
Method Article
Rapido isolamento dei Mitoribosome da cellule HEK
In questo articolo
Riepilogo
I mitocondri sono specializzati ribosomi che divergevano dalle loro controparti batteriche e citoplasmatici. Qui vi mostriamo come mitoribosomes può essere ottenuta dal loro vano nativo in cellule HEK. Il metodo prevede l'isolamento dei mitocondri da cellule in sospensione e conseguente purificazione di mitoribosomes.
Abstract
I mitocondri umani possiedono un set dedicato di ribosomi (mitoribosomes) che traducono 13 componenti proteici essenziali dei complessi della fosforilazione ossidativa codificati dal genoma mitocondriale. Poiché tutte le proteine sintetizzate da mitoribosomes umano sono proteine integrali di membrana, mitoribosomes umano sono legati alla membrana interna mitocondriale durante la traduzione. Rispetto al ribosoma citosolico il mitoribosome ha un coefficiente di sedimentazione di 55 anni, metà del contenuto di rRNA, nessun 5S rRNA e 36 altre proteine. Pertanto, un più alto rapporto di proteine-a-RNA e una struttura atipica rendono la mitoribosome umana sostanzialmente distinto dal suo omologo citosolico.
Nonostante l'importanza centrale della mitoribosome alla vita, nessun protocollo era disponibile per purificare il complesso intatto da linee cellulari umane. Tradizionalmente, mitoribosomes sono state isolate da tessuti animali ricche di mitocondri che richiedevano chilogrammi di materiale di partenza. Abbiamo ragionato che mitocondri nella divisione HEK293-derivato umano delle cellule coltivato in un terreno ricco di sostanze nutritive espressione avrebbero una traduzione attiva mitocondriale e, pertanto, potrebbero essere una fonte di materiale per gli studi strutturali e biochimici di il mitoribosome. Per studiare la sua struttura, abbiamo sviluppato un protocollo per la purificazione su grande scala di mitoribosomes intatto da cellule HEK. Qui, presentiamo il metodo cavitazione azoto come un più veloce, meno laborioso e più efficiente alternativa ai tradizionali metodi basati su taglio meccanici per lisi cellulare. Ciò ha provocato preparazioni a base di mitoribosome che ha permesso per la sua determinazione strutturale ad alta risoluzione, rivelando la composizione della mitoribosome umana intatta e suoi intermedi di assemblaggio. Qui, abbiamo follow-up su questo lavoro e presentare un ottimizzato e più metodo conveniente che richiedono solo ~ 1010 coltivate cellule HEK. Il metodo può essere impiegato per purificare umano mitoribosomal tradurre complessi, mutanti, assemblee di controllo di qualità e prodotti intermedi di subunità mitoribosomal. La purificazione può essere linearmente scalata dieci volte se necessario e applicato anche ad altri tipi di cellule.
Introduzione
Il processo della sintesi proteica mitocondriale si basa su 13 mt-mRNAs essenziali che vengono tradotti da una mitoribosome membrana-collegato specializzato per formare il nucleo catalitico della catena respiratoria. Il genoma mitocondriale alterato e la necessità per le proteine di inserire co-translationally nella membrana sostanzialmente hanno modellato l'architettura del ribosomi mitocondriali1. Strutture recenti ad alta risoluzione del mitoribosome dei mammiferi ha mostrato un aspetto sorprendentemente differente alla controparte batterica2,3. In particolare, vengono aggiunti almeno 36 mitocondri specifiche proteine, contribuendo ~ 1 MDa massa extra molecolare, mentre mt-rRNA è ridotto del duplice e altamente divergenti. I riarrangiamenti strutturali alterano quasi tutte le caratteristiche funzionali critiche che in precedenza erano generalmente accettate per essere universalmente conservati4.
Nuovi elementi principali sono stato acquistati da ciascuno delle subunità mitoribosomal, ad esempio, la piccola unità secondaria ha incorporato un'intrinseca mS29 proteine GTPasi nella sua regione di 'testa'. Attività GTPasica non è stato trovato in altri sistemi di traduzione, e la struttura indica che il GTPase potrebbe avere un ruolo nella subunità Assemblea2. il rRNA 5S che credeva di essere un punto di riferimento di tutte le subunità grande ribosomale note, formando il nucleo della protuberanza centrale, manca nella mitoribosome dei mammiferi e ha adottato mt-tRNA-Val come un blocco di costruzione integrale invece2. Divieto e colleghi hanno dimostrato che il mitoribosome suina ha mt-tRNA-Phe e non – Val5. Chrzanowska-Lightowlers, Minczuk e colleghi continuati su questi dati e trovano che mitoribosomes dai pazienti con compromessa mt-tRNA-Val stabilità può, in linea di principio, ospitare mt-tRNA-Phe6,7. Perché i mitoribosomes sono integrate questi elementi specifici e quali percorsi e trans-fattori sono necessari per questi assembly unici rimangono sconosciuti.
Nel complesso, l'elevata complessità della mitoribosome umana, nuovi componenti proteiche e l'associazione unica di mt-tRNA come elemento strutturale implica il coinvolgimento di fattori ancora sconosciuti mitocondrio-specifica trans. Tuttavia, poiché molte delle caratteristiche di questo sistema sono univoche ai mitocondri, che sono stati tradizionalmente difficili da indagare8, piccolo è conosciuto circa il meccanismo molecolare e controllo di qualità. Con lo sviluppo di analisi ad alta risoluzione singola particella elettrone cryo-microscopia (cryo-EM)20, opportunità sorgono ora completamente studiare i meccanismi molecolari sottostanti l'Assemblea, azione e controllo della qualità dell'essere umano mitoribosome. Il nostro rapporto della prima struttura dell'Assemblea umana mitoribosome intermedio fornisce il riconoscimento che è possibile visualizzare come si forma la mitoribosome umana e mostra che cryo-EM è fondamentale per l'identificazione di nuovi trans- recitazione assemblaggio fattori9.
Per espandere su questo sforzo iniziale, descriviamo un protocollo veloce per la purificazione di mitoribosome umana in dettaglio. Nella prima parte del protocollo, è descritto un isolamento su larga scala dei mitocondri altamente puri intatti da cellule in sospensione. Questa procedura richiede 9 h e può essere facilmente modificata e adattata alle scale e diversi tipi di cellule. Un passo significativo in questo protocollo è l'utilizzo di cavitazione di azoto per rompere le cellule. La seconda parte del protocollo è stata sviluppata per purificare mitoribosomes. Questa procedura richiede 7 h e produce una quantità sufficiente di mitoribosomes per gli studi biochimici e strutturali. Utilizzare dei mitocondri puri come materiale di partenza offre preparazioni finali di alta qualità e possa essere estrapolato per altre macromolecole mitocondriale.
Protocollo
Tutti i lavori di cultura di cellule di mammifero devono essere eseguito all'interno di una cappa di sicurezza biologica. Utilizzare attrezzature sterili se il contatto con le cellule. Indossare guanti in nitrile e un camice da laboratorio e seguire la pratica di buona coltura del tessuto.
1. coltura cellulare
- Mantenere le cellule in sospensione HEK293S in di Dulbecco per volta Eagle Medium (DMEM) completati con 10% siero bovino fetale privo di tetraciclina (FBS), 5 μg/mL Dyfed e azitromicina, di 200 μg/mL a 37 ° C e 5% CO2.
- Scala fino a 9 x T175 boccette. Al 90% confluency, raccogliere le cellule e li spin giù a 500 x g per 5 min. Risospendere le cellule pellettate in Freestyl completati con 5% FBS privo di tetraciclina. Contare le celle utilizzando un contatore di cellule automatizzato e regolare la concentrazione di cellule a 1,5 x 106 cellule/mL in un pallone di agitazione ventilato.
Nota: In genere questo corrisponderà ad un volume iniziale di 300 mL a 1.000 mL con ventilato. - Incubare le cellule in un'agitazione incubatore a 37 ° C e 5% di CO2 a 120 giri/min (50 mm di diametro d'agitazione).
- Dopo 2 giorni contare le celle e procedere per dividere le celle se la densità delle cellule è superiore a 3,0 x 106 cellule/mL. Dividere le celle facendo girare giù coltura cellulare 2 x 150 mL a 2 x 250 mL bottiglie coniche a 500 g per 5 min. Risospendere le cellule in media fresco 2 x 10 mL e trasferire su un 2 x 1, 000 mL ventilati beute contenenti il volume appropriato per una densità di cella finale di 1.5 x 106 cellule/mL, ad es. 2 x 300 mL da una densità iniziale delle cellule di 3.0 x 106 cellule/mL.
- Incubare le cellule in un'agitazione incubatore a 37 ° C e 5% di CO2 a 120 giri/min (50 mm di diametro d'agitazione).
- Dopo due giorni contare le celle e procedere per dividere le celle se la densità delle cellule è superiore a 3,0 x 106 cellule/mL. Split le cellule di filatura giù coltura cellulare 4 x 150 mL in 4 x 250 mL bottiglie coniche a 500 g per 5 min. Risospendere le cellule in media fresco 2 x 10 mL e trasferire 2 x 2.000 mL ventilato matraccio contenente il volume appropriato per una densità di cella finale di 1,5 x 106 cellule/mL, ad es. 2 x 700 mL da una densità iniziale delle cellule di 3.0 x 106 cellule/mL.
- Incubare le cellule in un'agitazione incubatore a 37 ° C e 5% di CO2 a 120 giri/min (50 mm di diametro d'agitazione).
- Dopo due giorni contare le celle e procedere per dividere le celle se la densità delle cellule è superiore a 3,0 x 106 cellule/mL. Dividere le celle versando il 2 x 700 mL coltura cellulare in 2 x 2800 mL ventilati boccette e rabbocco con il media fresco 2 x 300 mL per raggiungere un volume di 2 x 1 L.
- Incubare le cellule in un'agitazione incubatore a 37° C e 5% di CO2 a 120 giri/min (50 mm di diametro d'agitazione).
- Dopo 24 h contare le celle e raccogliere le cellule, se la densità delle cellule è tra 3,0-4,0 x 106 cellule/mL.
2. i mitocondri isolamento
- Buffer richiesti
Nota: Le quantità sono date per una preparazione da cellule L 2. Preparare le seguenti soluzioni stock davanti a tempo per buffer di isolamento mitocondriale (MIB), buffer di saccarosio/mannitolo (SM4), buffer sperimentale (MIBSM), tampone di risospensione (SEM).- Buffer di MIB di marca 0,5 L: 50mM HEPES-KOH, pH 7.5, 10 mM KCl, 1.5mM MgCl2, 1 mM EDTA, 1 mM EGTA, 1 millimetro DTT, inibitori della proteasi.
- Fare 100 mL SM4 di tampone: 280mm saccarosio, mannitolo 840 mM, 50 mM HEPES-KOH, pH 7.5, 10 mM KCl, 1.5 mM MgCl2, 1 mM EDTA, 1 mM EGTA, 1 m M DTT, inibitori della proteasi.
- 160 mL di tampone di MIBSM marca: 120 mL di tampone di MIB + 40 mL SM4 di tampone.
- Memorizzare 200 mL di PBS a 4 ° C.
- Rendere 5ml SEM buffer: saccarosio 250 mM, 20 mM HEPES-KOH, pH 7.5, 1 mM EDTA.
- Rendere le soluzioni di riserva 4 x 10 mL per il gradiente di saccarosio graduale contenente 20 mM HEPES-KOH, pH 7.5, 1 mM EDTA e 60% / 32% 23% e 15% saccarosio, rispettivamente.
- Isolamento di mitocondri
Nota: È importante lavorare rapidamente e mantenere tutto su ghiaccio durante tutta la procedura. - Preraffreddare l'azoto cavitazione camera prima dell'uso.
- Raccogliere le cellule L 2 x 1 (3-4 x 109 cellule per provetta da centrifuga) mediante centrifugazione a 1.000 x g per 7 min, 4 ° C.
- Decantare accuratamente il supernatante e risospendere le cellule pellettate rapidamente in 2 x 100 mL di PBS. Piscina le cellule.
- Centrifugare le cellule sedimento a 1.200 x g per 10 min, 4 ° C.
- Decantare il supernatante attentamente e pesare il pellet (~ 20 g).
- Risospendere il pellet in 120 mL di tampone MIB.
- Permettono che le cellule si gonfiano agitando delicatamente in una cella frigorifera per 10 min.
- Posizionare la camera di cavitazione di azoto sul ghiaccio e trasferire le cellule gonfiò alla camera di cavitazione di azoto. Aggiungere tampone di 45 mL SM4 (1/3 del volume finale del sedimento cellule, circa 140 mL, ottenendo una concentrazione finale di mannitolo di saccarosio e 210 mM 70 mM).
- Fissare la camera di azoto cavitazione e riempimento con azoto fino a quando la pressione raggiunge 500 psi. Chiudere i rubinetti e mantenere isolato il ghiaccio per 20 min.
- Lentamente la pressione nella camera di cavitazione di azoto e raccogliere il lisato (circa 185 mL).
- Centrifugare il materiale lisato per rimuovere i detriti cellulari e nuclei a 800 x g per 15 min, 4 ° C.
- Raccogliere il surnatante versandolo attraverso la garza in un becher tenuto su ghiaccio. Non gettare il pellet.
- Risospendere il pellet in 90 mL di tampone di MIBSM (1/2 il volume precedente). Omogeneizzare utilizzando Teflon/vetro lisata omogeneizzatore e ripetere la centrifugazione a 800 x g per 15 min, 4 ° C.
- Raccogliere il surnatante versandolo attraverso la garza in un becher tenuto su ghiaccio. Combinare questo supernatante con il surnatante dal passaggio 2.2.11.
- Centrifugare i surnatanti combinati a 1.000 x g per 15 min, 4 ° C.
- Raccogliere il surnatante versandolo attraverso la garza in un becher tenuto su ghiaccio.
- Scartare il pellet e centrifugare il supernatante contenente i mitocondri grezzi a 10.000 x g per 15 min, 4 ° C.
Nota: Il pellet in genere sarà composto da due parti: sciolto e stretto. - Lavare accuratamente il pellet sciolto senza disturbare la parte stretta. Risospendere il pellet stretto in 10 mL di tampone MIBSM.
- Eseguire un'analisi di concentrazione di proteina usando un commerciale Protein Assay Kit o un metodo simile. Rendimenti di concentrazione tipica da 2L a partire di cultura sono ~ 2 mg/mL.
- Aggiungere 200 unità di RNasi free DNase e farle per ruotare su un rullo in camera fredda per 20 min a mescolare uniformemente la dnasi per la rimozione del DNA genomic.
- Centrifugare a 10.000 x g per 15 min, 4 ° C e risospendere il pellet in buffer di 2 mL SEM. Mescolare delicatamente con un piccolo vetro/Teflon lisata omogeneizzatore per risospendere qualsiasi aggregazione rimanenti. Eseguire non più di cinque passaggi alti e bassi per evitare rotture. Tenere il ghiaccio.
- Preparare il gradiente di saccarosio in 14 mL provette SW40 pipettando attentamente 1,5 mL di buffer stock di 60% saccarosio nella parte inferiore del tubo. Aggiungere con cautela 4,5 mL di buffer stock 32% saccarosio in cima la band 60% senza disturbarla. Ripetere con 1,5 mL di tampone stock 23% di saccarosio e ancora con 1,5 mL di tampone stock 15% di saccarosio.
- Caricare l'intera sospensione mitocondriale (3 mL circa) sopra il gradiente di saccarosio.
- Centrifugare a SW40 rotore a 139, 065 x g per 60 min.
- Raccogliere con cura la fascia marrone la migrazione all'interfaccia del 32% e il 60% saccarosio utilizzando una pipetta di trasferimento, in genere 2-3 mL.
- Snap-congelare i mitocondri purificati in azoto liquido e conservare a-80 ° C.
3. Mitoribosome preparazione
- Buffer richiesti
Nota: Preparare le seguenti soluzioni stock davanti a tempo per il buffer di Lisi, cuscino/gradiente di saccarosio e tampone di risospensione.- Fare 10 mL di tampone di lisi: 25mm HEPES-KOH, pH 7.5, 150 mM KCl, 50mM MgOAc, 2% polietilene glicole octylphenyl etere, 2 mM DTT, inibitori della proteasi.
- Fare 10 mL di tampone per cuscino saccarosio: saccarosio 1m (34% w/v), 20 mM HEPES-KOH, pH 7.5, 100 mM KCl, 20 mM MgOAc, 1% polietilene glicole octylphenyl etere, 2 mM DTT.
- Fare 5 mL di tampone di risospensione: 20 mM HEPES-KOH, pH 7.5, 100 mM KCl, 20mM MgOAc, 2 mM DTT
- Fare 15% - 30% pendenze del saccarosio lineare con tampone di risospensione in tubi in policarbonato TLS-55.
- Purificazione di Mitoribosome
- Scongelare i mitocondri congelati sul ghiaccio.
- Aggiungere 2 volumi di tampone di Lisi,ad esempio aggiungere 6 mL di tampone di lisi dei mitocondri da 3 mL. Mescolare immediatamente capovolgendo la provetta diverse volte.
- Omogeneizzare con un omogeneizzatore di lisata Teflon/vetro piccolo per assistere la lisi e incubare per 5-10 min sul ghiaccio per completare la lisi.
- Centrifugare il materiale lisato (circa 9 mL) a 30.000 x g per 20 min, 4 ° C per rimuovere il materiale insolubile. Decantare il supernatante attentamente dal pellet e scartare il pellet.
- Ripetere la centrifugazione a 30.000 x g per 20 min a 4 ° C per garantire la chiarificazione del surnatante. Decantare il supernatante attentamente dal pellet e scartare il pellet.
- Preparare il cuscino di saccarosio in TLA 120,2 tubi (ultra-chiari): cuscino di saccarosio 0,4 mL per provetta. Preparare un filmato per mL di materiale lisato.
- Strato di lisato mitocondri dei cuscini di saccarosio: circa 1 mL per tubo, risultante in un lisato: rapporto di cuscino di 2,5: 1.
- Centrifugare il campione a 231, 550 x g per 45 min in TLA120.2 rotore a 4 ° C.
- Scartare il surnatante e sciacquare i tubi in modo sequenziale con 100 µ l di tampone di risospensione per rimuovere residuo saccarosio.
- Risospendere il pellet in totale 100 µ l di tampone di risospensione.
- Vortex a bassa velocità per 30 s a sciogliere i restanti aggregati e centrifugare a 17.949 x g per 10 min in conetti centrifuga a 4 ° C.
- Accuratamente raccogliere il surnatante e ripetere la centrifugazione.
- Misurare l'assorbimento del mitoribosome A260.
Nota: La resa tipica è 7 A260, con A260: A280 ratio di 1,3. - Caricare l'intero campione in un tubo di gradiente di saccarosio lineare singolo. Centrifugare a rotore TLS-55 al 213, 626 x g per 120 min a 4 ° C.
- Frazionare il gradiente, determinare la densità ottica a 260 e la piscina la frazione corrispondente al picco di acido nucleico insieme.
Nota: La tipica A260: A280 ratio del picco è > 1.6. - Scambiare il buffer, se necessario, utilizzando un metodo di scelta. Calcolare la concentrazione finale utilizzando la conversione 1 A260 = 0,1 mg/mL.
- Snap congelare il campione di mitoribosome purificato nel tampone di risospensione e conservare a-80 ° C.
Risultati
Cellule di divisione e altamente praticabile è un punto di partenza essenziale per la purificazione di mitoribosomes attivo. Questo protocollo è applicabile a qualsiasi cellule HEK293 in sospensione. Stiamo utilizzando in-House di cellula T501, che è che esprimono stabilmente un trasportatore sotto controllo tetraciclina-inducibile. La linea cellulare parentale è HEK293S-GnTI– cellule (Tabella materiali)10. Durante la crescita delle cellule e l'espansione nel FreeStyle 293 espressione medio della densità minima deve essere tenuta a 1,5 x 106 cellule/mL, al fine di garantire un tasso di raddoppio ogni due giorni, mentre la massima densità di cella non deve superare i 5 ~ x 106 celle / mL. dopo aver raggiunto una densità di cella superiori a 3 x 106 cellule/mL le cellule sono pellettate e risospesi in una media di fresco, preriscaldato in un volume espanso per ottenere la densità cellulare minima di 1.5 x 106 cellule/mL. Questa suddivisione viene eseguita ripetutamente ogni 2-3 giorni fino ad ottenere una massa di cella desiderata per la procedura. La densità di cella finale può variare nel range di 3-4.5 x 106 cellule/mL, e almeno 2 L è necessaria come punto di partenza per l'isolamento dei mitocondri. L'attuabilità delle cellule dovrebbe essere mantenuto generalmente > 90% e per la cultura finale che è raccolto > 95%. Trattamento della coltura delle cellule su larga scala con antibiotici non è raccomandato.
Dopo la serie di centrifugazioni differenziali, il volume della sospensione mitocondriale dopo passo 2.2.20 è tipicamente nella gamma di 3-5 mL. I mitocondri sono quindi separati su gradiente di saccarosio (Figura 1) da altri organelli. Il gradiente di saccarosio graduale è preparato tale che il volume della sfumatura e il volume della sospensione mitocondriale insieme a riempire il tubo di centrifugazione al suo massimo volume. Qui è richiesta particolare attenzione per raccogliere la fascia marrone la migrazione al 60% / 32% interfaccia con contaminazione minima dal buffer circostanti. Questo è importante al fine di mantenere il rapporto costante della proteina: detergente nel seguente passaggio di solubilizzazione di mitocondri. Si consiglia di valutare la concentrazione di proteina mitocondriale in questa fase, e un rendimento tipico del 15-20 mg di proteina mitocondriale per totale è previsto da ~ 1010 coltivate cellule HEK.
All'isolamento di successo mitocondri, vengono lisate mediante l'aggiunta di glicole polietilenico octylphenyl etere e mitoribosomes sono separati attraverso un cuscino di saccarosio (Figura 2). Per separare mitoribosomes dalla sostanza membranosa idrofobica, pellet sono risospese nel buffer non contenente detersivo e idrofobi complessi sono pellettati mediante centrifugazione. Questa procedura è ripetuta, e il grado di purificazione di mitoribosomes è quantificato mediante A260/280 rapporto diA, che dovrebbe essere ~ 1.3. Rendimento tipico è 7 A260 da un 2L inizia la coltura. Questa frazione mitoribosomal contiene anche ulteriori grandi complessi mitocondriali solubile, come della deidrogenasi del piruvato e glutammato deidrogenasi. Per separare mitoribosomes dai complessi mitocondriali solubili, il surnatante viene applicato a un gradiente di densità di saccarosio. Le frazioni contenenti il mitoribosome si trovano generalmente nella parte inferiore terzo del tubo. Il frazionamento del gradiente di saccarosio quindi può essere fatto con un pistone automatico o manualmente prendendo attentamente 50 frazioni µ l con una pipetta o punzonatura sul fondo del tubo con un ago 21G e raccogliendo le gocce.
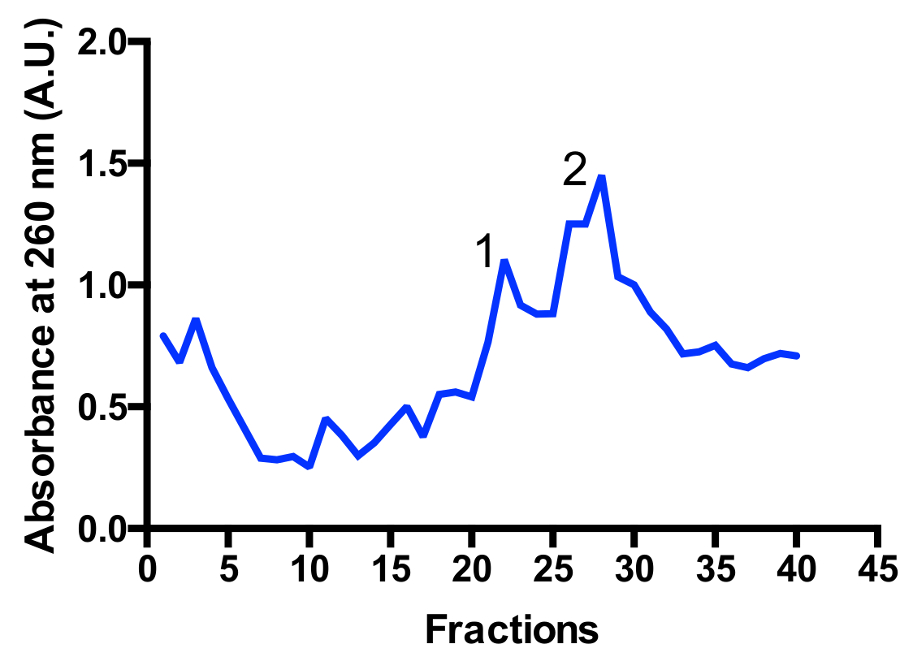
Due popolazioni principali mitoribosomal sono identificati nella sfumatura: monosome 55S e grande unità secondaria 39S, come illustrato in Figura 3 e Figura 4. La presenza della frazione grande unità secondaria suggerisce che le cellule sono raccolte in un altamente attiva divisione stato11. Il rapporto tra il monosome e picchi di grande unità secondaria può cambiare. Un ulteriore picco situato vicino alla parte inferiore può essere visualizzato nelle preparazioni con contaminanti ribosomi citoplasmatici degli anni ' 80. Per favore nota che il gradiente di saccarosio piccolo utilizzando l'oscillazione bucket rotore TLS-55 consente la rapida purificazione di ~ 1 mL di mitoribosomes ad una densità ottica di 0,4-1 un260. La separazione del monosome e picchi di grande unità secondaria può variare leggermente a seconda del frazionamento, ma di solito c'è una certa sovrapposizione dei due picchi. Quindi, quali frazioni da raccogliere, e piscina dovrebbe essere presi in considerazione al fine di garantire la più alta percentuale di monosome, o in alternativa grande unità secondaria, nel campione. Per gli studi di alta risoluzione cryo-EM, la separazione tra le due popolazioni di mitoribosomal non è assolutamente necessaria (a causa di operazioni aggiuntive in silico ). Tuttavia, se una separazione migliore è necessaria, è consigliabile utilizzare più grandi tubi e corrispondenti tempi di esercizio.

Figura 1 : Purificazione dei mitocondri su un gradiente di saccarosio. Organelli subcellulari da un 2L inizia la coltura sono stati frazionati attraverso una serie di centrifugazioni differenziali come descritto nel protocollo, ed i mitocondri sono stati separati su un gradiente di saccarosio discontinuo. I mitocondri purificati sono trovati nella banda inferiore all'interfaccia 32% / 60%.

Figura 2 : Purificazione del mitoribosomes su un cuscino di saccarosio. Mitoribosomes grezzo da una cultura di partenza 2 L vengono sedimentate attraverso 1 M cuscino di saccarosio. I pellet sono risospese in un amplificatore senza detersivo, e mitoribosomes sono chiariti tramite le due centrifugazioni come descritto nel protocollo. L'assorbanza è registrato per valutare la qualità della preparazione e tipici A260/280 rapporto diAdi ~1.3 (pannello di destra) certifica una frazione ricca di mitoribosome. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3 : Fine purificazione del mitoribosomes su un gradiente di saccarosio. Traccia di assorbanza da un 2L inizia la coltura. Le frazioni sono numerate dall'alto verso il basso della sfumatura. Vengono identificate due popolazioni principali mitoribosomal: grande unità secondaria (picco 1) e monosome 55S (picco 2). Potrebbe cambiare il rapporto tra le popolazioni. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4 : Micrografo elettronico, classi 2D e 3D ricostruzione. Pannello di sinistra: una microfotografia con il campione dal picco monosome 2 un ingrandimento calibrato di 1.23 A / pixel. Pannello centrale: Un post-elaborazione dati rappresentativi (classi 2D) rivelando monosomi intatti. Pannello di destra: ricostruzione 3D. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
Per quanto riguarda l'origine delle materie prime, anche se la relativamente facile disponibilità di tessuti animali che sono noti per essere una fonte ricca di mitocondri, lo rende una scelta popolare per mitoribosomes3,5,12, 13, non può essere modificati geneticamente e riprodotto in laboratorio facilmente. Quindi, c'è una necessità pratica chiaro per lo sviluppo di protocolli che coinvolgono linee cellulari umane coltivate in modo omogeneo. La differenza principale tra i protocolli si occupano di tessuti e cellule coltivate è la modalità di lisi e omogeneizzazione. Per le cellule coltivate cresciute come uno strato monomolecolare, un metodo tipico è Teflon/vetro lisata omogeneizzazione14. I protocolli di preparazione originale mentre efficienti sono stati sviluppati per piccole squame15. Un diretto la scalabilità che impiegano un omogeneizzatore con la capacità di 500 mL è possibile11, tuttavia, esso richiede ~ 2 h di lavoro manuale per raggiungere 80% lisi. Ciò ha introdotto almeno tre questioni: l'aggregazione di organelli a causa di lunghi tempi di attesa, Lisi disomogenea a causa di materiale pesante andare a fondo di un recipiente grande, riscaldamento del campione a causa della necessità di più tratti. Di conseguenza, un metodo preferibile di lisi utilizza cavitazione di azoto, che si basa sulla decompressione da un recipiente in pressione16,17. In questo metodo, le cellule sono in primo luogo si gonfiò in camera fredda al fine di ammorbidire le cellule e li rendono più suscettibili alla lisi. Come vengono inseriti nel dispositivo di cavitazione di azoto un buffer contenente saccarosio e mannitolo è aggiunto al fine di mantenere una pressione osmotica che contribuirà a mantenere intatti i mitocondri. Il dispositivo di cavitazione di azoto viene quindi pressurizzato con un grande volume di azoto privo di ossigeno, che si dissolve nelle cellule. Come la pressione è bolle di azoto rilasciato dalla soluzione con conseguente rottura delle membrane cellulari. Questo metodo offre numerosi vantaggi rispetto ai metodi di omogeneizzazione meccanici che comportano sollecitazioni di taglio e attrito come segue: 1) evitare qualsiasi stress fisico esterno sulle cellule; 2) un'espansione adiabatica che raffredda il campione, non assicura nessun danni dovuti al calore di organelli; 3) componenti delle cellule sono protetti dall'ossidazione per il gas inerte di azoto; 4) nessuna alterazione del pH del mezzo di sospensione; 5) il processo è uniforme e riproducibile, perché le stesse forze dirompenti vengono applicate all'interno di ogni cellula e in tutto il campione; 6) il processo è veloce e può essere completato entro 20-30 min.
Isolamento dei mitocondri da cellule coltivate e dei tessuti è stata descritta estesamente nella letteratura, e si basa su una rottura delle cellule dolce seguita da una serie di centrifugazioni differenziali. La maggior parte dei protocolli attualmente utilizzati seguire le procedure originale sviluppate nel mezzo del secolo precedente18. Mentre gli approcci biochimici di base siano corretti, ci sono parecchie idee sbagliate che sono evidenziati nella letteratura ed è rimasto inosservato. Per ottimizzare il protocollo per mitoribosomes, abbiamo studiato i principi generali riportati sistematicamente e concludere che: 1) l'inclusione di Mg2 + e K+ nel buffer non è cruciale. Si è argomentato che il KCl aiuta a impedire la formazione di un gel19, tuttavia, di proteine citoplasmatiche fornito una sufficiente diluizione come descritto nel nostro protocollo, questo fenomeno non si verifica. Inoltre, l'esclusione di Mg2 + è utile per ridurre le contaminazioni di ribosomi citoplasmatici20; 2) non vi è alcuna necessità di mantenere il rapporto di volume massima possibile delle cellule al medium per proteggere gli organelli rilasciati dall'ambiente IPO-osmotica. Il supporto osmotico nel nostro protocollo è sufficientemente fornito da buffer contenenti saccarosio e la diluizione delle cellule con buffer di isolamento mitocondri (fase 2.II.5) è fondamentale per la separazione efficiente di organelli in una preparazione su larga scala.
Il pH del buffer di isolamento mitocondri è vicino ai valori fisiologici, cioè 7.5, che è anche il pH ottimale per la preparazione di mitoribosome seguente. Agenti leganti EDTA ed EGTA vengono aggiunti ai media isolamento per chelare ioni contaminanti, ed essi chelato di magnesio libero e calcio, rispettivamente. Si è discusso che l'inclusione di EDTA può portare a danni della membrana mitocondriale interna14, la concentrazione è pertanto limitata a 1 mM come misura precauzionale. Non abbiamo osservato alcuna differenza quando si utilizza 5 mM EDTA nel passaggio di purificazione finale del gradiente di densità di saccarosio.
Il protocollo descritto nel presente documento utilizza cellule umane derivate da HEK293S per la purificazione della mitoribosome. La qualità della preparazione permette al campione ottenuto di essere studiato biochimicamente e strutturalmente a risoluzione atomica. Questo permette di applicare il metodo di mitoribosomal umano tradurre complessi, assiemi di controllo di qualità e prodotti intermedi di subunità mitoribosomal. Inoltre, poiché le cellule tumorali hanno amplificato capacità OXPHOS e traduzione di proteina mitocondriale elevata rispetto al tessuto adiacente stromal21, mitoribosomes sono bersagli stabiliti per cancro22. Pertanto, utilizzando questo protocollo per purificazione specifico di mitoribosomes in presenza di inibitori avrà applicazioni mediche. Inoltre, mutazioni di mitoribosomal sono state collegate a malattie mitocondriali ereditarie23. Poiché queste mutazioni hanno effetti diretti sulla struttura, l'approccio presentato qui sarà utile per la loro caratterizzazione strutturale. Il protocollo può essere espansa e applicato ad una varietà di domande scientifiche affrontare la comprensione fondamentale della traduzione in mitocondri umani e la sua importanza medica sperimentalmente.
Divulgazioni
Nessuno
Riconoscimenti
Questo lavoro è stato supportato dalla Fondazione svedese per la ricerca strategica (futuro leader Grant FFL15:0325), Ragnar Söderberg Foundation (Fellowship in medicina M44/16), Swedish Research Council (Starting Grant NT × 2015-04107), FEBS Long-Term Fellowship (SA) , Progetto H2020-MSCA-ITN-2016 721757 (VS).
Materiali
| Name | Company | Catalog Number | Comments |
| Dulbecco’s Modified Eagle Medium (DMEM) high glucose, GLUTAMAX supplement, pyruvate | Thermo Scientific | 31966-021 | |
| Fetal Bovine Serum (FBS) | Thermo Scientific | 16000-044 | |
| Blasticidin S HCl | Thermo Scientific | R210-01 | |
| Zeocin selection reagent | Thermo Scientific | R25001 | 100 mg/ml |
| Freestyle 293 Expression medium | Thermo Scientific | 12338026 | |
| T175 tissue culture flask with vented cap | Sarstedt | 83.3912.002 | |
| Shaker flasks with vented cap | Thermo Scientific | 4115-0500, 4115-1000, 4115-2000, 4115-2800 | |
| 250 ml conical bottle tubes, sterile | Corning | 430776 | |
| Eve automated cell counter | NanoEnTek | E1000 | |
| Nitrogen cavitation cell disruption vessel | Parr Instruments | 4635, 4639 | safety volume: 40ml, 600ml respectively |
| Dnase (RNA-free) | HT Biotechnology | N401a | |
| Teflon/glass dounce homogenizers | Cambridge Glassblowing Limited | Size designed upon request | |
| SW40 tubes for mitochondria gradient | Beckman | 344060 | Polypropylene, thin wall |
| Transfer pipettes | Sarstedt | 86.1171 | |
| TLA 120.2 tubes for cushion | Beckman | 343778 | Polycarbonate, thick wall |
| TLS-55 tubes for gradient | Beckman | 347356 | Ultra-clear |
| Gradient Station IP | BioComp | 153-002 |
Riferimenti
- Ott, M., Amunts, A., Brown, A. Organization and regulation of mitochondrial protein synthesis. Annual review of biochemistry. 85, 77-101 (2016).
- Amunts, A., Brown, A., Toots, J., Scheres, S. H., Ramakrishnan, V. The structure of the human mitochondrial ribosome. Science. 348 (6230), 95-98 (2015).
- Greber, B. J., Bieri, P., Leibundgut, M., Leitner, A., Aebersold, R., Boehringer, D., Ban, N. The complete structure of the 55S mammalian mitochondrial ribosome. Science. 348 (6232), 303-308 (2015).
- Greber, B. J., Ban, N. Structure and function of the mitochondrial ribosome. Annual review of biochemistry. 85, 103-132 (2016).
- Greber, B. J., Boehringer, D., Leitner, A., Bieri, P., Voigts-Hoffmann, F., Erzberger, J. P., Ban, N. Architecture of the large subunit of the mammalian mitochondrial ribosome. Nature. 505 (7484), 515-519 (2014).
- Rorbach, J., Gao, F., Powell, C. A., D'Souza, A., Lightowlers, R. N., Minczuk, M., Chrzanowska-Lightowlers, Z. M. Human mitochondrial ribosomes can switch their structural RNA composition. Proceedings of the National Academy of Sciences. , 201609338 (2016).
- Chrzanowska-Lightowlers, Z., Rorbach, J., Minczuk, M. Human mitochondrial ribosomes can switch structural tRNAs-but when and why?. RNA biology. 14 (12), 1668-1671 (2017).
- Gammage, P. A., Moraes, C. T., Minczuk, M. Mitochondrial Genome Engineering: The Revolution May Not Be CRISPR-Ized. Trends in Genetics. , (2017).
- Brown, A., Rathore, S., Kimanius, D., Aibara, S., Bai, X. C., Rorbach, J., Ramakrishnan, V. Structures of the human mitochondrial ribosome in native states of assembly. Nature Structural and Molecular Biology. 24 (10), 866 (2017).
- Reeves, P. J., Callewaert, N., Contreras, R., Khorana, H. G. Structure and function in rhodopsin: high-level expression of rhodopsin with restricted and homogenous N-glycosylation by tetracycline-inducible N-acetylglucosaminyltransferase I-negative HEK293S stable mammalian cell line. Proceedings of National Academy of Sciences USA. 99, 13419-13424 (2002).
- Brown, A., Amunts, A., Bai, X. C., Sugimoto, Y., Edwards, P. C., Murshudov, G., Ramakrishnan, V. Structure of the large ribosomal subunit from human mitochondria. Science. 346 (6210), 718-722 (2014).
- O'Brien, T. W., Kalf, G. F. Ribosomes from rat liver mitochondria I. Isolation procedure and contamination studies. Journal of Biological Chemistry. 242 (9), 2172-2179 (1967).
- Spremulli, L. L. Large-scale isolation of mitochondrial ribosomes from mammalian tissues. Mitochondria: Practical Protocols. , 265-275 (2007).
- Rice, J. E., Lindsay, J. G., Grahamand, J. M., Rickwood, D. Subcellular fractionation of mitochondria. Subcellular Fractionation: A Practical Approach. , 107-142 (1997).
- Attardi, G., Ching, E. Biogenesis of mitochondrial proteins in HeLa cells. Methods in enzymology. 56, 66-79 (1979).
- Gottlieb, R. A., Adachi, S. Nitrogen cavitation for cell disruption to obtain mitochondria from cultured cells. Methods in enzymology. 322, 213-221 (2000).
- Simpson, R. J. Disruption of cultured cells by nitrogen cavitation. Cold Spring Harbor Protocols. 2010 (11), (2010).
- Kennedy, E. P., Lehninger, A. L. Oxidation of fatty acids and tricarboxylic acid cycle intermediates by isolated rat liver mitochondria. Journal of Biological Chemistry. 179 (2), 957-972 (1949).
- Graham, J. M. Isolation of mitochondria from tissues and cells by differential centrifugation. Curr Protoc Cell Biol. , (2001).
- Amunts, A., Brown, A., Bai, X. C., Llácer, J. L., Hussain, T., Emsley, P., Ramakrishnan, V. Structure of the yeast mitochondrial large ribosomal subunit. Science. 343 (6178), 1485-1489 (2014).
- Sotgia, F., Martinez-Outschoorn, U. E., Howell, A., Pestell, R. G., Pavlides, S., Lisanti, M. P. Caveolin-1 and cancer metabolism in the tumor microenvironment: markers, models, and mechanisms. Annual Review of Pathology: Mechanisms of Disease. 7, 423-467 (2012).
- Škrtić, M., Sriskanthadevan, S., Jhas, B., Gebbia, M., Wang, X., Wang, Z., Lai, C. K. Inhibition of mitochondrial translation as a therapeutic strategy for human acute myeloid leukemia. Cancer cell. 20 (5), 674-688 (2011).
- Boczonadi, V., Horvath, R. Mitochondria: impaired mitochondrial translation in human disease. The international journal of biochemistry & cell biology. 48, 77-84 (2014).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati