
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
In vivo הדמיה של רקמת מוח פעילה לחלוטין בזחלי דגי זברה ערים וצעירים על ידי הסרת גולגולת ועור
In This Article
Summary
כאן אנו מציגים שיטה לדימוי המוח העוברי של דג הזברה בשלבים של זחלים ונוער. הליך מיקרו-פולשני זה, המותאם מגישות אלקטרופיזיולוגיות, מספק גישה לפרטים תאיים ותת-תאיים של נוירון בוגר וניתן לשלבו עם אופטוגנטיקה ומחקרים נוירופרמקולוגיים לאפיון תפקוד המוח והתערבות תרופתית.
Abstract
הבנת השינויים הארעיים המתרחשים במהלך התפתחות המוח וההתבגרות דורשת הדמיה מפורטת ברזולוציה גבוהה במרחב ובזמן ברזולוציה תאית ותת-תאית. ההתקדמות בטכנולוגיות מולקולריות והדמיה אפשרה לנו לקבל תובנות מפורטות רבות על מנגנונים תאיים ומולקולריים של התפתחות המוח בעובר דג הזברה השקוף. לאחרונה, תהליכים של עידון קישוריות עצבית המתרחשים בשלבים מאוחרים יותר של זחל מספר שבועות לאחר ההפריה, שהם למשל שליטה על התנהגות חברתית, קבלת החלטות או התנהגות מונעת מוטיבציה, עברו למוקד המחקר. בשלבים אלה, פיגמנטציה של עור דג הזברה מפריעה לחדירת אור לרקמת המוח, ופתרונות לשלבים עובריים, למשל, עיכוב פרמקולוגי של פיגמנטציה, אינם אפשריים יותר.
לכן, פתרון כירורגי זעיר פולשני לגישה מיקרוסקופית למוח של דגי זברה ערים מסופק כי נגזר גישות אלקטרופיזיולוגיות. בטלוסטים, עור וסחוס גולגולת רך ניתן להסיר בזהירות על ידי מיקרו קילוף שכבות אלה, חשיפת נוירונים הבסיסיים ועלונים אקסוניים ללא נזק. זה מאפשר הקלטה של מורפולוגיה עצבית, כולל מבנים סינפטיים ותכולתם המולקולרית, ותצפית על שינויים פיזיולוגיים כגון Ca2 + ארעי או אירועי תחבורה תאיים. בנוסף, חקירה של תהליכים אלה באמצעות עיכוב פרמקולוגי או מניפולציה אופטוגנטית היא אפשרית. גישה זו של חשיפה למוח מספקת מידע על שינויים מבניים ופיזיולוגיים בתאי העצב, כמו גם על המתאם והתלות ההדדית של אירועים אלה ברקמת מוח חיה בטווח של דקות או שעות. הטכניקה מתאימה להדמיה מוחית של זחלי דגי זברה עד 30 יום לאחר ההפריה, השלב ההתפתחותי האחרון שנבדק עד כה. זה, אם כן, מספק גישה לשאלות חשובות כגון עידון סינפטי ושינוי קנה מידה, הובלה אקסונית ודנדריטית, מיקוד סינפטי של מטען ציטוס שלד או ביטוי תלוי פעילות מקומית. לכן, ניתן לצפות לשימוש נרחב בגישת הרכבה והדמיה זו.
Introduction
במהלך העשורים האחרונים, דג הזברה (דניו rerio) התפתח כאחד האורגניזמים מודל החוליות הפופולרי ביותר עבור מחקרים התפתחותיים עובריים וזחלים. פוריותן הגדולה של נקבות דגי הזברה יחד עם התפתחות הרחם האקסית המהירה של העובר ושקיפותו בשלבים התפתחותיים עובריים מוקדמים הם רק כמה גורמים מרכזיים שהופכים את דג הזברה לאורגניזם מודל רב עוצמה כדי להפוך שאלות התפתחותיות1. התקדמות בטכנולוגיות גנטיות מולקולריות בשילוב עם רזולוציה גבוהה במחקרי הדמיה vivo מותר לטיפול מנגנונים ביולוגיים תאים שבבסיס תהליכים התפתחותיים2. בפרט, בתחום ההבחנה העצבית, הפיזיולוגיה, הקישוריות והתפקוד, דגי הזברה שפכו אור על יחסי הגומלין של דינמיקה מולקולרית, תפקודי מוח והתנהגות אורגניגנית בפירוט חסר תקדים.
עם זאת, רוב המחקרים הללו מוגבלים לשלבים עובריים וזחליים מוקדמים במהלך השבוע הראשון של ההתפתחות כמו שקיפות של רקמת מערכת העצבים הולך לאיבוד בהדרגה. בשלבים אלה, רקמת המוח נמנעת מגישה על ידי גישות מיקרוסקופיות ברזולוציה גבוהה להיות מוגן על ידי בידול הגולגולת פיגמנטציה3.
לכן, שאלות מרכזיות של הבחנה עצבית, התבגרות, ופלסטיות כגון עידון של קישוריות עצבית או קנה מידה סינפטי קשה ללמוד. תהליכים תאיים אלה חשובים על מנת להגדיר מנגנונים תאיים המניעים, למשל, התנהגות חברתית, קבלת החלטות או התנהגות מבוססת מוטיבציה, תחומים שבהם מחקר דגי זברה על זחלים בני מספר שבועות תרם לאחרונה ממצאים מרכזיים המבוססים על מחקרים התנהגותיים4.
גישות פרמקולוגיות לעכב פיגמנטציה בזחלי דגי זברה במשך מספר שבועות הם בקושי אפשרי או אפילו עלול לגרום השפעות מזיקות5,6,7,8. זנים מוטנטיים כפולים או משולשים עם פגמים פיגמנטציה ספציפיים, כגון קספר9 או קריסטל10, הפכו לכלים בעלי ערך רב, אך הם מייגעים ברבייה, מספקים צאצאים מעטים ומהווים את הסכנה של צבירת מומים גנטיים עקב הכלאה מופרזת.
כאן, הליך פולשני מינימלי כחלופה מסופק כי הוא חל על כל זן דג זברה. הליך זה הותאם ממחקרים אלקטרופיזיולוגיים כדי לתעד פעילות עצבית בזחלי דגי זברה חיים וערים. בטלוסטים, עור וסחוס גולגולת רך ניתן להסיר בזהירות על ידי מיקרו קילוף שכבות אלה, כי הם אינם שזורים היטב עם כלי המוח. זה מאפשר חשיפת רקמת מוח המכילה נוירונים ודרכיים אקסונליות ללא נזק להקלטת מורפולוגיה עצבית, כולל מבנים סינפטיים ותכולתם המולקולרית, אשר בתורם כוללים תצפית על שינויים פיזיולוגיים כגון Ca2 + ארעיים או אירועי תחבורה תאיים עד מספר שעות. יתר על כן, מעבר לאפיונים תיאוריים, הגישה הישירה לרקמת המוח מאפשרת חקירה של תפקודים עצביים בוגרים באמצעות מתן חומרים נוירופרמקולוגיים וגישות אופטוגנטיות. לכן, יחסי תפקוד מבנה אמיתיים יכולים להתגלות במוח דגי הזברה הצעירים באמצעות אסטרטגיית חשיפה זו למוח.
Protocol
כל עבודת בעלי החיים המתוארת כאן היא בהתאם לתקנות המשפטיות (EU-הוראה 2010/63). תחזוקה וטיפול בדגים אושרו על ידי הרשויות המקומיות ועל ידי נציג רווחת בעלי החיים של תותיית בראונשווייג.
1. הכנת נוזל עמוד שדרה מוחי מלאכותי (ACSF), אגוראז נמס נמוך ומחטי זכוכית חדות
- הכן את ACSF על ידי המסת הכימיקלים המפורטים בריכוזים הבאים במים מזוקקים. 134 מ"מ NaCl (58.44 גרם/ מול), 2.9 מ"מ KCl (74.55 גרם/ מול), 2.1 מ"מ CaCl2 (110.99 גרם/ מול), 1.2 מ"מ MgCl2 6x H2O (203.3 גרם / מול), 10 mM HEPES (238.31 g / mol), ו 10 mM d-גלוקוז (180.16 גרם / מול).
הערה: עבור MgCl2, CaCl2, ו- KCl, פתרונות מלאי 1 M מוכנים במים סטריליים מותפלים ומאוחסנים ב 4 °C (75 °F) להכנת ACSF טרי לאחר מכן. גלוקוז, HEPES ו- NaCl מומסים כתרכובות מוצקות בתמיסת ACSF הטרייה. להמסת כימיקלים, בצע את הוראות היצרן. - התאימו את רמת ה-pH של ה-ACSF ל-7.8 עם 10 M NaOH. הכנת ACSF דורשת מדידה מדויקת של כימיקלים והתאמה עדינה של pH כפי שהוא מחליף את נוזל השדרה במוח ושומר על התנאים הפיזיולוגיים הנדרשים עבור נוירונים להיות פונקציונלי באופן מלא אחר זה עלול לגרום לתעלול מוחי ומוות עצבי.
- יש לאחסן את ה-ACSF שהוכן זה עתה ב-4 °C (75°F) למשך 4 שבועות לכל היותר. עבור תנאי עבודה, aliquot את הנפח הנדרש של ACSF ליום / ניסוי וקדם מלחמה ב 25-28 °C (ובאופן אופציונלי לחמצן אותו, שלב 2.5)
הערה: ASCF מוכן טרי הוא בסדר ליום אחד. אם מתכננים להשתמש בו במשך מספר ימים, ACSF צריך להיות סטרילי מסונן. - להרדמה מאוחרת יותר של הזחלים, להכין פתרון מלאי 50 mM של d-טובוקרין במים מזוקקים ולאחסן את הפתרון ב -20 °C (100 μL aliquots במקפיא עד הצורך.
- כדי להטמיע את הדג, להכין 2.5% נמס נמוך (LM) agarose על ידי המסת 1.25 גרם LM-agarose(שולחן החומרים)ב 50 מ"ל ACSF להרתיח עד agarose מומס לחלוטין.
הערה: לחלופין, ריכוזים גבוהים או נמוכים יותר של LM-agarose ניתן להשתמש בהתאם לסט-אפ ניסיוני. עם זאת, אם האגורוז רך מדי, הוא לא יוכל להחזיק את הדג בעמדה בעת פתיחת הגולגולת. - לאחסן את agarose ב 37 °C מרחץ מים, כדי למנוע התגבשות ובגלל טמפרטורה זו גם לא תפגע בזחלים בעת הטבעה. לאחר האגרוז מבושל מקורר עד 37 °C (50 °F) ב waterbath, להוסיף את הכמות הדרושה של d-Tubocurarine לאגורוז aliquoted הדרוש ליום כדי להגיע לריכוז עבודה של 10 מיקרומטר. לשימוש עתידי, יש לאחסן את האגורוז שנותר ב-4 מעלות צלזיוס כדי למנוע זיהום.
- הכן מחטי זכוכית חדות ודקות מניני זכוכית (איור 1 משלים) באמצעות משיכת מיקרופיפט עם ההגדרות הבאות.
- פולר I, סוג נימי 1: חום 1: 65.8; חום 2: 55.1; משיכה דו-שלית
פולר II, נימי סוג 2: חום = 700; פיל = 4; Vel = 55; דל = 130; פול = 55; משיכה של צעד אחד.
הערה: היחידות ספציפיות עבור כל מושך ונימי זכוכית המשמשיםכאן, בהתאמה (ראה טבלת חומרים ). נימים ומשיכת אחרים יכולים לשמש גם להכנת מחטי הזכוכית. אבל מחטי הזכוכית לא צריך להיות דק מדי כפי שהם עלולים לשבור כאשר באים במגע עם הגולגולת. נימי: אורך: 100 מ"מ (4 אינץ '); מנת יתר: 1.5 מ"מ; תעודת זהות: 0.84 מ"מ; חוט: כן
- פולר I, סוג נימי 1: חום 1: 65.8; חום 2: 55.1; משיכה דו-שלית
2. הרדמה של זחלים והכנות להטמעה
- כאשר מתחילים את הניסוי במשך היום, להעביר את בעלי החיים הדרושים עם פיפטה פסטר פלסטיק צלחת פטרי בקוטר 90 מ"מ, אשר מלא או Danieau (עבור זחלים אשר עדיין נשמרים בצלחת פטרי עם Danieau) או מים ממתקן הדגים (עבור זחלים אשר ישנים יותר מ 7 dpf נשמרים במתקן הדגים).
- בעת צנרת דגים מעל 2 שבועות, ודא את פתיחת הפיפטה גדולה מספיק כדי למנוע פגיעה בדגים בעת העברתם. אין להשתמש ברשת כי זה יפגע פיזית במיוחד הזחלים הצעירים.
- הוסיפו רוטפרה או ארטמיה נאופלי מתאים לגודל הזחלים השמורים בצלחת פטרי, כדי להבטיח גישה חופשית למזון ומצב בריאותי מקסימלי של הזחלים ולהפחית את הלחץ.
- להטמעה, מעבירים את הזחלים שנבחרו לצלחת פטרי בקוטר 35 מ"מ במילוי ACSF. הוסף את הנפח הדרוש של d-Tubocurarine כדי להגיע ריכוז עבודה / מינון יעיל של 10 מיקרומטר ולחכות כמה דקות עד הזחלים משותקים לחלוטין11.
הערה: כאשר הדגים מזדקנים או אם יש צורך בהרדמה מלאה מהירה יותר (מתחת ל -5 דקות), ניתן להגדיל את הריכוז של d-טובוקרין (LD50 לעכברים הוא 0.13 מ"ג/ קילוגרם תוך ורידי12). ניתן גם להשתמש בהרדמה שונה, כגון α-בנגרוטוקסין (ריכוז עבודה: 1 מ"ג / מ"ל), אשר יש את אותה השפעה כמו curare וגם שומר על המוח פעיל לחלוטין13. אם מוח פעיל לחלוטין אינו נחוץ לנושא העניין, Tricaine במינון לא קטלני (0.02%) היא גם אפשרות למרדים את הזחלים באופן מלא. עם זאת, Tricaine חוסם נתרן ערוצים, ובכך פוגע בפעילות המוח14. - הכן את תא ההרכבה על ידי לקיחת המכסה של צלחת פטרי בקוטר 35 מ"מ, להפוך את המכסה הפוך, ומניח כיסוי זכוכית מרובעת (24 x 24 מ"מ) בתחתית המכסה. ראה איור 1 (החלק העליון) לקבלת תיאור סכמטי של שלבים אלה. המשטח החלק יותר של הזכוכית מונע החלקה מבלוק אגרוז, המכיל את הזחלים במהלך הליך פתיחת הגולגולת.
- Aliquot כמות ACSF הדרושה ליום בבקבוקון מתאים (למשל, צינור 50 מל,, בקבוק שוט וכו ') וחמצן אותו עם קרבוגן (5% CO2,95% O2). אם הדמיה רק מורפולוגיה (למשל, דפוסי פלואורסצנטיות) ACSF עדיין יש צורך להבטיח את שלמות המוח וכי תאים אינם מושפעים לרעה על ידי השפעות osmolarity אבל חמצון של ACSF אין צורך. שלב זה צריך להתבצע רק כאשר פעילות מוחית מלאה נחוצה להדמיה.
הערה: לרוויה אופטימלית של חמצן של המדיום, להוסיף אבן אוויר לסוף צינור קרבוגן. כדי להבטיח רמת חמצן גבוהה מספיק, יש צורך להחליף את ACSF בתאי ההדמיה עם ACSF מחומצן טרי כל 20-60 דקות, בהתאם למספר וגיל של זחלים מוטבע באותו תא הדמיה (למשל, עבור חילופי ACSF זחל מוטבע אחד כל שעה מספיק. עבור שישה זחלים מעל 14 dpf מוטבע במקביל, החלפת ACSF כל 20 דקות יש צורך) אז לתכנן את הכמות הדרושה של חמצן רווי ACSF על פי הניסוי המתוכנן.
3. הטמעת הזחלים
- מעבירים את הזחלים המרדים לחלוטין עם פיפטת פסטר מפלסטיק לתא הרכבה מוכן (בשלב 2.4). לאחר מכן, בזהירות להסיר את המדיום עודף כדי למנוע דילול של LM-agarose. כל השלבים הבאים צריכים להתבצע תחת מיקרוסקופ סטריאו עם הגדלה מספקת.
הערה: הטיית תא ההרכבה יכולה לסייע בהסרה מלאה של המדיום. - המשך מיד לשלב הבא, על ידי הוספת טיפת LM-אגרוז גדולה מספיק על גבי הזחלים (בסביבות 1 מ"ל, בהתאם לגודל הזחלים) כדי להגן על בעלי החיים מפני התייבשות ולהפחית מתח מיותר.
- מכוונים את הזחלים בעמדה לפני שהאגורז מתמצה. ודא כי החלק בגב של הזחלים מכוון כלפי מעלה. כמו כן, הקפד להטמיע את הזחלים קרוב לפני השטח של agarose ככל האפשר.
הערה: בהתאם לגודל ולמספר הזחלים המתוכננים להטמיע בו זמנית, ניתן להתאים את ריכוז האגרוז. לדוגמה, עבור 1-3 זחלים כי הם 30 dpf ישן, ריכוז של 1.8%-2% LM-אגרוז מומלץ. עבור 1-4 זחלים בני 7 dpf, זה הכי מעשי להשתמש 2.5% LM-agarose, ואילו, עבור 5-8 זחלים, 2% מתאים יותר. אם נדרש מוח פעיל לחלוטין, מומלץ להטמיע רק שלושה דגים בו זמנית כדי להפחית את הזמן הדרוש להפעלת הזחלים. באופן כללי, מומלץ להשתמש בריכוזים נמוכים יותר (1.8%-2%) ככל שהזחלים ישנים יותר או שהזחלים יותר מתוכננים להיות משובצים בו זמנית. - אם תמונות יצועדו באמצעות מיקרוסקופ הפוך, גזמו את בלוק האגורוז המכיל את הזחלים לצורה קובואידית קטנה. זה חשוב להעברת הזחלים לתא ההדמיה בהמשך. אם משתמשים במיקרוסקופ זקוף, אין צורך בגימור כזה, מכיוון שניתן להשתמש בתא ההרכבה גם כתא ההדמיה. באיור 1 (החלק העליון), ניתן למצוא תיאור סכמטי של שלבים אלה.
4. חשיפת המוח
הערה: כל השלבים הבאים צריכים להתבצע בזהירות רבה כדי לא לפגוע שלא לצורך בזחלים. אם נדרש מוח פעיל לחלוטין לניסוי, יש לזכור כי עם כל שנייה שעוברת, בעוד הדג עדיין מותקן במלואו באגרוז ויש לו גולגולת פתוחה ללא ACSF מחומצן, המוח יסבול מחוסר חמצן וגם להתייבש. ההשפעות של מחסור בחמצן יהפכו אפילו יותר דרמטיות, ככל שהזחלים המוטבעים ישנים יותר. לכן, חשוב לבצע את הניתוח לא רק בתוך הזמן הקצר ביותר האפשרי, אלא גם עם דיוק מרבי לא לעורר נזק מוחי מכני עם המחט. כאשר מאומן, צעדים 4.2-4.4 לא צריך לקחת יותר מ 30 s לכל דג.
- התחל את הניתוח ברגע שהאגורוז יתגבש. ראשית, לקצץ את כל agarose עודף מעל אזור המוח של עניין כדי לקבל גישה חופשית לראש ומרחב עבודה ברור. אם החלק החלק החלקי של הראש כבר בולט מתוך האגורוז, דלג על שלב זה.
- בהתאם לאזור העניין, לבחור מקום להתחיל עם הניתוח. קח את מחט הזכוכית ולעשות חתך קטן דרך העור אבל בלי לחדור עמוק מדי לתוך הרקמה. זו תהיה נקודת המוצא לקילוף העור המניח.
הערה: לקבלת תוצאות אופטימליות, לעולם אל תתחיל ישירות מעל אזור העניין כדי להפחית את הסיכון לפגיעה במבנים חשובים. במידת הצורך, ניתן אפילו להתחיל אחורית אל הבריח האחורי ומשם לעבוד קדימה עד האזור הלא רצוי של העור הוא קלוף משם. - המשך עם חתכים קטנים מאוד לאורך החלק של העור שמטרתם להסיר על ידי בקושי הזזת המחט ממש מתחת לפני השטח. רוב הזמן אין צורך לנוע לחלוטין סביב המוח ולחתוך חתיכה דמוית עיגול של עור וגולגולת, אלא פשוט לעשות שני חתכים לאורך הראש ולאחר מכן לדחוף את העור משם לצד זה או אחר. איור 2 מציג ייצוג סכמטי של אסטרטגיית החיתוך האופטימלית כדי לקבל גישה חופשית למוח הקטן.
הערה: ניתוח מיקרו זה הוא הליך עדין, סביר להניח שהוא יזדקק לאימון כלשהו כדי להסיר את העור בצורה מושלמת מבלי לפגוע במוח הבסיסי. מומלץ גם לגלות את אסטרטגיית החיתוך האופטימלית לאזור המוח של עניין ולהישאר עם זה לתקופת הניסוי. - מיד לאחר הסרת העור מכל הזחלים המוטמעים, המשך על ידי שפיכת ACSF (מחומצן) מעל האגורוז כדי להציף חלקיקי עור ודם לא רצויים ולשמור על המוח פעיל לחלוטין ולהגן עליו מפני התייבשות.
הערה: אם יש צורך במוח בריא לניסוי, מומלץ ללכת על מקסימום של שלושה דגים בכל פעם. - אם אתם משתמשים במיקרוסקופ זקוף, התחל ישירות עם הדמיה.
- בעת שימוש במיקרוסקופ הפוך, החלק מרית קטנה מתחת לבלוק אגרוז קוביות (שלב 3.4).
- מוסיפים טיפה קטנה של LM-agarose לתחתית תא ההדמיה (למשל, צלחת תחתית זכוכית) ומיד להפוך את בלוק אגרוז המכיל את הזחלים עם מרית עבור 180° ולדחוף אותו בעדינות לתחתית תא ההדמיה, בעוד טיפת agarose הנוזל משמש כמו דבק.
- כאשר agarose התגבש, למלא את תא ההדמיה עם ACSF (מחומצן), ולאחר מכן להתחיל הדמיה. ראה איור 1 (חלק תחתון) לקבלת תיאור סכמטי.
- כאשר נדרשת פעילות מוחית מלאה לניסוי, ודא תמיד כי ACSF בתא ההדמיה יש רמת חמצן גבוהה מספיק. כדי להבטיח זאת, החליפו את המדיום בזהירות עם ACSF מחומצן טרי כאשר הדבר אפשרי כל 20-60 דקות (בהתאם למספר וגודל הדג, הגודל והמשטח של תא ההדמיה, ומשך ההדמיה).

איור 1: הליך סכמטי להכנת דגי זברה פתוחים של הגולגולת להדמיית in vivo באופן צעד. הוראות העבודה עבור השלבים השונים ניתן למצוא בגרפיקה עצמה. גרפיקה שעוצבה על ידי פלוריאן הטש ועיבדה את פול שראם. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: ייצוג סכמטי מפורט של המיקרו-ניתוח שבוצע להסרת חתיכות עור וגולגולת מעל אזור העניין במוח. העיגול האדום מסמן את המקום שבו צריך לבצע את החתך הראשון. הקו האדום המקווקו תוחם את הנתיב האופטימלי לחתוך יחד עם המחט כדי לקבל גישה חופשית למוח הקטן מבלי לפגוע בו. החץ הירוק מסמן את הכיוון שבו ניתן לדחוף בקלות את חלקי העור והגולגולת המוגזמים. הקפד לעולם לא לחדור לתוך רקמת המוח במהלך כל ההליך. לאחר קילוף מוצלח של העור, אזור העניין במוח (כאן, המוח הקטן) יהיה נגיש באופן חופשי לכל סוג של הדמיית ויו ברזולוציה גבוהה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
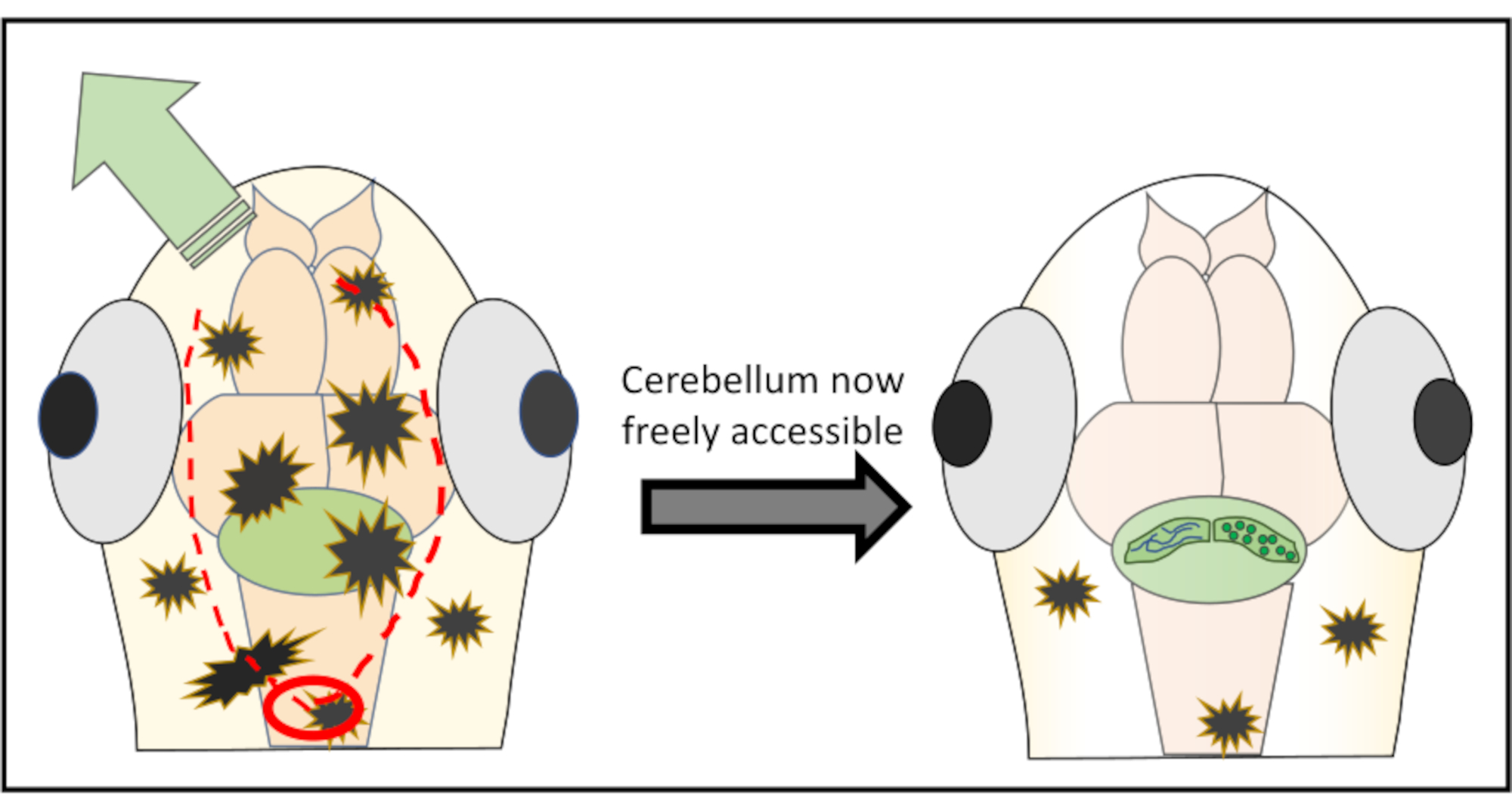{kind=link}
תוצאות
איור 3Aג הצג זחל 14 dpf של הקו הטרנסגני Tg[-7.5Ca8:GFP]bz12[15] כשהגולגולת עדיין שלמה. תאי הפיגמנט בעור השכבת-על מפוזרים על כל הראש ומפריעים לאות הפלואורסצנטיות באזור העניין (כאן, המוח הקטן). עם הזחל במצב זה, לא ניתן להשיג תמונות ברזולוצ...
Discussion
השיטה המוצגת מספקת גישה חלופית לבידוד המוח או לטיפול בזחלי דגי זברה עם תרופות המעכבות פיגמנטציה להקלטת תמונות ברזולוציה גבוהה של נוירונים בסביבת הוויו שלהם. איכות התמונות המתועדת בשיטה זו דומה לתמונות ממוחות מוסברים, אך בתנאים טבעיים.
יתר על כן, אובדן בעוצמת הפלואורס?...
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
אנו מודים במיוחד לטימו פריטש על טיפול מעולה בבעלי חיים והרמן דרינג, מוחמד אלסיי, סול פוזה - מנדז, יעקב פון טרוטה, קומאלי ולישטי וברברה וינטר על תמיכתם המועילה. אנו גם אסירי תודה לכל שאר חברי מעבדת קסטר על המשוב שלהם. הפרויקט מומן בחלקו על ידי קרן המחקר הגרמנית (DFG, KO1949/7-2) פרויקט 241961032 (ל- RWK) ובונדסמיניסטריום פיור בילדונג או פורשונג (BMBF; פרויקט CIPRESS נוירון II ERA-Net 01EW1520 ל- JCM) הוא הודה.
Materials
| Name | Company | Catalog Number | Comments |
| Calcium chloride | Roth | A119.1 | |
| Confocal Laser scanning microscope | Leica | TCS SP8 | |
| d-Glucose | Sigma | G8270-1KG | |
| d-Tubocurare | Sigma-Aldrich | T2379-100MG | |
| Glass Capillary type 1 | WPI | 1B150F-4 | |
| Glass Capillary type 2 | Harvard Apparatus | GC100F-10 | |
| Glass Coverslip | deltalab | D102424 | |
| HEPES | Roth | 9105.4 | |
| Hoechst 33342 | Invitrogen (Thermo Fischer) | H3570 | |
| Imaging chamber | Ibidi | 81156 | |
| Potassium chloride | Normapur | 26764298 | |
| LM-Agarose | Condalab | 8050.55 | |
| Magnesium chloride (Hexahydrate) | Roth | A537.4 | |
| Microscope Camera | Leica | DFC9000 GTC | |
| Needle-Puller type 1 | NARISHIGE | Model PC-10 | |
| Needle-Puller type 2 | Sutter Instruments | Model P-2000 | |
| Pasteur-Pipettes 3ml | A.Hartenstein | 20170718 | |
| Sodium chloride | Roth | P029.2 | |
| Sodium hydroxide | Normapur | 28244262 | |
| Tricain | Sigma-Aldrich | E10521-50G | |
| Waterbath | Phoenix Instrument | WB-12 | |
| 35 mm petri dish | Sarstedt | 833900 | |
| 90 mm petri dish | Sarstedt | 821473001 |
References
- Embryology. Zebrafish Development Available from: https://embryology.med.unsw.edu.au/embryology/index.php/Zebrafish_Development (2020)
- Sassen, W. A., Köster, R. W. A molecular toolbox for genetic manipulation of zebrafish. Advances in Genomics and Genetics. Dove Medical Press. 2015 (5), 151-163 (2015).
- Singh, A. P., Nüsslein-Volhard, C. Zebrafish stripes as a model for vertebrate colour pattern formation. Current Biology. 25 (2), 81-92 (2015).
- Kalueff, A. V., et al. Time to recognize zebrafish 'affective' behavior. Brill: Behaviour. 149 (10-12), 1019-1036 (2012).
- Karlsson, J., von Hofsten, J., Olsson, P. -. E. Generating transparent zebrafish: a refined method to improve detection of gene expression during embryonic development. Marine Biotechnology. 3, 522-527 (2001).
- Bohnsack, B. L., Gallina, D., Kahana, A. Phenothiourea sensitizes zebrafish cranial neural crest and extraocular muscle development to changes in retinoic acid and IGF signaling. PloS One. 6, 22991 (2011).
- Elsalini, O. A., Rohr, K. B. Phenylthiourea disrupts thyroid function in developing zebrafish. Development Genes and Evolution. 212, 593-598 (2003).
- Baumann, L., Ros, A., Rehberger, K., Neuhauss, S. C. F., Segner, H. Thyroid disruption in zebrafish (Danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions. Aquatic Toxicology. 172, 44-55 (2016).
- White, R., et al. Transparent adult zebrafish as a tool for in vivo transplantation analysis. Cell Stem Cell. 2, 183-189 (2008).
- Antinucci, P., Hindges, R. A crystal-clear zebrafish for in vivo imaging. Scientific Reports. 6, 29490 (2016).
- Burr, S. A., Leung, Y. L. Curare (d-Tubocurarine). Encyclopedia of Toxicology (3rd Edition). , 1088-1089 (2014).
- Gesler, H. M., Hoppe, J. 3,6-bis(3-diethylaminopropoxy) pyridazine bismethiodide, a long-acting neuromuscular blocking agent. The Journal of Pharmacology and Experimental Therapeutics. 118 (4), 395-406 (1956).
- Furman, B. . Alpha Bungarotxin. Reference Module in Biomedical Sciences. , (2018).
- Attili, S., Hughes, S. M. Anaesthetic tricaine acts preferentially on neural voltage-gated sodium channels and fails to block directly evoked muscle contraction. PLoS One. 9 (8), 103751 (2014).
- Namikawa, K., et al. Modeling neurodegenerative spinocerebellar ataxia type 13 in zebrafish using a Purkinje neuron specific tunable coexpression system. Journal of Neuroscience. 39 (20), 3948-3969 (2019).
- Hennig, M. Theoretical models of synaptic short term plasticity. Frontiers in Computational Neuroscience. 7 (45), (2013).
- Wang, Y., et al. Moesin1 and Ve-cadherin are required in endothelial cells during in vivo tubulogenesis. Development. 137, 3119-3128 (2010).
- Hobro, A., Smith, N. An evaluation of fixation methods: Spatial and compositional cellular changes observed by Raman imaging. Vibrational Spectroscopy. 91, 31-45 (2017).
- Knogler, L. D., Kist, A. M., Portugues, R. Motor context dominates output from purkinje cell functional regions during reflexive visuomotor behaviours. eLife. 8, 42138 (2019).
- Hsieh, J., Ulrich, B., Issa, F. A., Wan, J., Papazian, D. M. Rapid development of Purkinje cell excitability, functional cerebellar circuit, and afferent sensory input to cerebellum in zebrafish. Frontier in Neural Circuits. 8 (147), (2014).
- Scalise, K., Shimizu, T., Hibi, M., Sawtell, N. B. Responses of cerebellar Purkinje cells during fictive optomotor behavior in larval zebrafish. Journal of Neurophysiology. 116 (5), 2067-2080 (2016).
- Harmon, T. C., Magaram, U., McLean, D. L., Raman, I. M. Distinct responses of Purkinje neurons and roles of simple spikes during associative motor learning in larval zebrafish. eLife. 6, 22537 (2017).
- Zehendner, C. M., et al. Moderate hypoxia followed by reoxygenation results in blood-brain barrier breakdown via oxidative stress-dependent tight-junction protein disruption. PLoS One. 8 (12), 82823 (2013).
- Dhabhar, F. S. The short-term stress response - mother nature's mechanism for enhancing protection and performance under conditions of threat, challenge, and opportunity. Frontiers of Neuroendocrinology. 49, 175-192 (2018).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved