
Method Article
Dissezione, cultura e analisi delle cellule staminali primarie dal mouse per lo studio della delaminazione e della migrazione delle cellule creste neurali
* Questi autori hanno contribuito in egual misura
In questo articolo
Riepilogo
Questo protocollo descrive la dissezione e la coltura delle cellule creste neurali craniche dai modelli murini, principalmente per lo studio della migrazione cellulare. Descriviamo le tecniche di imaging dal vivo utilizzate e l'analisi dei cambiamenti di velocità e forma delle cellule.
Abstract
Negli ultimi decenni c'è stata una maggiore disponibilità di modelli murini geneticamente modificati utilizzati per imitare le patologie umane. Tuttavia, la capacità di studiare i movimenti cellulari e la differenziazione in vivo è ancora molto difficile. Le neurocristopatie, o disturbi del lignaggio della cresta neurale, sono particolarmente difficili da studiare a causa della mancanza di accessibilità delle fasi embrionali chiave e delle difficoltà nel separare la cresta neurale mesenchyme dal mesenchyme mesodermico adiacente. Qui, abbiamo deciso di stabilire un protocollo di routine ben definito per la coltura delle cellule primarie di cresta neurale cranica. Nel nostro approccio sezionamo il bordo della piastra neurale del topo durante la fase iniziale di induzione della cresta neurale. La regione di confine della piastra neurale viene espiantata e coltivata. Le cellule della cresta neurale si formano in un foglio epiteliale che circonda il bordo della piastra neurale, e da 24 h dopo l'espianto, iniziano a delaminare, sufacendo una transizione epiteliale-mesenchymal (EMT) per diventare cellule della cresta neurale completamente motile. Grazie al nostro approccio di coltura bidimensionale, le popolazioni di tessuti distinti (piastra neurale contro cresta neurale premigratoria e migratoria) possono essere facilmente distinte. Utilizzando approcci di imaging dal vivo, possiamo quindi identificare i cambiamenti nell'induzione della cresta neurale, nell'EMT e nei comportamenti migratori. La combinazione di questa tecnica con mutanti genetici sarà un approccio molto potente per comprendere la biologia delle cellule della cresta neurale normale e patologica.
Introduzione
Il lignaggio della cresta neurale (NC) è una popolazione transitoria, multipotente e migratoria di cellule che appare esclusivamente nei vertebrati durante lo sviluppo embrionale precoce1,2. I derivati della cresta neurale sono estremamente diversi e includono glia, muscolo liscio, melanociti, neuroni e ossa craniofacciali e cartilagine3,4. Poiché la cresta neurale contribuisce alla funzione di molti sistemi di organi, questo lignaggio è essenziale per l'embriogenesi umana. Lo sviluppo di Aberrant NC è implicato in una vasta gamma dei più comuni difetti di nascita umana (ad esempio labbro e palato)5, e anche disturbi come la malattia di Hirschsprung (HSCR), la sindrome di Wardensburg (WS), la sindrome di CHARGE e la sindrome di Williams6 ,7,8,9.
Lo sviluppo NC è stato esplorato in una serie di sistemi modello non mammiferi tra cui modelli Xenopus, pulcino e pesce zebra. Nei mammiferi, il lavoro nei modelli murini ha identificato alcuni degli eventi genetici chiave alla base dello sviluppo della cresta neurale; tuttavia, è stato più difficile seguire la biologia cellulare della migrazione della cresta neurale, a causa dell'inaccessibilità dell'embrione di topo (rivisto altrove10,11). Inoltre, mentre gli studi su pulcini, Xenopus e pesci zebra hanno stabilito una rete di regolazione genica per NC, gli studi sulla perdita di funzioni in questi modelli animali a volte non presentano un fenotipo comparabile nel topo. Ad esempio, in Xenopus, zebrefish e pulcino, segnalazione Wnt non canonica è uno dei meccanismi cellulari che permette al NC di acquisire la sua capacità migratoria12,13,14,15 . Tuttavia, nel mouse, la perdita di segnalazione Wnt non canonica non sembra influire sulla migrazione16. Poiché la migrazione NC in vivo è stata difficile da rintracciare per lunghi periodi nel topo, non è chiaro se queste differenze di specie riflettano diversi modi di migrazione o differenze nella regolazione molecolare.
Come notato, gli studi NC nel topo sono stati molto impegnativi perché la coltura ex utero degli embrioni è laboriosa. Inoltre, il NC è costantemente in intimo contatto con tessuti adiacenti come mesoderma e neurectoderm. Il recente uso di driver Cre specifici della cresta neurale o coloranti esogeni ci ha permesso di etichettare fluorescentmente il NC migratorio; tuttavia, questi approcci sono ancora limitati. Nonostante più report che descrivano diverse tecniche per visualizzare la migrazione NC17,18, è stato difficile risolvere queste tecniche in una procedura semplice e di routine.
È chiaro che c'è bisogno di tecniche che consentano la manipolazione e la caratterizzazione dei mammiferi NC. Abbiamo concentrato i nostri sforzi sul mouse cranial NC in quanto è il modello primario per lo studio dello sviluppo craniofacciale umano e neurocristopatie. Abbiamo perfezionato il nostro approccio sulla base di diversi rapporti interessanti che descrivono la coltura primaria delle cellule NC19,20,21. Qui, descriviamo accuratamente le tecniche di coltura ottimali per espiantare le cellule NC primarie. Dimostriamo il metodo di imaging delle cellule vive e l'uso ottimale di diverse matrici per rivestire le piastre di coltura. Il nostro protocollo descrive come catturare la migrazione di cellule NC vive utilizzando un microscopio invertito, che è inteso come linea guida per l'uso con altri microscopi, nonché un riepilogo dettagliato delle nostre analisi cellulari.
Il risultato atteso dall'espianto dovrebbe essere una distribuzione ben disposta delle cellule che si distinguono chiaramente al microscopio, dove si possono vedere tre diverse popolazioni di cellule che rappresentano (i) piastra neurale, (ii) premigratory, e, (iii) cellule di cresta neurale migratorie. Dimostriamo come analizzare i comportamenti cellulari al confine della popolazione premigratoria delle cellule durante la transizione epiteliale-mesenchymal. Abbiamo anche concentrato il nostro impegno sullo studio delle cellule completamente migratorie per la velocità cellulare, la distanza e la morfologia cellulare.
Protocollo
Tutto il lavoro sugli animali è stato sottoposto ad approvazione etica dal King's College London Ethical Review Process ed è stato eseguito in conformità con UK Home Office Project License P8D5E2773 (KJL).
1. Preparazione dei reagenti
- Preparare soluzioni e strumenti generali, tra cui salina buffer salina (PBS), 70% etanolo, strumenti di dissezione (lame di forza e dissezione o aghi sterili), piastre di plastica o vetrini rivestiti con una matrice extracellulare disponibile in commercio ( Idrogel o fibronectina (vedere la Tabella dei materiali) e supporti di cresta neurale (vedi sotto).
- Preparare il mezzo basale della cresta neurale utilizzando il mezzo aquila modificato di Dulbecco (DMEM, 4500 mg/L di glucosio), il 15% del siero bovino fetale (FBS), 0,1 mM minimo di amminoamine medio-essenziale (MEM NEAA 100X), 1 mM di pirevate di sodio, 55 m -mercaptoetanolo, 100 penicillina mL, 100 unità/mL streptomicina e 2 mM L-glutamina.
- Condizionare i supporti durante la notte utilizzando le cellule dell'alimentatore STO inibibili dalla crescita21.
- Preparare le celle STO (vedere la Tabella dei Materiali)dei supporti per contenere il DMEM integrato da 10% FBS e 100 U/mL penicillina, 100 U/mL streptomycin. Far crescere ed espandere le cellule STO fino alla confluenza in flaconi da 25 cm2 rivestiti con 0,1% di gelatina. Applicare 5000 rad di irradiazione gamma.
- Seme circa 3 x 106 cellule inibite dalla crescita su un piatto di 10 cm2 o 25 cm2 fiaschetta (dal punto 1.2.1.1). Aggiungere circa 10-12 mL di cresta neurale mezzo basale e incubare durante la notte.
NOTA: Le cellule di semi possono essere utilizzate per produrre mezzi condizionali per un massimo di 10 giorni. Controllare regolarmente l'aspetto delle celle
- Filtrare il mezzo (dimensione dei pori di 0,22 m) e integrare con 25 ng/mL fattore di crescita fibroblasto di base (bFGF) e 1000 U di fattore inibitore della leucemia (LIF).
NOTA: Conservare a 4 gradi centigradi e utilizzare entro un mese o conservare a -20 gradi centigradi e utilizzare entro 3 mesi.
- Condizionare i supporti durante la notte utilizzando le cellule dell'alimentatore STO inibibili dalla crescita21.
- Rivestire le superfici di coltura dei tessuti con matrice extracellulare.
NOTA: A seconda della domanda biologica posta, la matrice può essere rivestita su piatti di coltura con fondo di vetro, piatti di coltura di tessuti plastici o slittamenti di copertura in vetro. Vedi sotto per le diverse diluizioni di idrogel a base di ECM a seconda del substrato a matrice. La fibronectina è stata testata su piatti con fondo di vetro e copre le liste solo alle concentrazioni specificate di seguito. Qui, ci riferiremo alle superfici rivestite di substrato come "piastre rivestite".- Rivestire le superfici di coltura dei tessuti con idrogel a base elatamente.
NOTA: Mantenere il substrato freddo fino a quando non si sovrappiatto, raffreddando il supporto o mantenendo il ghiaccio.- Scongela l'idrogel a 4 gradi durante la notte. Aggiungere 5 mL di 10% FBS in DMEM a 5 mL di idrogel per un volume finale di 10 mL (vedere la Tabella dei Materiali).
- Rendere l'aliquota di 0,5–1 mL più comode e conservarle a -20 gradi centigradi.
- Scongelare le aliquote idrogel sul ghiaccio.
- Utilizzare una diluizione 1:20 del brodo di idrogel per rivestire la plastica.
- Utilizzare una diluizione 1:5 del brodo di idrogel per rivestire vetrini di vetro e piastre di coltura di tessuti dal fondo di vetro.
NOTA: Diluire l'idrogel nel DMEM freddo. - Applicare un numero sufficiente di idrogel diluito per coprire l'area desiderata su piastre/vetrini e incubare per 30-45 min a 37 gradi centigradi.
- Utilizzare piastre/scivoli rivestiti immediatamente o conservare i vetrini rivestiti a 4 gradi centigradi durante la notte.
- Rimuovere gli eccessi e risciacquare i vetrini con DMEM ad alto glucosio (opzionale) prima dell'uso.
- Rivestire le superfici di coltura dei tessuti con fibronectina.
- Fare aliquote di 1 soluzione di stock di fibronectina mg/mL e conservare a -80 gradi centigradi. Diluire la fibronectina con pbS (dPBS) di Dulbecco ad una concentrazione finale di 1 g/mL.
- Applicare fibronectina sufficiente per coprire l'area desiderata e incubare a temperatura ambiente per 15 min.
- Rimuovere la fibronectina residua e lasciare asciugare il vetro per 30-45 min.
- Sciacquare pozzi o coprire gli slittamenti con DMEM ad alto glucosio (opzionale) prima dell'uso.
- Rivestire le superfici di coltura dei tessuti con idrogel a base elatamente.
2. Giorno 1: dissezione degli embrioni dello stadio di somite precoce
NOTA: utilizzare strumenti sterili e soluzioni sterili. Se è necessaria la genotipizzazione, raccogliere il corpo dell'embrione per l'estrazione del DNA.
- La dissezione della piastra neurale cranica è limitata agli embrioni a 8,5 giorni dopo il coitum (dpc). Selezionare gli embrioni allo stadio di somitia 5-8. Dissezionare l'utero in PBS e tagliare il mesotrium per separare ogni embrione (Figura 1A). La parete muscolare dell'utero si contrae e il tessuto deciduale diventerà visibile (Figura 1B).
NOTA: Mantenere gli embrioni nell'utero in PBS ghiacciato mentre le dissezioni vengono eseguite un embrione alla volta. Spostare gli embrioni con una pipetta Pasteur in vetro in PBS sterile fresco per migliorare la visibilità e ridurre la contaminazione. - Far scorrere le pinze tra lo strato muscolare e il tessuto deciduale e rimuovere lo strato muscolare con una seconda coppia di pinze (Figura 1C).
- Usando le pinze, forare il deciduo ai bordi del palo mesomico e con una seconda coppia di pinze strappare per aprire perpendicolarmente al palo.
- Sbucciare il tessuto deciduale con le pinze per visualizzare la membrana del Reichert.
- Rimuovere con attenzione la membrana di Reichert. Il sacco del tuorlo viscerale diventa visibile e l'embrione può essere visto all'interno (Figura 1D).
- Rimuovere il sacco del tuorlo viscerale e l'amnione (Figura 1E) e posizionare l'embrione per visualizzare la piega della testa (Figura 1F).
- Tagliare la testa sopra il cuore e raschiare via il mesoderma sottostante utilizzando pinze e/o strumenti ciglia per ottenere una piastra neurale pulita (NP) (Figura 1H).
NOTA: Il NP può essere mantenuto intero o diviso lungo l'asse anteroposteriore in modo che ogni lato possa essere placcato singolarmente. Il bordo della piastra neurale può essere ulteriormente tagliato lontano dalla piastra neurale al fine di ridurre al minimo i contributi neuronali agli espianti. - Utilizzare una pipetta pasteur di vetro per trasferire la piastra neurale sezionata su una piastra rivestita di idrogel piena di supporti di cresta neurale condizionati.
- Ruotare delicatamente il piatto per posizionare il NP al centro del pozzo. Questo è importante per massimizzare la qualità della fase per l'imaging a cellule vive (il giorno 2).
- Incubare durante la notte (o al punto temporale desiderato) a 37 gradi centigradi nel 5% di CO2. Le cellule della cresta neurale dovrebbero migrare visibilmente dalla piastra neurale.
NOTA: le celle di solito si attaccano entro 6-8 h. Dopo l'attacco dell'espianto, concedere più tempo per visualizzare le celle di migrazione. Di solito per 24 h dopo espianto, possiamo trovare tre popolazioni distinguibili di cellule. La prima popolazione, al centro dell'espianta, è la piastra neurale (NP). La seconda popolazione, il premigratory NC (pNC), circonda l'NP in un foglio epiteliale di cellule. La terza popolazione, nell'anello esterno, è formata da NC migratori (mNC), che sono di dimensioni maggiori, e appaiono completamente mesenchymal (Figura 2).
3. Giorno 2: Imaging cellulare vivo di cellule craniche
NOTA: l'imaging deve essere eseguito a 24 ore dopo l'espiantamento per creare un'immagine e quantificare in modo ottimale la migrazione delle celle della cresta neurale. Non è necessario aggiornare i supporti di induzione NC prima dell'imaging delle cellule vive. È necessario l'accesso a un microscopio invertito, con uno stadio motorizzato e una camera ambiente incorporata. Utilizzare piatti di coltura dei tessuti multi-well adatti per l'imaging (Tabella dei materiali).
-
Configurazione del microscopio
- Impostare la camera d'ambiente a 37 gradi centigradi e al 5% di CO2.
- Forare un buco nel coperchio del coperchio della piastra di coltura dei tessuti per consentire all'ago di CO2, collegato alla camera di umidificazione CO2, di sedersi all'interno della piastra.
- Posizionare il piatto di coltura dei tessuti nel supporto del campione e nastro adesivo lungo il coperchio della piastra e l'agoCO2 per evitare agitazione durante l'acquisizione multi-bene.
- Accendere il controller del microscopio, il controller dello stage e il software di imaging.
- Concentrarsi sulle cellule NC craniche all'ingrandimento di 10 volte (con l'anello di fase corrispondente nel condensatore selezionato).
- Impostare il contrasto di fase di alta qualità sul microscopio regolando il campo iris diaframma, l'iris diaframma di apertura e il telescopio di centratura, come specificato nel manuale di configurazione del microscopio.
-
Contrasto di fase dell'imaging a celle vive
- Impostare la directory o il percorso del file in cui verranno salvati i file time-lapse.
- Impostare il tempo di esposizione, il binning e l'area della fotocamera.
- Impostare il numero di punti di tempo, la durata dell'imaging e l'intervallo di tempo tra i fotogrammi.
- Per quantificare la capacità migratoria delle cellule NC, impostare il microscopio su un ingrandimento di 10 x, prendendo 1 fotogramma ogni 5 min (217 punti temporali superiori a 18 h). Per quantificare la morfologia cellulare, impostare l'ingrandimento su 40x, prendendo 1 fotogramma / min (61 punti temporali su 1 h). Per quantificare la dinamica lamellipodiale, impostare l'ingrandimento a 40x o 60x ingrandimento, prendendo 1 telaio ogni 10 s (oltre 10 min).
- Per l'imaging multi-pozzo, impostare la fase meccanica per spostarsi tra le posizioni di interesse XY selezionate. Verificare che le celle NC cranial siano a fuoco e che le posizioni dello stage siano corrette.
- Utilizzare il comando Acquisisci per avviare l'imaging time-lapse.
- Una volta completata la creazione di immagini time-lapse, esaminare i dati multidimensionali ed esportare i file .stk per l'analisi.
NOTA: .stk è un file di stack TIFF. - Uscire dal software, spegnere il computer e spegnere i controller dello stage, della fotocamera e del microscopio.
4. Analisi dell'imaging: quantificazione della migrazione delle cellule della cresta neurale
NOTA: Per definire meglio i comportamenti cellulari esposti dalla migrazione delle cellule della cresta neurale cranica murinale, abbiamo analizzato una serie di parametri migratori quantificabili, concentrandosi in particolare sulla capacità migratoria e sulla dinamica della forma cellulare. (1) La migrazione (distanza accumulata) è la lunghezza totale del percorso effettuata dalla cella (m); (2) La migrazione (distanza euclidea) è la distanza in linea retta tra la posizione iniziale e quella finale della cella( (3) La migrazione (velocità cellulare) è la distanza percorsa per cella per unità di tempo (m/min); (4) Forma cella (area di cella) è la superficie totale coperta dalla cella. Impostare la scala del pixel in base al microscopio di imaging. (A - Apx x Npx, dove Unpx è l'area dei pixel e Npx - numero di pixel. Unità:s2; (5) La forma della cella (circolarità cellulare) è la deviazione della forma della cella da un cerchio perfetto che è indicata da un valore di circolarità di 1,0 (4ì (A/P2))in cui A è l'area e la P - perimetro.
-
Tracciamento a cella singola
NOTA: per misurare la migrazione delle celle NC, vengono generate coordinate XY delle singole celle in tutti i fotogrammi time-lapse. Ciò consente una successiva analisi delle misure di distanza, velocità e persistenza della migrazione cellulare.- Aprire ImageJ e importare i dati come file di stack TIFF.
- Fare clic su Analizza . Impostare Scala per calibrare i file .stk in base alle impostazioni del microscopio, lavorando in pixel/m.
- Fare clic su Plugins Proprietà Tracking . Manual Tracking per aprire il plugin di tracciamento manuale delle celle di Image J. Per iniziare il tracciamento delle celle, selezionare Aggiungi traccia.
- Traccia le celle attraverso tutti i fotogrammi dei filmati time-lapse, usando il nucleo come punto di riferimento.
NOTA: è necessario tenere traccia di 10-20 celle per espianto, per un totale di 60 celle tracciate (n . 3). Le cellule che subiscono la divisione cellulare nel corso del time-lapse dovrebbero essere escluse dall'analisi. - Salvare ed esportare i risultati come file CSV. I risultati rappresentano il numero della traccia cellulare, il numero di sezione e le coordinate XY su tutti i fotogrammi.
-
Quantificazione della capacità migratoria delle cellule creste neurali
- Aprire i dati di tracciamento a singola cella (vedere sopra). Convertire i file .csv in formato .txt.
- Aprire il software dimigrazione (tabella dei materiali ). Fare clic sulla scheda Importa dati per importare i dati di rilevamento delle celle come file con estensione txt.
- In Set di dati - Inizializzazione, selezionare il numero di sezioni o fotogrammi da analizzare e impostare la calibrazione XY e l'intervallo di tempo tra i fotogrammi. Selezionare Applica impostazioni per salvare le impostazioni.
- Selezionare il simbolo Dati di stampa per formare i grafici delle traiettorie. Selezionare il simbolo Statistiche per quantificare le misure di distanza e velocità.
- Salvare i grafici di traiettoria come file bitmap (.bmp) e le misure di distanza e velocità come file .txt. Selezionare il simbolo Rimuovi dati. Ripetere l'operazione per gli altri file time-lapse.
NOTA: i grafici di traiettoria possono essere utilizzati per visualizzare la immediatezza dei singoli percorsi cellulari per una determinata condizione o stato di cella nel corso dei filmati time-lapse (Figura 4A). I dati relativi alla distanza e alla velocità memorizzati nei file .txt possono quindi essere utilizzati per ulteriori analisi.
-
Quantificazione dell'area della cresta neurale e della circolarità
- Aprire i file .stk time-lapse in ImageJ e calibrare in base alle impostazioni del microscopio, lavorando in pixel/m.
- In Analizza Imposta Misure, fare clic per selezionare i parametri della forma della cella: area della cella, perimetro e descrittore della forma.
- Usate lo strumento selezione a mano libera per disegnare manualmente intorno a ciascuna cella, utilizzando i contorni della membrana cellulare come guida.
- Premere i tasti Ctrl e B sulla tastiera per mantenere il contorno della cella sovrastato sull'immagine. Ripetere l'operazione per le celle su ogni intervallo di time-lapse.
- Utilizzare l'immagine Proprietà Overlay . A Gestione ROI per memorizzare i valori.
- Una volta delineate tutte le celle di interesse per fotogramma, fare clic su Misura. Salvare i risultati come file CSV.
NOTA: devono essere delineate da 10 a 20 celle per filmato, per un totale di 30-60 celle analizzate per condizione (n : 3). I dati delle forme di cella (file .csv) possono essere utilizzati per quantificare il modo in cui le dinamiche della forma delle cellule cambiano nel tempo (Figura 4C) o come la morfologia può essere modificata in diversi trattamenti cellulari.
Risultati
Utilizzando la procedura illustrata qui, gli embrioni di topo sono stati sezionati dall'utero e i tessuti extraembryonici sono stati rimossi (Figura 1A–D). Gli embrioni sono stati somiti messi in scena (utilizzando solo embrioni a 5-8 somiti (ss), Figura 1E,F). La piastra neurale cranica è stata poi sezionata e il neuroepitelio è stato isolato. Le cellule mesodermiche, identificate come cellule sciolte, circolari e mesenchice, sono state delicatamente spazzolate (Figura 1G-L). La piastra neurale anteriore può essere espiantata intera, nel qual caso il tessuto della cresta neurale emergerà lateralmente ed espanderà radialmente intorno all'espianto, oppure ogni bordo della piastra neurale (destra e sinistra) può essere espiantato separatamente. Ciò è particolarmente utile quando si espiantano da mutanti genetici.
Entro 24 h, una regione di cresta neurale cranica premigratoria (epiteliale) può essere chiaramente vista circondando l'espianto della piastra neurale (Figura 2B). Inoltre, una sottopopolazione di cellule della cresta neurale ha subito una transizione epiteliale a quella mesenchica e appare completamente mesenchymica (Figura 2). Così, abbiamo diversi anelli concentrici di celle distinte, con la piastra neurale (NP) al centro, la cresta neurale premigratory (pNC) nel cerchio intermedio, e una popolazione di cresta neurale migratoria (mNC) nell'anello esterno (Figura 2B). Al fine di tracciare le cellule NC, è possibile utilizzare modelli murini geneticamente modificati come si mostra Figura 2C. In questo caso, abbiamo usato la cresta neurale specifica Wnt1::Cre; RosamTmG che si traduce in celle NC essere etichettati in verde. In questi topi, le cellule esprimono il pomodoro a membrana (mT, in rosso) a meno che non esprimano Cre ricombinase. La ricombinazione porta alle cellule che esprimono la proteina fluorescente verde membrana (GFP, in verde). Le celle rosse mostrate al centro dell'espianto sono cellule di lamiere neurali. Alcune cellule di piastra neurale dorsale esprimono anche GFP; per la cultura a lungo termine, allestiamo tutte le cellule al centro. Per i nostri scopi, la purezza dell'espianto è sufficiente per tracciare le diverse popolazioni di cellule della cresta neurale. Dove è necessaria una maggiore purezza della cresta neurale, questa strategia di etichettatura genetica può essere combinata con lo smistamento fluorescente delle cellule attivate (FACS) per garantire la purezza della popolazione. In alternativa, è possibile fissare gli espianti e identificare la popolazione NC con l'etichettatura degli anticorpi.
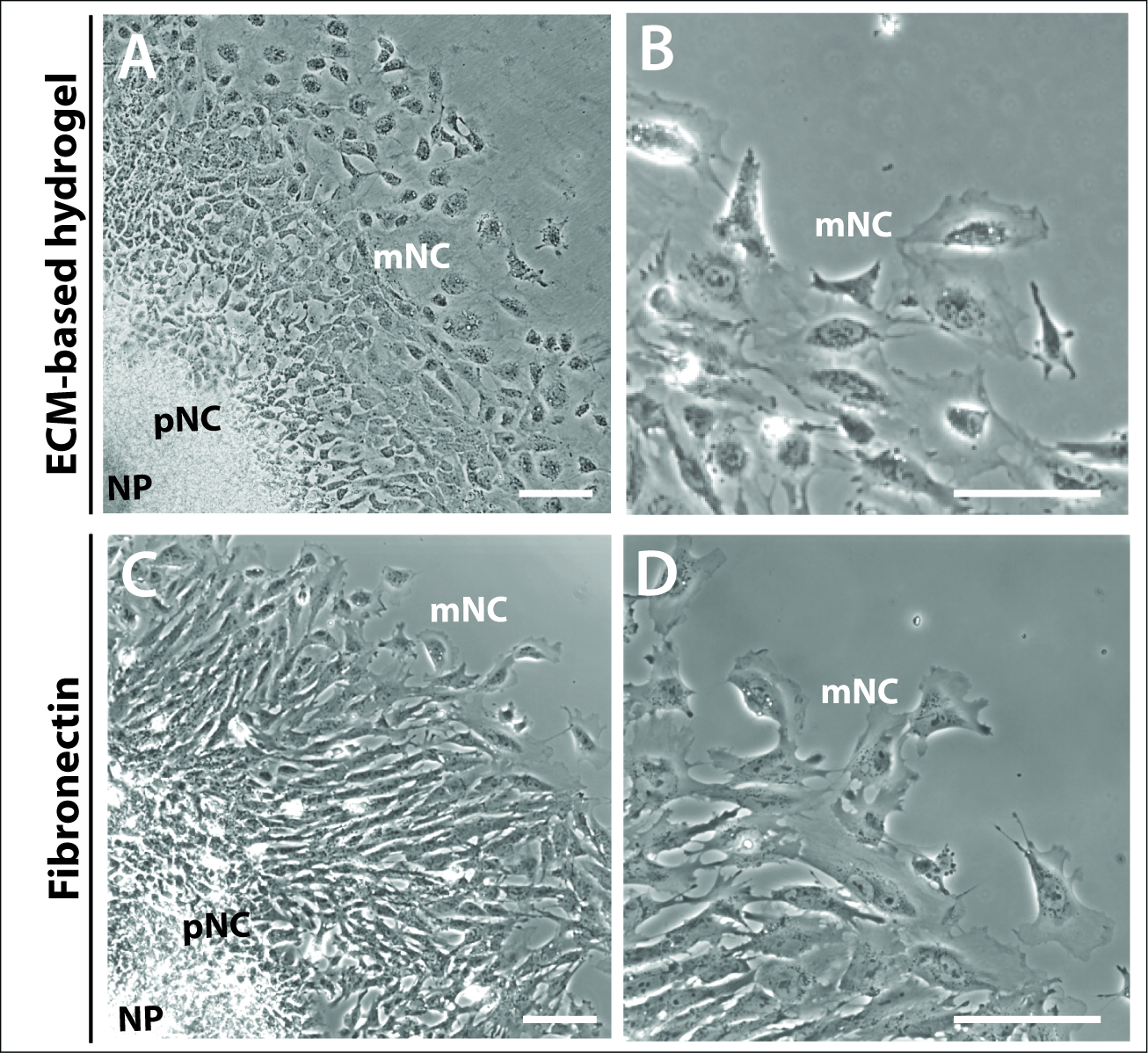
Era inoltre evidente da 24 h che i caratteristici anelli concentrici delle cellule NC premigratori e completamente migratori delle colture explant non dipendevano né erano governati dalla scelta della matrice (Figura 3). Le colture espiante placcate su un idrogel a base e cm e fibronectina formavano strutture espiante comparabili, che comprendevano le tre popolazioni cellulari, NP, pNC e mNC (Figura 3A,C). La morfologia delle cellule della cresta neurale era anche paragonabile a quelle placcate sull'idrogel e sulla fibrognactina a base ecm (Figura 3B,D). Tuttavia, gli expianta placcati sulla fibronectin a base di fibronectin prodotte cellule con lamellipopodi a più alta enza sul bordo della cellula, apparentemente più polarizzate nella direzione della migrazione (Figura 3B,D).
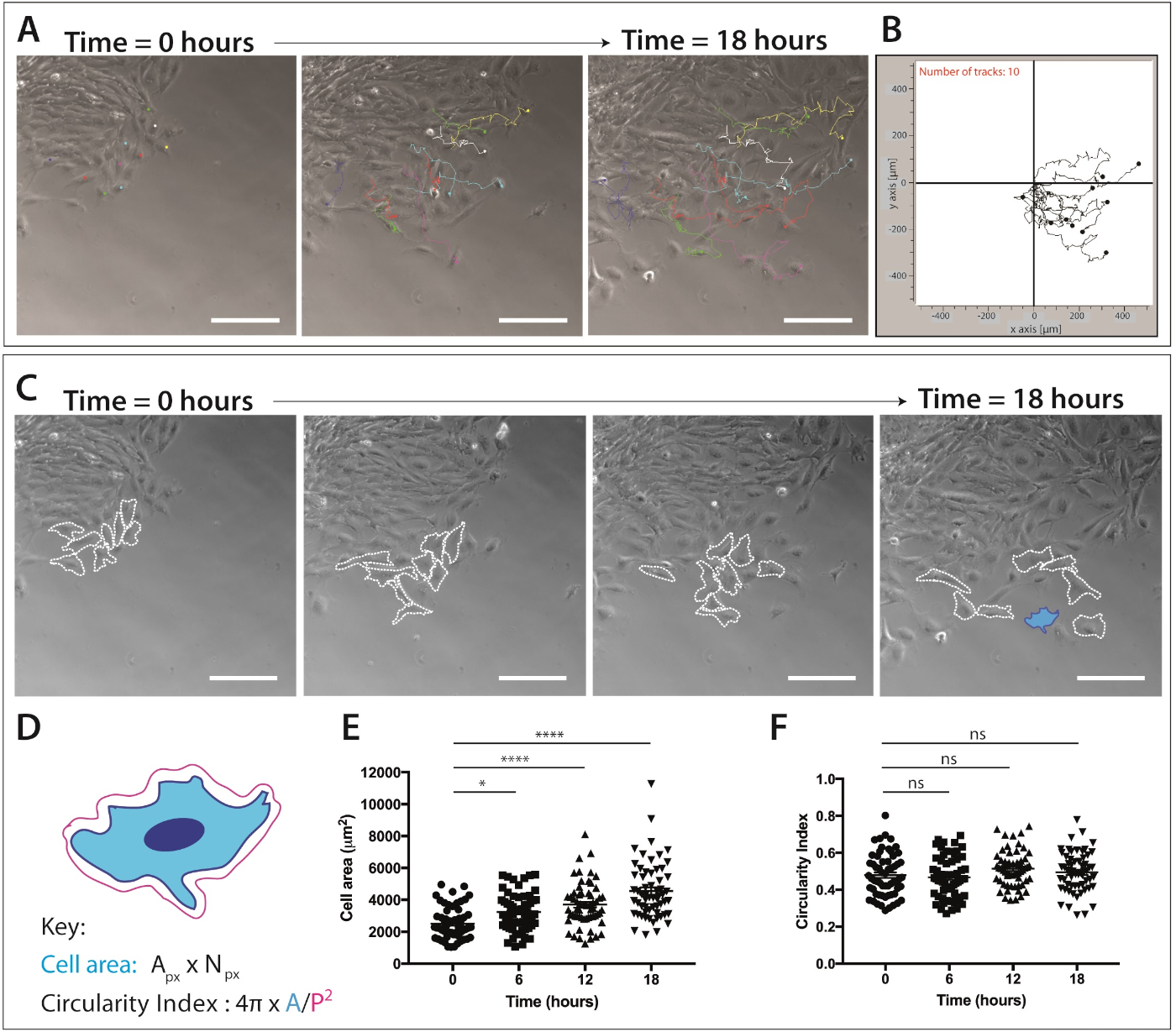
Una volta che una popolazione di cellule della cresta neurale migratoria è evidente, l'imaging delle cellule vive può essere completato. La microscopia time-lapse è impostata su un ingrandimento di 10 x (18 h, 1 fotogramma/5 min) per la successiva analisi della migrazione delle celle NC (Figura 4A). Il plug-in ImageJ Manual Tracking genera coordinate XY di singole celle su tutti i fotogrammi dei filmati time-lapse (Figura 4B). Queste coordinate possono essere elaborate utilizzando il software di migrazione. Questo software consente la visualizzazione di singole tracce cellulari nel tempo (Figura 4B) e può essere utilizzato per quantificare la distanza accumulata ed euclidea, così come la velocità delle cellule.
I dati di imaging time-lapse forniscono anche una grande quantità di informazioni sulla morfologia cellulare durante la migrazione delle cellule creste neurali craniche (Figura 4C). Delineando singole membrane cellulari, le misure dell'area cellulare e del perimetro possono essere calcolate da tutti i frame dei filmati (Figura 4C). Queste misurazioni consentono la successiva quantificazione dell'area cellulare e della circolarità (Figura 4D). La figura 4C mostra un'analisi delle variazioni della forma delle cellule su 18 h. Si noti che quando le cellule si allontanano dall'espianto, l'area cellulare aumenta in modo significativo mentre la circolarità delle cellule rimane relativamente costante (ANOVA unidirezionale, il test di confronto multiplo di Tukey) ( Figura 4E,F). Ciò suggerisce che quando le cellule si allontanano dal bordo epiteliale e perdono contatti tra cellule, mostrano un aumento dell'area di diffusione cellulare. Le misure di circolarità cellulare non sono cambiate in modo significativo nel tempo; tuttavia, i cambiamenti a breve termine nella circolarità possono essere osservati se viene quantificato un numero maggiore di punti temporali. Le misure di circolarità cellulare possono anche fornire dati interessanti sulle dinamiche della forma delle cellule in presenza di un segnale chemiotattico o in condizioni confinate.

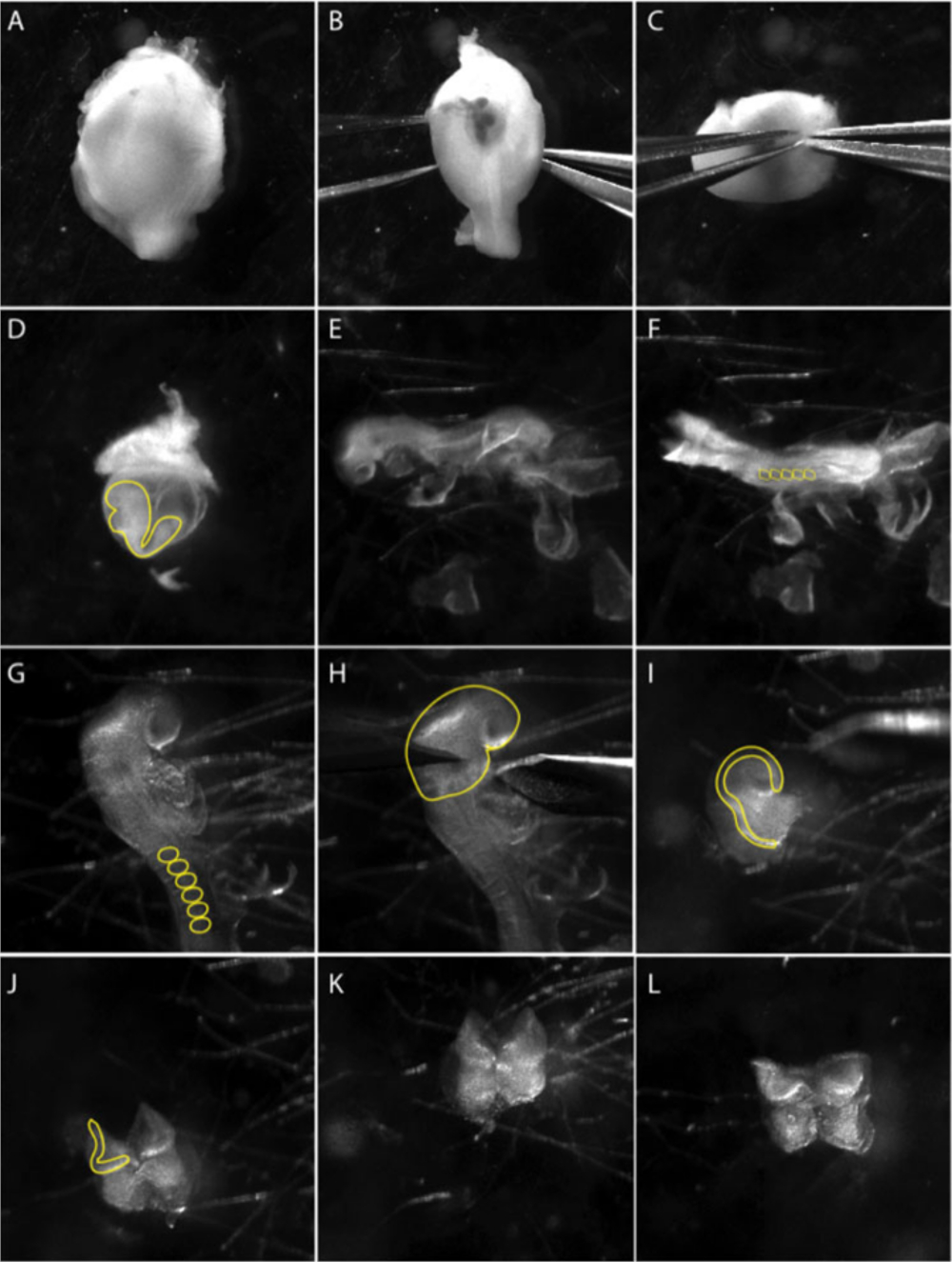
Figura 1: Isolamento degli espianti della cresta neurale cranica da un embrione e8.5.
Le immagini sono immagini di un video che documenta la tecnica della micro-dissezione. (A-C) Dissezione dell'embrione dall'utero. (B-C) Utilizzando due pinze affilate, allontanare delicatamente lo strato muscolare. Il pannello (D) mostra l'embrione all'interno del sacco del tuorlo viscerale (linea gialla). Estrarre l'embrione dal sacco del tuorlo viscerale. (E) Vedute laterali dell'embrione allo stadio 8,5 laterali. (F) Vista dorsale dell'embrione allo stadio 8.5. Contare i somiti (ss) per determinare l'età degli embrioni; di solito 5-8 ss (cerchi gialli in F). (G) Osservare da vicino la regione cranica dell'embrione. Rimuovere le membrane extraembryonici dalla regione cranica; somiti sono contrassegnati da una linea gialla. (H) Le dissezioni della piastra neurale anteriore vengono eseguite sotto il primo arco ramificato (linea gialla). (I) Vista laterale della dissezione della piastra neurale anteriore. Le pieghe neurali, dove sorgono le cellule neurali della cresta, sono contrassegnate da una linea gialla. (J-L) Rimuovere il tessuto mesodermico (cellule mesenchymal soffici) alla base delle pieghe neurali anteriori il più possibile prima di placcare NP su piatti di coltura preparati. Il filmato è stato ripreso utilizzando uno stereo-microscopio con una lente apocromatica widefield a zoom 3.0X (vedere la tabella dei materiali). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Espianto di cresta neurale cranica Murine.
(A) Rappresentazione schematica della vista dorsale di un embrione di topo e8.5. La regione cranica dell'embrione viene tagliata alla linea tratteggiata. Il bordo della piastra neurale (evidenziato in rosso) è isolato dal tessuto del mesoderma circostante e coltivato per 24 h per consentire alla cresta neurale cranica di emigrare. Schema adattato da22,23. (B) Sinistra: Rappresentazione immagine di campo luminoso di una cresta neurale cranica explant 24 ore dopo la placcatura. Si osservano tre popolazioni di cellule, che sono anche schematizzate a destra. NP - piastra neurale, pNC - cresta neurale pre-migrantia e mNC - cresta neurale migratoria. Barra della scala: 250 m. (C) Immagini di ingrandimento più elevato di un'espianto da topo geneticamente etichettato (Wnt1::cre; RosamTmG). Le celle senza driver Cre esprimono il pomodoro a membrana (mT) in rosso. Espressione di Cre sotto il controllo di una cresta neurale specifico Wnt1 promotore porta all'escissione della cassetta mT e l'espressione della membrana GFP (mG) in verde. I nuclei sono macchiati di Hoescht (in blu). Barra di scala : 200 m. (D-D') Immagini di ingrandimento superiore delle cellule migratorie che esprimono la Membrana GFP (D). (D') Il DNA è etichettato con Hoescht (blu). (D'') Unione di D e D'. Barra di scala 20 m. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Espiante coltivate su diversi substrati.
(A–B) Immagini di contrasto di fase di espianti coltivati su ECM-hydrogel commerciale. (C-D) Espiantate coltivate su fibronectina 1 g/mL. Le cellule della piastra neurale (NP), le cellule della cresta neurale premigratoria (pNC) e migratoria (mNC) possono essere distinte dalle loro diverse morfologie cellulari. Barra della scala: 100 m. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Quantificazione della migrazione delle cellule creste neurali craniche e delle dinamichedella forma cellulare.
(A) I fotogrammi a contrasto di fase dell'imaging time-lapse di colture ex-lapse sovrapposte a singole tracce di cellule di cresta neurale, utilizzando il plug-in ImageJ/Fiji Manual Cell Tracking. Dieci celle mNC rappresentative sono state tracciate manualmente su 18 h (217 fotogrammi) e xY coordinate sono state esportate. I dati sono rappresentati come un punto sovrapposto e grafici a linee. Le cellule sono state placcate su 1 fibronectina g/mL. Barra della scala: 200 m. (B) Grafico di traiettoria rappresentativo di 10 celle mNC, generato utilizzando un software di migrazione. (C) Fotogrammi a contrasto di fase tratti dall'analisi time-lapse delle colture espiante. Le linee tratteggiate delineano 8 cellule rappresentative della cresta neurale migratoria analizzate per la dinamica della forma delle cellule quando sono placcate su fibronectina da 1 g/mL. Barra della scala: 200 m. (D) Rappresentazione schematica dei calcoli utilizzati per quantificare l'area cellulare e la circolarità. La morfologia cellulare dello schema è quella della cellula evidenziata in blu (C). Unpx : area dei pixel, Npx , numero di pixel, A , area, P e perimetro (1 pixel , 1,60772 , m2). (E-F) La quantificazione delle misure di circolarità cellulare e di circolarità cellulare nel tempo. I dati rappresentano media : SEM. Ogni punto rappresenta una cella (n x 60), preso da 3 esperimenti indipendenti e analizzato a 0 h, 6 h, 12 h e 18 h (ns non significativo, s<0.05, s.p< 0.0001 unidirezionale ANOVA, test di confronto multiplo di Tukey). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
Studiare le cellule della cresta neurale dei mammiferi è stata una sfida per gli scienziati a causa della natura in utero dello sviluppo dei mammiferi. Gli studi in vivo sono difficili da stabilire, in quanto l'embrione deve essere manipolato in condizioni che imitano la vita nell'utero. In pratica, è quasi impossibile riprodurre questi embrioni (E8) per più di 24 ore, specialmente per l'imaging dal vivo. Inoltre, l'induzione e la migrazione della cresta neurale avvengono contemporaneamente alla chiusura del tubo neurale e alla svolta embrionale nel topo; questo è un evento morfogenetico cruciale e stressante, che spesso fallisce quando gli embrioni vengono coltivati ex utero. Pertanto, il tasso di successo degli approcci ex utero è generalmente basso. L'uso di cellule NC immortalate21 è uno strumento utile per ridurre l'uso animale e può fornire una migliore fonte di cellule di cresta neurale per l'analisi a lungo termine, la trasfezione e gli studi di arricchimento. Tuttavia, c'è chiaramente la necessità di coltura in modo affidabile cellule di cresta neurale primaria. Il nostro metodo è applicabile ai modelli genetici condizionati o knock-out del topo. Un metodo paragonabile al nostro è stato descritto per altre popolazioni di creste neurali20; tuttavia, il nostro metodo descrive a fondo l'isolamento passo-passo delle cellule NC craniche murine. Descriviamo anche in dettaglio l'uso di matrici diverse e la procedura di analisi della migrazione.
Per ottenere risultati coerenti, abbiamo scoperto che una particolare attenzione prestata alla messa in scena durante la selezione degli embrioni. Non sorprende che il numero di somiti sia correlato alle diverse fasi dello sviluppo cranico NC. Pertanto, la conoscenza dell'anatomia embrionale è molto importante prima di acquisire dati sperimentali. Questo approccio può quindi essere adattato per isolare popolazioni discrete di cellule della cresta neurale, a seconda della questione biologica e delle cellule bersaglio.
Una volta che gli embrioni sono selezionati ed sezionati, le cellule mesodermiche possono essere facilmente distinte e devono essere rimosse per consentire una migliore visualizzazione e ridurre la contaminazione. Per le colture a lungo termine, il tessuto della piastra neurale può essere rimosso a 24 h di placcatura al fine di prevenire la contaminazione da tessuti neurali. Un ulteriore perfezionamento potrebbe essere l'uso dell'etichettatura fluorescenza del lignaggio (ad esempio, utilizzando un caricatore Wnt1::cre o Sox10::creERT combinato con giornalisti fluorescenti24,25) per distinguere le cellule della cresta neurale da altri tessuti, come illustrato nella figura 2C.
Precedenti relazioni hanno evidenziato il potenziale di placcatura topo NC espiantare colture su diverse matrici, più comunemente su idrogel commerciali ECM, fibronectina e collagene I20,21,26. Nelle nostre mani, le colture di espiante NC cranial del tono vengono coltivate con successo su tutte e tre le matrici, a concentrazioni specificate nei rapporti originali (dati non mostrati). L'approccio raffinato iniziale che ci siamo adattati per le nostre colture espiante NC utilizzava un idrogel commerciale come matrice di scelta, che consisteva principalmente di laminina e collageni21(Figura 3A–B). Tuttavia, la composizione di questo idrogel non è chiaramente definita, con fattore di crescita sconosciuto e contenuto proteico. Come tale, da allora abbiamo spostato il nostro approccio alla placcatura dei topi NC espiantare le colture sulla fibronectina (Figura 3C-D). La fibronectina è ben definita e altamente espressa nelle membrane ECM e basement lungo le quali le cellule NC migranoin vivo28,29,30. Per ottimizzare una matrice di fibronectina che meglio replica la migrazione delle cellule della cresta neurale e la morfologia come visto utilizzando l'idrogel, abbiamo confrontato i comportamenti delle cellule NC esposti sull'idrogel con una titolazione di 0,25-30 fibrosictina , e definito 1 fibronectina g/mL come fornendo proprietà ideali (dati non mostrati). Crediamo che questo lavoro preliminare possa contribuire a stabilire un quadro per il confronto sistematico delle matrici, come la fibronectina, con quelle descritte in precedenza, vale a dire collagene e laminina32,33,34. Sarebbe particolarmente interessante confrontare la capacità migratoria delle cellule NC del topo sulla fibronectina contro collagene I, dato che il collagene-IA1 è endogenamente secreto da cellule NC di topo, aviarie e umane28,30,31,32. Il collagene I è quindi rilevante quanto la fibronectina nella considerazione della scelta della matrice. Vale anche la pena riconoscere che la biodisponibilità dei fattori di crescita nei media può essere alterata da diversi componenti di matrice, soprattutto dato l'elevato contenuto di siero dei nostri media. Per superare questo problema, stiamo attualmente lavorando per produrre condizioni di cultura definita senza siero. Questi supporti definiti sono utilizzati con successo nei protocolli di induzione della cresta neurale nel campo delle cellule staminali pluripotenti, ma richiedono un'ulteriore ottimizzazione per il nostro sistema di coltura degli espiantatori NC33,34. Il nostro lavoro può anche servire come punto di partenza per le condizioni di raffinazione per altri tipi di cellule NC come il dn cardiaco e del tronco, e per i successivi studi di differenziazione NC. Ancora più importante, questo protocollo consente l'isolamento delle cellule NC craniche per una varietà di applicazioni. Imvedremo studi sulla migrazione diretta, la migrazione 3D e l'invasione. Le cellule isolate in questo modo possono essere trattatein vitroper una serie di analisi. Ad esempio, le cellule possono essere facilmente trattate utilizzando diverse piccole molecole per colpire specifiche proteine, possono essere trattate in punti temporali definiti e gli esperimenti di washout possono essere progettati per determinare il recupero dei comportamenti cellulari (Figura 4). La coltura a più lungo termine per i saggi di trasfezione e differenziazione è possibile, così come il passaggio delle cellule (dati non mostrati). Tuttavia, la fattibilità, la capacità di rinnovamento cellulare e multipotenza deve essere convalidata dopo il passaggio. Le cellule placcate su coperture in vetro possono essere utilizzate anche in protocolli di colorazione immunofluorescente, seguendo l'imaging dal vivo. Infine, questo approccio rappresenta un sistema estremamente potente per studiare la migrazione di NC da modelli murini genetici22,23,24,25.
Divulgazioni
Gli autori non hanno conflitti di interesse.
Riconoscimenti
Siamo grati al King's College London Biological Services Unit, in particolare a Tiffany Jarvis e Lynsey Cashnella per il loro continuo sostegno. Ringraziamo Derek Stemple, Mamoru Ishii e Robert Maxson per consigli e aiuto con i reagenti durante la definizione iniziale di questo protocollo. Ringraziamo Dheraj Taheem per l'aiuto con l'irradiazione gamma delle cellule STO. Ringraziamo i laboratori Liu e Krause, in particolare Tommy Pallett, per un grande supporto. Questo lavoro è stato finanziato da sovvenzioni del BBSRC (BB/R015953/1 a KJL/MK), un'astronave del MRC Doctoral Training Programme (LD), Newland Pedley Fund (ALM) e KRIPIS II, Segretariato Generale della Ricerca e Tecnologia (GSRT), Del Ministero dell'Istruzione e delle Grecia e Fondation Santé (a SGGM).
Materiali
| Name | Company | Catalog Number | Comments |
| bFGF | R&D systems | 233-FB | |
| b-mercaptoethanol | Gibco | 31350-010 | |
| Tissue culture flasks | Corning | 430639 | 25cm2 culture flasks |
| DMEM (4500 mg/L glucose) | Sigma | D5671 | |
| Dulbecco's phosphate-buffered saline (dPBS) | Sigma | D8537 | |
| Ethanol | Fisher Chemicals | E/0650DF/C17 | |
| Fetal Bovine Serum | Sigma | TFS AA 10-155 | |
| Fetal Bovine Serum (ES Cell FBS) | Gibco | 26140079/16141-079 | |
| Fibronectin bovine plasma | Sigma | F1141 | |
| Gelatin from bovine skin | Sigma | G9382 | |
| L-glutamine | Sigma | G7513 | |
| Glass-bottomed, multi-well 24-well tissue culture plate | Ibidi | 82406 | Glass-bottomed, No 1.5, 24-well tissue culture dish with black sides |
| Cell migration analysis tool | Ibidi | v2.0 | Manuals for this software can be found at: https://ibidi.com/manual-image-analysis/171-chemotaxis-and-migration-tool.html. |
| Circularity Plug-in | ImageJ | v1.29 or later | The circularity plug-in is an extended version of ImageJ/Fiji’s Measure command, designed by Rasband, W., (2000) (wsr@nih.gov). |
| LIF | ESGRO by Millipore | ESG1106 | |
| Manual Cell Tracking Plug-in | ImageJ | v1.34k or later | ref Cordelieres F. Institut Curie, Orsay (France) 2005 |
| Microscope Image Analysis Software | Universal Imaging Corporation | 6.3r7 | MetaMorph Software Series 6.3r7 |
| MEM non-essential aminoacids 100X | Gibco | 11140-050 | |
| Matrigel (ECM-based Hydrogel) | Corning | 356234 | |
| Microscope | Olympus | 1X81 | Olympus 1X81 inverted microscope |
| Microscope camera | Photometrics | 512B | Photometrics Cascadell 512B camera (4x UPlanFL N, 10x UPlanFL N, 20x UPlanFL N, 40x UPlanFL N objectives) |
| Microscope controller | Olympus | 1X2-UCB | Olympus 1X2-UCB microscope controller |
| Microscope temperature controller | Solent Scientific | RS232 | Maintained at 37°C |
| Penicillin/streptomycin | Sigma | A5955 | |
| Sodium Pyruvate | Sigma | S8636 | |
| Statistics software | Graphpad Prism | v7.0 | |
| STO feeder cells | ATCC | CRL-1503 | |
| Stereomicroscope | Nikon | SMZ | |
| XY microscope stage controller | Applied Scientific Instrumentation (ASI) | MS-4400 |
Riferimenti
- Duband, J. L. Diversity in the molecular and cellular strategies of epithelium-to-mesenchyme transitions: Insights from the neural crest. Cell Adhesion & Migration. 4, 458-482 (2010).
- Mayor, R., Carmona-Fontaine, C. Keeping in touch with contact inhibition of locomotion. Trends in Cell Biology. 20, 319-328 (2010).
- Le Douarin, N., Kalcheim, C. The neural crest. 2nd edn. , Cambridge University Press. (1999).
- Trainor, P. A. Neural Crest Cells : Evolution, Development, and Disease. , Academic Press. (2014).
- Jiang, R., Bush, J. O., Lidral, A. C. Development of the upper lip: morphogenetic and molecular mechanisms. Developmental dynamics : an official publication of the American Association of Anatomists. 235, 1152-1166 (2006).
- Ahola, J. A., et al. Increased incidence of Hirschsprung's disease in patients with hypoplastic left heart syndrome--a common neural crest-derived etiology? Journal of Pediatric Surgery. 44, 1396-1400 (2009).
- Bajpai, R., et al. CHD7 cooperates with PBAF to control multipotent neural crest formation. Nature. 463, 958-962 (2010).
- Inoue, K., et al. Congenital hypomyelinating neuropathy, central dysmyelination, and Waardenburg-Hirschsprung disease: phenotypes linked by SOX10 mutation. Annals of Neurology. 52, 836-842 (2002).
- Kim, H., Kang, K., Ekram, M. B., Roh, T. Y., Kim, J. Aebp2 as an epigenetic regulator for neural crest cells. PloS one. 6, e25174(2011).
- Barriga, E. H., Trainor, P. A., Bronner, M., Mayor, R. Animal models for studying neural crest development: is the mouse different? Development. 142, 1555-1560 (2015).
- Bronner, M. E., LeDouarin, N. M. Development and evolution of the neural crest: an overview. Developmental Biology. 366, 2-9 (2012).
- De Calisto, J., Araya, C., Marchant, L., Riaz, C. F., Mayor, R. Essential role of non-canonical Wnt signalling in neural crest migration. Development. 132, 2587-2597 (2005).
- Carmona-Fontaine, C., et al. Contact inhibition of locomotion in vivo controls neural crest directional migration. Nature. 456, 957-961 (2008).
- Matthews, H. K., et al. Directional migration of neural crest cells in vivo is regulated by Syndecan-4/Rac1 and non-canonical Wnt signaling/RhoA. Development. 135, 1771-1780 (2008).
- Banerjee, S., et al. A novel role for MuSK and non-canonical Wnt signaling during segmental neural crest cell migration. Development. 138, 3287-3296 (2011).
- Pryor, S. E., et al. Vangl-dependent planar cell polarity signalling is not required for neural crest migration in mammals. Development. 141, 3153-3158 (2014).
- Serbedzija, G. N., Bronner-Fraser, M., Fraser, S. E. Vital dye analysis of cranial neural crest cell migration in the mouse embryo. Development. 116, 297-307 (1992).
- Kawakami, M., Umeda, M., Nakagata, N., Takeo, T., Yamamura, K. Novel migrating mouse neural crest cell assay system utilizing P0-Cre/EGFP fluorescent time-lapse imaging. BMC Developmental Biology. 11, 68(2011).
- Stemple, D. L., Anderson, D. J. Isolation of a stem cell for neurons and glia from the mammalian neural crest. Cell. 71, 973-985 (1992).
- Etchevers, H. Primary culture of chick, mouse or human neural crest cells. Nature protocols. 6, 1568-1577 (2011).
- Ishii, M., et al. A stable cranial neural crest cell line from mouse. Stem Cells and Development. 21, 3069-3080 (2012).
- Gonzalez Malagon, S. G., Liu, K. J. ALK and GSK3: Shared Features of Neuroblastoma and Neural Crest Cells. Journal of Experimental Neuroscience. 12, 1179069518792499(2018).
- Gonzalez Malagon, S. G., et al. Glycogen synthase kinase 3 controls migration of the neural crest lineage in mouse and Xenopus. Nature Communications. 9, 1126(2018).
- Danielian, P. S., Muccino, D., Rowitch, D. H., Michael, S. K., McMahon, A. P. Modification of gene activity in mouse embryos in utero by a tamoxifen-inducible form of Cre recombinase. Current Biology : CB. 8, 1323-1326 (1998).
- Laranjeira, C., et al. Glial cells in the mouse enteric nervous system can undergo neurogenesis in response to injury. Journal of Clinical Investigation. 121, 3412-3424 (2011).
- Pfaltzgraff, E. R., Mundell, N. A., Labosky, P. A. Isolation and culture of neural crest cells from embryonic murine neural tube. Journal Of Visualized Experiments : JoVE. , e4134(2012).
- Sternberg, J., Kimber, S. J. The relationship between emerging neural crest cells and basement membranes in the trunk of the mouse embryo: a TEM and immunocytochemical study. Journal of Embryology and Experimental Morphology. 98, 251-268 (1986).
- Sternberg, J., Kimber, S. J. Distribution of fibronectin, laminin and entactin in the environment of migrating neural crest cells in early mouse embryos. Journal of Embryology and Experimental Morphology. 91, 267-282 (1986).
- George, E. L., Georges-Labouesse, E. N., Patel-King, R. S., Rayburn, H., Hynes, R. O. Defects in mesoderm, neural tube and vascular development in mouse embryos lacking fibronectin. Development. 119, 1079-1091 (1993).
- Henderson, D. J., Copp, A. J. Role of the extracellular matrix in neural crest cell migration. Journal of Anatomy. 191 (Pt 4), 507-515 (1997).
- Thomas, S., et al. Human neural crest cells display molecular and phenotypic hallmarks of stem cells. Human Molecular Genetics. 17 (21), 3411-3425 (2008).
- Greenberg, J. H., Seppa, S., Seppa, H., Hewitt, T. Role of collagen and fibronectin in neural crest cell adhesion and migration. Developmental Biology. 87 (2), 259-266 (1981).
- Leung, A. W., et al. WNT/β-catenin signaling mediates human neural crest induction via a pre-neural border intermediate. Development. 143 (3), 398-410 (2016).
- Zhu, Q., Lu, Q., Gao, R., Cao, T. Prospect of Human Pluripotent Stem Cell-Derived Neural Crest Stem Cells in Clinical Application. Stem Cells International. 2016, 7695836(2016).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati