
このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
目覚めしたゼブラフィッシュ幼虫と頭蓋骨と皮膚除去による若年組織の完全に活性な脳組織のインビボイメージング
要約
ここでは、幼虫および若年期までの生体内でゼブラフィッシュ胚性脳を画像化する方法を提示する。この微侵襲的な手順は、電気生理学的アプローチから適応され、成熟したニューロンの細胞および細胞内の詳細へのアクセスを提供し、脳機能および薬物介入を特徴付けるための光遺伝学および神経薬理学的研究と組み合わせることができる。
要約
脳の発達と成熟の間に起こる一時的な変化を理解するには、細胞および細胞内解像度での空間と時間における詳細な高解像度イメージングが必要です。分子および画像技術の進歩により、透明ゼブラフィッシュ胚における脳の発達の細胞および分子メカニズムに関する多くの詳細な洞察を得ることができています。最近では、受精の数週間後に起こる神経細胞の結合性の改善プロセスは、例えば社会的行動、意思決定、動機づけによる行動の制御など、研究の焦点に移っています。これらの段階では、ゼブラフィッシュ皮膚の色素沈着は、脳組織への光の浸透を妨げ、胚性段階の溶液、例えば、色素沈着の薬理学的阻害は、もはや実現不可能である。
従って、目覚めたゼブラフィッシュの脳への顕微鏡アクセスのための低侵襲外科的解決は、電気生理学的アプローチに由来する提供される。テレオストでは、皮膚および柔らかい頭蓋骨軟骨は、これらの層をマイクロ剥離することによって慎重に除去することができ、根底にあるニューロンおよび軸索管を損傷することなく露出させる。これにより、シナプス構造とその分子内容を含む神経形態、およびCa2+ 過渡性または細胞内輸送事象などの生理学的変化の観察を記録することができます。さらに、薬理学的阻害または光遺伝学的操作によってこれらのプロセスの尋問が可能である。この脳暴露アプローチは、ニューロンの構造的および生理学的変化に関する情報だけでなく、分または時間の範囲で生きている脳組織におけるこれらの事象の相関関係および相互依存性に関する情報を提供する。この技術は、受精後30日までのゼブラフィッシュ幼虫のインビボ脳イメージングに適している、これまでにテストされた最新の発達段階。したがって、シナプスの洗練とスケーリング、軸索および樹状輸送、細胞骨格貨物のシナプスターゲティング、または局所活動依存式などの重要な質問へのアクセスを提供します。したがって、この取り付けおよびイメージングアプローチの幅広い使用が期待できます。
概要
ここ数十年、ゼブラフィッシュ(Danio rerio)は、胚性および幼虫発生研究のための最も人気のある脊椎動物モデル生物の一つとして進化してきました。初期胚発生期の胚の急速な 元子宮 の発達とその透明性と相まってゼブラフィッシュメスの大きな胎児性は、ゼブラフィッシュを発達の問題を和らげるための強力なモデル生物にする重要な要因のほんの一部である1。分子遺伝学的技術の進歩と高分解能 インビボ イメージング研究を組み合わせることで、発達過程の基礎となる細胞生物学的メカニズムに対処することが可能となった2.特に、神経細胞の分化、生理、接続性、機能の分野において、ゼブラフィッシュは、分子動力学、脳機能、および生物的挙動の相互作用をこれまでにない詳細に明示しています。
しかし、これらの研究のほとんどは、神経系組織の透明性が徐々に失われるため、発達の最初の週の間に胚および初期の幼虫段階に限定されている。これらの段階では、脳組織は、頭蓋骨の分化と色素沈着によって遮蔽される高分解能顕微鏡アプローチによってアクセスを妨げられる。
したがって、神経の分化、成熟、および神経の結合性の改良やシナプススケーリングなどの可塑性の主要な問題は研究が困難である。これらの細胞プロセスは、例えば、社会的行動、意思決定、または動機に基づく行動を駆動する細胞メカニズムを定義するために重要であり、数週間前の幼虫に関するゼブラフィッシュ研究が最近行動研究4に基づいて重要な知見を提供している領域。
ゼブラフィッシュ幼虫の色素沈着を数週間阻害する薬理学的アプローチは、ほとんど実現可能ではないか、5、6、7、8の有害な影響を引き起こす可能性さえある。キャスパー9や結晶10のような特定の色素沈着欠陥を有する二重または三重突然変異株は、非常に貴重な道具となっているが、繁殖に骨の折れる、少数の子孫を提供し、過剰な近親交配による遺伝的奇形を蓄積する危険性がある。
ここでは、任意のゼブラフィッシュ株に適用可能な代替として最小限の侵襲的手順が提供される。この手順は、生きていると目覚めたゼブラフィッシュ幼虫の神経活動を記録するために電気生理学的研究から適応されました.テレオストでは、皮膚および柔らかい頭蓋骨軟骨は、脳血管構造と密接に織り交ぜられていないため、これらの層をマイクロ剥離することによって慎重に除去することができる。これにより、ニューロンおよび軸索管を含む脳組織を損傷なく暴露し、シナプス構造とその分子内容物を含む神経形態を記録することができ、Ca2+ 過渡または細胞内輸送事象などの生理学的変化の観察を数時間まで含む。さらに、脳組織への直接的なアクセスは、記述的特徴付け以外にも、神経薬理学的物質の管理と光遺伝学的アプローチによる成熟した神経機能の問い合わせが可能です。したがって、この脳暴露戦略を用いて、真の構造機能関係を若年ゼブラフィッシュ脳で明らかにすることができる。
Access restricted. Please log in or start a trial to view this content.
プロトコル
ここに記載されているすべての動物の仕事は、法的規制(EU指令2010/63)に従っています。魚の維持と取り扱いは、地元当局と技術大学ブラウンシュヴァイクの動物福祉代表によって承認されています。
1. 人工脳脊髄液(ACSF)、低融解アガロース、鋭いガラス針の調製
- 蒸留水中の以下の濃度で記載された化学物質を溶解することによってACSFを準備します。134 mM NaCl (58.44 g/mol), 2.9 mM KCl (74.55 g/mol), 2.1 mM CaCl2 (110.99 g/mol), 1.2 m M MgCl2 6x H2O (203.3 g/mol), 10 mM HEPES (238.31 g/mol), および 10 mM d-グルコース (180.16 g/mol).
注: MgCl2、CaCl2、および KCl の場合、1 M ストック溶液は脱塩された無菌水で調製され、その後新しい ACSF を準備するために 4 °C で保管されます。グルコース、HEPES、およびNaClは、新鮮なACSF溶液中で固体化合物として溶解される。化学物質を溶解するには、製造元の指示に従ってください。 - ACSF の pH を 10 M NaOH で 7.8 に調整します。ACSFの調製には、脳脊髄液を置き換え、ニューロンが完全に機能するために必要な生理学的条件を維持し、脳の誤機能および神経死を引き起こす可能性があるため、化学物質の正確な測定とpHの微調整が必要です。
- 作りたてのACSFを4°Cで最大4週間保管してください。作業条件の場合、日/実験に必要なACSFの量をアリクォートし、25-28 °Cでプリウォームします(必要に応じて、ステップ2.5)
注:作りたてのASCFは1日間で大丈夫です。数日間にわたって使用する場合、ACSF は無菌フィルタ処理する必要があります。 - 幼虫の後の麻酔のために、蒸留水にd-Tubocurarineの50 mMストック溶液を調製し、必要になるまで100μLのアリコートとして-20°Cで溶液を保存します。
- 魚を埋め込むには、1.25gのLM-アガロース(材料表)を50mLACSFに溶解して2.5%低融解(LM)アガロースを調製し、アガロースが完全に溶解するまで沸騰させます。
注:あるいは、LM-アガロースの濃度が高いか低いかは、実験的なセットアップに応じて使用することができます。しかし、アガロースが柔らかすぎると、頭蓋骨を開くときに魚を位置に保持することはできません。 - アガロースは37°Cの水浴で保管し、固化を避け、この温度も埋め込むときに幼虫に害を与えないので保管してください。沸騰アガロースを水浴中で37°Cに冷却した後、10μMの作業濃度に達するために必要なアリクォートアガロースに必要量のd-Tubocurarinを加えます。今後の使用のために、汚染を避けるために、残ったアガロースを4°Cで保管してください。
- 次の設定でマイクロピペットプーラーを使用して、ガラスの毛細い毛細い針(補足図1)から鋭いガラス針を準備します。
- プーラーI、キャピラリータイプ1:ヒート1:65.8;熱2:55.1;2ステップ引っ張る
プーラーII、キャピラリータイプ2:熱=700;フィル = 4;Vel = 55;デル = 130;Pul = 55;1ステップの引っ張り。
注:単位は、ここで使用される各プーラーとガラスキャピラリーごとに固有のものです( 資料表を参照)。他の毛細血管および引き手はまたガラスの針を準備するために使用することができる。しかし、頭蓋骨に接触すると壊れる可能性があるため、ガラス針は薄すぎないようにしてください。毛細管:長さ:100ミリメートル(4インチ)。OD:1.5ミリメートル。ID:0.84ミリメートル。フィラメント: はい
- プーラーI、キャピラリータイプ1:ヒート1:65.8;熱2:55.1;2ステップ引っ張る
2. 幼虫の麻酔と埋め込みのための準備
- その日の実験を開始する際には、プラスチック製のパスツールピペットで必要な動物を直径90mmのペトリ皿に移し、ダノー(まだダノーと一緒にペトリ皿に保管されている幼虫用)または魚の施設からの水(7dpfより古く、魚の施設に保管されている幼虫の場合)で満たされています。
- 2週間以上の魚をピペット化する場合は、ピペットの開口部が魚を移管する際に魚を傷付けないように十分な大きさであることを確認してください。特に若い幼虫に物理的に損傷を与えるので、ネットを使用しないでください。
- ペトリ皿に保管されている幼虫の大きさに適した ロチフェラ または アルテミア ナウプリを加え、食べ物への自由なアクセスと幼虫の最大の健康状態を確保し、ストレスを軽減します。
- 埋め込みのために、選択した幼虫をACSFで満たされた直径35mmのペトリ皿に移す。10 μMの有効量の働き濃度/有効用量に達するために必要な量のd-Tubocurarinを加え、幼虫が完全に固定されるまで数分待ちます11.
注:魚が古くなったり、より速い完全麻酔が必要な場合(5分以下)、d-Tubocurarineの濃度を高めることが可能です(マウスのLD50 は静脈内12mg/kg)。また、α-bungarotoxin(働き濃度:1 mg/mL)などの異なる麻酔薬を使用することも可能であり、これはキュラレと同じ効果を有し、また脳を完全に活性13に保つ。完全に活性な脳が対象の対象に必要でない場合、非致死量のトリケーヌ(0.02%)は、幼虫を完全に麻酔するオプションでもあります。しかしながら、トリケーヌはナトリウムチャネルをブロックし、それによって脳活動14を損なう。 - 直径35mmのペトリ皿の蓋を取って取り付けチャンバーを準備し、蓋を裏返し、蓋の底部に正方形のガラスカバースリップ(24 x 24 mm)を置きます。これらの手順の概略的な説明については、 図 1 (上部) を参照してください。ガラスのより滑らかな表面は、頭蓋骨の開口手順中に幼虫を含むアガロースブロックの滑り落ちを防ぎます。
- アリコートは、適切なバイアル(例えば、50mLチューブ、ビーカー、ショットボトルなど)で一日に必要なACSFの量を、カルボーゲン(5%CO2、95%O2)で酸素化する。もし、画像化が形態(例えば蛍光パターン)だけであれば、ACSFは脳の完全性を確保するために必要であり、細胞は浸透性効果によって悪影響を受けず、ACSFの酸素化は必要ない。このステップは、イメージングに完全な脳活動が必要な場合にのみ実行する必要があります。
注:媒体の最適な酸素飽和を得るために、カルボーゲン管の端にエアストーンを加えます。十分に高い酸素レベルを保証するためには、同じイメージングチャンバーに埋め込まれた幼虫の数と年齢に応じて、20〜60分ごとにイメージングチャンバー内のACSFを酸素化し、20〜60分ごとに交換する必要があります(例えば、1回毎に単一の埋め込まれた幼虫ACSF交換に対して十分です)。14dpfより古い6匹の幼虫が並列に埋め込まれた場合、20分ごとにACSFを交換する必要があります)ので、計画された実験に従って必要な量の酸素飽和ACSFを計画してください。
3. 幼虫の埋め込み
- プラスチック製のパスツールピペットを使用して完全麻酔をした幼虫を(ステップ2.4)準備された取り付けチャンバーに移します。次いで、過剰培地を慎重に除去し、LM-アガロースの希釈を避ける。以下の手順はすべて、十分な倍率でステレオ顕微鏡で行う必要があります。
注:取り付けチャンバーを傾けると、媒体を完全に取り外すことができます。 - 次のステップに進み、幼虫の上に十分に大きなLM-アガロースの滴を加えることによって(幼虫の大きさに応じて1mL近く)動物を乾燥から保護し、不必要なストレスを軽減する。
- アガロースが固まる前の位置に幼虫の向きを変えて下さる。幼虫の後ろ側の部分が上向きであることを確認します。また、幼虫をできるだけアガロースの表面に近づけるようにしてください。
注:同時に埋め込む予定の幼虫の大きさと数に応じて、アガロース濃度を調整することができます。例えば、1-3の幼虫が30dpf古い場合、1.8%-2%のLM-アガロースの濃度が推奨される。1-4古い幼虫の場合、2.5%のLM-アガロースを使用するのが最も実用的ですが、5-8の幼虫では2%がより適しています。完全に活性な脳が必要な場合は、幼虫の操作に必要な時間を短縮するために、同時に3匹の魚のみを埋め込むのが推奨されます。一般に、幼虫が古いほど低濃度(1.8%-2%)を使用するか、幼虫が同時に埋め込まれる予定です。 - 反転顕微鏡を使用して画像を記録する場合は、幼虫を含むアガロースブロックを小さな立方体の形にトリミングします。これは、後で幼虫をイメージングチャンバーに移す上で重要です。直立顕微鏡を用いる場合、このようなトリミングは必要ないが、取り付けチャンバは撮像室としても使用できるからである。 図 1 (上部) では、これらの手順の概略説明を見つけることができます。
4. 脳を暴露する
注:以下の手順はすべて、幼虫を不必要に傷つけないように最大限の注意を払って行ってください。実験に完全に活性な脳が必要な場合は、1秒ごとに魚がアガロースに完全に取り付けられ、酸素化されたACSFのない開いた頭蓋骨を持っている間、脳は酸素不足に苦しみ、また乾燥することを覚えておいてください。酸素欠乏の影響はさらに劇的になり、埋め込まれた幼虫は古くなります。したがって、可能な限り最短時間で行うだけでなく、極めて正確に針で機械的な脳損傷を起こさないという手術を行うことが重要です。訓練を受けた場合, ステップ 4.2 -4.4 魚あたり 30 s 以上を取るべきではありません.
- アガロースが固まったらすぐに手術を開始します。まず、関心のある脳領域の上に余分なアガロースのすべてをトリミングして、頭と明確な作業空間への自由なアクセスを得る。頭部の後ろ側が既にアガロースから突き出ている場合は、このステップをスキップします。
- 関心のある地域に応じて、手術から始める場所を選びます。ガラスの針を取り、皮膚を通して小さな切開を行うが、組織に深く浸透しすぎない。これは、重ね合わせの皮膚を剥がす出発点となります。
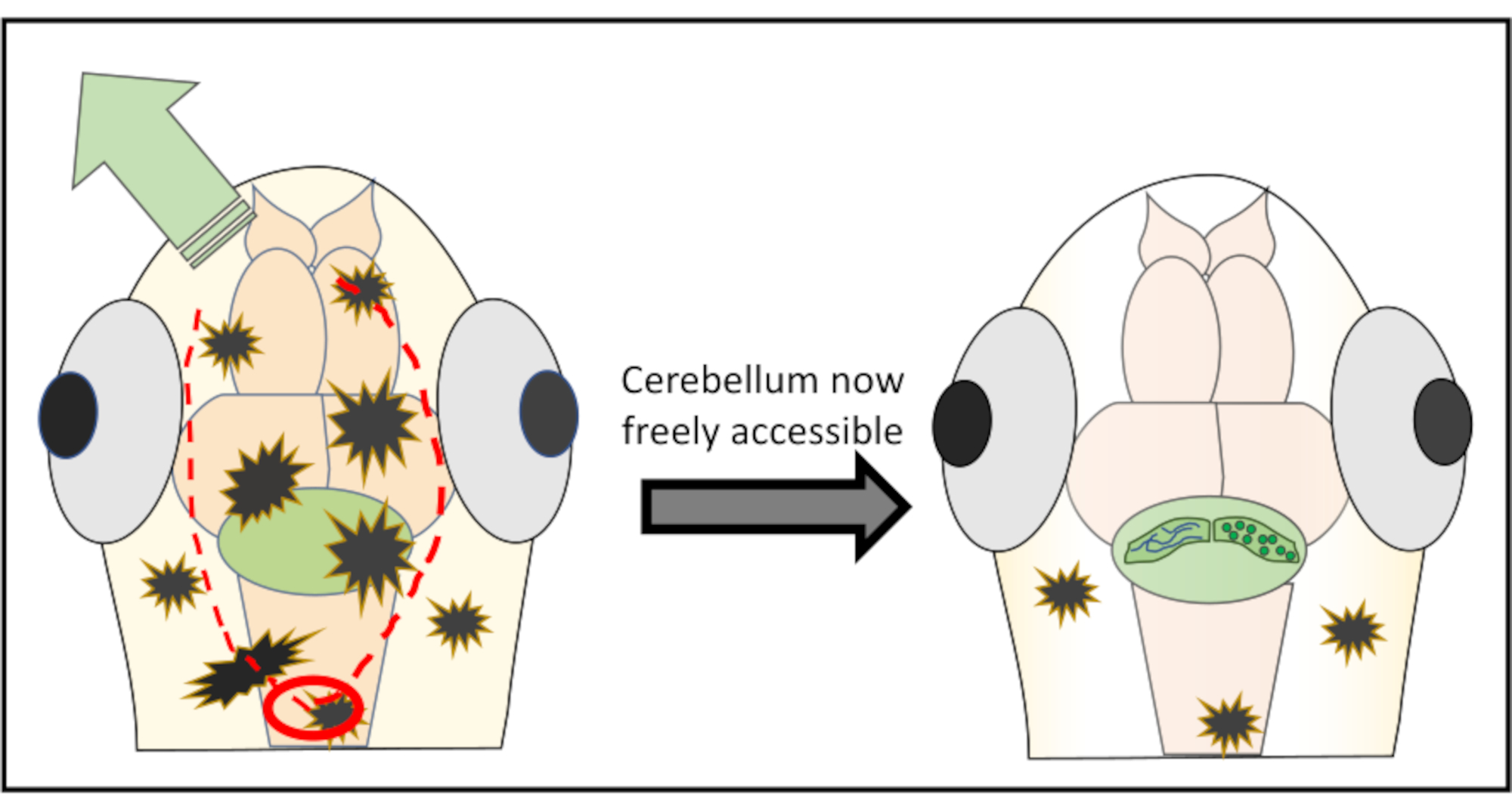
注: 最適な結果を得るには、重要な構造を損傷するリスクを減らすために、対象領域の真上から開始しないでください。必要に応じて、後脳に後部を開始することさえでき、そこから皮膚の不要な領域が剥がれるまで前方に働くことさえできます。 - 表面のすぐ下で針をかろうじて動かして取り除くことを目指して、皮膚の部分に沿って非常に小さなカットを続けます。ほとんどの場合、脳の周りを完全に移動し、円のような皮膚と頭蓋骨を切り取る必要はありませんが、むしろ頭に沿って2つの切開を行い、皮膚を片側または他方の側に押し出す必要があります。 図2 は、小脳への自由なアクセスを得るための最適な切断戦略の概略図を示す。
注:このマイクロ手術は繊細な手順であり、基礎となる脳を損傷することなく皮膚を完全に除去するためのトレーニングが必要になる可能性が最も高いです。また、関心のある脳領域の最適な切断戦略を見つけ、実験の期間に固執することをお勧めします. - 埋め込まれたすべての幼虫から皮膚を取り除いた直後に、アガロースの上にACSFを注いで不要な皮膚粒子や血液をあふれさせ、脳を完全に活性に保ち、乾燥から保護します。
注:実験に健康な脳が必要な場合は、一度に最大3匹の魚を食べます。 - 直立顕微鏡を使用する場合は、イメージングから直接開始します。
- 逆顕微鏡を使用する場合は、小さなへらを立方体アガロースブロックの下にスライドさせます(ステップ3.4)。
- LM-アガロースの小さな滴をイメージングチャンバーの底(例えば、ガラス底皿)に加え、幼虫を含むアガロースブロックをヘラで180°反転させ、撮影室の底にそっと押し込み、液体アガロースドロップは接着剤として機能します。
- アガロースが固まったら、イメージングチャンバーを(酸素化された)ACSFで満たし、次にイメージングを開始します。概略的な説明については 、図 1 (下部) を参照してください。
- 実験に完全な脳活動が必要な場合は、イメージングチャンバーのACSFが十分に高い酸素レベルを有していることを常に確認してください。これを確実にするために、可能な限り20〜60分ごとに酸素化したばかりのACSFと慎重に媒体を交換します(魚の数と大きさ、イメージングチャンバーのサイズと表面、イメージング時間に応じて異なります)。

図1:イン ビボ イメージング用開いた頭蓋骨ゼブラフィッシュを段階的に調製するための概略的手順 さまざまな手順の作業手順は、グラフィック自体にあります。フロリアン・ヘッチがデザインし、ポール・シュラムが脚色したグラフィック。 この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図2:対象となる脳領域の上の皮膚や頭蓋骨の破片を除去するために行われたマイクロ手術の詳細な模式図表 赤い円は、最初のカットを行う必要がある場所を示します。赤い点線は、小脳に損傷を与えることなく、小脳への自由なアクセスを得るために針と一緒に切断するのに最適なパスを表します。緑色の矢印は、過度の皮膚や頭蓋骨の部分を簡単に押し出すことができる方向を示しています。全体の手順の間に脳組織に浸透しないようにしてください。.皮膚の剥がれに成功した後、目的の脳領域(ここでは小脳)は、あらゆる種類の高解像度 インビボ イメージングのために自由にアクセスできるようになります。 この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
Access restricted. Please log in or start a trial to view this content.
結果
図3A、C トランスジェニックラインTg[-7.5Ca8:GFP] の14 dpf幼虫を示すbz12[15] 頭蓋骨はまだそのままです。重ね合わせの皮膚の色素細胞は頭全体に分布し、対象領域(ここでは小脳)における蛍光シグナルに干渉している。この状態では、この状態では、脳の高解像度画像を得ることは不可能です。
Access restricted. Please log in or start a trial to view this content.
ディスカッション
提示された方法は、脳の単離またはそれらの インビボ 環境でニューロンの高解像度画像を記録するための色素沈着を阻害する医薬品とゼブラフィッシュ幼虫の治療に代わるアプローチを提供する。この方法で記録された画像の品質は、自然な条件下で、植えられた脳からの画像に匹敵します。
さらに、固定剤18での治療は必要としないため、蛍?...
Access restricted. Please log in or start a trial to view this content.
開示事項
著者らは開示するものは何もない。
謝辞
特に、ティモ・フリッチの優れた動物ケアとヘルマン・ドーリング、モハメド・エルザイ、ソル・ポーズ・メンデス、ヤコブ・フォン・トロサ、コマリ・ヴァリシェッティ、バーバラ・ウィンターの皆様の支援に感謝します。また、Kösterラボの他のすべてのメンバーの皆様のフィードバックに感謝しています。このプロジェクトの一部は、ドイツ研究財団(DFG、KO1949/7-2)プロジェクト241961032(RWK)とブンデスミニウム・フュル・ビルドゥン・ウント・フォルシュン(BMBF;エラネットニューロンII CIPRESSプロジェクト01EW1520にJCM)が認められる。
Access restricted. Please log in or start a trial to view this content.
資料
| Name | Company | Catalog Number | Comments |
| Calcium chloride | Roth | A119.1 | |
| Confocal Laser scanning microscope | Leica | TCS SP8 | |
| d-Glucose | Sigma | G8270-1KG | |
| d-Tubocurare | Sigma-Aldrich | T2379-100MG | |
| Glass Capillary type 1 | WPI | 1B150F-4 | |
| Glass Capillary type 2 | Harvard Apparatus | GC100F-10 | |
| Glass Coverslip | deltalab | D102424 | |
| HEPES | Roth | 9105.4 | |
| Hoechst 33342 | Invitrogen (Thermo Fischer) | H3570 | |
| Imaging chamber | Ibidi | 81156 | |
| Potassium chloride | Normapur | 26764298 | |
| LM-Agarose | Condalab | 8050.55 | |
| Magnesium chloride (Hexahydrate) | Roth | A537.4 | |
| Microscope Camera | Leica | DFC9000 GTC | |
| Needle-Puller type 1 | NARISHIGE | Model PC-10 | |
| Needle-Puller type 2 | Sutter Instruments | Model P-2000 | |
| Pasteur-Pipettes 3ml | A.Hartenstein | 20170718 | |
| Sodium chloride | Roth | P029.2 | |
| Sodium hydroxide | Normapur | 28244262 | |
| Tricain | Sigma-Aldrich | E10521-50G | |
| Waterbath | Phoenix Instrument | WB-12 | |
| 35 mm petri dish | Sarstedt | 833900 | |
| 90 mm petri dish | Sarstedt | 821473001 |
参考文献
- Hill, M. A. Embryology. Zebrafish Development. , Available from: https://embryology.med.unsw.edu.au/embryology/index.php/Zebrafish_Development (2020).
- Sassen, W. A., Köster, R. W. A molecular toolbox for genetic manipulation of zebrafish. Advances in Genomics and Genetics. Dove Medical Press. 2015 (5), 151-163 (2015).
- Singh, A. P., Nüsslein-Volhard, C. Zebrafish stripes as a model for vertebrate colour pattern formation. Current Biology. 25 (2), 81-92 (2015).
- Kalueff, A. V., et al. Time to recognize zebrafish 'affective' behavior. Brill: Behaviour. 149 (10-12), 1019-1036 (2012).
- Karlsson, J., von Hofsten, J., Olsson, P. -E. Generating transparent zebrafish: a refined method to improve detection of gene expression during embryonic development. Marine Biotechnology. 3, 522-527 (2001).
- Bohnsack, B. L., Gallina, D., Kahana, A. Phenothiourea sensitizes zebrafish cranial neural crest and extraocular muscle development to changes in retinoic acid and IGF signaling. PloS One. 6, 22991(2011).
- Elsalini, O. A., Rohr, K. B. Phenylthiourea disrupts thyroid function in developing zebrafish. Development Genes and Evolution. 212, 593-598 (2003).
- Baumann, L., Ros, A., Rehberger, K., Neuhauss, S. C. F., Segner, H. Thyroid disruption in zebrafish (Danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions. Aquatic Toxicology. 172, 44-55 (2016).
- White, R., et al. Transparent adult zebrafish as a tool for in vivo transplantation analysis. Cell Stem Cell. 2, 183-189 (2008).
- Antinucci, P., Hindges, R. A crystal-clear zebrafish for in vivo imaging. Scientific Reports. 6, 29490(2016).
- Burr, S. A., Leung, Y. L. Curare (d-Tubocurarine). Encyclopedia of Toxicology (3rd Edition). , 1088-1089 (2014).
- Gesler, H. M., Hoppe, J. 3,6-bis(3-diethylaminopropoxy) pyridazine bismethiodide, a long-acting neuromuscular blocking agent. The Journal of Pharmacology and Experimental Therapeutics. 118 (4), 395-406 (1956).
- Furman, B. Alpha Bungarotxin. Reference Module in Biomedical Sciences. , (2018).
- Attili, S., Hughes, S. M. Anaesthetic tricaine acts preferentially on neural voltage-gated sodium channels and fails to block directly evoked muscle contraction. PLoS One. 9 (8), 103751(2014).
- Namikawa, K., et al. Modeling neurodegenerative spinocerebellar ataxia type 13 in zebrafish using a Purkinje neuron specific tunable coexpression system. Journal of Neuroscience. 39 (20), 3948-3969 (2019).
- Hennig, M. Theoretical models of synaptic short term plasticity. Frontiers in Computational Neuroscience. 7 (45), (2013).
- Wang, Y., et al. Moesin1 and Ve-cadherin are required in endothelial cells during in vivo tubulogenesis. Development. 137, 3119-3128 (2010).
- Hobro, A., Smith, N. An evaluation of fixation methods: Spatial and compositional cellular changes observed by Raman imaging. Vibrational Spectroscopy. 91, 31-45 (2017).
- Knogler, L. D., Kist, A. M., Portugues, R. Motor context dominates output from purkinje cell functional regions during reflexive visuomotor behaviours. eLife. 8, 42138(2019).
- Hsieh, J., Ulrich, B., Issa, F. A., Wan, J., Papazian, D. M. Rapid development of Purkinje cell excitability, functional cerebellar circuit, and afferent sensory input to cerebellum in zebrafish. Frontier in Neural Circuits. 8 (147), (2014).
- Scalise, K., Shimizu, T., Hibi, M., Sawtell, N. B. Responses of cerebellar Purkinje cells during fictive optomotor behavior in larval zebrafish. Journal of Neurophysiology. 116 (5), 2067-2080 (2016).
- Harmon, T. C., Magaram, U., McLean, D. L., Raman, I. M. Distinct responses of Purkinje neurons and roles of simple spikes during associative motor learning in larval zebrafish. eLife. 6, 22537(2017).
- Zehendner, C. M., et al. Moderate hypoxia followed by reoxygenation results in blood-brain barrier breakdown via oxidative stress-dependent tight-junction protein disruption. PLoS One. 8 (12), 82823(2013).
- Dhabhar, F. S. The short-term stress response - mother nature's mechanism for enhancing protection and performance under conditions of threat, challenge, and opportunity. Frontiers of Neuroendocrinology. 49, 175-192 (2018).
Access restricted. Please log in or start a trial to view this content.
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved