
Method Article
כתמים וכימות של היברידיזציה באתרו בתוך דג זברה
In This Article
Summary
טכנולוגיות לעריכת גנים אפשרו לחוקרים ליצור מוטציות דג זברה לחקור פונקציה גנטית בקלות יחסית. כאן, אנו מספקים מדריך לבצע היפוך העובר מקבילי וכימות של אותות היברידיזציה באתרו ב zebrafish. גישה זו לא משוחדת מספק דיוק גדול יותר בניתוחים פנוטיניים המבוססים על היברידיזציה באתרו.
Abstract
היברידיזציה באתרו (ISH) היא טכניקה חשובה המאפשרת לחוקרים ללמוד התפלגות mRNA באתרו והייתה טכניקה קריטית בביולוגיה התפתחותית במשך עשורים. באופן מסורתי, רוב המחקרים ביטוי גנים הסתמך על הערכה חזותית של האות ISH, שיטה הנוטה הטיה, במיוחד במקרים שבהם זהויות לדוגמה ידועים בפריורי. אנו דיווחו בעבר על שיטה לעקוף הטיה זו ולספק כימות מדויקות יותר של אותות ISH. כאן, אנו מציגים מדריך פשוט כדי להחיל שיטה זו כדי לכמת את רמות הביטוי של גנים של עניין עוברים ויטראז ולתאם את זה עם גנוסוגים המתאימים שלהם. השיטה היא שימושית במיוחד כדי לכמת ביטוי מוגבל של גנים מוגבלים בדגימות של גנוטיפים מעורבים והוא מספק חלופה משוחדת ומדויקת לשיטות הניקוד המסורתי ויזואלי.
Introduction
המבוא של טכנולוגיות עריכת הגנום (ZFN, TALENs ולאחרונה, CRISPR/Cas9) הובילה לעלייה מסיבית במספר מעבדות ברחבי העולם העושים שימוש במערכות אלה כדי ללמוד את הפונקציה של גנים ספציפיים vivo. Zebrafish במיוחד הם לקלה מניפולציה גנטית ומוטציות רבות נוצרו בעבר האחרון1,2. עבור ביולוגים התפתחותיים, אחת השיטות הנפוצות ביותר כדי להעריך את ההשלכות הפנותיות של מוטציות גנים בפיתוח עובריים הוא באתרו היברידיזציה (ISH). בהעדר פגמים מורפולוגיים ברורים המפרידים בין מוטציות homozygous מסוג פראי שלהם או heterozygous אחים, זה חיוני כדי לזהות כראוי גנוטיפים שונים במדויק.
ה-ISH הקלאסי מסתמך על ניתוחים איכותניים של עוצמות האות כדי לגזור מסקנות בנוגע לאינטראקציות רגולטוריות בין גן הריבית והגנים המסומנים בסמן. למרות שהיא שימושית, מנתח זה סובל מווריאציה טכנית ועשוי להיות מוטה בציפיות החוקרים. לפיכך, פותחה שיטה לכמת ביטוי גנים לאחר שעברו הדמיה של העוברים ויטראז ', ללא ידע מוקדם של גנוטיפ המקביל. זה היה לאחר הפקת דנ א יעיל והקלדה שאפשרו לנו גנוטיפ כמותית עם ביטוי גנים3. בעוד את ה-גנופיקציה של העוברים פוסט ish כבר בשימוש לפני4,5, התמונה מבוסס הכמת של תבניות ISH לא היה בשימוש נרחב מלבד כמה מחקרים6,7. החלופות הפופולריות ביותר מסתמכות על הניקוד החזותי או על הספירה של תאים מוכתמים בעלי ערך8,9,10, שניהם נוטים להיות מוכתמים ירודים וחוקר הטיה. שיטה זו שימושית במיוחד כדי ללמוד שינויים בגנים עם דפוסי הביטוי כי הם מוגבלים מרחב, כגון runx1 או gata2b, שניהם מבוטא מערכת מוגבלת של תאי הרצפה של אבי העורקים המכונה המוגלובין אנדותל11,12.
כאן, אנו שואפים לספק מדריך מעשי ליישום הקוונפיקציה על ידי ניתוח תמונה באמצעות פיג'י13, כמו גם את הפקת ה-DNA ואת פרוטוקול גנוהקלדה. זה נועד להמחיש חזותית שיטה שפורסמה בעבר שלנו3. השיטה שלנו מאפשרת ייצוג מדויק של הווריאציה ביטוי הגנים שזוהו על ידי ISH ו הקצאה משוחדת של רמות ביטוי גנים לגנוסוגים ספציפיים.
Protocol
הליכים הכרוכים בנושאי בעלי חיים מוסדרים על ידי בעלי החיים (הליכים מדעיים) חוק 1986 ואושר על ידי המשרד הביתי ואת החיים המקומיים רווחה וביקורת אתית גוף.
1. העוברים ויטראז
- הכינו פתרון גליצרול (50% – 80% ב-1x מאגר PBS) וערבבו כדי להגון את התמיסה (למשל, להשאיר ברולר לפחות 5 דקות). פתרון זה ניתן לשמור במשך חודשים בטמפרטורת החדר.
- לאחר באתרו היברידיזציה14,15,16,17, העברת עוברים לפתרון גליצרול עם הפיפטה 3 מ ל ולצאת להתפשר לפחות 5 דקות. אם הדמיה עוברים מעלה 24 hpf (שעות הפריה פוסט), הם עשויים להיות מולבן כמתואר18.
- הכן ותייג מספיק צינורות PCR כדי להעביר עוברים ויטראז ' לאחר הדמיה.
- הוסף 100% גליצרול לחלק התחתון של הבאר בשקופית שקופית של זכוכית עם שלושה מ ל של פסטר.
- שימוש 3 mL פסטר הצינורות, העברת עובר מוכתם איש אחד לשקופית זכוכית האוריינט כנדרש תחת stereomicroscope מצויד עם מצלמה דיגיטלית האור התחתון העליון.
הערה: ג'ל לשימוש הטענת הצינורות למיקום העוברים להדמיה, אך כלים אחרים (למשל, מלקחיים, המחט מבתר) יהיו נאותים באותה מידה. - באמצעות העובר הראשון, כוונן את זמן ההארה והחשיפה בהגדלה הרצויה. השתמש בתנאים אלה עבור כל העוברים באותו ניסוי (כלומר, אם הדמיה 40 עוברים מתוך heterozygous מוטציה incross, לוודא את ההארה, זמן חשיפה ההגדלה זהה עבור כל).
- תמונה כמו עוברים רבים ככל הנדרש. סמן כל תמונה באמצעות מספר ייחודי. לאחר הדמיה, להעביר את העובר לצינור PCR/צלחת מתויג עם אותו מספר.
הערה: יש לשמור תמונות כקבצי TIF, אך גם תבניות אחרות מתאימות.- במקרה הצורך, הסר גליצרול עודפות בצינורות/צלחות ה-PCR.
הערה: בשלב זה ניתן לאחסן את העוברים בצינורות ה-PCR במשך מספר שבועות בטמפרטורת החדר.
- במקרה הצורך, הסר גליצרול עודפות בצינורות/צלחות ה-PCR.
2. לחלץ DNA ו גנוטיפ העוברים ויטראז
הערה: כאן, להשתמש בשיטה אמינה וזולה כדי לבודד דנ א גנומית מבוסס על שיטה המאצ19 עם יעילות חילוץ ה-dna של 95%-100%3.
- לאחר השלמת ההדמיה, הוסף 40-75 μL של מאגר לפירוק אלקליין (למשל, מצליחן) לכל צינורית.
- מודטה ב 95 ° c עבור כ 30 דקות ומצננים את הצינורות עד 4 ° צ' לפני הוספת נפח שווה של ניטרול מאגר. דגירה של לילה ב-4 ° צ' עשויה לשפר את יעילות ה-PCR.
הערה: בשלב זה, ה-DNA של גנומית יכול לשמש עבור הקלדה או מאוחסן ב-20 ° c עד שהוא נדרש. - מדגימות גנוטיפ עם שיטה מתאימה (למשל, hrma, RFLP)3,20,21 כנדרש עבור מוטציה של עניין.
- הערה ה-גנוטיפ המתאים לכל דוגמה (לדוגמה, באמצעות תוכנת גיליון אלקטרוני).
3. מכמת את עוצמת הפיקסל של העוברים ויטראז ' (ניתוח תמונה באמצעות תוכנת פיג'י)
- כדי לכמת את היברידיזציה באתרו (ISH) מכתים את עוצמת האות, להמיר את כל התמונות לגווני אפור 8-bit כמתואר3. אם התמונות נשמרו בתור. TIF קבצים, השתמש במאקרו פיג'י עבור המרת אצווה3. לחילופין, להמיר תמונות בפורמטים אחרים (למשל,. JPG) ל-. טיף באמצעות תוכנה מתאימה ולאחר מכן המרה לגווני אפור של 8 סיביות באמצעות פיג'י. לנוחיותך, הנה ההליך צעד אחר צעד שפורסם בעבר3, עם כמה שינויים.
- פתחו תמונות בפיג והיפוך התמונה בעריכה > היפוך. לאחר מכן שנו את סוג התמונה ל-8 סיביות (תמונה ≫ סוג > 8 סיביות).
- בעזרת כלי בחירת המצולע, ציירו את אזור הריבית (ROI) באופן ידני בתמונה שמסביב לאזור המכיל את האות.
- הקש t כדי לפתוח את מנהל הROI. השתמש בפקודה מדידה של מנהל הroi כדי למדוד את עוצמת ההחזר. העתק את הערך הממוצע מחלון התוצאות לתוכנת גיליון אלקטרוני.
- להעביר את אותו ROI, להבטיח את אותו גודל וצורה כמו האזור המקורי, לאזור של דג שאינו מכיל כתמים כלשהם. חזור על שלב 3.4 כדי למדוד את הרקע.
- כדי להשיג את עוצמת הפיקסל הממוצע של האות ISH, הפחת את ערך העוצמה הממוצע של אזור הרקע מתוך זה של האזור הוכתם עבור כל עובר.
- הקצה כל ערך אינטנסיביות ל-גנוטיפ (משלב 2.3).
4. ניתוח התוצאות במבחנים סטטיסטיים מתאימים
- התווה את כל הערכים בהתוויה אחת של Q-Q כדי לזהות סטיות מהתפלגות רגילה.
הערה: ניתן גם לאמת את ההפצה הרגילה עם מבחן הקולמוגורוב-סמירנוף או מבחן שפירו-וילק. עם זאת, עבור גדלים גדולים במדגם יש סיכון גבוה של תוצאות חיוביות שגויות בבדיקות אלה. - אם קיימות סטיות חזקות מהתפלגות רגילה, הפוך את כל הערכים (באמצעות פונקציות ln או sqrt) כדי לוודא שהם מופצים בדרך כלל לפני שתמשיך.
- לנתח את ההבדלים בין הערכים (שינוי אם הכרחי) מוקצה לכל גנוטיפ (wt נגד הטרוזיציטים נגד מוטציה) עם 2-זנבית ANOVA עם 95% רמות ביטחון, חשבונאות לשוויון של סטיות עם בדיקה של לוונו תיקון וולש. עבור השוואות זיווגים בין שני סוגי גנוטיפים, השתמש בבדיקה של Tukey (סטיות שוות) או משחקים-האוול (שונויות לא שוות).
- אם הערכים לא מופצים בדרך כלל למרות הטרנספורמציה, השתמש במבחן לא פרמטרי (קרוסה-ווליס) כדי לנתח את ההבדלים בין ערכים מדורגים לבין מבחן השוואות מרובות של דאן לאחר הניתוח עם בונפררוני תיקון עבור זיווגים שוואות.
- התווה את הערכים שאינם משתנים (משלב 3.6) כחלקות נקודה עבור הייצוג הטוב ביותר של התוצאות.
תוצאות
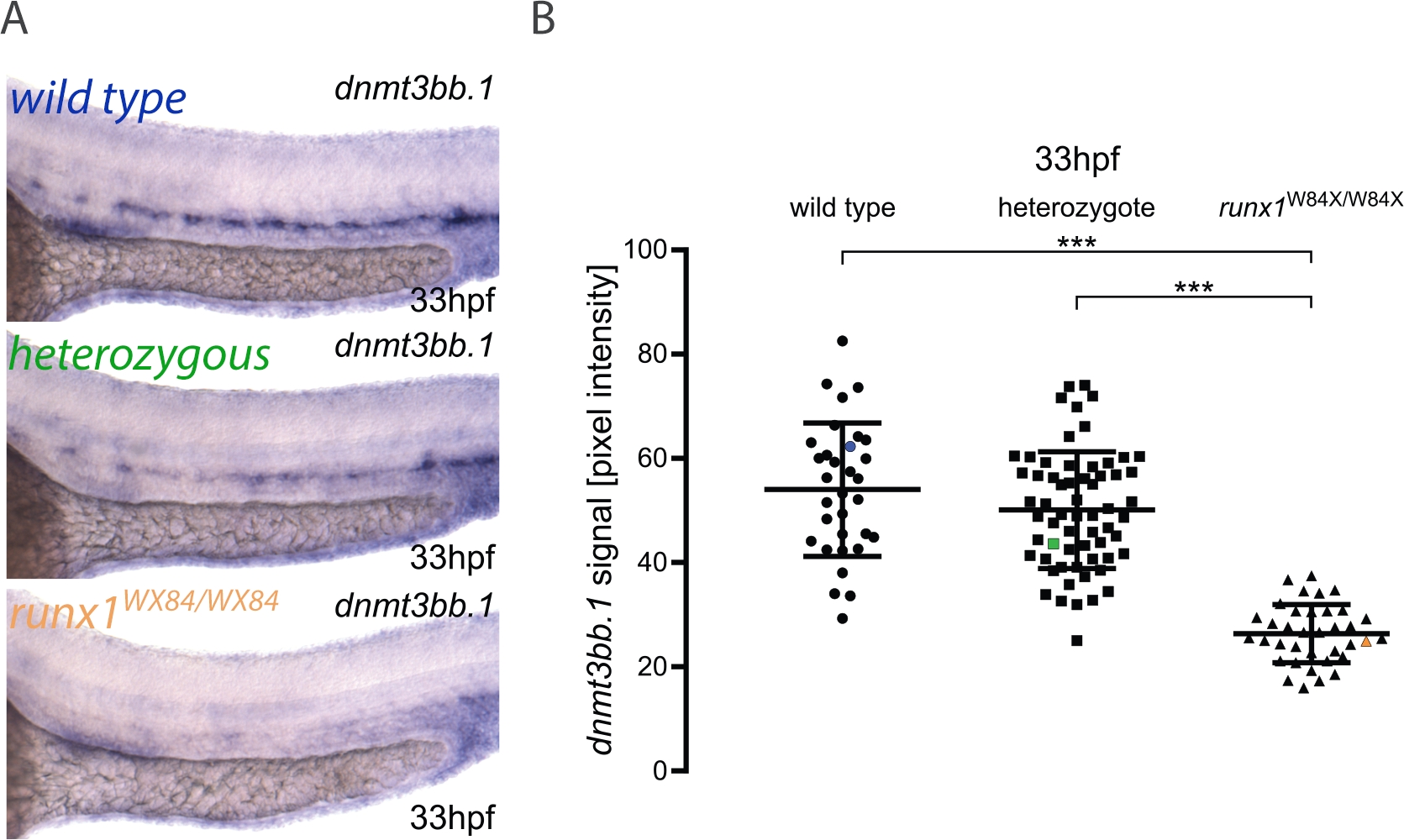
כאן, אנו מתארים את היישום המעשי של צינור עבור כימות התמונה והעובר הקלדה כפי שפורסם במקום אחר3. זרימת העבודה עבור פעולת השירות מוצגת באיור 1. כדי להמחיש כיצד להשתמש בשיטה זו, התבצע ערך עבור dnmt3bb. 1 ב- 33 hpf עוברים מ runx1W84X/+22 incross (איור 2). 130 עוברים שנוצרו באמצעות אותם מצבי תאורה כמפורט בפרוטוקול ומתייג אותם במספר ייחודי. לאחר הדמיה, כל עובר הועבר לצינור ה-PCR לצורך הקלדה. בשלב זה, ניתוח התמונה בוצע כדי לייחס ערך של עוצמת פיקסלים לכל תמונה. ה-גנוטיפ הוקצה לתמונה המתאימה וערכי עוצמת הפיקסלים שקובצו בהתאם ל-גנוטיפ שלהם לניתוח סטטיסטי. ירידה בביטוי dnmt3bb .1 זוהה ב RUNX1W84X/W84X מוטציות (איור 2א, ב)3, בהסכם עם תצפיות קודמות5. מעניין, runx1W84X/+ heterozygous העוברים לא הראו הבדלים משמעותיים dnmt3bb. 1 ביטוי (איור 2א, ב) לעומת אחים סוג פראי שלה, הרומז כי עותק אחד של runx1 מספיק כדי לשמור על dnmt3bb. 1 ביטוי ברמות המתאימות.
רבים מוטציות דג זברה להיכשל להראות פנוטיפ מעובריים שניתן לזהות בדרך אחרת באמצעות אובדן אחר של טכנולוגיות פונקציה כמו ליגוראוסטינו olig, הגאות (MOs). פער זה ניתן לייחס מספר גורמים כולל אפקטים מחוץ ליעד, פיצוי חלבון אימהי, היפואומורפית אלל23 או התופעה שהתגלתה לאחרונה של פיצוי גנטי24,25,26,27. בדוגמה זו, שאלנו אם runx1 ביטוי הופחת או אבד ב lmo4auob100 מוטציות מאז שפורסמו בעבר נתונים באמצעות lmo4a מו הציע כי runx1 הצטמצמה ב lmo4a שמורנים28. כאן, הניתוח לא חשף הבדלים משמעותיים בביטוי runx1 בין סוג פראי ו lmo4auob100 homozygous מוטציות3 (איור 3A, ב). ניתוח נוסף על ידי העובר היחיד qPCR הראה כי היה ירידה קטנה אך משמעותית בביטוי runx1 lmo4auob100 מוטציות (איור 3ג). לכן, ייתכן שקוונפיקציה של התמונה לא תוכל לזהות הבדלים קטנים ברמות הביטוי. לחילופין, חוסר הבדל בין הגנוטיפים שגילינו הוא אמיתי והניסויים qPCR מזהים שינויים runx1 ביטוי ברקמות אחרות כמו telenchephalon שבו הן lmo4a והן runx1 מבוטא. החוקרים צריכים תמיד לאמת את התוצאות שלהם עם שיטה עצמאית כמו qPCR, אבל באופן אידיאלי מעשיר את רקמת העניין על ידי הזרם cy, למשל.
במקרים נדירים שבהם ל-ISH יש רקע גבוה (איור 3D), ערך עוצמת הפיקסלים של אזור זה גבוה כל כך שהחיסור מערך האות מפיק מספר שלילי ובמקרים כאלה העוברים לא ייכללו בניתוח. בניסיון שלנו, זה התרחש כ 0.4% העוברים בנחקר3 אבל עשוי להשתנות בין ניסויים, מרגשים או קבוצות של ריאגנטים. למרות שזה יכול להיות מגבלה של השיטה, התדר הנמוך של הרקע הגבוה הוא לא סביר מאוד להשפיע על התוצאות הכלליות.
כדי לבדוק את ההשפעה של בחירת אזורים שונים לתיקוני רקע, אנו מדדו לראשונה את עוצמת הפיקסל של האות runx1 ISH ב -28 העוברים השונים, באמצעות אזורים שונים לתיקוני רקע (איור 4). ארבעה אזורים שונים נבחרו: שניים באזור הגזע (R1, ו-R2), אחד באזור החלמון (בלתי מוכתם, אבל סביר לצבור כתמים ברקע) ואזור קטן יותר הקדמי של התשואה (אר סי, איור 4ב). מדידת עוצמת הפיקסלים באזורים אלה הראתה הבדל יציב יחסית באינטנסיביות בין התשואה לאזור הרקע (איור 4ג). עם זאת, R3 תמיד הראו ערכים גבוהים מאוד (מעל אלה ב-ROI). לאחר היפוך והמרה ל-8-bit, אזור החלמון נראה בהיר מאוד ולכן אינו מתאים לשימוש כתיקון רקע. R2 היה קרוב יותר התשואה אבל הכיל כמה אות ISH, ושימוש בו עבור תיקון ירד את עוצמת פיקסל ממוצע בהשוואה או R1 (ממוקם דורסלי נוספת, הרחק מהאות ISH) או אר אר. כך, או R1 או אר אר הם האזורים המתאימים שניתן להשתמש בהם עבור תיקון רקע (למרות השטח של אר 4 להיות קטן יותר מזה של R1). הבא, ביקשו להשוות את השימוש R1 או אר 4 השפיעו על התוצאות בעת השוואת runx1 expression. בשביל זה, אנחנו לחצות dll4+/- הטרוזיטים29 וניתח runx1 ביטוי באקראי סוג פראי נבחר ו Dll4-/-עוברים (איור 4E). למרות שהשימוש ב-R1 או בארה ב לתיקון רקע מושפע מערכים בודדים, עוצמות הפיקסלים הממוצע באותו גנוטיפ לא היו שונות באופן משמעותי (איור 4E). יתר על כן, השוואת runx1 ביטוי עדיין מניב ערכי עוצמה ממוצע דומה בין גנוטיפים באמצעות האזורים R1 או 4 כתיקון רקע (μR1= 16.3 ו-μ4= 18.2, בהתאמה). יחד, הגענו למסקנה כי למרות הבחירה של אזור הרקע חשוב, הקריטריונים העיקריים היא כי הוא אינו כולל אזורי חלמון (מועדים להצטברות של כתמים ברקע) וכי הוא לא צריך להכיל כל (ספציפי) מכתים שעשוי להטות את ערכי עוצמת הפיקסל של הרקע.

איור 1: זרימת העבודה של כימות התמונה המקבילית ופרוטוקול ההקלדה. עוברים שנאספו מתוך incross של דגים heterozygous עבור אלל מוטציה נחקר עבור הגן הנמדד עם פרוטוקול ISH סטנדרטי. לאחר הדמיה, דנ א גנומית מופק באמצעות פרוטוקול מצליחן על ידי הוספת מאגר הליזה ישירות על העובר בצינור 0.2 mL PCR, ואחריו 30 דקות דגירה ב 95 ° c. ה-DNA הזה משמש להקלדה של העוברים על ידי ה-pcr, pcr ו הגבלת אורך של קטע ההגבלה (RFLP), kasp בחני או כל שיטה מתאימה אחרת. במקביל, התמונות עבור כל עובר הן הפוכות ומומרות לסולם אפור של 8 סיביות. ROIs של צורה וגודל זהים המכילים את האות ISH (צהוב) והרקע (כחול) נבחרים ונמדדים באופן ידני. , המידות, המוקצות לגנוטיפים מתאימים. מנותחות מבחינה סטטיסטית האיור הותאם מ Dobrzycki et al.3אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: כימות התמונה ב-runx1 מוטציות חושף רמות מופחתת של dnmt3bb. 1 ביטוי על ידי ISH. (A) לדוגמה תמונות של ISH ב-33 hpf סוג פראי (כחול), runx1+/w84 x (ירוק) ו runx1W84X/W84X (כתום) עוברים, מציג dnmt3bb. 1 ביטוי בראש העורקים הראש. (ב) ערכי עוצמת פיקסלים של dnmt3bb. 1 mrna ב runx1W84X/W84Xעוברים (n = 36) ירדו באופן משמעותי בהשוואה לסוגי בר (n = 32) ו הטרוזיטים (n = 62) (ANOVA, p < 0.001). המקדמים של וריאציה הם 24%, 22% ו -21% עבור סוג פראי, הטרוזיציטים וקבוצות מוטציה, בהתאמה. נקודת נתונים כחול, ירוק וכתום מתאימות לתמונות לדוגמה מהחלונית A. הסורגים מייצגים ממוצע ± ש גויטיין * * *p< 0.001 (משחקים-האוול לאחר הבדיקה הפוסט-הוק). האיור הותאם מ Dobrzycki et al.3אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: מדידת רמות ביטוי runx1 על ידי ISH ב lmo4auob100מוטציות. (א) דמויות מייצגות של ISH עבור runx1 בסוג של 28 hpf פראי (כחול), heterozygous (ירוק) ו lmo4auob100/uob100 (כתום) עוברים, מציג את הביטוי בראש העורקים הראש. (ב) כימות האות runx1 mrna, זיהה על ידי ISH, מן 28 hpf פראי סוג (n = 15), heterozygous lmo4a+/- (הט) (n = 34) ו lmo4auob100/uob100 מוטציה (n = 18) עוברים ממצמד אחד מראה לא הבדל משמעותי בעוצמת פיקסל runx1 בין גנוסוגים שונים (ANOVA,> p 0.6) נקודת נתונים כחול, ירוק וכתום מתאימות לתמונות לדוגמה מהחלונית A. הסורגים מייצגים ממוצע ± ש גויטיין (C) boxplots הצגת רמות runx1 mrna מנורמל (2-ΔCt) בסוג פראי יחיד (כחול; n = 12) ו lmo4auob100/uob100 (מוט, כתום; n = 12) עוברים, שנמדדו על ידי רביעיית ה-PCR, מראה רמות נמוכות של runx1 במוטציות בהשוואה לסוג פראי. *p < 0.05 (t מבחן). (ד) דוגמה לניסוי ISH על העובר 28 (מוכתם בrunx1, ראשי חץ צהובים) המראה רקע גבוה. האיור הותאם מ Dobrzycki et al.3אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: ההשפעה של תיקון עוצמת הרקע על תוצאות המדידה. (א) דמות מייצגת של runx1 ISH כתמים ב העובר סוג פראי ב 28 hpf. (ב) אותה תמונה לאחר היפוך והמרה ל-8 סיביות. אזור הריבית (ROI) מודגש בירוק ובארבעה אזורים שונים המשמשים לתיקון רקע (R1-פור) מודגשים בצהוב. (ג) מדידות של עוצמת פיקסלים גולמיים בכל האזורים המוצגים בלוח B. הערה העוצמה ב-R3 (חלמון) היא גבוהה באופן עקבי מאשר האות ISH בפועל של ההחזר (n = 11). (D) Runx1 ביטויים בתוך ההחזר באמצעות R1, R2 ו-4 אזורי רקע. אזורים עבור ROI, R1, R2 ו R3 ~ 28500 פיקסלים; . אר. 8500 פיקסלים שים לב כי R3 רקע לא נעשה שימוש עבור השוואה זו כתיקון הרקע (ROI-R3) בעקביות הניב ערכים שליליים. (ה) Runx1 רמות ביטויים בסוג פראי ו- dll4-/- מוטציות באמצעות R1 או 4 עבור תיקון הרקע (n = 10 עבור כל דוגמה). ניתוח סטטיסטי בלוחות D ו-E בוצע באמצעות מבחן מחקר מקרוקאל-ווליס, בהנחה כי ערכי עוצמת הפיקסל לא מופצים בדרך כלל. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Discussion
יש לשקול מספר גורמים כאשר משתמשים בשיטה זו לכמת את הביטוי הגנטי. יש לשמור על תנאי ההדמיה במהלך הניסוי (לדוגמה, תאורה, זמני חשיפה ומיצוב העובר) כדי לצמצם את השונות בין המידות. נקודה קריטית היא להימנע מצביעת הדגימות, מכיוון שהבדלים בין הדגימות עשויים להיות רעולי פנים. למשל, הירידה בביטוי של ולפה בהעדר Eto2 ב קסנפוס זריזה העוברים30 ניתן להבחין רק על ידי כתמים ניטור בזהירות על פני תקופה של 24 שעות. לכן, זה אימון טוב כדי מדעית לקבוע רמות מכתים הולם עבור כל הגן הטוב ביותר לייצג את הביטוי שלה, מבלי להגיע לרוויה. כתמים מיותר גם להגדיל באופן מלאכותי את עוצמת פיקסלים ברקע בתמונות בגווני אפור שהומרו 8-bit ולהטות את תוצאות הקוונרה. במקרים קיצוניים, רמת הרקע ברקמות עובריים עשויה להיות גבוהה יותר מאשר האות ISH ב-ROI שנבחר ואין לכלול בדוגמאות אלה מהניתוח. תופעה דומה נצפתה כאשר בדקנו את התאמתו של החלמון הבלתי מוכתם לתיקון רקע (איור 4). לאחר היפוך והמרה של 8-bit פיקסלים כהים באזור החלמון להיות בהירים יותר מאשר אות ISH בעובר ולהפוך את הרקע מתוקן ערכים שליליים. כך, למנוע את השימוש בחלמון לתיקון רקע. מדידת אות הרקע באזורי פיגמנט בעובר (למשל, את העיניים או את החלק המאחור של הגזע מ 26/28 hpf ואילך) יהיה להטות באופן שווה את תוצאות כימות וצריך גם להימנע. ישנם פרוטוקולים זמינים עבור עוברי הלבנת דגים, או לפני או אחרי ISH18 ו עוברים הלבנת מעלה 24 hpf לפני מומלץ הדמיה.
מאחר ששיטה זו מסתמכת על מדידת עוצמת הפיקסלים באזור מוגדר כנגד עוצמת פיקסלים ברקע באזור שאינו מוכתם באופן שווה, אין זה מתאים לכמת של גנים מסוימים או כמעט בלתי-מוגדרים. במקום זאת, הוא מתאים היטב למדידת ביטוי של גנים עם התפלגות מוגבלת מרחב שבו אזור למדידת עוצמת פיקסל ברקע ניתן לזהות בקלות. הניתוח הנוסף שלנו מרמז כעת כי באמצעות שטח קטן יותר (3-4x קטן יותר) עבור תיקון רקע תשואות תוצאות דומות לשימוש באזור שווה ערך לזה של ההחזר. זה מרחיב את הישימות של השיטה לגנים הביע תחומים רחבים יותר (ובכך דורשים ROIs גדול יותר עבור מדידות אינטנסיביות), כל עוד אפשר להשתמש באזורים ברורים בלתי מוכתמים של העובר לתיקון הרקע.
בסופו של דבר, אנו מציעים שהגנוטיפים יבוצעו במקביל או לאחר כימות התמונה כדי למזער את הטיית הניסויים. בקשת ניסויים שנייה כדי לחזור על הפחתת הדגימה על דגימות אנניטיות ולהשוות עם הערכה הראשונה של המידות תסייע גם להפחית את ההטיה באמצעות מזהה. אם התמונות שניתן לכמת הן מהשוואה בין טיפולים שאינם דורשים גנוטיפ (למשל, סוג פראי לעומת מעכב כימי או סוג פראי לעומת MO הסתרה), הניסויים לבצע את המידות צריך לעוור את זהותו של ה דוגמה.
Disclosures
המחברים מצהירים שאין להם אינטרסים פיננסיים מתחרים.
Acknowledgements
ברצוני להודות לצוות השירותים הביו-רפואיים באוקספורד וברמינגהאם עבור גידול דגים מעולה. T.D. היה ממומן על ידי כרומוזום אמון ברוכים ה#WT102345 בביולוגיה התפתחותית מלגה לדוקטורט (בשנת המאה השלוש/13/Z). עקב חומרי גלם ו-מ. מ. מומן על ידי קרן הלב הבריטית (BHF IBSR מלגת/13/50/30436) ומודים על תמיכתם הנדיבה. עקב חומרי גלם מכיר תמיכה ממרכז BHF של מצוינות מחקר (RE/13/1/30181), אוקספורד.
Materials
| Name | Company | Catalog Number | Comments |
| 0.2 mL PCR tubes (8-strips with lids) | StarLab | A1402-3700 | 96-well plates are equally appropriate for sample handling but beware of cross contamination between samples |
| 3 mL Pasteur pipettes | Alpha Laboratories | LW4114 | |
| Cavity slides | Brand | BR475535-50EA | |
| Digital Camera (Qimaging Micropublisher 5.0) | Qimaging | ||
| Eppendorf Microloader tips | Eppendorf | 10289651 | the tips are used to orient the embryos for imaging in glycerol |
| Excel | Microsoft | ||
| F3000 Fiber Optic Cold Light Source | Photonic | ||
| Fiji | |||
| Glycerol | Sigma | G5516-1L | |
| Graphpad Prism 8.01 | GraphPad Software, Inc. | we prefer to use Graphpad, but other statistics software packages are also suitable (e.g. SigmaPlot or SPSS) | |
| HotSHOT alkaline lysis buffer | 25 mM NaOH, 0.2 mM disodium EDTA, pH 12 | ||
| HotSHOT neutralization buffer | Tris HCl 40 mM, pH 5 | ||
| PBS (10X) pH 7.4 | Thermofisher | 70011044 | |
| Stereomicroscope with illumination stand (Nikon SMZ800N) | Nikon | ||
| Thermocycler | Thermofisher |
References
- Varshney, G. K., et al. High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome Research. 25 (7), 1030-1042 (2015).
- Varshney, G. K., et al. CRISPRz: a database of zebrafish validated sgRNAs. Nucleic Acids Research. 44 (D1), D822-D826 (2016).
- Dobrzycki, T., Krecsmarik, M., Bonkhofer, F., Patient, R., Monteiro, R. An optimised pipeline for parallel image-based quantification of gene expression and genotyping after in situ hybridisation. Biology Open. 7 (4), bio031096 (2018).
- Bresciani, E., et al. CBFbeta and RUNX1 are required at 2 different steps during the development of hematopoietic stem cells in zebrafish. Blood. 124 (1), 70-78 (2014).
- Gore, A. V., et al. Epigenetic regulation of hematopoiesis by DNA methylation. Elife. 5, e11813 (2016).
- Fan, Y., et al. Tissue-Specific Gain of RTK Signalling Uncovers Selective Cell Vulnerability during Embryogenesis. PLoS Genetics. 11 (9), e1005533 (2015).
- Wen, B., et al. GATA5 SUMOylation is indispensable for zebrafish cardiac development. Biochimica et Biophysica Acta. 1861 (7), 1691-1701 (2017).
- Espin-Palazon, R., et al. Proinflammatory signaling regulates hematopoietic stem cell emergence. Cell. 159 (5), 1070-1085 (2014).
- Peterkin, T., Gibson, A., Patient, R. Redundancy and evolution of GATA factor requirements in development of the myocardium. Developmental Biology. 311 (2), 623-635 (2007).
- Genthe, J. R., Clements, W. K. R-spondin 1 is required for specification of hematopoietic stem cells through Wnt16 and Vegfa signaling pathways. Development. 144 (4), 590-600 (2017).
- Kalev-Zylinska, M. L., et al. Runx1 is required for zebrafish blood and vessel development and expression of a human RUNX1-CBF2T1 transgene advances a model for studies of leukemogenesis. Development. 129 (8), 2015-2030 (2002).
- Butko, E., et al. Gata2b is a restricted early regulator of hemogenic endothelium in the zebrafish embryo. Development. 142 (6), 1050-1061 (2015).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Lleras Forero, L., et al. Segmentation of the zebrafish axial skeleton relies on notochord sheath cells and not on the segmentation clock. Elife. 7, (2018).
- Jowett, T., Yan, Y. L. Double fluorescent in situ hybridization to zebrafish embryos. Trends in Genetics. 12 (10), 387-389 (1996).
- Thisse, C., Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nature Protocols. 3 (1), 59-69 (2008).
- Narayanan, R., Oates, A. C. Detection of mRNA by Whole Mount in situ Hybridization and DNA Extraction for Genotyping of Zebrafish Embryos. Bio-protocol. , e3193 (2019).
- Monteiro, R., Pouget, C., Patient, R. The gata1/pu.1 lineage fate paradigm varies between blood populations and is modulated by tif1gamma. EMBO JOURNAL. 30 (6), 1093-1103 (2011).
- Truett, G. E., et al. Preparation of PCR-quality mouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). Biotechniques. 29 (1), 52-54 (2000).
- Wilkinson, R. N., Elworthy, S., Ingham, P. W., van Eeden, F. J. A method for high-throughput PCR-based genotyping of larval zebrafish tail biopsies. Biotechniques. 55 (6), 314-316 (2013).
- Parant, J. M., George, S. A., Pryor, R., Wittwer, C. T., Yost, H. J. A rapid and efficient method of genotyping zebrafish mutants. Developmental Dynamics. 238 (12), 3168-3174 (2009).
- Jin, H., et al. Definitive hematopoietic stem/progenitor cells manifest distinct differentiation output in the zebrafish VDA and PBI. Developement. 136 (4), 647-654 (2009).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. 13 (10), e1007000 (2017).
- El-Brolosy, M. A., Stainier, D. Y. R. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genetics. 13 (7), e1006780 (2017).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. 524 (7564), 230-233 (2015).
- Ma, Z., et al. PTC-bearing mRNA elicits a genetic compensation response via Upf3a and COMPASS components. Nature. 568 (7751), 259-263 (2019).
- El-Brolosy, M. A., et al. Genetic compensation triggered by mutant mRNA degradation. Nature. 568 (7751), 193-197 (2019).
- Meier, N., et al. Novel binding partners of Ldb1 are required for haematopoietic development. Development. 133 (24), 4913-4923 (2006).
- Kettleborough, R. N., et al. A systematic genome-wide analysis of zebrafish protein-coding gene function. Nature. 496 (7446), 494-497 (2013).
- Leung, A., et al. Uncoupling VEGFA functions in arteriogenesis and hematopoietic stem cell specification. Developmental Cell. 24 (2), 144-158 (2013).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved