
Un abonnement à JoVE est nécessaire pour voir ce contenu. Connectez-vous ou commencez votre essai gratuit.
Method Article
Manipulation subcellulaire spatio-temporelle du cytosquelette de microtubules dans l’embryon de souris préimplantatoire vivant à l’aide de photostatines
Dans cet article
Résumé
Les inhibiteurs typiques des microtubules, largement utilisés dans la recherche fondamentale et appliquée, ont des effets de grande portée sur les cellules. Récemment, les photostatines sont apparues comme une classe d’inhibiteurs de microtubules photoscommutables, capables de manipulation instantanée, réversible et spatio-temporelle précise des microtubules. Ce protocole étape par étape détaille l’application de photostatines dans un embryon de souris préimplantatoire vivant en 3D.
Résumé
Le cytosquelette des microtubules forme le cadre d’une cellule et est fondamental pour le transport intracellulaire, la division cellulaire et la transduction du signal. La perturbation pharmacologique traditionnelle du réseau de microtubules omniprésents en utilisant, par exemple, le nocodazole peut avoir des conséquences dévastatrices pour n’importe quelle cellule. Les inhibiteurs de microtubules photocommutables de manière réversible ont le potentiel de surmonter les limites en permettant aux effets des médicaments d’être mis en œuvre de manière spatio-temporelle contrôlée. L’une de ces familles de médicaments est la photostatine à base d’azobenzène (PST). Ces composés sont inactifs dans des conditions sombres et, lors de l’éclairage par la lumière UV, ils se lient au site de liaison à la colchicine de la β-tubuline et bloquent la polymérisation des microtubules et le renouvellement dynamique. Ici, l’application de PST dans l’embryon de souris préimplantatoire vivant en 3 dimensions (3D) vise à perturber le réseau de microtubules au niveau subcellulaire. Ce protocole fournit des instructions pour la configuration expérimentale, ainsi que des paramètres d’activation et de désactivation de la lumière pour les PST utilisant la microscopie confocale à cellules vivantes. Cela garantit la reproductibilité et permet à d’autres d’appliquer cette procédure à leurs questions de recherche. Des photocommutateurs innovants comme les PST peuvent évoluer en tant qu’outils puissants pour faire progresser la compréhension du réseau dynamique de microtubules intracellulaires et pour manipuler le cytosquelette de manière non invasive en temps réel. En outre, les PST peuvent s’avérer utiles dans d’autres structures 3D telles que les organoïdes, les blastoïdes ou les embryons d’autres espèces.
Introduction
L’architecture des microtubules varie considérablement selon les différents types de cellules pour prendre en charge diverses fonctions 1,2. Sa nature dynamique de croissance et de rétrécissement permet une adaptation rapide aux signaux extra- et intracellulaires et de répondre aux besoins en constante évolution d’une cellule. Par conséquent, il peut être considéré comme « l’empreinte morphologique » jouant un rôle clé dans l’identité cellulaire.
Le ciblage pharmacologique du cytosquelette des microtubules à l’aide d’inhibiteurs de petites molécules a conduit à une pléthore de découvertes fondamentales en biologie du développement, en biologie des cellules souches, en biologie du cancer et en neurobiologie 3,4,5,6,7. Cette approche, bien qu’indispensable, présente diverses limites telles que la toxicité et les effets hors cible. Par exemple, l’un des agents ciblant les microtubules les plus utilisés, le nocodazole, est un puissant médicament dépolymérisant les microtubules8. Cependant, les inhibiteurs de petites molécules tels que le nocodazole sont actifs dès le moment de l’application et, compte tenu de la nature essentielle du cytosquelette de microtubules à de nombreuses fonctions cellulaires critiques, la dépolymérisation globale des microtubules peut produire des effets hors cible, ce qui peut ne pas convenir à de nombreuses applications. De plus, le traitement au nocodazole est irréversible à moins que les échantillons ne soient lavés sans médicament, ce qui empêche l’imagerie continue en direct et, par conséquent, le suivi précis des filaments de microtubules individuels.
Le développement de composés activés par la lumière a commencé avec la création de molécules photoconçues et a annoncé une nouvelle ère dans le ciblage et la surveillance des effets de l’inhibition de la croissance des microtubules de manière précise et contrôlée par spatiotemporalité. Une famille de médicaments photocommutables réversibles, les photostatines (PST), a été développée en remplaçant le composant stilbène de la combretastatine A-4 par de l’azobenzène9. Les PST sont inactifs jusqu’à l’éclairage par lumière UV, la trans-configuration inactive se convertissant en configuration cis active par isomérisation réversible. Les Cis-PST inhibent la polymérisation des microtubules en se liant au site de liaison à la colchicine de la β-tubuline, bloquant son interface avec la β-tubuline et empêchant la dimérisation nécessaire à la croissance desmicrotubules 10. Parmi une cohorte de PST, le PST-1P est apparu comme un composé de plomb car il a la puissance la plus élevée, est entièrement soluble dans l’eau et montre un début rapide de bioactivité après l’illumination.
La trans- à cis-isomérisation la plus efficace des PST se produit à des longueurs d’onde comprises entre 360 et 420 nm, ce qui permet deux options pour l’activation PST. Une ligne laser de 405 nm sur un microscope confocal typique peut être administrée pour un ciblage spatial optimal de l’inhibition de la croissance des microtubules. La capacité de localiser l’emplacement et le moment de l’activation PST grâce à un éclairage laser de 405 nm facilite un contrôle temporel et spatial précis, permettant une perturbation de la dynamique des microtubules au niveau subcellulaire, dans des temps de réponse inférieurs àla seconde 9. Alternativement, une lumière UV LED abordable permet à l’éclairage de l’organisme entier d’induire une perturbation de l’architecture des microtubules à l’échelle de l’organisme. Cela peut être une alternative rentable pour les chercheurs pour qui l’objectif est l’apparition précise de l’inhibition, plutôt que le ciblage spatial. Une autre caractéristique des PST est leur inactivation à la demande en appliquant la lumière verte d’une longueur d’onde comprise entre 510 et 540 nm9. Cela permet de tracer les filaments de microtubules avant, pendant et après l’inhibition de la croissance médiée par PST.
Les PST, bien qu’ils soient encore relativement récents, ont été utilisés dans de nombreuses applications in vitro dans divers domaines de recherche11, y compris l’étude de nouveaux mécanismes de migration cellulaire dans les amiboïdes12, dans les neurones isolés du cerveau de la souris nouveau-née13, et le développement de l’épithélium alaire chez Drosophila melanogaster14 . D’autres médicaments réactifs à la lumière se sont avérés être des outils précieux pour perturber de manière ciblée la fonction cellulaire. Par exemple, un analogue de la blébbistatine, l’azidoblébbistatine, a été utilisé pour améliorer l’inhibition de la myosine sous éclairage15,16. Cela met en évidence le potentiel de nouvelles découvertes en raison de la capacité d’inhibition spatiotemporalement contrôlée de la fonction cellulaire.
Les organismes 3D vivants présentent des systèmes superbes mais plus délicats pour manipuler la dynamique des microtubules au niveau de l’animal entier, d’une seule cellule ou subcellulaire dans des conditions physiologiques. En particulier, l’embryon de souris préimplantatoire offre un aperçu exceptionnel du fonctionnement interne de la cellule ainsi que des relations intercellulaires au sein d’un organisme17. Des cycles consécutifs d’activation et de désactivation des PST ciblés temporellement et spatialement ont contribué à la caractérisation du pont interphasique, une structure post-cytocinétique entre les cellules, en tant que centre d’organisation de microtubules non centrosomiques dans l’embryon de souris préimplantatoire16. Une configuration expérimentale similaire a démontré l’implication de microtubules en croissance dans l’étanchéité de l’embryon de souris pour permettre la formation de blastocystes18. En outre, les PST ont également été utilisés dans des embryons entiers de poisson-zèbre pour étudier la migration des cellules neuronales en inhibant la croissance des microtubules dans un sous-ensemble de cellules du cerveau postérieur19.
Ce protocole décrit la configuration expérimentale et l’utilisation de PST-1P dans l’embryon de souris préimplantatoire. Les instructions présentées ici peuvent également guider l’application des PST pour un large éventail d’objectifs tels que l’étude de la ségrégation chromosomique et de la division cellulaire, le trafic de cargaison intracellulaire et la morphogenèse et la migration cellulaires. En outre, de telles études aideront à la mise en œuvre des PST dans les systèmes organoïdes, les blastoïdes et d’autres modèles embryonnaires tels que Caenorhabditis elegans et Xenopus laevis, ainsi qu’à étendre potentiellement l’utilisation des PST pour les technologies de fécondation in vitro .
Protocole
Les expériences ont été approuvées par le Monash Animal Ethics Committee sous le numéro d’éthique animale 19143. Les animaux ont été logés dans des conditions spécifiques de maison d’animaux exempts d’agents pathogènes à l’animalerie (Monash Animal Research Platform) dans le strict respect des directives éthiques.
1. Collecte d’embryons de souris préimplantatoires
- Superovulez et accouplez les souris comme décrit précédemment16,18, conformément aux directives institutionnelles en matière d’éthique animale.
REMARQUE: Les souches les plus couramment utilisées pour la collecte d’embryons vivants sont les souris C57BL / 6 ou FVB / N. Toutes les données présentées ici ont été générées à l’aide de souris FVB/N. - Le matin après l’accouplement, rincez les zygotes de l’oviducte à l’aide d’un milieu M2 tel que décrit20 ou d’un milieu de liquide tubaire humain (HTF). À l’aide d’un appareil de pipette buccale tel que décrit 21,22, transférer les zygotes dans des gouttelettes fraîches de potassium Simplex Optimised Medium (KSOM), préavertiquées à 37 °C et équilibrées à 5 % de CO2, dans une boîte de culture de 35 mm recouverte d’un volume suffisant d’huile minérale pour assurer une couverture médiatique.
- Microinjectez des zygotes comme décrit20 avec codage d’ARNc pour un microtubule rouge marqué par fluorescence plus marqueur d’extrémité. Ici, l’ARNc pour la protéine de liaison terminale 3 (EB3)-dTomato a été utilisé à une concentration de 30 ng / μL après préparation et purification comme décrit16,18 et dilution dans un tampon de microinjection.
REMARQUE: Il est recommandé de préparer l’ARNc à l’avance et de le conserver à -20 ° C jusqu’à ce que nécessaire. - Cultiver des embryons dans l’obscurité à 37 °C et 5 % de CO2 jusqu’à ce que les embryons aient atteint le stade de développement souhaité pour le traitement PST-1P.
REMARQUE: Pour une ressource complète des temps de culture requis pour différents stades embryonnaires, voir23. Pour les embryons au stade 16 cellules utilisés ici, la culture jusqu’au jour embryonnaire 3 (E3) après la fécondation.
2. Préparation de médicaments et de plats d’imagerie
REMARQUE: Pour les étapes 2.1 à 2.10, travaillez exclusivement dans des conditions sombres ou rouges pour éviter l’activation involontaire de PST-1P. Du papier d’aluminium ou des couvercles sombres doivent être utilisés pour tous les tubes et plats contenant des PST.
- Préparer une concentration de stock de 50 mM PST-1P dans de l’eau ultrapure.
REMARQUE: Le poids moléculaire de PST-1P est de 440 g / mol. La solution mère est stable à -20 °C jusqu’à 1 an. Le PST-1P est soluble dans l’eau ou le tampon aqueux, mais ne se dissout pas facilement dans le DMSO. - À partir de l’étape 2.1, préparer une concentration de travail intermédiaire de 800 μM PST-1P dans de l’eau ultrapure.
- Diluer pst-1P à une concentration finale de 40 μM dans du KSOM frais. Comme une expérience typique nécessite environ 20 μL de KSOM traité au PST-1P, diluer 1 μL de 800 μM PST-1P dans 19 μL de KSOM pour un volume final de 20 μL afin de s’assurer que suffisamment de milieu est préparé à l’avance, en utilisant uniquement de la lumière rouge pour la visibilité.
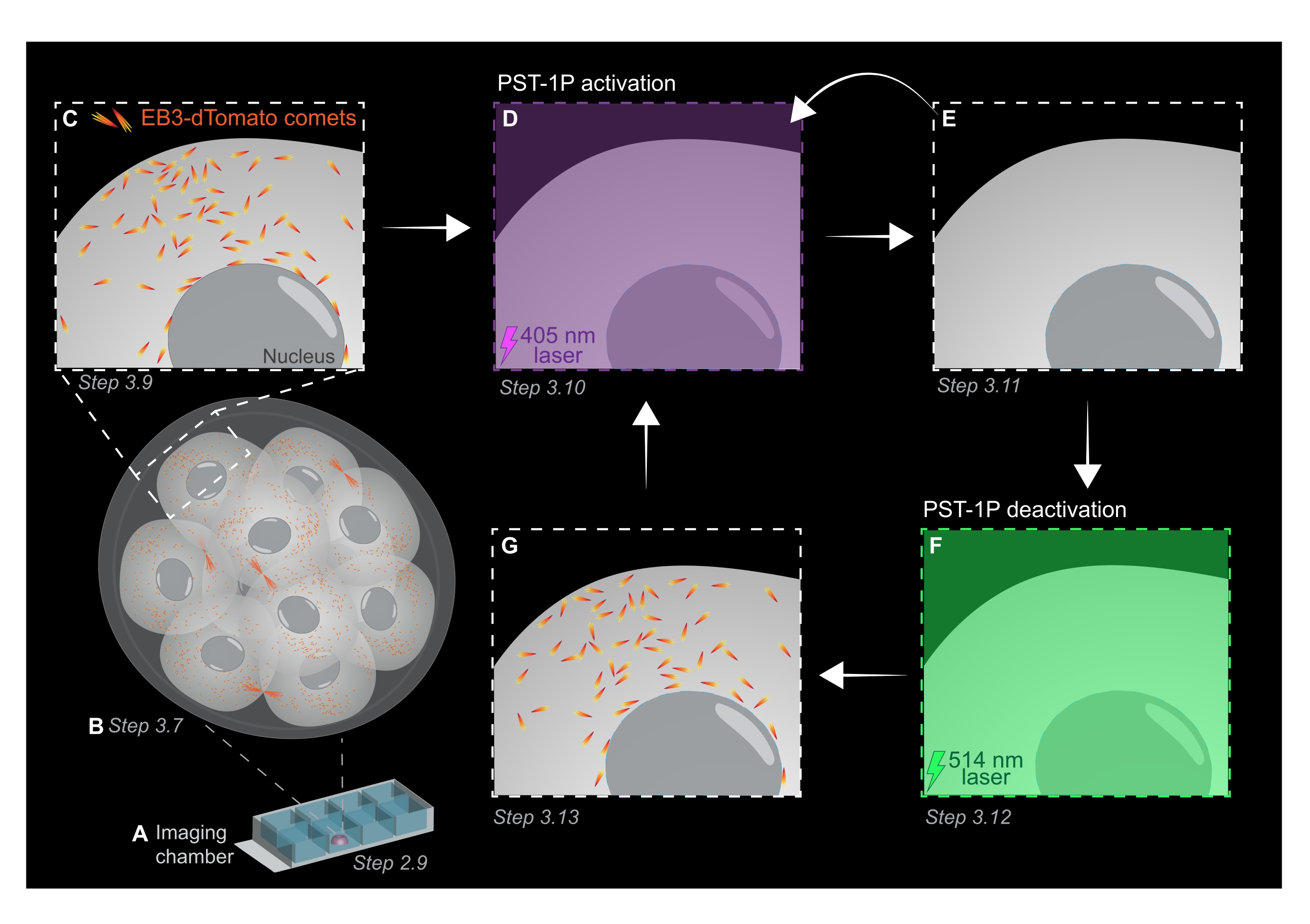
REMARQUE: La concentration et l’état d’activation d’une dilution PST-1P peuvent être vérifiés en prenant un spectre d’absorbance UV-Vis à l’aide d’un spectrophotomètre qui doit être effectué lors de l’établissement du dosage. L’absorbance à 380 nm (A380) d’une dilution entièrement trans de 40 μM dans une cuvette de 1 cm doit être d’environ 0,8. Les formes cis et trans ont la même absorbance à 455 nm (A455). Lorsque le rapport de l’A380 à l’A455 est d’environ 9:1, la dilution est inactive (entièrement trans). Lorsque le rapport A380:A455 est de 1:2, la dilution est complètement activée (entièrement cis). Les ratios intermédiaires reflètent les états intermédiaires d’activation. - Pour préparer la lame de la chambre à l’imagerie en direct, pipettez 10 μL de KSOM traité pst-1P au centre d’un puits pour former une gouttelette hémisphérique (Figure 1A).
- Ajoutez doucement suffisamment d’huile minérale pour couvrir la gouttelette, en veillant à ce qu’elle ne disperse pas le milieu. Cela garantira que la gouttelette ne s’évapore pas.
- Préchauffer et équilibrer le plat coulissant de la chambre dans un incubateur à 37 °C et 5 % de CO2 pendant au moins 3 h ou au plus, pendant la nuit.
- À la fin de la période d’équilibrage, à l’étape 2.6, préparez une boîte de culture de 35 mm avec une gouttelette de 10 μL de KSOM traité au PST-1P, préchauffé et équilibré comme étape de lavage. Ne pas superposer avec de l’huile.
- Transférer les embryons par pipetage buccal dans la gouttelette KSOM traitée par PST-1P à partir de l’étape 2.7.
REMARQUE: Les étapes 2.7 et 2.8 sont recommandées pour le pipetage buccal des embryons, mais sont facultatives. - Transférer immédiatement les embryons par pipetage buccal au centre de la gouttelette KSOM traitée par PST-1P dans la lame de la chambre d’imagerie préparée aux étapes 2.4-2.6 (Figure 1A).
- Incuber des embryons à 37 °C et 5 % de CO2 dans du KSOM traité au PST-1P dans la lame de la chambre d’imagerie pendant au moins 1 h avant l’imagerie. Si possible, montez la lame de chambre sur le microscope à l’intérieur d’une chambre environnementale à 37 °C et 5 % de CO2, et dans l’obscurité totale pour vous assurer que tous les PST-1P sont dans la configuration trans inactive et que les embryons peuvent couler au fond de la boîte.
3. Imagerie en direct et photoactivation PST-1P
REMARQUE: Les étapes 3.1 à 3.13 sont effectuées sur un microscope confocal à balayage laser équipé de détecteurs de photodiodes d’avalanche (APD) et d’une chambre environnementale sombre. Ces instructions se réfèrent spécifiquement à la configuration de l’imagerie à l’aide du logiciel d’acquisition décrit dans la Table des matériaux; cependant, ils peuvent également être appliqués à d’autres systèmes de microscopie confocale.
- Préparez un objectif d’immersion dans l’huile d’eau 63x/1,2 NA avec le milieu d’immersion prescrit.
- À l’aide d’une torche à lumière rouge pour guider le positionnement, avancez l’objectif pour entrer en contact avec le milieu d’immersion. À ce stade, évitez d’utiliser une lumière blanche ou brillante pour trouver les embryons, car cela pourrait activer précocement PST-1P.
- À l’aide de l’oculaire et sous éclairage à la lumière rouge, localisez le bord de la gouttelette de milieu et positionnez l’objectif directement sur cet endroit. Cela peut aider l’utilisateur à établir l’orientation et à trouver le plan focal.
- Ensuite, à travers l’oculaire ou sur la numérisation en mode live activée par logiciel, utilisez un filtre de longueur d’onde rouge ou un laser de 561 nm pour localiser les embryons dans la gouttelette.
- Utilisez des contrôleurs de scène et le mode de balayage en direct pour définir les points de départ et d’arrivée pour l’acquisition d’une pile z de l’embryon entier.
- Ajustez les paramètres de puissance laser (généralement, avec des détecteurs très sensibles tels que les APD, une puissance laser de 561 nm inférieure à 5% est suffisante), le décalage numérique (généralement à -0,900) pour optimiser l’apparence des comètes EB3-dTomato et minimiser le bruit de fond, le sténopé à 2 μm, la résolution des pixels de 512 x 512 et le temps de séjour des pixels de 3,15 μs.
- Acquérir une pile z de l’embryon entier avec des intervalles de section de 1 μm pour évaluer les zones de croissance des microtubules dans l’organisme entier (Figure 1B).
- Utilisez l’image z-stack 3D de l’étape 3.7 pour identifier les régions d’intérêt (ROI) pour les expériences de suivi EB3-dTomato. Augmentez le zoom à 3x et dessinez un retour sur investissement rectangulaire autour de la zone subcellulaire spécifique d’intérêt.
- Acquérir un film time-lapse d’un seul plan z en utilisant les valeurs typiques des paramètres d’imagerie: puissance laser de 561 nm à 5%, décalage numérique de -0,900, résolution en pixels de 512 x 512, sténopé à 3 μm, temps de séjour en pixels de 3,15 μs, zoom de 3x, intervalle de temps de 500 ms.
REMARQUE: 120 délais fourniront un film de suivi de 1 min et devraient être suffisants pour l’analyse des données. L’acquisition peut se poursuivre plus longtemps, à condition que le blanchiment du fluorophore soit minime (Figure 1C). - Pour activer PST-1P, passez à un laser 405 nm et acquérez un autre film time-lapse avec le laser 405 nm réglé sur 10 % de puissance, une résolution de pixels de 512 x 512, un sténopé ouvert au maximum, un temps de séjour en pixels de 3,15 μs, un zoom de 3x, un intervalle de temps de 500 ms et un total de 20 images (Figure 1D).
- Revenez au laser 561 nm et répétez l’acquisition comme à l’étape 3.9 pour confirmer la perte des comètes EB3-dTomato après activation de PST-1P (Figure 1E). Assurez-vous que cette acquisition a lieu le plus tôt possible après l’activation.
REMARQUE: L’étape 3.10 peut être effectuée de manière répétitive pour une inhibition plus longue des comètes EB3-dTomato. Cependant, les embryons doivent être surveillés attentivement pour éviter tout dommage causé par la lumière UV. - Pour inverser le PST-1P à son état trans inactif, engagez un laser 514 nm à 10% de puissance. Obtenez un film en accéléré avec le laser 514 nm réglé sur 10 % de puissance, une résolution en pixels de 512 x 512, un sténopé ouvert au maximum, un temps de séjour en pixels de 3,15 μs, un zoom de 3x, un intervalle de temps de 500 ms et un total de 20 périodes (Figure 1F).
- Répétez l’étape 3.11 pour visualiser la récupération des comètes EB3-dTomato (Figure 1G).
4. Analyse des données d’image
- Pour analyser et quantifier l’inhibition de la polymérisation des microtubules par les PST, utilisez des logiciels à la disposition des chercheurs pour répondre à leurs besoins spécifiques. Ceux recommandés pour une utilisation posséderont un outil de suivi, qui peut suivre manuellement ou automatiquement le mouvement, la direction et la vitesse des comètes EB3-dTomato16,18.

Figure 1 : Représentation schématique de la photoactivation et de la désactivation du PST-1P dans l’embryon de souris préimplantatoire 3D vivant. Toutes les expériences sont réalisées dans l’obscurité totale (fond noir) ou uniquement par éclairage à la lumière rouge. (A) Des embryons de souris préimplantatoires vivants exprimant EB3-dTomato sont cultivés au stade 16 cellules, puis transférés sur une gouttelette de KSOM contenant 40 μM PST-1P dans une lame de chambre d’imagerie. (B) Une image 3D de l’embryon entier permet d’évaluer la croissance des microtubules en visualisant la distribution des comètes EB3-dTomato. (C) Pour commencer l’expérience, les comètes EB3-dTomato sont suivies dans une région subcellulaire à l’aide de l’imagerie time-lapse. (D) La photoactivation ultérieure de PST-1P dans la même région subcellulaire à l’aide d’un laser de 405 nm entraîne la perte de comètes EB3-dTomato (E). L’activation intensifiée du PST-1P peut être mise en œuvre, si nécessaire, par un éclairage lumineux séquentiel de 405 nm. (F-G) Pour inverser PST-1P à son état inactif et restaurer les comètes EB3-dTomato, un laser de 514 nm est appliqué à la même région subcellulaire. Si nécessaire, plusieurs cycles de photoactivation et de désactivation peuvent être effectués. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Résultats
Conformément au protocole, des embryons de souris préimplantatoires ont été microinjectés avec de l’ARNc pour EB3, marqués avec du dTomato fluorescent rouge (EB3-dTomato). Cela permet de visualiser les microtubules en croissance lorsque EB3 se lie au microtubule polymérisant plus se termine24.
Les expériences ont été réalisées 3 jours après la fécondation (E3) lorsque l’embryon de souris est composé de 16 cellules. Tout autre stade de développement p...
Discussion
Le réseau de microtubules fait partie intégrante du fonctionnement interne fondamental d’une cellule. Par conséquent, cela présente des défis dans la manipulation de la dynamique des microtubules dans les organismes vivants, car toute perturbation du réseau a tendance à avoir des conséquences généralisées pour tous les aspects de la fonction cellulaire. L’émergence de composés de ciblage de microtubules photocommutables présente un moyen de manipuler avec précision le cytosquelette au niveau subcellula...
Déclarations de divulgation
Les auteurs ne déclarent aucun intérêt concurrent ou financier.
Remerciements
Les auteurs tiennent à remercier le Dr Oliver Thorn-Seshold et Li Gao de nous avoir fourni des photostatines et des conseils sur la préparation des manuscrits, Monash Production pour le soutien au tournage et Monash Micro Imaging pour le soutien à la microscopie.
Ce travail a été soutenu par la subvention de projet APP2002507 du Conseil national de la santé et de recherches médicales (CRSM) à J.Z. et la bourse Azrieli de l’Institut canadien de recherches avancées (ICRA) à J.Z. L’Australian Regenerative Medicine Institute est soutenu par des subventions du gouvernement de l’État de Victoria et du gouvernement australien.
matériels
| Name | Company | Catalog Number | Comments |
| Aspirator tube | Sigma-Aldrich | A5177 | For mouth aspiration apparatus |
| Chamber slides - LabTek | Thermo Fisher Scientific | NUN155411 | |
| cRNA encoding for EB3-dTomato | N/A | N/A | Prepared according to manufacturers instructions using mMessage in vitro Transcription kit |
| Culture dishes - 35mm | Thermo Fisher Scientific | 150560 | |
| Human chorionic growth hormone | Sigma-Aldrich | C8554 | |
| Human Tubal Fluid (HTF) medium | Cosmo-Bio | CSR-R-B071 | |
| Imaris Image Analysis Software | Bitplane | ||
| Immersion Oil W 2010 | Carl Zeiss | 444969-0000-000 | For use with microscope immersion objective |
| LED torch - Red light | Celestron | 93588 | |
| M2 medium | Sigma-Aldrich | M7167 | |
| Mice - wild-type FVB/N, males and females | N/A | N/A | Females 8-9 weeks old. Males 2-6 months old. |
| Microcapillary Pipettes - Kimble | Sigma-Aldrich | Z543306 | For mouth aspiration apparatus |
| Microinjection buffer | N/A | N/A | 5 mM Tris, 5 mM NaCl, 0.1 mM EDTA, pH 7.4 |
| Mineral oil | Origio | ART-4008-5P | |
| mMessage In vitro Transcription kit | Thermo Fisher Scientific | AM1340 | |
| NanoDrop Spectrophotometer | Thermo Fisher Scientific | ||
| Potassium Simplex Optimised Medium (KSOM) medium | Cosmo-Bio | CSR-R-B074 | |
| Pregnant mare serum gonadotrophin | Prospec Bio | HOR-272 | |
| PST-1P | N/A | N/A | Borowiak, M. et al., Photoswitchable Inhibitors of Microtubule Dynamics Optically Control Mitosis and Cell Death. Cell. 162 (2), 403-411, doi:10.1016/j.cell.2015.06.049, (2015). |
| RNA purification kit | Sangon | B511361-0100 | |
| Ultrapure water | Sigma-Aldrich | W1503 | |
| ZEN Black Software | Carl Zeiss |
Références
- Hawdon, A., Aberkane, A., Zenker, J. Microtubule-dependent subcellular organisation of pluripotent cells. Development. 148 (20), (2021).
- Sanchez, A. D., Feldman, J. L. Microtubule-organizing centers: from the centrosome to non-centrosomal sites. Current Opinion in Cell Biology. 44, 93-101 (2017).
- Galli, M., Morgan, D. O. Cell size determines the strength of the spindle assembly checkpoint during embryonic development. Developmental Cell. 36 (3), 344-352 (2016).
- Vazquez-Diez, C., Paim, L. M. G., FitzHarris, G. Cell-size-independent spindle checkpoint failure underlies chromosome segregation error in mouse embryos. Current Biology. 29 (5), 865-873 (2019).
- Baudoin, J. P., Alvarez, C., Gaspar, P., Metin, C. Nocodazole-induced changes in microtubule dynamics impair the morphology and directionality of migrating medial ganglionic eminence cells. Developmental Neuroscience. 30 (1-3), 132-143 (2008).
- Munz, F., et al. Human mesenchymal stem cells lose their functional properties after paclitaxel treatment. Scientific Reports. 8 (1), 312 (2018).
- Jordan, M. A., Wilson, L. Microtubules as a target for anticancer drugs. Nature Reviews Cancer. 4 (4), 253-265 (2004).
- Vasquez, R. J., Howell, B., Yvon, A. M., Wadsworth, P., Cassimeris, L. Nanomolar concentrations of nocodazole alter microtubule dynamic instability in vivo and in vitro. Molecular Biology of the Cell. 8 (6), 973-985 (1997).
- Borowiak, M., et al. Photoswitchable inhibitors of microtubule dynamics optically control mitosis and cell death. Cell. 162 (2), 403-411 (2015).
- Gaspari, R., Prota, A. E., Bargsten, K., Cavalli, A., Steinmetz, M. O. Structural basis of cis- and trans-Combretastatin binding to tubulin. Chem. 2 (1), 102-113 (2017).
- Thorn-Seshold, O., Meiring, J. Photocontrolling microtubule dynamics with photoswitchable chemical reagents. ChemRxiv. , (2021).
- Kopf, A., et al. Microtubules control cellular shape and coherence in amoeboid migrating cells. Journal of Cell Biology. 219 (6), 201907154 (2020).
- Sawada, M., et al. PlexinD1 signaling controls morphological changes and migration termination in newborn neurons. The EMBO Journal. 37 (4), 97404 (2018).
- Singh, A., et al. Polarized microtubule dynamics directs cell mechanics and coordinates forces during epithelial morphogenesis. Nature Cell Biology. 20 (10), 1126-1133 (2018).
- Kepiro, M., et al. Azidoblebbistatin, a photoreactive myosin inhibitor. Proceedings of the National Academy of Sciences of the United States of America. 109 (24), 9402-9407 (2012).
- Zenker, J., et al. A microtubule-organizing center directing intracellular transport in the early mouse embryo. Science. 357 (6354), 925-928 (2017).
- White, M. D., Zenker, J., Bissiere, S., Plachta, N. Instructions for assembling the early mammalian embryo. Developmental Cell. 45 (6), 667-679 (2018).
- Zenker, J., et al. Expanding actin rings zipper the mouse embryo for blastocyst formation. Cell. 173 (3), 776-791 (2018).
- Theisen, U., et al. Microtubules and motor proteins support zebrafish neuronal migration by directing cargo. Journal of Cell Biology. 219 (10), 201908040 (2020).
- Rulicke, T. Pronuclear microinjection of mouse zygotes. Methods in Molecular Biology. 254, 165-194 (2004).
- Greaney, J., Subramanian, G. N., Ye, Y., Homer, H. Isolation and in vitro culture of mouse oocytes. Bio-protocol. 11 (15), 4104 (2021).
- Subramanian, G. N., et al. Oocytes mount a noncanonical DNA damage response involving APC-Cdh1-mediated proteolysis. Journal of Cell Biology. 219 (4), 201907213 (2020).
- Mihajlovic, A. I., Bruce, A. W. The first cell-fate decision of mouse preimplantation embryo development: integrating cell position and polarity. Open Biology. 7 (11), 170210 (2017).
- Roostalu, J., et al. The speed of GTP hydrolysis determines GTP cap size and controls microtubule stability. Elife. 9, 51992 (2020).
- Gao, L., et al. In vivo photocontrol of microtubule dynamics and integrity, migration and mitosis, by the potent GFP-imaging-compatible photoswitchable reagents SBTubA4P and SBTub2M. BioRxiv. bioRxiv. , (2021).
- Tichy, A. M., Gerrard, E. J., Legrand, J. M. D., Hobbs, R. M., Janovjak, H. Engineering strategy and vector library for the rapid generation of modular light-controlled protein-protein interactions. Journal of Molecular Biology. 431 (17), 3046-3055 (2019).
- van Haren, J., Adachi, L. S., Wittmann, T. Optogenetic control of microtubule dynamics. Methods in Molecular Biology. 2101, 211-234 (2020).
- Adikes, R. C., Hallett, R. A., Saway, B. F., Kuhlman, B., Slep, K. C. Control of microtubule dynamics using an optogenetic microtubule plus end-F-actin cross-linker. Journal of Cell Biology. 217 (2), 779-793 (2018).
- Kogler, A. C., et al. Extremely rapid and reversible optogenetic perturbation of nuclear proteins in living embryos. Developmental Cell. 56 (16), 2348-2363 (2021).
- Maghelli, N., Tolic-Norrelykke, I. M. Laser ablation of the microtubule cytoskeleton: setting up and working with an ablation system. Methods in Molecular Biology. 777, 261-271 (2011).
- Bukhari, S. N. A., Kumar, G. B., Revankar, H. M., Qin, H. L. Development of combretastatins as potent tubulin polymerization inhibitors. Bioorganic Chemistry. 72, 130-147 (2017).
- Gilazieva, Z., Ponomarev, A., Rutland, C., Rizvanov, A., Solovyeva, V. Promising applications of tumor spheroids and organoids for personalized medicine. Cancers (Basel). 12 (10), 2727 (2020).
- Scherer, K. M., et al. Three-dimensional imaging and uptake of the anticancer drug combretastatin in cell spheroids and photoisomerization in gels with multiphoton excitation. Journal of Biomedical Optics. 20 (7), 78003 (2015).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.