peuvent être suivis sélectivement après l’étiquetage des marqueurs de surface avec des anticorps fluorescents. Nous avons utilisé cette approche de l’image du sang périphérique humain CD14- cellules (monocytes) migrer dans un fMLP chemotactic (N-formylmethionine-leucyl-phenylalanine) gradient38. De même, les anticorps anti-F4/80 fluorescents pourraient être utilisés pour imager les macrophages de souris migrer dans un gradient chemotactic de C5a. La phototoxicité est un inconvénient potentiel de l’utilisation de l’imagerie par fluorescence41. Cela peut être réduit par divers moyens42, y compris l’utilisation de fluorophores excités avec des longueurs d’onde plus longues et l’ajout d’antioxydants au milieu. Alternativement, les cellules étiquetées pouvaient d’abord être identifiées par microscopie de fluorescence et ensuite représentées par microscopie en temps-temps et en contraste de phase. Cependant, dans la pratique, les cellules se déplaçant à des vitesses modérément basses, telles que 1 m/min (macrophages) ou 4 m/min (monocytes), peuvent être images par intermittence par microscopie à fluorescence à intervalles de minutes, ce qui est bien toléré38. Nous avons précédemment utilisé la microscopie de fluorescence et la diapositive de chimiotaxis décrite ici pour les essais 3D de chimiotaxis38,43. Dans ce cas, les deux réservoirs ont été préremutés avec moyen et 15 'L chimioattractant-contenant milieu a été entraîné dans l’un des réservoirs immédiatement avant de piper lentement cellules étiquetées fluorescentes suspendus dans le type de collagène moyen contenant dans la zone d’observation. La partie difficile de cette procédure est la manipulation du type de collagène I, qui est concentré dans la solution acide. Le pH de la solution de collagène doit être neutralisé par l’ajout de solution alcaline avant de mélanger la solution de collagène glacé avec la suspension cellulaire. Le transfert du mélange collagène-cellule dans un incubateur à 37 oC initiera la polymérisation du collagène. Pendant l’incubation, la glissière doit être lentement tournée autour de son long axe afin que les cellules restent réparties uniformément dans les directions des axes X, Y et Z tandis que le collagène polymérise en gel. Un toboggan de chimiotaxis fermé connexe adapté aux essais de chimiotaxis 3D, avec six prises au lieu de quatre prises, a récemment été décrit29. Ce système permet d’introduire le mélange collagène-cellule dans la zone d’observation avant de remplir indépendamment chacun des réservoirs de flan de bois, parce que chaque réservoir dispose de deux ports de remplissage, plutôt que d’un seul port.
En résumé, nous décrivons un essai de chimiotaxis en temps réel qui permet la visualisation de cellules naviguant dans un gradient chemotactic sur une période de 6 heures ou plus. Ici, nous nous concentrons sur les macrophages, qui jouent un rôle majeur dans les maladies inflammatoires, mais ont été sous-représentés dans les essais chimiotaxis en temps réel par rapport aux cellules mobiles plus rapides comme les neutrophiles et les amibes Dictyostelium.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Ce travail a été soutenu par une subvention (HA 3271/3-2) de la DFG (Deutsche Forschungsgemeinschaft).
matériels
| Name | Company | Catalog Number | Comments |
| µ-Slide (anodized aluminium) rack | Ibidi, Martinsried, Germany | 80003 | Autoclavable stackable rack for channel slides |
| µ-Slide Chemotaxis 2D (chemotaxis slide) | Ibidi, Martinsried, Germany | 80306 | Slide containing chemotaxis chambers (tissue culture treated) |
| 100x penicillin/streptomycin | Thermo Fisher Scientific | 15140122 | Used as supplement for RPMI 1640 media |
| 10-100 µL pipette with volume control ring | Eppendorf | 3123000047 | Eppendorf Research plus pipette |
| 10-200 µL pipette tips | Greiner Bio-One International | 739261 | Pipette tips with beveled tips (96 pieces per rack: sterile) |
| 14 mL polypropylene round bottom tubes | BD Falcon | 352059 | Used to collect peritoneal cells |
| 14-bit Hamamatsu C9100-50 Electron Multiplying-Charged Couple Device (EM-CCD) peltier-cooled camera | Hamamatsu Photonics Inc., Japan | EM-CCD camera of the spinning disk confocal microscope system | |
| 2-20 µL pipette with volume control ring | Eppendorf | 3123000039 | Eppendorf Research plus pipette |
| 24 G plastic catheter | B Braun Mesungen AG, Germany | 4254503-01 | Used for peritoneal lavage |
| 405 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (405 nm) source of spinning disk confocal microscope system | |
| 488 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (488 nm) source of spinning disk confocal microscope system | |
| 561 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (561 nm) source of spinning disk confocal microscope system | |
| Alexa Fluor 488-conjugated rat (IgG2a) monoclonal (clone BM8) anti-mouse F4/80 antibody | Thermo Fisher Scientific | MF48020 | Mouse macrophage marker and plasma membrane label |
| Alexa Fluor 594-conjugated rat (IgG2a) monoclonal (clone 6D5) anti-mouse CD19 antibody | BioLegend | 115552 | Mouse B cell marker |
| C-Chip disposable (improved Neubauer) hemocytometer | NanoEnTek (distributed by VWR International) | 631-1098 | Used to count cells |
| CSU-X1 spinning disk scanner | Yokogawa Electric Corporation, Japan | Nipkow spinning disk unit | |
| Hank’s buffered salt solution without Ca2+ and Mg2+ | Thermo Fisher Scientific | 14170120 | Used for peritoneal lavage |
| Heat-inactivated fetal bovine serum | Thermo Fisher Scientific | 10082139 | Used as supplement for RPMI 1640 media |
| Hoechst 34580 | Thermo Fisher Scientific | H21486 | Cell permeable, blue fluorescent nucleic acid stain |
| ImageJ (image processing and analysis in Java) | National Institutes of Health (NIH) | Image analysis software | |
| Lipopolysaccharides from Escherichia coliO111:B4 | Sigma-Aldrich | L4391-1MG | Toll-like receptor 4 ligand |
| Nikon Eclipse Ti inverse microscope | Nikon, Japan | Inverted microscope | |
| Patent Blue V, sodium salt | Sigma-Aldrich | 21605-10G | Blue-colored dye used as visual indicator of gradient formation |
| Recombinant mouse complement C5a protein | R&D Systems | 2150-C5-025 | Chemoattractant for mouse macrophages |
| RPMI 1640 medium containing 20 mM Hepes | Sigma-Aldrich | R7388 | Basis medium for assays |
| UltraVIEW Vox 3D live cell imaging system + Volocity software | Perkin Elmer, Rodgau, Germany | Spinning disk confocal microscope system | |
| Zeiss LSM 510 + Axiovision software | Carl Zeiss Microscopy, Oberkochen, Germany | Confocal laser scanning microscope (LSM) adapted for phase-contrast microscopy |
Références
- Lammermann, T., Germain, R. N. The multiple faces of leukocyte interstitial migration. Seminars in Immunopathology. 36, 227-251 (2014).
- Lammermann, T., Sixt, M. Mechanical modes of 'amoeboid' cell migration. Current Opinion in Cell Biology. 21, 636-644 (2009).
- Woodham, E. F., Machesky, L. M. Polarised cell migration: intrinsic and extrinsic drivers. Current Opinion in Cell Biology. 30, 25-32 (2014).
- Devreotes, P. N., et al. Excitable Signal Transduction Networks in Directed Cell Migration. Annual Review of Cell and Developmental Biology. 33, 103-125 (2017).
- Kamp, M. E., Liu, Y., Kortholt, A. Function and Regulation of Heterotrimeric G Proteins during Chemotaxis. International Journal of Molecular Sciences. 17 (1), 90 (2016).
- Miao, Y., et al. Wave patterns organize cellular protrusions and control cortical dynamics. Molecular Systems Biology. 15, 8585 (2019).
- Ridley, A. J., et al. Cell migration: integrating signals from front to back. Science. 302, 1704-1709 (2003).
- Lammermann, T., et al. Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature. 453, 51-55 (2008).
- Mullins, R. D., Heuser, J. A., Pollard, T. D. The interaction of Arp2/3 complex with actin: nucleation, high affinity pointed end capping, and formation of branching networks of filaments. Proceedings of the National Academy of Sciences of the United States of America. 95, 6181-6186 (1998).
- Leithner, A., et al. Diversified actin protrusions promote environmental exploration but are dispensable for locomotion of leukocytes. Nature Cell Biology. 18, 1253-1259 (2016).
- McDonald, B., et al. Intravascular danger signals guide neutrophils to sites of sterile inflammation. Science. 330, 362-366 (2010).
- Leber, T. Ueber die Entstehung der Entzündung und die Wirkung der entzündungserregenden Schädlichkeiten. Fortschritte der Medizin. 6, 460-464 (1888).
- Tauber, A. I. Metchnikoff and the phagocytosis theory. Nature Reviews Molecular Cell Biology. 4, 897-901 (2003).
- Clark, E. R., Linton Clark, E. Reactions of cells in the tail of amphibian larvae to injected croton oil (aseptic inflammation). American Journal of Anatomy. 27, 221-254 (1920).
- Clark, E. R., Linton Clark, E. The reaction of living cells in the tadpole's tail toward starch, agar-agar, gelatin, and gum arabic. The Anatomical Record. 24, (1922).
- Comandon, J. Phagocytose in vitro des Hématozoaires du Calfat (enregistrement cinématographique). Comptes Rendus Hebdomadaires des Séances et Mémoires de la Société de Biologie. 69, 314-316 (1917).
- McCutcheon, M. Chemotaxis in leukocytes. Physiological Reviews. 26, 319-336 (1946).
- Boyden, S. The chemotactic effect of mixtures of antibody and antigen on polymorphonuclear leucocytes. The Journal of Experimental Medicine. 115, 453-466 (1962).
- Horwitz, D. A., Garrett, M. A. Use of leukocyte chemotaxis in vitro to assay mediators generated by immune reactions. I. Quantitation of mononuclear and polymorphonuclear leukocyte chemotaxis with polycarbonate (nuclepore) filters. Journal of Immunology. 106, 649-655 (1971).
- Bignold, L. P. A novel polycarbonate (Nuclepore) membrane demonstrates chemotaxis, unaffected by chemokinesis, of polymorphonuclear leukocytes in the Boyden chamber. Journal of Immunological Methods. 105, 275-280 (1987).
- Zigmond, S. H. Ability of polymorphonuclear leukocytes to orient in gradients of chemotactic factors. The Journal of Cell Biology. 75, 606-616 (1977).
- Zicha, D., Dunn, G. A., Brown, A. F. A new direct-viewing chemotaxis chamber. Journal of Cell Science. 99, 769-775 (1991).
- Muinonen-Martin, A. J., Veltman, D. M., Kalna, G., Insall, R. H. An improved chamber for direct visualisation of chemotaxis. PLoS One. 5, 15309 (2010).
- Zengel, P., et al. mu-Slide Chemotaxis: a new chamber for long-term chemotaxis studies. BMC Cell Biology. 12, 21 (2011).
- Valentim, A. M., Guedes, S. R., Pereira, A. M., Antunes, L. M. Euthanasia using gaseous agents in laboratory rodents. Lab Animal. 50, 241-253 (2016).
- Franks, N. P. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal. Nature Reviews. Neuroscience. 9, 370-386 (2008).
- Cordelieres, F. P., et al. Automated cell tracking and analysis in phase-contrast videos (iTrack4U): development of Java software based on combined mean-shift processes. PLoS One. 8, 81266 (2013).
- Zantl, R., Horn, E. Chemotaxis of slow migrating mammalian cells analysed by video microscopy. Methods in Molecular Biology. 769, 191-203 (2011).
- Biswenger, V., et al. Characterization of EGF-guided MDA-MB-231 cell chemotaxis in vitro using a physiological and highly sensitive assay system. PLoS One. 13, 0203040 (2018).
- Austyn, J. M., Gordon, S. F4/80, a monoclonal antibody directed specifically against the mouse macrophage. European Journal of Immunology. 11, 805-815 (1981).
- Hanley, P. J., et al. Motorized RhoGAP myosin IXb (Myo9b) controls cell shape and motility. Proceedings of the National Academy of Sciences of the United States of America. 107, 12145-12150 (2010).
- Wilkinson, P. C. Cell Locomotion and Chemotaxis: Basic Concepts and Methodological Approaches. Methods. 10, 74-81 (1996).
- Pfeffer, W. Locomotorische Richtungsbewegungen durch chemische Reize. Untersuchungen aus dem Botanischen Institut zu Tübingen. 1, 363 (1884).
- Konigs, V., et al. Mouse macrophages completely lacking Rho subfamily GTPases (RhoA, RhoB, and RhoC) have severe lamellipodial retraction defects, but robust chemotactic navigation and altered motility. The Journal of Biological Chemistry. 289, 30772-30784 (2014).
- Horsthemke, M., et al. Multiple roles of filopodial dynamics in particle capture and phagocytosis and phenotypes of Cdc42 and Myo10 deletion. The Journal of Biological Chemistry. 292, 7258-7273 (2017).
- Bachg, A. C., et al. Phenotypic analysis of Myo10 knockout (Myo10(tm2/tm2)) mice lacking full-length (motorized) but not brain-specific headless myosin X. Scientific Reports. 9, 597 (2019).
- Horsthemke, M., et al. A novel isoform of myosin 18A (Myo18Agamma) is an essential sarcomeric protein in mouse heart. The Journal of Biological Chemistry. 294, 7202-7218 (2019).
- Bzymek, R., et al. Real-time two- and three-dimensional imaging of monocyte motility and navigation on planar surfaces and in collagen matrices: roles of Rho. Scientific Reports. 6, 25016 (2016).
- Wang, G. G., et al. Quantitative production of macrophages or neutrophils ex vivo using conditional Hoxb8. Nature Methods. 3, 287-293 (2006).
- Gran, S., et al. Imaging, myeloid precursor immortalization, and genome editing for defining mechanisms of leukocyte recruitment in vivo. Theranostics. 8, 2407-2423 (2018).
- Magidson, V., Khodjakov, A. Circumventing photodamage in live-cell microscopy. Methods in Cell Biology. 114, 545-560 (2013).
- Icha, J., Weber, M., Waters, J. C., Norden, C. Phototoxicity in live fluorescence microscopy, and how to avoid it. BioEssays : News and Reviews in Molecular, Cellular and Developmental Biology. 39 (8), 1700003 (2017).
- Isfort, K., et al. Real-time imaging reveals that P2Y2 and P2Y12 receptor agonists are not chemoattractants and macrophage chemotaxis to complement C5a is phosphatidylinositol 3-kinase (PI3K)- and p38 mitogen-activated protein kinase (MAPK)-independent. The Journal of Biological Chemistry. 286, 44776-44787 (2011).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.