
Method Article
תרומה של Na+/ K+ משאבה להתפוצצות קצבית, נחקרת עם דוגמנות ניתוחי מהדק דינמיים
In This Article
Erratum Notice
Summary
מוצג כאן היא שיטה לחקירה של התפקידים של Na+/ K+ משאבה מתמשכתNa + הנוכחי ineurons לב עלוקה באמצעות מלחציים דינמיים.
Abstract
משאבת Na+/ K+ , הנחשבת לעתים קרובות כפונקציית רקע בפעילות העצבית, תורמת זרם חיצוני (Ipump) המגיב לריכוז הפנימי של Na+ ([Na+]i). בנוירונים מתפוצצים, כגון אלה שנמצאו ברשתות עצביות של מחולל דפוסים מרכזי (CPG) המייצרות תנועות קצביות, ניתן לצפות שמשאבתה- I [Na+] ולכן משאבת I תשתנה לאורך מחזור ההתפרצות. היענות זו לפעילות חשמלית, בשילוב עם עצמאות מפוטנציאל הממברנה, מעניקה לימשאבה עם תכונות דינמיות שאינן נפוצות בזרמים מבוססי ערוץ (למשל, ערוצי מתח או משדר מגודרים או דליפה). יתר על כן, בנוירונים רבים, פעילות המשאבה מווסתתת על ידי מגוון של אפננים, מה שמרחיב עוד יותר את התפקיד הפוטנציאלי של אנישואב בפעילות מתפרצת קצבית. מאמר זה מראה כיצד להשתמש בשילוב של מודלים ושיטות הידוק דינמיות כדי לקבוע כיצד אנימשאבה ואת האינטראקציה שלה עם Na+ פעילות קצבית השפעה נוכחית מתמשכת ב- CPG. באופן ספציפי, מאמר זה יתמקד בפרוטוקול מהדק דינמי ושיטות מידול חישוביות בלב פנימי של עלוקות רפואיות.
Introduction
פעימות לב בעלוקות מונעות על ידי CPG המורכב מ-9 זוגות דו-צדדיים של אינטראורונים לב (HNs) המופצים על פני גרעינים מגזריים בינוניים רבים. בליבת ה-CPG נמצאים זוגות מעכבים הדדית של אינטראורונים הממוקמים בגרעיניהחלק השלישי וה-4 המהווים מתנדים חצי-מרכזיים (HCOs)(איור 1A). נוירונים אלה ממשיכים להתפוצץ כאשר מבודדים באופן סינפטי באמצעות bicuculline1. אחרים, כגון הזוג בגרעיניהמגזרי 7 (המוקד של פרוטוקול זה), הם גם מתפרצים, המסוגלים לייצר פעילות מתפרצת כאשר מבודדים באופן סינפטי. הם אינם מחוברים זה לזה ומקבלים רק קלט יורד, ולכן מבודדים בקלות על ידי ניתוק הגנגליון משאר חוט העצבים. פעילות מתפרצת עצמאית זו רגישה לזרם הדליפה שהוכנס שנגרם על ידי חדירה עם מיקרו-ectrodes חד להקלטה אך מתפוצצת במרץ כאשר היא מוקלטת בשיטות תיקון רופף1.
הן נוירוני HN בודדים והן HN HCOs עוצבו (מודלים של תא איזופוטנציאלי יחיד מבוסס הודג'קין-האקסלי של נוירוני HN המכילים את כל הזרמים המגודרים והסינפטיים שזוהו באופן ניסיוני), וכל מאפייני הפרץ של המערכת החיה נתפסו בהצלחה2. מיומודולין, נוירופפטיד אנדוגני בעלוקות, מקטין באופן משמעותי את התקופה (T) של קצב הפרץ של נוירונים HN מבודדים ו- HN HCOs. אפנון זה פועל כדי להגדיל את זרם h (היפר-קוטביות מופעל זרם פנימה, אניh) כדי להקטין אנימשאבה3. תצפית זו הובילה לחקירה של איך אנימשאבה אינטראקציה עם Ih, וכיצד אפנון המשותף שלהם תורם לפעילות הקצבית של נוירונים HN. הפעלת המשאבה על ידי הגדלת [Na+]i (באמצעות מונסין ionophore) מאיצה את קצב פרץ HN הן HCOs HN נוירונים HN מבודדים4. האצה זו הייתה תלויה ב- ih. כאשר נחסמתי (2 מ"מ צלז צלזיון+), תקופת הפרץ לא שונתה בשיטה זו של הפעלת המשאבה; עם זאת, משך הפרץ (BD) צומצם, ומרווח הבין-גזעי (IBI) גדל הן ב- HN HCOs והן בנוירונים מבודדיםHN 4.
עבור פרוטוקול זה, כל הזרמים של נוירון HN(7) חי, כולל זרם המשאבה, אנימשאבה, משולבים במודל HN כדלקמן:
 (1)
(1)
כאשר C הוא קיבוליות הממברנה (ב- nF), V הוא פוטנציאל הממברנה (ב- V), t הוא זמן (ב s). תיאורים ומשוואות זרם יוני מפורטים תוארו במקום אחר2,4. נוירון הדגם המלא של דגם HN פועל בזמן אמת(איור 2). התוכנה תהיה זמינה על GitHub עם הפרסום ויהיה מתאים לרוץ על לוח עיבוד האות הדיגיטלי המתואר בטבלת החומרים. כאן, מוקד החקירה הוא זרם Na+/ K+ משאבה (אנימשאבה) ואת הזרמים מגודר מתח תורם משמעותי Na+ שטף: זרם Na+ מהיר (INa) ו זרם Na+ מתמשך (IP). ההולכות המקסימליות של זרמים אלה הן 
 בהתאמה. Na+/ K+ משאבה מחליף שלושה Na תאי+ יונים עבור שני K חוץ תאי+ יונים, ובכך לייצר זרם חיצוני נטו. חשוב לציין, הוא שואב 3 פעמים Na+ מתוך הנוירון כפי שזרם זה מציין, אשר חשוב לחישוב Naתאי + ריכוז.
בהתאמה. Na+/ K+ משאבה מחליף שלושה Na תאי+ יונים עבור שני K חוץ תאי+ יונים, ובכך לייצר זרם חיצוני נטו. חשוב לציין, הוא שואב 3 פעמים Na+ מתוך הנוירון כפי שזרם זה מציין, אשר חשוב לחישוב Naתאי + ריכוז.
זרם המשאבה Na+/K+ תלוי בריכוזים תאיים של Na+ והוא בא לידי ביטוי על ידי הפונקציה הסיגמוידלית הבאה:
 (2)
(2)
כאשר [Na]i הוא נה תאי+ ריכוז,  הוא מקסימום Na+/ K+ משאבה הנוכחי, [Na]ih הוא נה תאי+ ריכוז עבור חצי הפעלה של Na+/ K+ משאבה, ו [Na]הוא הרגישות של Na+/ K+ משאבה ל [Na]i. [Na]אני בונה כתוצאה של Na+ זרם נישא על ידי IP ואני Na והוא פחת על ידי Na+ שפכים של Na+/ K+ משאבה. התרומה של Ih ואנידולף סך Na+ שטף הוא קטן ולא נחשב במודל בזמן אמת.
הוא מקסימום Na+/ K+ משאבה הנוכחי, [Na]ih הוא נה תאי+ ריכוז עבור חצי הפעלה של Na+/ K+ משאבה, ו [Na]הוא הרגישות של Na+/ K+ משאבה ל [Na]i. [Na]אני בונה כתוצאה של Na+ זרם נישא על ידי IP ואני Na והוא פחת על ידי Na+ שפכים של Na+/ K+ משאבה. התרומה של Ih ואנידולף סך Na+ שטף הוא קטן ולא נחשב במודל בזמן אמת.
 (3)
(3)
שבו, v הוא הנפח (~ 6.7 pL) של המאגר Na+ תאי תאי, F הוא הקבוע של פאראדיי, ואת נה+ ריכוז חוץ תאי נשמר קבוע.
מוליכות מגודרות מתח ודליפות הובחנו - אלה מגיבים לפוטנציאל הממברנה - מזרם המשאבה, המוסדר על ידי ריכוז Na+ תאי מחושב ([Na+]i). [נה+] i בנוי דרך Na+ כניסה באמצעות זרם Na+ מהיר (INa) המייצר פוטנציאל פעולה (קוצים) ואת זרם Na+ מתמשך (IP) המספק את depolarization כדי לתמוך spiking. [נה+] אני, בתורו, מופחת על ידי הפעולה של המשאבה דרך שחול של Na+. ערכי HN חיים בסיסיים של  (5nS) ו
(5nS) ו  - (150 nS) הונחו, ואנו לוקחים בחשבון כל מהדק דינמי נוסף .
- (150 nS) הונחו, ואנו לוקחים בחשבון כל מהדק דינמי נוסף .
מטרת הפרוטוקול המתואר כאן היא לתמרן אנילשאוב בדיוק והפכה בזמן אמת כדי לגלות כיצד הוא אינטראקציה עם זרמים מגודרים מתח (זרם Na+ מתמשך בפרוטוקול הנוכחי) כדי לשלוט פרץ קצבי ב- HNs יחיד. כדי להשיג מטרה זו, נעשה שימוש במצדק דינמי, אשר מציג באופן מלאכותי, על הפקודה, כמות מדויקת של כל זרם שניתן לחשב כמו המודל פועל. לשיטה זו יש יתרונות על פני מניפולציה פרמקולוגית של המשאבה, המשפיעה על הרקמה כולה, יכול להיות השפעות מחוץ למטרה כי הם לעתים קרובות קשה להפוך, ולא ניתן מניפולציה מדויקת. מהדק דינמי5,6 קורא את המתח של נוירון מוקלט בזמן אמת ( איור1B) ומחשב ומזריק, בזמן אמת, את כמות כל זרם המבוסס על משוואות מודל ואת הערכים שנקבעו של כל  או
או  . שיטות דומות ניתן ליישם בקלות על כל נוירון שניתן להקליט תאיים. עם זאת, פרמטרים יצטרכו להיות rescaled הנוירון שנבחר, ואת הנוירון צריך להיות מבודד תשומות סינפטיות, למשל, פרמקולוגית.
. שיטות דומות ניתן ליישם בקלות על כל נוירון שניתן להקליט תאיים. עם זאת, פרמטרים יצטרכו להיות rescaled הנוירון שנבחר, ואת הנוירון צריך להיות מבודד תשומות סינפטיות, למשל, פרמקולוגית.
Protocol
הערה: נושאים ניסיוניים בעלי חיים לא חולייתיים אינם מוסדרים על ידי האוניברסיטאות NIH או אמורי וג'ורג'יה. בכל זאת ננקטו כל האמצעים כדי למזער את סבל העלוקות המשמשות בעבודה זו.
1. הכן גנגליון מבודד 7 מחבל העצבים העלוקה
- שמור על עלוקות Hirudo ורבנה במי בריכה מלאכותיים (המכילים 0.05% w /v של מלח ים) מדולל במים דה מיוננים ב 16 °C (על מחזור 12:12 אור כהה.
- הכן את העלוקות לניתוח על ידי קר-anesthizing אותם במיטה של קרח כתוש במשך >10 דקות עד immobile.
- מלאו צלחת ניתוח שחורה מרופד שרף לעומק של כ-1 ס"מ עם תמיסת מלח צוננת המכילה 115 מ"מ NaCl, 4 מ"מ KCl, 1.7 מ"מ CaCl2, 10 מ"מ D-גלוקוז, ו-10 מ"מ HEPES במים נטולי יינון; pH מותאם ל 7.4 עם 1 M NaOH. מצמידים את צד העלוקה לתא מצופה השרף השחור (לפחות 20 ס"מ על 10 ס"מ עם עומק של לפחות 2 ס"מ מעל השרף בעובי של לפחות 2 ס"מ).
- תחת סטרומיקרוסקופ בהגדלה של פי 20 עם תאורת מדריך אור עקשני, בצע חתך אורך של לפחות 3 ס"מ עם מספריים קפיציים 5 מ"מ דרך דופן הגוף בחלק 1/3של הגוף. השתמש סיכות כדי למשוך הצידה את דופן הגוף ולחשוף את האיברים הפנימיים.
הערה: כל ארוחת דם מאוחסנת ניתן להסיר על ידי שאיבה עם פיפטת פסטר מלוטש אש. - לבודד גנגליון גוף בינוני בודד 7 (גנגליון פלחוני חופשי שביעי למוח).
- פתח את הסינוס שבו חבל העצבים שוכן באמצעות מספריים קפיציים 5 מ"מ. הקפידו לפצל את הסינוסים באופן שרירי ומשאירים שתי רצועות סינוס. השתמש במלקחיים חדים #5 כדי לעזור להנחות את החיתוך ולהחזיק את הסינוס.
- שמור את הסינוס מחובר לכל אחד משני שורשי העצבים הדו-צדדיים שיוצאים מן הגנגליון (הוא דבק בחוזקה לכל שורש) כדי להשתמש ברצועות אלה של סינוס להצמדת הגנגליון.
- הסר את הגנגליון מהגוף על ידי חיתוך חבילות עצב החיבור rostral ו caudal המקשרות את הגרעינים (רחוק מןהגנגליון 7 ככל האפשר) ואת רצועות הסינוס, ולאחר מכן לחתוך את השורשים לרוחב למקום שבו הם מגיחים מן הסינוס.
- הצמידו את הגנגליון (באמצעות מלקחיים ישנים קהים מס '5) עם סיכות חרקים מקוצרות, בצד הגחון למעלה, בצלחות פטרי ברורות ומרופדים בשרף. הכנס סיכות ברצועות של סינוס ורקמות רופפות דבקות בשורשים ואת חיבורי rostral ו caudal, רחוק ככל האפשר מן הגנגליון.
הערה: השרף חייב להיות לא עבה יותר מ 3 מ"מ אם תאורה טובה מלמטה הוא להיות מושגת במהלך ההקלטה. ודאו שההנגליון מתוח, הן לאורך והן לרוחב. - הגדל את ההגדלה של סטריאומיקרוסקופ ל 40x או יותר, ולהתאים את התאורה האלקטורלית כך שגופי התא העצבי ניתן לראות בקלות על פני השטח הגחון של הגנגליון ממש מתחת פרינוריום.
- הסר את פרינוריום של הגנגליון (desheath) עם מיקרו-סיטסיטורים.
- התחל את desheathing על ידי חיתוך נדן רופף בין השורשים בצד אחד, ולהמשיך את החתך לרוחב לצד השני, הקפד לשמור על להבי המספריים שטחי ולא לפגוע בגופי התא העצבי ישירות מתחת נדן.
- לעשות חתך שטחי דומה caudally מן החתך לרוחב לאורך קו האמצע.
- עכשיו לתפוס את דש caudolateral של נדן בצד אחד עם מלקחיים #5 בסדר, למשוך אותו מן הגנגליון, ולחתוך אותו עם microscissors.
- חזור על הפעולה בצד השני; הליך זה חושף את שני הנוירונים HN(7) להקלטה עם microelectrodes.
- מניחים את צלחת ההכנה במערך ההקלטה, ומניחים עם תמיסת מלח בקצב זרימה של 5 מ"ל / דקה בטמפרטורת החדר.
2. לזהות ולהקליט אינטראורונים לב עלוקה עם מיקרואלקטרודים חדים
- למשך ההקלטה של נוירון HN(7) (הקלטות נמשכות בין 30 ל -60 דקות), לרכוש ודיגיטציה את עקבות הזרם התאי והמתח מדגם אלקטרומטר נוירופיזיולוגי בקצב של 5 kHz עם רכישת נתונים דיגיטליים (אנלוגי לדיגיטל, A עד D) וגירוי (דיגיטלי לאנלוגי, D עד A) מערכת, ולהציג על מסך מחשב.
הערה: ניתן להשתמש בכל תוכנה מסחרית או מותאמת אישית ולוח A עד D ל- A לרכישת נתונים (A עד D). D עד A ותוכנה שנבנתה בהתאמה אישית נדרשות עבור מהדק דינמי. - תחת סטרומיקרוסקופ ב 50-100x עם תאורת שדה כהה מלמטה, לזהות באופן זמני נוירון HN(7) של הזוג הדו צדדי על ידי מיקומו הקנוני בעמדה posteriolateral בגנגליון באמצע גוף שבע.
- כעת שואפים לחדור את הנוירון HN(7) putative עם microelectrode חד מלא 2 M אשלגן אצטט ו 20 mM KCl באמצעות מיקרומניפולטור.
- הנח את המיקרו-קטרוד קרוב מאוד לגוף תא המטרה.
- בחן ללא הרף את הפוטנציאל המוקלט עם האלקטרומטר, והגדר את הפוטנציאל הזה לאפס mV לפני החדירה לנוירונים.
- לחדור את הנוירון עם microelectrode, על ידי נהיגה איטית האלקטרודה לאורך ציר ארוך שלה עם המניפולטור. באמצעות פונקציית זמזום אלקטרומטר, להגדיר את משך זמזום 100 ms, עד שינוי שלילי בפוטנציאל הממברנה ופעילות spiking נמרצת הוא ציין.
- הגדר את האלקטרומטר במצב מהדק זרם רצסיבי (DCC) ≥ 3 kHz כדי לתעד בו זמנית את פוטנציאל הממברנה ולעבור זרם עם microelectrode יחיד (פיצוי קיבולת מוגדר רק מתחת לצלצול ולאחר מכן חייג בחזרה 10%).
- לפקח על ההתיישבות של האלקטרודה במהלך DCC על אוסצילוסקופ.
- הזרק זרם קבוע של -0.1 nA עם מזרק זרם קבוע אלקטרומטר במשך דקה או שתיים כדי לייצב את ההקלטה.
- זהה בוודאות את הנוירון HN(7) לפי צורת הספייק האופיינית לו ופעילות מתפרצת חלשה (איור 1Ci).
- בצע כל ניתוח נתונים במצב לא מקוון לאחר השלמת הניסוי ושמור את כל הנתונים בדיסק.
3. לבנות HN בזמן אמת או נוירון מודל אחר
- בנה תוכנה מותאמת אישית באמצעות לוח עיבוד אותות דיגיטלי (DSB; D עד A ו- A עד D) במחשב שולחני כדי ליישם בזמן אמת את זרמי המודל המתוארים בזרמי מודל2,4 או שונים עבור נוירונים או ניסויים אחרים.
- השתמש משוואות בסגנון הודג'קין-האקסלי מכיוון שהן השיטה המועדפת בדרך כלל לייצוג זרמי דגם.
- ראה7 לקבלת תיאור מפורט של היישום של מודל HN בזמן אמת ומהדק דינמי לפני הוספת זרם המשאבה. עיין בסעיף המבוא לתיאור הזרמים, ריכוז Na+ תאיים, ומוליכות של נוירון HN(7) חי במודל HN.
4. ליישם ולגוון מוליכות/זרמים דינמיים
- השתמש בתוכנת הידוק דינמי שנבנתה בהתאמה אישית עבור ה- DSB כדי ליישם ולשנות בזמן אמת מהדק כל ממשק משתמש גרפי (איור 3) (GUI) מוליכות מתוכנתות וזרמים של מודל HN בזמן אמת של הנוירון HN(7).
הערה: כתזכורת, והם ההולכה המקסימלית של זרם Na+ המתמשך (IP) וזרם המשאבה המקסימלי (אנימשאבה), בהתאמה. - השתמש בתיבות הזנת GUI בתוכנה כדי לבצע שינויים, כאשר המודל פועל, בתיבה (PumpMaxL) ו -(GpinHNLive) (איור 3).
הערה: תיבות הקלט של GUI מקבלות ערכים מוקלדים, ושלבים של 0.1 nA מומלצים עבור ושלבים של 1 nS מומלצים עבור .- הוסף כמויות קטנות של ועם מהדק דינמי כדי לייצב את פרץ של נוירון HN(7), אשר נחלש על ידי דליפה המושרה microelectrode, כפי שמוצג באיור 1Cii.
הערה: חדירת מיקרו-ectרודe גורמת לנזק בממברנה המתבטא כמוליכת דליפה מוגברת או ירידה בהתנגדות לקלט. - התחל על ידי הוספת ערך של 0.1-0.2 nA, אשר מפצה על הדליפה הנגרמת microelectrode, אבל מדכא עירור, ולאחר מכן להגדיל בהדרגה , אשר מגביר את העירור, עד מתפרץ רגיל מתפתח, בדרך כלל ב ~ 1-4 nS (איור 4A).
- הוסף כמויות קטנות של
- באופן שיטתי לשנות זרמים אלה (במרווחים של 0.1 nA עבור ו 1 nS עבור ) כדי נוירון HN(7) המוקלט עם מהדק דינמי (איור 3), ולהעריך את ההשפעות שלהם על מאפייני פרץ: תדירות ספייק (ו: ההדדיות של הממוצע של מרווח interspike במהלך פרץ), מרווח interburst (IBI: הזמן בין העלייה האחרונה בפרץ אחד לספייק הראשון בפרץ הבא), משך ההתפרצות (BD: הזמן בין העלייה הראשונה בפרץ לבין העלייה האחרונה בפרץ), ותקופת פרץ (T: הזמן בין העלייה הראשונה בפרץ לבין העלייה הראשונה בפרץ הבא).
- לשנות את הערכים של , כמו בהדגמת הווידאו, כדי להכיר את הטכניקה ולאחר מכן לצאת החוצה.
- החזק בערך קבוע מסוים וטאטא בהפרשים קבועים של 1 nS על-פני טווח של פעילות מתפרצת רגילה תומכת.
- כעת הגדל את הערך הקבוע של 0.1 nA ושוב לטאטא על מגוון של פעילות מתפרצת רגילה תומכת.
- עבור כל זוג פרמטרים מיושם, לאסוף נתונים המכילים לפחות 8 התפרצויות כך שניתן יהיה לבצע מדדים ממוצעים אמינים של f, IBI, BD ו- T.
- המשך עם גורפים כל עוד הנוירון נשאר בר קיימא, כפי שהוערך על ידי spiking חזק ופוטנציאל בסיסי יציב של תנודה.
- אסוף נתונים ממספר נוירונים (מבעלי חיים שונים) כדי ליצור גרף מורכב(איור 5).
- החזק
- לשנות את הערכים של
תוצאות
דוגמנות עם תוספת של משאבה4 הביאה את הממצאים הניסיוניים שהוצגו בסעיף המבוא לפוקוס חד יותר והחלה להסביר את מנגנון התפוצצות בסיוע המשאבה. המודל בזמן אמת שהודגם כאן מכוון ( ופרמטרים נבחרו) כך שהוא מייצר פעילות קצבית סדירה הנופלת בגבולות הפעילות הרגילה כפי שנצפו בניסויים - f, IBI, BD, T - וממשיך לייצר פעילות כזו כאשר הפרמטרים המופננים למיומודולין (זרם המשאבה המקסימלי) ו  (מוליכות מקסימלית של h-זרם) מגוונים או מגוונים במודל. ערכי הפרמטר שנקבעו יכולים לשמש כמדד או ערכת קנונים לניסויי מידול. במקרי מודל אלה, אנישואב מתנדות לאורך מחזור הפרץ כמו [Na+]i סביב רמה בסיסית. אנימשאבה תורמת סיום פרץ במהלך שלב הפרץ, ואת hyperpolarization זה מייצר מפעיל אניh במהלך IBI; שימו לב לרמה המקסימלית של Ih ליד חניכת פרץ(איור 2).
(מוליכות מקסימלית של h-זרם) מגוונים או מגוונים במודל. ערכי הפרמטר שנקבעו יכולים לשמש כמדד או ערכת קנונים לניסויי מידול. במקרי מודל אלה, אנישואב מתנדות לאורך מחזור הפרץ כמו [Na+]i סביב רמה בסיסית. אנימשאבה תורמת סיום פרץ במהלך שלב הפרץ, ואת hyperpolarization זה מייצר מפעיל אניh במהלך IBI; שימו לב לרמה המקסימלית של Ih ליד חניכת פרץ(איור 2).
למרות שמודל ה-HN בזמן אמת כולל את כל הזרמים המיושמים2,4 הזמינים להדק דינמי, ההתמקדות כאן הייתה ב- ו- , אשר זמינים לשינויים בזמן שהמודל פועל ב- GUI המהדק הדינמי (איור 3). מהדק דינמי מאפשר לנסיין להוסיף (או להחסיר עם שלילי או ) כל מוליכות או זרם לתוך נוירון באופן מלאכותי המחקה את המתח והתלות היונית של מוליכות אמיתית או זרם. לכן, ניתן לחקור באופן מלא כיצד מוליכות מסוימת / זרם אינטראקציה עם מוליכות אנדוגני / זרמים בתוך תאים (איור 1). מודל HN בזמן אמת מציין כי זרם Na+ מתמשך (IP) בנוירונים HN תורם הרבה של Na+ ערך משפיע מאוד [Na+]i (איור 2)ולכן, אנימשאבה. מכיוון ש-IP פעילה בפוטנציאל ממברנה שלילי יחסית, היא מתנגדת לכךשאשאוב אפילו במהלך ה-IBI.
תצפיות אלה מצביעות על כך שזה מאלף לחקור אינטראקציות בין נוירונים מבודדים HN עם מהדק דינמי כפי שנדון בעבר8,9,10. ניסויים אלה (מתמשכים) מבוצעים עם הקלטות מיקרו-ectrode חדות בנוירונים בודדים ומבודדים סינפטיים HN(7) (גנגליון שביעי מנותק מחבל העצבים). עד כה, ניסויים אלה מראים כי התפרצות חזקה משוחזרת בנוירונים HN פעילים טוניק (בשל חדירת microelectrode הציג דליפה) על ידי תוספת משותפת של IP ואנימשאבה עם מהדק דינמי(איור 4). זוהי תצפית חשובה המציינת כי מנגנון מתפרץ זמין נוירונים אלה (גם כאשר דליפה נפגעת) הנובעת מהאינטראקציה של משאבת I ו- IP. תוצאות ראשוניות מצביעות על האינטראקציה המורכבת החזקה שלהם, שניתן לחקור במודל ובניסויים (איור 5).
לסיכום, אנישואב בתגובה לעלייה תקופתית ב[Na+]i במהלך פעילות מתפרצת תורם לקצב הפרץ באמצעות סיום פרץ (הפחתת BD). האינטראקציה של IP ואנימשאבה מהווה מנגנון כי הוא מספיק כדי לתמוך פעילות מתפרצת אנדוגני; מנגנון זה יכול להחזיר התפרצות חזקה ב HN interneurons נרשם תאית גנגליון 7. האינטראקציה בין IP ואנילשאוב דרך [Na+]אני משפיע על תקופת פרץ HN באופן לא מונוטוני ומבטיח חוסן של פרץ אוטונומי. מסקנות אלה עולות בקנה אחד עם ניסויים ומידול במערכות חוליות11,12.

איור 1: פעילות חשמלית של לב עלוקה ויישום של I pump and IP עם מלחציים דינמיים. (A)פעילות מתפרצת רגילה נרשמה בו זמנית, באופן חוץ-תאי (למעלה) ובאופן תאי (למטה), בדופק עלוקה HCO מגנגליון שלישי, סכמטי של הנוירונים המתועדים והקשרים הסינפטיים המעכבים ההדדית שלהם מימין. (B)סכמטי מהדק דינמי בעת הקלטת HN(7) פנימי בגנגליון מבודד 7; הערה אין אינטראקציה סינפטית בין שני HN(7) interneurons. (Ci) מתפוצץ באינטריון HN(7) שנחשף על ידי דליפה. (Cii) פרץ חזק יותר יכול להיות מיוצר על ידי הוספת מהדק דינמי Iמשאבה ( = 0.1 nA), אשר מפצה על הדליפה המושרה microelectrode, אבל מדכא את העירור, ו (1 nS), אשר מגביר את העירור. קווים מקווקווים שחורים מציינים ערכי בסיס. קיצורים: HN = הלב interneuron; HCO = מתנד חצי מרכז; אנימשאבה = זרם כלפי חוץ; IP = נה מתמיד+ זרם; = נה מקסימאלית+/ K+ זרם משאבה; = מוליכות מקסימלית של זרם Na+ המתמשך; Vm = פוטנציאל ממברנה; [נה+] i = ריכוז פנימי של Na+. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: דגם פנימי יחיד של HN המציג עקבות לפוטנציאל הממברנה (Vm), Ih, Ipump, [Na+]i, ו- IP. זרמים היפרפולריים כלפי חוץ הם שליליים, וזרמים מפורק הם חיוביים. קווים מקווקווים שחורים מציינים ערכי בסיס. קיצורים: HN = הלב interneuron; אנימשאבה = זרם כלפי חוץ; IP = נה מתמיד+ זרם; = נה מקסימאלית+/ K+ זרם משאבה; Ih = היפרפולריזציה מופעל זרם פנימי; = מוליכות מקסימלית של זרם Na+ המתמשך; = מוליכות מקסימלית של זרם פנים המופעל על-ידי היפרפולריזציה; Vm = פוטנציאל ממברנה; [נה+] i = ריכוז פנימי של Na+. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: ממשק משתמש גרפי של מודל הלב (HN) בזמן אמת ומהדק דינמי המיושם בלוח עיבוד אותות דיגיטלי. משמאל עליון: תיבות מתמטיקה אדומה הן תיבות פרמטר שנקבעו על-ידי המשתמש עבור המודל בזמן אמת, ואילו תיבות Blue Live הן תיבות פרמטר שנקבעו על-ידי המשתמש המשמשות ב- מהדק הדינאמי. El = פוטנציאל ההיפוך של זרם הדליפה; Gl = מוליכות דליפה; Gh = h-הנוכחי מוליכות מקסימלית; Gp = P מוליכות מקסימלית נוכחית; GCaS = מוליכות מקסימלית זרם סידן איטי; PumpMax = משאבה זרם מקסימלי; [מוליכות סינפטית מקסימלית GSyn2 לנוירונים בהתאמה; סף חציית ספייק ThreshSyn2 לתיווך פוטנציאל סינפטי - אלה המשמשים להכנת מתנד חצי מרכז היברידי (חי / דגם) לא מאויר כאן.]. שמאל תחתון עבור מהדק דינמי. בצד שמאל נמצאים 5 ערכים ממוחשבים של משתני מהדק דינמיים: אנימשאבה = זרם משאבה מוזרק; Ih = h-הנוכחי מוזרק (לא בשימוש כאן); IP = זרם P מוזרק; NaI = מחושב פנימי Na+ ריכוז; ENa = פוטנציאל היפוך נתרן מחושב. שמאל תחתון עבור מהדק דינמי. מימין למשתנים המשושבים יש 6 תיבות פרמטר שנקבעו על ידי המשתמש: GNa = שימוש מרבי בנתרן מלוכסן אנדוגני לחישוב Na+ flux הקשורים לפוטנציאל פעולה; PumpMaxL = זרם משאבה מקסימלי להזרקה על ידי מהדק דינמי; נאי ראה משוואה (2); Gh = מוליכות מקסימלית כדי לקבוע h-זרם להיות מוזרק על ידי מהדק דינמי; Gp = שימוש מוליכות מקסימלית זרם אנדוגני P מניח כדי לחשב Na+ שטף הקשורים זרם P אנדוגני; GpinHNLive = מוליכות מקסימלית כדי לקבוע זרם P להיות מוזרק על ידי מהדק דינמי. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: ניתוח מהדק דינמי של פריצה עצמאית של HN(7). עלייה של מ- (A) 4.0 nS ל (B) 9.0 nS מאטה את קצב פרץ HN העצמאי. עקבות ניסיוניות מראות התפרצות קצבית בנוירונים מבודדים HN(7) עם מהדק דינמי. טווחי התנודה של [Na+]i ו- Vm גדלים עם upregulated . עקבות מלמעלה למטה: Vm מוקלט, מוזרק לימשאבה,מחושב [Na+]i,והזריק IP. קווים מקווקווים שחורים מציינים ערכי בסיס. קיצורים: HN = הלב interneuron; אנימשאבה = זרם כלפי חוץ; IP = נה מתמיד+ זרם; = נה מקסימאלית+/ K+ זרם משאבה; = מוליכות מקסימלית של זרם Na+ המתמשך; Vm = פוטנציאל ממברנה; [נה+] i = ריכוז פנימי של Na+. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
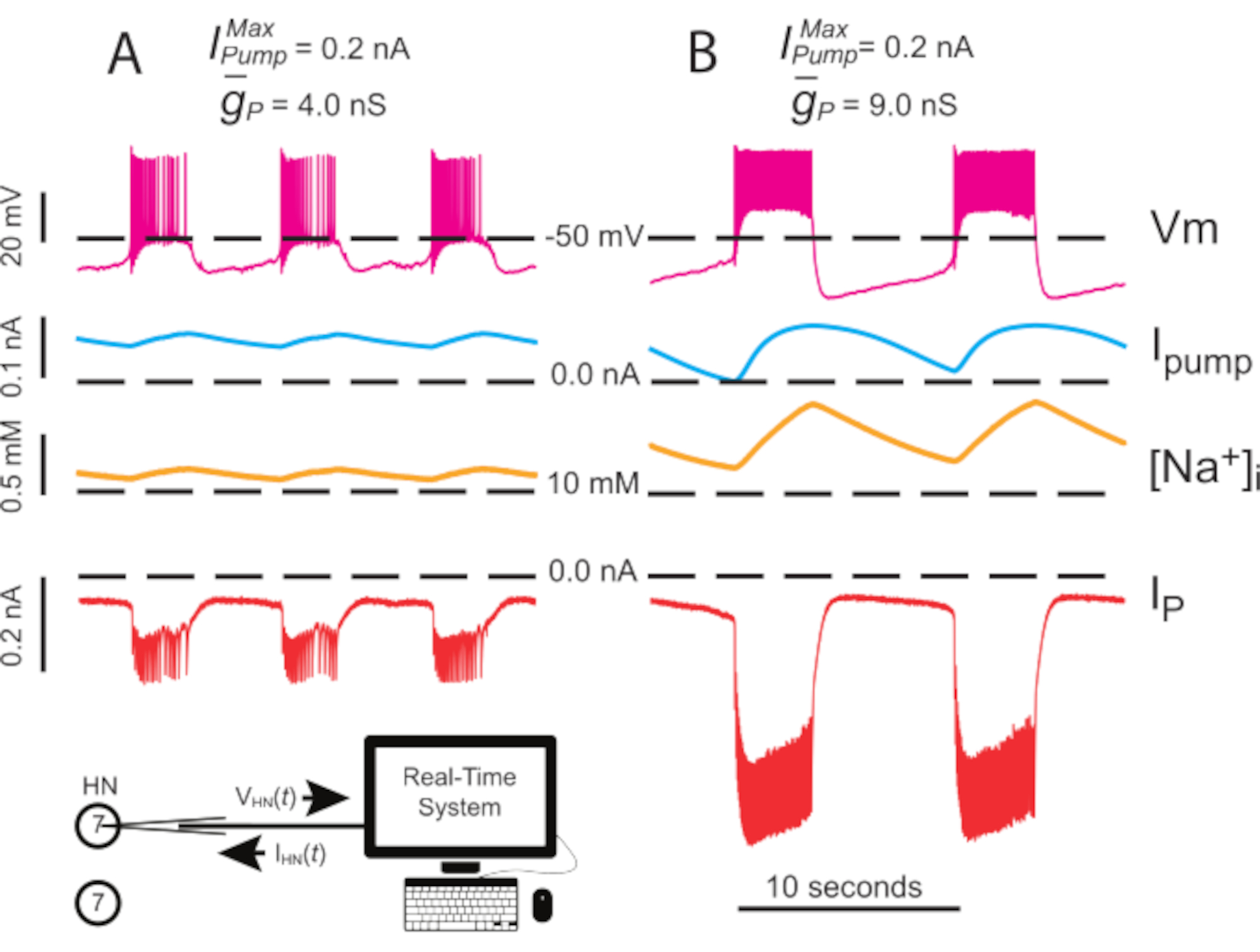{kind=link}

איור 5: ניתוח מהדק דינמי של פריצה עצמאית של HN(7). Upregulation של נוטה להקטין, ואחריו תקופת התפרצות HN מוגברת. בניסויים בודדים (נקודות המחוברות באמצעות קווים) באמצעות מהדק דינמי, ערכים נסחפו בעודם מוחזקים קבועים. צבעים מייצגים רמות קבועות שונות של הוסיף בשימוש בניסויים שונים. קיצורים: HN = הלב interneuron; = נה מקסימאלית+/ K+ זרם משאבה; = מוליכות מקסימלית של זרם Na+ המתמשך. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Discussion
מידול, מהדק דינמי והניתוחים המתקבלים שהם מאפשרים הם טכניקות שימושיות לחקר האופן שבו יחידים וקבוצות של מוליכות/זרמים יוניים תורמים לפעילות החשמלית של תאי העצב (איור 1, איור 2,איור 4 ואיור 5). השימוש בטכניקות אלה מראה כיצד זרם Na+/K+ pump (Ipump) מקיים אינטראקציה עם זרמים מגודרים במתח, במיוחד זרם Na+ המתמשך (IP), כדי לקדם התפרצות חזקה ב- HNs הליבה של מחולל פעימות הלב של עלוקה. על ידי שילוב של ניסויי מלחציים דינמיים ודוגמנות, ניתן לבחון מודלים באופן ישיר יותר מהאפשרי עם הקלטת מתח רגילה וטכניקות מהדק זרם. התוצאות שנאספו מניסויי המלחציים הדינמיים (איור 5) ישמשו כדי לחדד עוד יותר את מודל ה-HN. השיטה הבסיסית של הידוק דינמי המודגם כאן ניתן להתאים אישית כדי לשקף את המאפיינים של כל נוירון תחת מחקר אם מודל מתמטי של זרמים עצביים ניתן לקבוע עם ניסויי מלחצי מתח.
השלמה מוצלחת של הניסויים מהסוג המוצג כאן דורשת שיפוד זהיר של HN או נוירון אחר בעת שימוש במיקרואלקטרודה חדה, מכיוון שהתפרצות חזקה מצטמצמת על ידי חדירת אלקטרודה1. (טכניקות הקלטה של תיקון תאים שלמים, אשר ממזערות את הדליפה שהוצגה, חלות גם על נוירונים אחרים, אך אינן פועלות היטב על נוירונים עלוקה.) זה קריטי כי שיגור של נוירון HN גורם נזק מינימלי הנוירון (דליפה נוספת), והתנגדות קלט צריך להיות במעקב חייב להיות בטווח של 60-100 MOhms לניסויים מוצלחים4.
מהדק דינמי הוא טכניקה רבת עוצמה, אבל יש לו מגבלות שהוטלו על ידי גיאומטריה עצבית כי מוליכות מלאכותיות מיושמות באתר של אלקטרודה הקלטה - בדרך כלל גוף התא - לא באתר שבו זרמים יוצרי קצב הם בדרך כללמקומיים 5,6,10. בנוירונים HN עלוקה, גוף התא קרוב חשמלית לאזור האינטגרציה (נוריט עיקרי) של הנוירון שבו רוב הזרמים הפעילים הם מקומיים, קוצים מופעלים.
Disclosures
ללא
Acknowledgements
אנו מודים לכריסטיאן ארקסלבן על ניסויי מלחציים דינמיים ראשוניים על נוירונים HN(7) שהדגימו את יכולותיהם המתפוצצות. אנג'לה ונינג סייעה לניסויים בעצות מומחים. אנו מכירים NIH למימון עבודה זו באמצעות גרנט 1 R21 NS11355 ל- GSC ו- RLC.
Materials
| Name | Company | Catalog Number | Comments |
| ANIMALS | |||
| Hirudo verbana | Leech.com, https://www.leech.com/collections/live-leeches | live leeches 2-3 grams | |
| CHEMICALS | |||
| ARTIFICIAL POND WATER | |||
| CaCl2 | Sigma Aldrich | C5670-100G | 1.8 mM add last after adjusting pH |
| glucose | Sigma Aldrich | G7021-100G | 10 mM |
| HEPES | Sigma Aldrich | H4034-100G | 10 mM |
| Instant Ocean (sea salt ) | Spectrum Brands Inc., Madison, WI | 0.05% (w/v) diluted in deionized water | |
| KCl | Sigma Aldrich | P9333-500G | 4 mM |
| NaCl | Sigma Aldrich | S7653-250G | 115 mM |
| NaOH 0.1 N Solution | Sigma Aldrich | 2105-50ML | Adjust to pH 7.4 with NaOH |
| MICROELECTRODES | |||
| K Acetate | Sigma Aldrich | P1190-100G | 2 M |
| KCl | Sigma Aldrich | P9333-500G | 20 mM |
| SALINE | |||
| EQUIPMENT | |||
| #5 Forceps | Fine Science Tools Dumont | 11251-30 OR 11251-20 | For general leech dissection |
| AxoClamp 2A/2B DCC electrometer | Axon Instruments Molecular Devices | 2A/2B | For recording of neuronal membrane potential and discontinuous current clamp |
| Black resin | Dow Sylguard | 170 | Lines general dissect dish |
| Capilary glass 1 mm outer diameter, 0.75 mm inner diameter | A-M Systems | 615000 | For fabricating sharp microelectrodes |
| Clear resin | Dow Sylguard | 184 | Lines Petri dish used to mount ganglion for electrophysilogy |
| Dark field condenser | Nikon | Dry 0.95-0.80 MBL 1210 | For illuminating the ganglion preparation during cell impalement |
| Digidata 1440A | Axon CNS Molecular Devices | 1440A | Performs A to D and D to A for data acquisition and stimulation during electrophysiology |
| Digital signal processing board | dSpace | CLP1104 | Our software implements all the conductances/currents in our model HN neuron on a DS1103 dSPACE PPC Controller Board in real-time at a rate of 20 kHz with a ControlDesk GUI (dSPACE, Paderborn, Germany)9. |
| Falming/Brown Microelectrode Puller | Sutter Instruments | P-97 | For fabricating sharp microelectrodes |
| Fiber-Lite high intensity illuminator | Dolan Jenner Industries | 170D | For illuminating the general dissection and for illuminating the ganglion preparation during cell impalement |
| Headstage amplifier for AxoClamp 2A | Axon Instruments | HS-2A Gain:0.1LU | Now part of Molecular Devices for recording of neuronal membrane potential and discontinuous current clamp |
| Light guide | Dolan Jenner Industries | Rev R 38 08 3729107 | For illuminating the general dissection and for illuminating the ganglion preparation during cell impalement |
| Micromanipulator | Sutter Instruments | MPC-385 | Micromanipulator for cell impalement with microelectrodes |
| Micromanipulator controller | Sutter Instruments | MPC-200 | Controls micromanipulators for cell impalement with microelectrodes |
| Minuten pins | BioQuip | 0.15 mm diameter 1208SA | Should be shortened by curtting to ~5 mm |
| Optical Breadboard 3' x 5' x 8" | Newport | Obsolete | With the 4 pneumatic Isolators below used to construct a vibration free workspace for electrophysiology |
| Oscilloscope | HAMEG Instruments | HM303-6 | To monitor electrode setteling during DCC |
| Pascheff-Wolff spring scissors | Moria | Supplied by Fine Science Tools (Foster City, CA) catalog # 15371-92 | |
| pClamp 9 Software | Axon Instruments | 9 | Now part of Moleculear Devices uses the Digidata 1440 for data acquisition and stimulation during electrophysiology |
| Pneumatic Isolators 28" | Newport | Obsolete | With optical breadboard used to construct a vibration free workspace for electrophysiology |
| Simulink / MATLAB software | MathWorks | 2006 (Obsolete) | Implements dynamic clamp on the digital signal processing board |
| Stereomicroscope | Wild | M5A | 10x Eye Pieces used for dissecting the leech and removingand desheathing ganglia |
| Steromicroscope | Wild | M5 | 20x Eye Pieces used in electrophysiologcal station to visualize neuron for microelectrode penetration |
| Student Vannas Spring Scissors | Fine Science Tools | 91500-09 | For general leech dissection |
References
- Cymbalyuk, G. S., Gaudry, Q., Masino, M. A., Calabrese, R. L. Bursting in leech heart interneurons: cell-autonomous and network-based mechanisms. Journal of Neuroscience. 22, 10580-10592 (2002).
- Hill, A. A., Lu, J., Masino, M. A., Olsen, O. H., Calabrese, R. L. A model of a segmental oscillator in the leech heartbeat neuronal network. Journal of Computational Neuroscience. 10, 281-302 (2001).
- Tobin, A. E., Calabrese, R. L. Myomodulin increases Ih and inhibits the NA/K pump to modulate bursting in leech heart interneurons. Journal of Neurophysiology. 94, 3938-3950 (2005).
- Kueh, D., Barnett, W. H., Cymbalyuk, G. S., Calabrese, R. L. Na(+)/K(+) pump interacts with the h-current to control bursting activity in central pattern generator neurons of leeches. eLife. 5, 19322(2016).
- Sharp, A. A., O'Neil, M. B., Abbott, L. F., Marder, E. Dynamic clamp: computer-generated conductances in real neurons. Journal of Neurophysiology. 69, 992-995 (1993).
- Prinz, A. A., Abbott, L. F., Marder, E. The dynamic clamp comes of age. Trends in Neuroscience. 27, 218-224 (2004).
- Barnett, W., Cymbalyuk, G. Hybrid systems analysis: real-time systems for design and prototyping of neural interfaces and prostheses. Biohybrid systems: nerves, interfaces, and machines. Jung, R. , Wiley VCH Verlag. Weinheim. 115-138 (2011).
- Sorensen, M., DeWeerth, S., Cymbalyuk, G., Calabrese, R. L. Using a hybrid neural system to reveal regulation of neuronal network activity by an intrinsic current. Journal of Neuroscience. 24, 5427-5438 (2004).
- Olypher, A., Cymbalyuk, G., Calabrese, R. L. Hybrid systems analysis of the control of burst duration by low-voltage-activated calcium current in leech heart interneurons. Journal of Neurophysiology. 96, 2857-2867 (2006).
- Calabrese, R. L., Prinz, A. A. Realistic modeling of small neuronal networks. Computational Modeling Methods for Neuroscientists. DeSchutter, E. , MIT Press. MA, USA. 285-316 (2010).
- Rybak, I. A., Molkov, Y. I., Jasinski, P. E., Shevtsova, N. A., Smith, J. C. Rhythmic bursting in the pre-Bötzinger complex: mechanisms and models. Progress in Brain Research. 209, 1-23 (2014).
- Picton, L. D., Nascimento, F., Broadhead, M. J., Sillar, K. T., Miles, G. B. Sodium pumps mediate activity-dependent changes in mammalian motor networks. Journal of Neuroscience. 37, 906-921 (2017).
Erratum
Formal Correction: Erratum: Contribution of the Na+/K+ Pump to Rhythmic Bursting, Explored with Modeling and Dynamic Clamp Analyses
Posted by JoVE Editors on 7/28/2021. Citeable Link.
An erratum was issued for: Contribution of the Na+/K+ Pump to Rhythmic Bursting, Explored with Modeling and Dynamic Clamp Analyses. An author name was updated.
The name of the first author was updated from:
Ricardo Javier Erazo Toscano
to:
Ricardo Javier Erazo-Toscano
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved