
Method Article
Genotipagem e Quantificação da Coloração de Hibridização in Situ em Zebrafish
Neste Artigo
Resumo
As tecnologias de edição de genes permitiram que os pesquisadores gerassem mutantes de zebrafish para investigar a função genética com relativa facilidade. Aqui, fornecemos um guia para realizar genotipagem e quantificação de embriões paralelos em sinais de hibridização in situ em zebrafish. Essa abordagem imparcial proporciona maior precisão em análises fenotípicas baseadas na hibridização do situ.
Resumo
No situ hybridization (ISH) é uma técnica importante que permite aos pesquisadores estudar distribuição de mRNA in situ e tem sido uma técnica crítica em biologia do desenvolvimento há décadas. Tradicionalmente, a maioria dos estudos de expressão genética dependia da avaliação visual do sinal ISH, um método que é propenso a viés, particularmente nos casos em que as identidades amostrais são conhecidas como priori. Já reportamos anteriormente um método para contornar esse viés e fornecer uma quantificação mais precisa dos sinais ISH. Aqui, apresentamos um guia simples para aplicar este método para quantificar os níveis de expressão de genes de interesse em embriões manchados de ISH e correlacionar isso com seus genótipos correspondentes. O método é particularmente útil para quantificar sinais de expressão genética espacialmente restritos em amostras de genótipos mistos e fornece uma alternativa imparcial e precisa aos métodos tradicionais de pontuação visual.
Introdução
A introdução de tecnologias de edição de genomas (ZFN, TALENs e, mais recentemente, CRISPR/Cas9) levou a um aumento maciço no número de laboratórios em todo o mundo que fazem uso desses sistemas para estudar a função de genes específicos in vivo. Zebrafish, em particular, são favoráveis à manipulação genética e muitos mutantes foram gerados nos últimos1,2. Para biólogos de desenvolvimento, um dos métodos mais comuns para avaliar as consequências fenotípicas das mutações genéticas no desenvolvimento embrionário está na hibridização situ (ISH). Na ausência de defeitos morfológicos óbvios que separam mutantes homozigosos de seu tipo selvagem ou irmãos heterozigosos, é essencial ser capaz de identificar corretamente diferentes genótipos com precisão.
O ISH clássico conta com análises qualitativas de intensidades de sinal para obter conclusões sobre interações regulatórias entre o gene de interesse e genes marcadores selecionados. Embora úteis, essas análises sofrem de variação técnica e podem ser tendenciosas pelas expectativas dos pesquisadores. Assim, um método foi desenvolvido para quantificar a expressão genética após a imagem de embriões manchados de ISH, sem conhecimento prévio do genótipo correspondente. Isso foi seguido por uma eficiente extração de DNA e genotipagem que nos permitiu correlacionar quantitativamente genótipo com expressão genética3. Embora a genotipagem de embriões pós ISH tenha sido usada antes de4,5, quantificação baseada em imagem de padrões ISH não tem sido amplamente utilizada além de um par de estudos6,7. As alternativas mais populares dependem da pontuação visual ou da contagem de células manchadas de ISH8,9,10,ambas propensas à má reprodutibilidade e viés de pesquisador. Este método é particularmente útil para estudar mudanças em genes com padrões de expressão que são espacialmente restritos, como runx1 ou gata2b, ambos expressos em um subconjunto restrito de células do piso aórtico chamada endotelio hematogênico11,12.
Aqui, nosso objetivo é fornecer um guia prático para a implementação da quantificação por análise de imagem utilizando Fiji13, bem como o protocolo de extração de DNA e genotipagem. Isso é feito para ilustrar visualmente nosso método3publicado anteriormente. Nosso método permite uma representação precisa da variação na expressão genética detectada pelo ISH e uma atribuição imparcial dos níveis de expressão genética a genótipos específicos.
Protocolo
Os procedimentos envolvendo os sujeitos animais são regulamentados pela Lei de Animais (Procedimentos Científicos) de 1986 e foram aprovados pelo Home Office e pelo Órgão local de Bem-Estar Animal e Revisão Ética.
1. Embriões manchados de imagem ISH
- Prepare uma solução de glicerol (50%-80% no buffer 1x PBS) e misture para homogeneizar a solução (por exemplo, deixe em um rolo por pelo menos 5 min). Esta solução pode ser mantida por meses em temperatura ambiente.
- Depois da hibridização situ14,15,16,17, transferir embriões para a solução glicerol com uma pipeta pasteur de 3 mL e sair para se contentar com pelo menos 5 min. Se embriões de imagem com mais de 24 hpf (horas após a fertilização), eles podem ser branqueados conforme descrito18.
- Prepare e rotule tubos PCR suficientes para transferir embriões manchados de ISH após a imagem.
- Adicione 100% de glicerol ao fundo do poço em um slide de depressão de vidro com uma pipeta pasteur de 3 mL.
- Usando uma pipeta pasteur de 3 mL, transfira um único embrião manchado de ISH para o slide de vidro e oriente conforme necessário um estereótipo equipado com uma câmera digital e iluminação inferior e superior.
NOTA: Use as pontas de pipeta de carregamento de gel para posicionar os embriões para imagem, mas outras ferramentas (por exemplo, fórceps, agulha dissecando) serão igualmente adequadas. - Usando o primeiro embrião, ajuste o tempo de iluminação e exposição na ampliação desejada. Use essas condições para TODOS os embriões no mesmo experimento (ou seja, se a imagem de 40 embriões de uma incross mutante heterozigosa, certifique-se de que a iluminação, o tempo de exposição e a ampliação são os mesmos para todos).
- Imagem quantos embriões necessários. Rotule cada imagem com um número único. Após a imagem, transfira o embrião para um tubo/placa PCR rotulado com o mesmo número.
NOTA: As imagens devem ser salvas como arquivos TIF, mas outros formatos também são adequados.- Se necessário, remova o excesso de glicerol nos tubos/placas PCR.
NOTA: Neste ponto, os embriões podem ser armazenados nos tubos PCR por várias semanas à temperatura ambiente.
- Se necessário, remova o excesso de glicerol nos tubos/placas PCR.
2. Extrair DNA e genótipo dos embriões manchados de ISH
NOTA: Aqui, use um método confiável e barato para isolar o DNA genômico baseado no método HoTSHOT19 com uma eficiência de extração de DNA de 95%-100%3.
- Após a imagem estiver completa, adicione 40-75 μL de tampão de lise alcalina (por exemplo, HoTSHOT) a cada tubo.
- Incuba r$ 95 °C por aproximadamente 30 min e esfrie os tubos a 4 °C antes de adicionar um volume igual de tampão de neutralização. Uma incubação noturna a 4 °C pode melhorar a eficiência do PCR.
NOTA: Neste ponto, o DNA genômico pode ser usado para genotipagem ou armazenado a -20 °C até que seja necessário. - Amostras de genótipo com método adequado (por exemplo, HRMA, RFLP)3,20,21 conforme necessário para a mutação de interesse.
- Observe o genótipo correspondente a cada amostra (por exemplo, usando um software de planilha).
3. Quantificar a intensidade dos pixels de embriões manchados de ISH (análise de imagem usando software Fiji)
- Para quantificar a intensidade do sinal de coloração in situ (ISH), converta todas as imagens em cinza de 8 bits conforme descrito3. Se as imagens fossem salvas como. Arquivos TIF, use uma macro Fiji para conversão de lote3. Alternativamente, converta imagens em outros formatos (por exemplo, . JPG) para . TIF usando software apropriado e converta-se em escala cinza de 8 bits usando Fiji. Por conveniência, aqui está o procedimento passo a passo que foi publicado anteriormente3, com algumas alterações.
- Abra imagens em Fiji e inverta a imagem com Edit > Invert. Em seguida, mude o tipo de imagem para 8 bits (Imagem > Type > 8 bits).
- Utilizando a ferramenta de seleção de polígonos, desenhe a região de interesse (ROI) manualmente na imagem ao redor da região contendo o sinal.
- Pressione para abrir o gerente do ROI. Use o comando de Medida do gerente do ROI para medir a intensidade do ROI. Copie o valor médio da janela resultados para um software de planilha.
- Mova o mesmo ROI, garantindo o mesmo tamanho e forma da região original, para uma região do zebrafish que não contém nenhuma coloração. Repita o passo 3.4 para medir o fundo.
- Para obter a intensidade média dos pixels do sinal ISH, subtraia o valor médio de intensidade da região de fundo da região manchada para cada embrião.
- Atribuir cada valor de intensidade a um genótipo (a partir da etapa 2.3).
4. Analisar os resultados com testes estatísticos apropriados
- Traçar todos os valores em um gráfico Q-Q para identificar quaisquer desvios da distribuição normal.
NOTA: A distribuição normal também pode ser verificada com o teste Kolmogorov-Smirnov ou o teste Shapiro-Wilk. No entanto, para grandes tamanhos amostrais há um alto risco de falsos positivos nestes testes. - Se houver fortes desvios da distribuição normal, transforme todos os valores (usando funções de LN ou sqrt) para garantir que eles sejam normalmente distribuídos antes de prosseguir.
- Analise as diferenças entre os valores (transformados, se necessário) atribuídos a cada genótipo (wt vs. heterozygote vs. mutante) com ANOVA de 2 caudas com 95% de níveis de confiança, representando a igualdade de variâncias com um teste de Levene e correção de Welch. Para comparações parwise entre cada par de genótipos, use o teste pós-hoc de Tukey (variações iguais) ou Games-Howell (variâncias desiguais).
- Se os valores não forem normalmente distribuídos apesar da transformação, use um teste não paramétrico (Kruskall-Wallis) para analisar as diferenças entre valores classificados e um teste de comparações múltiplas pós-hoc Dunn com correção de Bonferroni para parwise Comparações.
- Traçar os valores não transformados (a partir da etapa 3.6) como parcelas de ponto para a melhor representação dos resultados.
Resultados
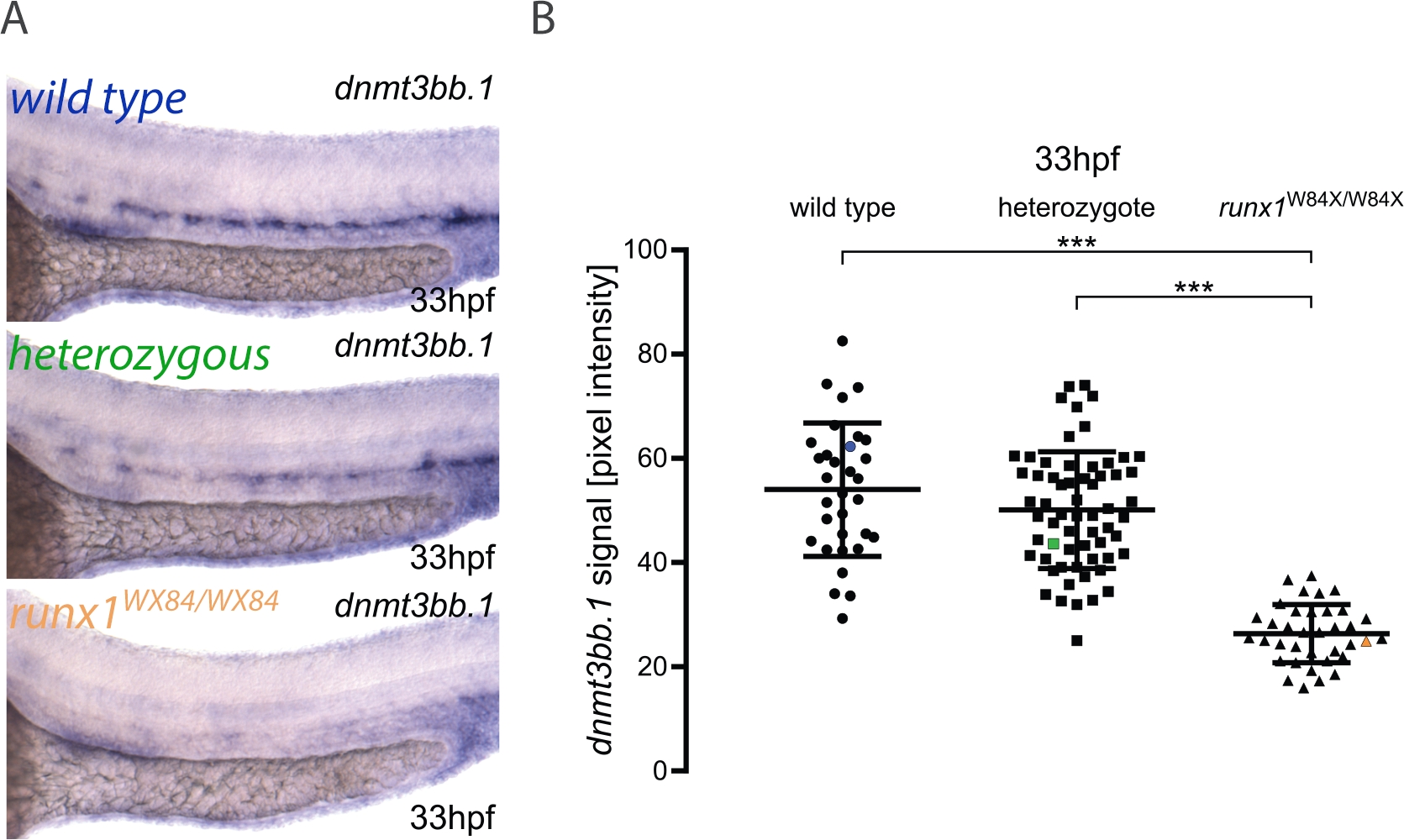
Aqui, descrevemos a aplicação prática do gasoduto para quantitação de imagens e genotipagem de embriões conforme publicado em outros lugares3. O fluxo de trabalho para o método é mostrado na Figura 1. Para ilustrar como usar esse método, ish foi realizado para dnmt3bb.1 em 33 embriões hpf de um runx1W84X/+22 incross ( Figura2). 130 embriões foram imagens usando as mesmas condições de iluminação detalhadas no protocolo e rotulando-os com um número único. Após a imagem, cada embrião foi transferido para um tubo PCR para genotipagem. Neste ponto, a análise de imagem foi realizada para atribuir um valor de intensidade de pixel a cada imagem. O genótipo foi então atribuído à sua imagem correspondente e aos valores de intensidade dos pixels agrupados de acordo com seu genótipo para análise estatística. Foi detectada uma diminuição na expressão dnmt3bb.1 nos mutantes Runx1W84X/W84X (Figura 2A,B)3,de acordo com observações anteriores5. Curiosamente, os embriões heterozigos runx1W84X/+ não mostraram diferenças significativas na expressão dnmt3bb.1 (Figura 2A,B) em comparação com seus irmãos do tipo selvagem, sugerindo que uma cópia do Runx1 é suficiente para manter a expressão dnmt3bb.1 em níveis apropriados.
Muitos mutantes de zebrafish não mostram um fenótipo embrionário que pode ser detectado usando outras tecnologias de perda de função, como oligonucleotídeos de morfolino (MOs). Essa discrepância pode ser atribuída a uma série de causas, incluindo efeitos fora do alvo, compensação de proteínamaterna, alelo hipomórfico23 ou ao fenômeno recém-descoberto de compensação genética24,25,26,27. Neste exemplo, perguntamos se a expressão runx1 foi reduzida ou perdida em mutantes lmo4auob100 desde que dados publicados anteriormente usando um MO4A MO sugeriram que o runx1 é diminuído em morfantes lmo4a 28. Aqui, a análise não revelou diferenças significativas na expressão runx1 entre o tipo selvagem eos mutanteshomozigos 3(Figura 3A,B). Análises posteriores por um único embrião qPCR mostraram que houve uma pequena, mas significativa diminuição na expressão runx1 em mutantes lmo4auob100 (Figura 3C). Assim, é possível que a quantificação da imagem não seja capaz de detectar pequenas diferenças nos níveis de expressão. Alternativamente, a falta de diferença entre genótipos detectados é real e os experimentos qPCR estão detectando alterações na expressão runx1 em outros tecidos como o telenchephalon onde tanto lmo4a quanto runx1 são expressos. Os pesquisadores devem sempre verificar seus resultados com um método independente como o qPCR, mas idealmente enriquecendo para o tecido de interesse por citometria de fluxo, por exemplo.
Em raros casos em que o ISH tem alto fundo (Figura 3D), o valor de intensidade de pixel sessa área é tão alto que a subtração do valor do sinal produz um número negativo e, nesses casos, esses embriões seriam excluídos da análise. Em nossa experiência, isso ocorreu em cerca de 0,4% dos embriões 3 sondados runx1,mas pode variar entre experimentos, sondas ou lotes de reagentes. Embora isso possa ser uma limitação do método, a baixa frequência de alto nível é muito improvável de influenciar os resultados gerais.
Para testar o efeito de selecionar diferentes áreas para correções de fundo, primeiro medimos a intensidade do pixel do sinal runx1 ISH em 28 embriões de hpf, utilizando diferentes regiões para correções de fundo(Figura 4). Foram selecionadas quatro regiões diferentes: duas na região do tronco (R1 e R2), uma na região da gema (não manchada, mas com probabilidade de acumular manchas de fundo) e uma área menor anterior ao ROI (R4, Figura 4B). Medir a intensidade dos pixels nessas regiões mostrou uma diferença relativamente estável de intensidade entre o ROI e qualquer área de fundo(Figura 4C). No entanto, o R3 sempre apresentou valores muito altos (acima dos do ROI). Após a inversão e conversão para 8 bits, a região da gema parece muito brilhante e, portanto, não é adequada para uso como correção de fundo. R2 estava mais perto do ROI, mas continha algum sinal ISH, e usá-lo para correção diminuiu a intensidade média do pixel quando comparado com qualquer R1 (localizado ainda mais dorsal, longe do sinal ISH) ou R4. Assim, ou R1 ou R4 são áreas apropriadas que podem ser utilizadas para correção de antecedentes (apesar da área de R4 ser menor que a do R1). Em seguida, queríamos comparar como o uso de R1 ou R4 afetou os resultados ao comparar a expressão runx1. Para isso, cruzamos dll4+/- heteroziggotes29 e analisamos a expressão runx1 em tipo selvagem selecionado aleatoriamente e dll4-/-embriões(Figura 4E). Embora o uso de R1 ou R4 para correção de antecedentes afetou valores individuais, as intensidades médias de pixel sem o mesmo genótipo não foram significativamente diferentes(Figura 4E). Além disso, comparar a expressão runx1 ainda produz valores de intensidade média semelhantes entre genótipos usando áreas R1 ou R4 como correção de fundo (μR1=16,3 e μR4=18,2, respectivamente). Juntos, concluímos que, embora a escolha da área de fundo seja importante, o principal critério é que ela não inclua regiões de gema (propensas ao acúmulo de manchas de fundo) e que não deve conter nenhuma mancha (específica) que possa distorcer os valores de intensidade do pixel do fundo.

Figura 1: Fluxo de trabalho do protocolo de quantitação e genotipação de imagem paralela. Embriões coletados de uma entrada de peixes heterozigos para um alelo mutante são sondados para o gene medido com um protocolo ISH padrão. Após a imagem, o DNA genômico é extraído usando o protocolo HotSHOT adicionando o tampão de lise diretamente ao embrião em um tubo PCR de 0,2 mL, seguido por uma incubação de 30 min a 95 °C. Este DNA é usado para genotipagem dos embriões por PCR, PCR e restrição de polimorfismo de comprimento de fragmento (RFLP), ensaios KASP ou qualquer outro método apropriado. Paralelamente, as imagens de cada embrião são invertidas e convertidas em cinza saem de 8 bits. RoIs de forma e tamanho idênticos contendo o sinal ISH (amarelo) e fundo (azul) são selecionados manualmente e medidos. As medidas, atribuídas aos genótipos correspondentes, são estatisticamente analisadas. Figura adaptada de Dobrzycki et al.3Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Quantitação de imagem em mutantes runx1 revela níveis reduzidos de expressão dnmt3bb.1 por ISH. (A) Exemplo imagens de ISH em 33 hpf tipo selvagem (azul), runx1+/W84X (verde) e runx1W84X/W84X (laranja) embriões, mostrando expressão dnmt3bb.1 na aorta dorsal. (B) Os valores de intensidade do Pixel de dnmt3bb.1 mRNA em runx1W84X/W84Xembriões (n=36) são significativamente reduzidos em comparação com tipos selvagens (n=32) e heteroziggotes (n=62) (ANOVA, p < 0,001). Os coeficientes de variação são 24%, 22% e 21% para grupos selvagens, heterozigos e mutantes, respectivamente. O ponto de dados azul, verde e laranja corresponde às imagens de exemplo do painel A. As barras representam média ± s.d. ***p<0.001 (Teste pós-hoc Games-Howell). Figura adaptada de Dobrzycki et al.3Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Medir níveis de expressão runx1 por ISH em mutantes lmo4auob100. (A) Imagens representativas do ISH para runx1 em 28 hpf tipo selvagem (azul), heterozigoso (verde) e lmo4auob100/uob100 (laranja) embriões, mostrando a expressão na aorta dorsal. (B ) Quantificação do sinal runx1 mRNA, detectado sacando por ISH, do tipo selvagem de 28 hpf (n=15), embriões heterozigos lmo4a+/- (het) (n=34) e lmo4auob100/uob100 embriões mutantes (n=18) de uma embreagem não mostra diferença significativa na intensidade de pixels runx1 entre os diferentes genótipos (ANOVA,> p 0.6). O ponto de dados azul, verde e laranja corresponde às imagens de exemplo do painel A. As barras representam média ± s.d. (C) Boxplots exibindo níveis normalizados runx1 mRNA (2-ΔCt) em tipo selvagem único (azul; n=12) e lmo4auob100/uob100 (mu, laranja; n=12) embriões, medidos pelo qRT-PCR, mostrando níveis reduzidos de runx1 nos mutantes em comparação com o tipo selvagem. *p < 0,05(t teste). (D) Exemplo de um experimento ISH em um embrião de 28 hpf (manchado para runx1, pontas de flecha amarela) mostrando fundo alto. Figura adaptada de Dobrzycki et al.3Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Efeito da correção da intensidade de fundo nos resultados de medição. (A)Imagem representativa da coloração runx1 ISH em um embrião do tipo selvagem a 28 hpf. (B)Mesma imagem após inversão e conversão para 8 bits. A região de interesse (ROI) é destacada em verde e quatro áreas diferentes utilizadas para correção de antecedentes (R1-R4) são destacadas em amarelo. (C) As medidas de intensidade de pixels brutos em todas as regiões mostradas no painel B. Observe que a intensidade no R3 (gema) é consistentemente maior do que o sinal ISH real no ROI (n=11). (D) Níveis de expressão Runx1 no ROI utilizando áreas de fundo R1, R2 e R4. Áreas para ROI, R1, R2 e R3~28500 pixels; R4~8500 pixels. Observe que o fundo R3 não foi utilizado para essa comparação, pois a correção de antecedentes (ROI-R3) consistentemente produziu valores negativos. (E) Níveis de expressão Runx1 no tipo selvagem e dll4-/- mutantes usando R1 ou R4 para correção de fundo (n=10 para cada amostra). A análise estatística nos painéis D e E foi realizada usando um teste kruskal-wallis não paramétrico, assumindo que os valores de intensidade do pixel não são normalmente distribuídos. Clique aqui para ver uma versão maior deste valor.
{kind=link}
Discussão
Alguns fatores devem ser considerados ao usar este método para quantificar a expressão genética. As condições de imagem devem ser mantidas ao longo do experimento (por exemplo, iluminação, tempos de exposição e posicionamento de embriões) para reduzir a variabilidade entre as medidas. Um ponto crítico é evitar a supermancha das amostras, pois as diferenças na coloração entre amostras podem ser mascaradas. Por exemplo, a diminuição da expressão VegfA na ausência de Eto2 em Xenopus laevis embriões30 só poderia ser detectada monitorando cuidadosamente a coloração durante um período de 24 horas. Assim, é uma boa prática determinar empiricamente níveis adequados de coloração para cada gene que melhor representem sua expressão, sem atingir a saturação. A supercoloração também aumentará artificialmente a intensidade dos pixels de fundo nas imagens convertidas em escala de cinza de 8 bits e distorcerá os resultados de quantitação. Em casos extremos, o nível de fundo em tecidos embrionários pode ser maior do que o sinal ISH no ROI selecionado e essas amostras devem ser excluídas da análise. Um fenômeno semelhante foi observado quando testamos a adequação da gema não manchada para correção de fundo (Figura 4). Após a inversão e conversão para 8 bits, os pixels mais escuros na região da gema tornam-se mais brilhantes do que o sinal ISH no embrião e tornam os valores corrigidos por fundo negativos. Assim, evite o uso da gema para correção de fundo. Medir o sinal de fundo em áreas pigmentadas no embrião (por exemplo, os olhos ou a parte dorsal do tronco a partir de 26/28 hpf em diante) irá igualmente distorcer os resultados de quantitação e também deve ser evitado. Existem protocolos disponíveis para branqueamento de embriões de zebrafish, antes ou depois de ISH18 e embriões branqueadores com mais de 24 hpf antes da recomendação da imagem.
Como este método depende de medir a intensidade dos pixels em uma área definida contra uma intensidade de pixel de fundo em uma área equivalente não manchada, não é apropriado para quantitação de genes onipresentes ou quase onipresentemente expressos como está. Em vez disso, é bem adequado para medir a expressão de genes com uma distribuição espacialmente restrita onde uma área para medir a intensidade dos pixels de fundo pode ser prontamente identificada. Nossa análise adicional agora sugere que o uso de uma área menor (3-4x menor) para correção de antecedentes produz resultados semelhantes ao uso de uma área equivalente à do ROI. Isso estende a aplicabilidade do método aos genes expressos em domínios espaciais mais amplos (e, portanto, exigindo ROIs maiores para medidas de intensidade), desde que se possa usar áreas claramente não manchadas do embrião para correção de fundo.
Por fim, sugerimos que o genotipamento seja realizado em paralelo ou após a quantitação da imagem para minimizar o viés do experimentador. Pedir a um segundo experimentador para repetir a quantitação em amostras anonimizadas e comparar com o primeiro conjunto de medições também ajudará a reduzir o viés do experimentador. Se as imagens a serem quantificadas são de uma comparação entre tratamentos que não requerem genotipagem (por exemplo, tipo selvagem versus inibidor químico ou tipo selvagem vs. knockdown mo), o experimentador realizando as medidas deve ser cego à identidade do Amostra.
Divulgações
Os autores declaram que não têm interesses financeiros concorrentes.
Agradecimentos
Gostaríamos de agradecer aos funcionários dos Serviços Biomédicos de Oxford e Birmingham por excelente criação de zebrafish. T.D. foi financiado por uma bolsa de doutorado em cromossomo e biologia do desenvolvimento da Wellcome Trust (#WT102345/Z/13/Z). R.M. e M.K. foram financiados pela British Heart Foundation (BHF IBSR Fellowship FS/13/50/30436) e são gratos por seu generoso apoio. R.M. reconhece o apoio do Centro de Excelência em Pesquisa da BHF (RE/13/1/30181), Oxford.
Materiais
| Name | Company | Catalog Number | Comments |
| 0.2 mL PCR tubes (8-strips with lids) | StarLab | A1402-3700 | 96-well plates are equally appropriate for sample handling but beware of cross contamination between samples |
| 3 mL Pasteur pipettes | Alpha Laboratories | LW4114 | |
| Cavity slides | Brand | BR475535-50EA | |
| Digital Camera (Qimaging Micropublisher 5.0) | Qimaging | ||
| Eppendorf Microloader tips | Eppendorf | 10289651 | the tips are used to orient the embryos for imaging in glycerol |
| Excel | Microsoft | ||
| F3000 Fiber Optic Cold Light Source | Photonic | ||
| Fiji | |||
| Glycerol | Sigma | G5516-1L | |
| Graphpad Prism 8.01 | GraphPad Software, Inc. | we prefer to use Graphpad, but other statistics software packages are also suitable (e.g. SigmaPlot or SPSS) | |
| HotSHOT alkaline lysis buffer | 25 mM NaOH, 0.2 mM disodium EDTA, pH 12 | ||
| HotSHOT neutralization buffer | Tris HCl 40 mM, pH 5 | ||
| PBS (10X) pH 7.4 | Thermofisher | 70011044 | |
| Stereomicroscope with illumination stand (Nikon SMZ800N) | Nikon | ||
| Thermocycler | Thermofisher |
Referências
- Varshney, G. K., et al. High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome Research. 25 (7), 1030-1042 (2015).
- Varshney, G. K., et al. CRISPRz: a database of zebrafish validated sgRNAs. Nucleic Acids Research. 44 (D1), D822-D826 (2016).
- Dobrzycki, T., Krecsmarik, M., Bonkhofer, F., Patient, R., Monteiro, R. An optimised pipeline for parallel image-based quantification of gene expression and genotyping after in situ hybridisation. Biology Open. 7 (4), bio031096 (2018).
- Bresciani, E., et al. CBFbeta and RUNX1 are required at 2 different steps during the development of hematopoietic stem cells in zebrafish. Blood. 124 (1), 70-78 (2014).
- Gore, A. V., et al. Epigenetic regulation of hematopoiesis by DNA methylation. Elife. 5, e11813 (2016).
- Fan, Y., et al. Tissue-Specific Gain of RTK Signalling Uncovers Selective Cell Vulnerability during Embryogenesis. PLoS Genetics. 11 (9), e1005533 (2015).
- Wen, B., et al. GATA5 SUMOylation is indispensable for zebrafish cardiac development. Biochimica et Biophysica Acta. 1861 (7), 1691-1701 (2017).
- Espin-Palazon, R., et al. Proinflammatory signaling regulates hematopoietic stem cell emergence. Cell. 159 (5), 1070-1085 (2014).
- Peterkin, T., Gibson, A., Patient, R. Redundancy and evolution of GATA factor requirements in development of the myocardium. Developmental Biology. 311 (2), 623-635 (2007).
- Genthe, J. R., Clements, W. K. R-spondin 1 is required for specification of hematopoietic stem cells through Wnt16 and Vegfa signaling pathways. Development. 144 (4), 590-600 (2017).
- Kalev-Zylinska, M. L., et al. Runx1 is required for zebrafish blood and vessel development and expression of a human RUNX1-CBF2T1 transgene advances a model for studies of leukemogenesis. Development. 129 (8), 2015-2030 (2002).
- Butko, E., et al. Gata2b is a restricted early regulator of hemogenic endothelium in the zebrafish embryo. Development. 142 (6), 1050-1061 (2015).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Lleras Forero, L., et al. Segmentation of the zebrafish axial skeleton relies on notochord sheath cells and not on the segmentation clock. Elife. 7, (2018).
- Jowett, T., Yan, Y. L. Double fluorescent in situ hybridization to zebrafish embryos. Trends in Genetics. 12 (10), 387-389 (1996).
- Thisse, C., Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nature Protocols. 3 (1), 59-69 (2008).
- Narayanan, R., Oates, A. C. Detection of mRNA by Whole Mount in situ Hybridization and DNA Extraction for Genotyping of Zebrafish Embryos. Bio-protocol. , e3193 (2019).
- Monteiro, R., Pouget, C., Patient, R. The gata1/pu.1 lineage fate paradigm varies between blood populations and is modulated by tif1gamma. EMBO JOURNAL. 30 (6), 1093-1103 (2011).
- Truett, G. E., et al. Preparation of PCR-quality mouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). Biotechniques. 29 (1), 52-54 (2000).
- Wilkinson, R. N., Elworthy, S., Ingham, P. W., van Eeden, F. J. A method for high-throughput PCR-based genotyping of larval zebrafish tail biopsies. Biotechniques. 55 (6), 314-316 (2013).
- Parant, J. M., George, S. A., Pryor, R., Wittwer, C. T., Yost, H. J. A rapid and efficient method of genotyping zebrafish mutants. Developmental Dynamics. 238 (12), 3168-3174 (2009).
- Jin, H., et al. Definitive hematopoietic stem/progenitor cells manifest distinct differentiation output in the zebrafish VDA and PBI. Developement. 136 (4), 647-654 (2009).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. 13 (10), e1007000 (2017).
- El-Brolosy, M. A., Stainier, D. Y. R. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genetics. 13 (7), e1006780 (2017).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. 524 (7564), 230-233 (2015).
- Ma, Z., et al. PTC-bearing mRNA elicits a genetic compensation response via Upf3a and COMPASS components. Nature. 568 (7751), 259-263 (2019).
- El-Brolosy, M. A., et al. Genetic compensation triggered by mutant mRNA degradation. Nature. 568 (7751), 193-197 (2019).
- Meier, N., et al. Novel binding partners of Ldb1 are required for haematopoietic development. Development. 133 (24), 4913-4923 (2006).
- Kettleborough, R. N., et al. A systematic genome-wide analysis of zebrafish protein-coding gene function. Nature. 496 (7446), 494-497 (2013).
- Leung, A., et al. Uncoupling VEGFA functions in arteriogenesis and hematopoietic stem cell specification. Developmental Cell. 24 (2), 144-158 (2013).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados