
Method Article
Isolamento delle cellule mononucleate del sangue periferico umano e delle cellule T CD4+ da pazienti con sindrome di Sézary per la profilazione trascrittomica
In questo articolo
Riepilogo
Presentiamo un semplice protocollo per l'isolamento di cellule mononucleate del sangue periferico da sangue intero ottenuto da pazienti con diagnosi di Sindrome di Sézary, seguito dalla selezione di cellule T CD4+, dalla loro stimolazione con phorbol12-myristate13-acetato e A23187 ionoforo, e dalla preparazione dell'RNA per la profilazione trascrittomica.
Abstract
I linfomi cutanei a cellule T (CTCL) derivano dalla trasformazione e dalla proliferazione incontrollata di cellule T mature che presentano la pelle, e la micosi fungoide (MF) e la sindrome di Sézary (SS) rappresentano i sottotipi più comuni. Nonostante una serie di studi sulla caratterizzazione dell'espressione genica, delle alterazioni genetiche e delle anomalie epigenetiche della CTCL, la patogenesi molecolare di MF/SS rimane poco chiara. MF si riferisce al CTCL più comune con una predominanza cutanea, ed è solitamente limitato alla pelle, mentre SS è una variante leucemica aggressiva di CTCL con diffuso coinvolgimento cutaneo ed è caratterizzata da una distribuzione neoplastica che coinvolge principalmente sangue, pelle e linfonodi. Concentrandosi sulla pratica clinica, l'identificazione dei biomarcatori di espressione genica ha un enorme potenziale per migliorare la diagnosi e il trattamento di MF / SS. In effetti, recenti studi trascrittomici hanno identificato potenziali biomarcatori diagnostici dalle differenze nell'espressione genica tra cellule T normali e maligne, che possono migliorare la nostra comprensione della biologia SS e rivelare potenziali bersagli terapeutici. Questo manoscritto descrive un protocollo riproducibile dettagliato per l'isolamento delle cellule mononucleate del sangue periferico dal sangue intero fresco da pazienti con diagnosi di SS, selezione di cellule T di memoria CD4 + (cellule T CD4 + CD45RO +), stimolazione chimica e preparazione di RNA adatto per la profilazione trascrittomica per scoprire nuovi marcatori molecolari prognostici per ottenere ulteriori informazioni sull'eziologia della malattia. La stimolazione con agonista chimico per attivare la regolazione nucleare fornisce una valutazione più specifica per i percorsi importanti nella regolazione dinamica della trascrizione e nell'espressione genica ed elimina i difetti confondenti che possono derivare da difetti di segnalazione a monte derivanti dalla perdita di antigene TCR sulla membrana cellulare. I dati ottenuti dal confronto del trascrittoma delle cellule T SS non stimolate e stimolate smascherano i difetti funzionali regolatori dell'espressione genica non evidenti dall'analisi delle cellule quiescenti non stimolate. Inoltre, il metodo delineato da questo approccio può essere adattato per studiare i difetti di espressione genica delle cellule T in altre malattie immunitarie delle cellule T.
Introduzione
Il linfoma cutaneo a cellule T (CTCL), compresi i sottotipi più comuni micosi fungoide (MF) e sindrome di Sézary (SS), è un gruppo eterogeneo di malattie derivate dalla trasformazione e dalla proliferazione incontrollata di cellule T mature 1,2. Le cellule T neoplastiche hanno un CD4+CD45RO+ maturo, fenotipodi memoria 3, ed esprimono marcatori di adesione all'homing cutaneo, aumentando l'epidermotropismo4, che si manifesta come un'eruzione cutanea in particolare nella malattia precoce. Il decorso clinico della MF è spesso indolente quando si è sotto cure gestite di routine, ma un sottogruppo di pazienti può progredire verso una malattia più avanzata. In questi casi di MF, le lesioni cutanee crescono e si ispessiscono in tumori di grandi dimensioni e le cellule T neoplastiche possono diffondersi ai linfonodi e agli organi viscerali. Al contrario, la SS è una variante leucemica più aggressiva della CTCL5, caratterizzata da una triade di sintomi: eritroderma generalizzato (definito come che colpisce >80% della superficie corporea totale), linfoadenopatia e presenza di più di 1000/mm3 cellule T clonali atipiche circolanti con nuclei cerebriformi, le cosiddette cellule di Sézary 6,7 . La prognosi per i pazienti con SS è significativamente peggiore di MF. La SS è rara con un tasso di incidenza di 0,1/100.000 e rappresenta circa il 3% dei casi totali di CTCL 8,9. La CTCL si presenta tipicamente negli anziani con un'età media di circa 60 anni10. Il tasso di incidenza per CTCL era in aumento e mentre la causa non è chiara, il tasso si è stabilizzato dal 199811,12.
La patogenesi molecolare della SS rimane poco chiara. Gli studi genetici, epigenetici e di espressione genica hanno prodotto una ricchezza di nuovi dati, tuttavia rimangono risultati incoerenti, principalmente a causa delle piccole coorti di pazienti studiate2, nonché delle differenze nella progettazione sperimentale e nelle popolazioni di controllo13,14. Una migliore caratterizzazione genomica e trascrittomica può far luce sia sui meccanismi della malattia che su obiettivi terapeutici precedentemente inesplorati. Pertanto, sono necessari ulteriori studi da una popolazione di pazienti più ampia per comprendere meglio questa neoplasia eterogenea. I pannelli di biomarcatori altamente sensibili e specifici in una coorte SS hanno funzionato in modo meno uniforme in altre coorti15, il che rappresenta un serio ostacolo nello sviluppo di biomarcatori diagnostici e prognostici affidabili per SS16. I biomarcatori diagnostici ideali saranno costantemente e altamente sovraespressi nelle cellule T maligne, mentre assenti o quasi assenti nelle cellule T normali17. La scoperta di biomarcatori specifici della malattia è importante per il progresso dei protocolli diagnostici e terapeutici per le SS.
Dati trascrittomici di alta qualità sia per le cellule T maligne che per quelle normali richiedono un approccio efficiente e affidabile alla preparazione del campione. Qui, discuteremo una strategia dettagliata ma semplice per ottenere campioni di RNA da popolazioni di cellule T rilevanti per SS. Discuteremo l'isolamento delle cellule mononucleate del sangue periferico (PBMC) dal sangue intero, la selezione magnetica negativa delle popolazioni di cellule T CD4 + CD45RO + rilevanti per la malattia, l'attivazione chimica per rivelare differenze nelle risposte funzionali e la preparazione dell'RNA per la profilazione trascrittomica. Nel protocollo attuale, l'attivazione chimica è stata eseguita utilizzando phorbol myristate acetate (PMA) e calcio ionoforo (A23187)18,19, perché studi precedenti hanno mostrato una segnalazione difettosa del recettore delle cellule T in CTCL e la stimolazione con PMA / A23187 bypassa il recettore delle cellule T20,21. Inoltre, PMA/A23187 consente un'attivazione prossimale più diretta dei segnali nucleari necessari per l'attivazione del gene delle citochine. Infine, la stimolazione delle cellule T fornisce un ulteriore livello di comprensione della regolazione dell'espressione genica che non potrebbe essere ottenuto dalle cellule T a riposo in cui il cambiamento dinamico è assente.
Protocollo
Le cellule umane sono potenzialmente infettive. Pertanto, gli esperimenti vengono eseguiti rigorosamente in conformità con le precauzioni e le procedure richieste discusse come amministrazione per la sicurezza e la salute sul lavoro (OSHA) e dispositivi di protezione individuale (DPI).
1. Isolamento dei PBMC dal sangue intero
- Raccogliere tutti i materiali necessari dalla Tabella 1 e portarli a temperatura ambiente (RT). Riscaldare RP10F a 37 °C. Regolare la centrifuga su RT. Fatta eccezione per le centrifughe e il conteggio delle cellule, eseguire tutti i passaggi utilizzando cellule vitali in un armadio di sicurezza biologica.
- Ottenere il sangue in cinque tubi da 10 ml (quantità desiderata) contenenti anticoagulante. Conservare il sangue intero a temperatura ambiente (18\u201224 °C). Etichettare 50 mL di provette di separazione con il numero del soggetto di ricerca umano per il campione di sangue da elaborare.
- Trasferire 10\u201215 mL di sangue in ciascun tubo di separazione con il numero del soggetto corrispondente. Diluire il sangue almeno 2 volte con la soluzione salina bilanciata di Hank (HBSS). Non superare i 35 ml di sangue diluito per tubo.
- Con attenzione e lentamente sottofondare il sangue con ~ 13 ml di mezzo di densità. Guarda attraverso il mezzo di densità trasparente nella parte inferiore del tubo e interrompi il pipettaggio quando la pipetta è quasi vuota (per evitare il rilascio di bolle). Rimuovere con attenzione la pipetta per evitare di mescolare il sangue e gli strati medi di densità.
- Trasferire con cura i tubi di separazione riempiti alla centrifuga senza disturbare gli strati.
- Centrifuga a 500 x g per 30 min con il freno della centrifuga spento (decelerazione impostata a zero).
NOTA: se la centrifuga visualizza solo rpm, consultare le specifiche del rotore per stimare l'equivalente rpm per 500 x g. - Rimuovere con cura i tubi di separazione dalla centrifuga senza disturbare gli strati. Osservare il buffy coat, che si è formato tra il mezzo di densità e gli strati di plasma.
- Pipetta dall'alto per rimuovere e scartare la maggior parte della frazione plasmatica superiore, in modo che 10 ml rimangano sopra il rivestimento buffy. Raccogli con cura e lentamente il cappotto buffy. Trasferire i buffy coats da due tubi di separazione in un nuovo tubo da 50 mL pre-marcato e sterile, come mostrato nella Figura 1.
- Diluire i PBMC di almeno 2 volte con HBSS, portando il volume in ogni nuovo tubo fino a 50 ml. Ricordarsi di cambiare il freno della centrifuga al massimo. PBMC a pellet mediante centrifugazione a 400 x g per 10 min. Rimuovere il surnatante il più possibile e toccare il fondo del tubo per allentare il pellet.
- Per lisare i globuli rossi residui (RBC), risospesciare ogni pellet cellulare in 1\u20122 mL di tampone di lisi ammonio-cloruro-potassio (ACK) per 10 mL di volume di sangue originale. Incubare esattamente per 5 minuti. Arrestare tempestivamente la lisi con un volume uguale o superiore di HBSS e regolare il volume a 50 ml. Centrifuga a 400 x g per 10 min.
- Rimuovere il surnatante e picchiettare il fondo del tubo per allentare il pellet cellulare. Raggruppare le cellule dello stesso donatore. Portare il volume fino a 50 ml con HBSS. Centrifuga a 400 x g per 10 min.
- Rimuovere il surnatante e picchiettare il fondo del tubo per allentare il pellet cellulare. Risospese le cellule in 10 ml di mezzo caldo RP10F e prendere un'aliquota per il conteggio delle cellule vitali usando il tripano blu.
- Calcola il numero totale di cellule in ogni campione usando un emocitometro.
2. Purificazione di cellule T CD4+CD45RO+ da PBMC
NOTA: La purificazione delle cellule T CD4+CD45RO+ dai PBMC viene effettuata utilizzando la separazione magnetica disponibile in commercio (vedere Tabella dei materiali) con modifiche minori. Si preferisce seguire il manuale del kit per il tempo di incubazione in quanto ogni kit commerciale ha le proprie istruzioni.
- Lavare la quantità desiderata di PBMC in 10 ml di buffer di selezione. Centrifuga a 400 x g per 10 min. Rimuovere il surnatante e picchiettare il fondo del tubo per allentare il pellet.
- Diluire i PBMC a 5 x 107 celle/mL nel tampone di selezione e trasferirli in un tubo a fondo tondo da 5 mL di polistirene (12 x 75 mm).
- Aggiungere 50 μL di cocktail di anticorpi per 1 mL di campione e mescolare delicatamente. Incubare a temperatura ambiente per 5 min.
- Immediatamente prima dell'uso, vortici di particelle magnetiche per 30 secondi ad alta velocità. Aggiungere 50 μL di particelle magnetiche per 1 mL di campione al tubo contenente PBMC e mescolare delicatamente.
- Portare il volume fino a 2,5 ml con tampone di selezione e mescolare delicatamente. Posizionare il tubo (senza coperchio) nel magnete e incubare a RT per 2,5 minuti.
- Prelevare il magnete e, con un movimento continuo, invertire il magnete e il tubo per versare la sospensione cellulare arricchita in un nuovo tubo sterile.
- Per aumentare il recupero, aggiungere 2,5 mL di tampone di selezione al tubo rimanente nel magnete, senza disturbare le perline immobilizzate. Tenere il magnete per altri 2,5 minuti e ripetere il passaggio 2.6 per recuperare celle aggiuntive.
- Prendi un'aliquota per il conteggio delle cellule vitali usando il tripano blu. Calcola il numero totale di cellule usando un emocitometro o un contatore di cellule.
- Confermare la purezza mediante citometria a flusso (Figura 3).
3. Attivazione chimica
- Regolare le cellule T CD4 + CD45RO + a 5 x 106 celle / mL con mezzo CALDO RP10F e distribuire le cellule in piatti di coltura della dimensione desiderata. Riposa le celle in un incubatore umidificato a 37 °C 5% CO2 durante la notte.
- Celle a riposo a pellet mediante centrifugazione a 400 x g per 10 min. Rimuovere il surnatante e picchiettare il fondo del tubo per allentare il pellet cellulare.
- Regolare la concentrazione cellulare a 5 x 106 celle/mL con mezzo caldo RP10F e distribuire 0,5\u20121 x 107 celle in ciascuno dei tre tubi sterili con tappo a vite.
NOTA: Se sono disponibili cellule sufficienti, nel progetto sperimentale possono essere incluse stimolazioni duplicate o punti temporali aggiuntivi. - Stimolare le cellule nei tubi 2 e 3 con PMA e A23187. Aggiungere PMA a 25 ng/mL e A23187 a 500 ng/mL e mescolare delicatamente. Aggiungere un volume uguale di dimetilsolfossido (DMSO) alle cellule nel tubo 1 che servirà come veicolo (controllo). Ad esempio, se si utilizzano 1 μL di PMA e 1 μL di A23187, aggiungere 2 μL di DMSO al tubo del veicolo. Mantenere la concentrazione finale di DMSO al di sotto dello 0,5% in tutti i tubi.
- Allentare i tappi sui tubi e riportare le celle all'incubatore a CO2 a 37 °C 5% per 2 ore (tubo 2) e 6 ore (tubi 1 e 3). All'ora indicata, centrifugare le celle a 500 x g per 10 min.
- Prima della lisi, scartare il più possibile il surnatante, senza disturbare il pellet cellulare.
- Lisare tempestivamente le cellule, come indicato dalle istruzioni del kit di isolamento dell'RNA disponibile in commercio (vedere Tabella dei materiali). Procedere con l'isolamento dell'RNA o congelare il lisato a -80 °C per elaborarlo successivamente con campioni aggiuntivi.
NOTA: la purezza e l'integrità dell'RNA possono essere verificate utilizzando l'elettroforesi microcapillare. - Opzionale: per rimuovere completamente tutte le tracce di DNA dal campione di RNA purificato, utilizzare il kit di pulizia dell'RNA (vedere Tabella dei materiali), secondo le istruzioni del produttore.
Risultati
Questo protocollo include procedure per l'isolamento delle PBMC dal sangue SS, la purificazione delle cellule T CD4 + CD45RO + mediante selezione negativa, la stimolazione delle cellule T purificate e l'isolamento dell'RNA totale per la profilazione trascrittomica. La Figura 1 descrive il processo di isolamento della PBMC dal sangue intero. Si prega di notare che la resa totale delle SS PBMC varierà con il volume iniziale del sangue e il carico tumorale circolante di ciascun paziente. Nel nostro laboratorio, la resa media delle SS PBMC è stata di 4,6 × 106 cellule/mL di sangue intero (1,85 × 106 – 3,25 x 107 cellule/mL per 7 SS). La redditività media dei PBMC isolati è stata del 95\u201299%. La Figura 2 mostra l'elevata purezza e vitalità delle cellule T di memoria CD4+CD45RO+ selezionate. La resa media di cellule T CD4+CD45RO+ da PBMC SS è stata del 75% (75,6% – 84%), rispetto al 15,9% (3% – 30%) di PBMC donatori normali (ND) ottenuti da camere del sistema di leucoriduzione (LRS). La vitalità e la purezza delle cellule T CD4+CD45RO+ ottenute da questo protocollo di selezione negativa sono state costantemente elevate (Figura 3).
In precedenza abbiamo combinato il protocollo di attivazione di cui sopra con microarray per studiare i cambiamenti funzionali nei trascrittomi di entrambe le cellule T SS e ND e abbiamo dimostrato che le cellule T di memoria SS e le PBMC SS esprimono scarsamente citochine e altri geni di risposta immunitaria rispetto alle cellule T ND e PBMC 19,22,23. La Figura 4 mostra la robusta attivazione di diversi geni delle citochine tra cui IL4, IL 10, IL13 e IL22 nelle cellule T ND, ma non nelle cellule T SS. Questo difetto nell'espressione genica funzionale nelle cellule T SS è stato confermato da altri gruppi24. Inoltre, molti geni non normalmente espressi nelle cellule T ND sono altamente espressi nelle cellule T SS, sia a riposo che dopo stimolazione (Figura 4). Questi includono i geni biomarcatori SS precedentemente descritti DNM3, PLS3, TOX e TWIST1 25,26,27, così come ANK1 e SGCE, che sono stati segnalati per la prima volta dal nostro gruppo. Questi biomarcatori positivi sono altamente espressi in SS, ma non in ND, ed evitano insidie tecniche associate a biomarcatori negativi.

Figura 1: Isolamento PBMC dal sangue intero. Fare clic qui per visualizzare una versione più grande di questa figura.
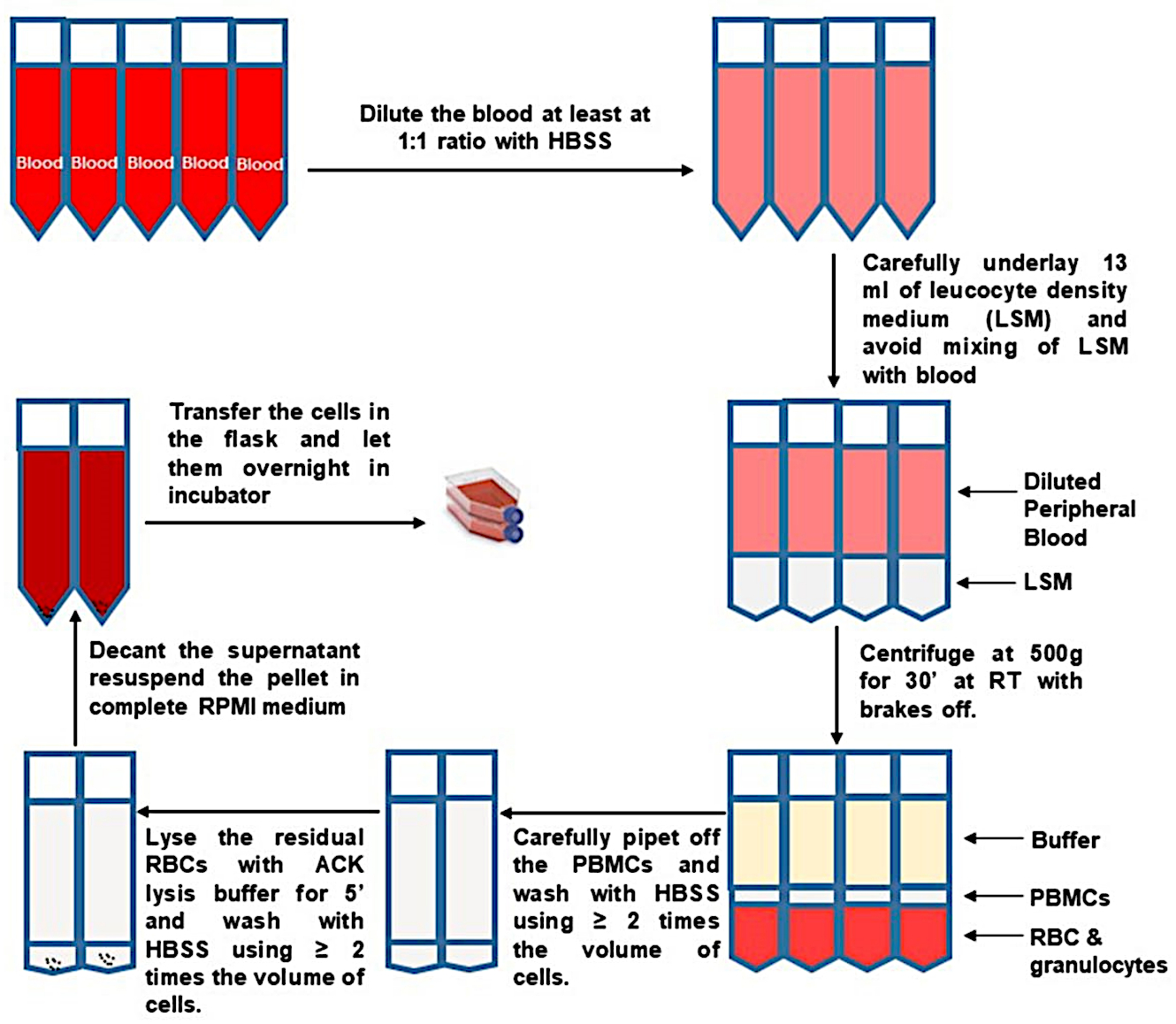{kind=link}

Figura 2: Selezione negativa di cellule T di memoria CD4+ da PBMC isolati. Fare clic qui per visualizzare una versione più grande di questa figura.
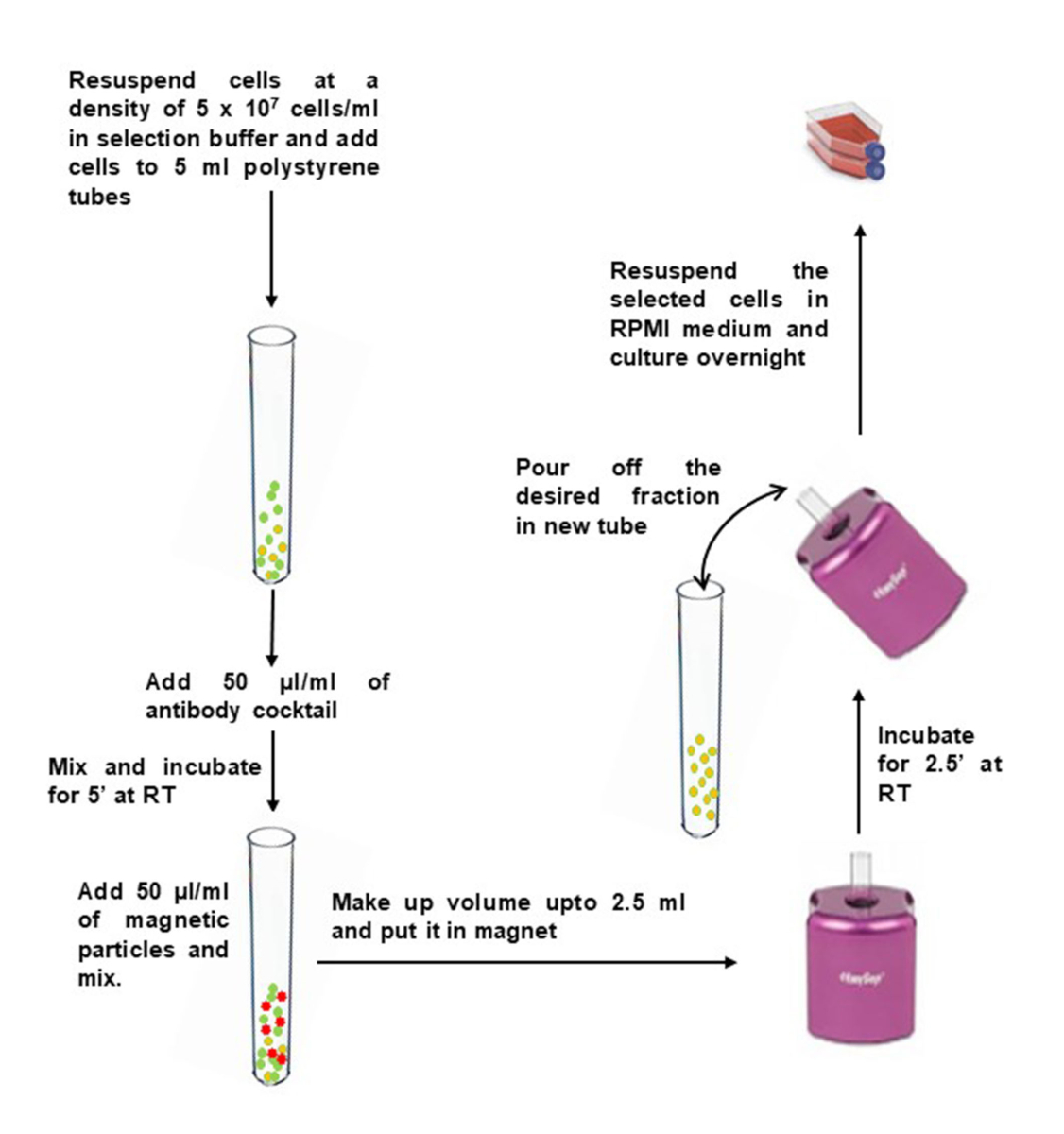{kind=link}

Figura 3: La purezza delle cellule T CD4+CD45RO+ è stata confermata dalla citometria a flusso. I linfociti sono stati controllati da diffusione della luce (A), i linfociti vivi hanno escluso il colorante di vitalità eFluor780 (B) e (C) rappresenta donatori normali non selezionati (ND). La selezione negativa ha portato a popolazioni quasi pure di cellule T CD45RO+ in pazienti ND (D) e SS (E). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Espressione genica differenziale in cellule T di memoria CD4+CD45RO+ a riposo e attivate da SS e ND. L'espressione genica z-score è rappresentata da una scala di colori dal rosso (alta espressione) al verde (bassa espressione). Le barre colorate nella parte superiore della mappa di calore rappresentano i trattamenti cellulari: finto / veicolo trattato (rosso), 2 ore stimolate (blu) e 6 ore stimolate (giallo). Diversi geni biomarcatori SS sono altamente espressi e i geni delle citochine sono scarsamente espressi nelle cellule T SS rispetto alle cellule T ND. Fare clic qui per visualizzare una versione più grande di questa figura.
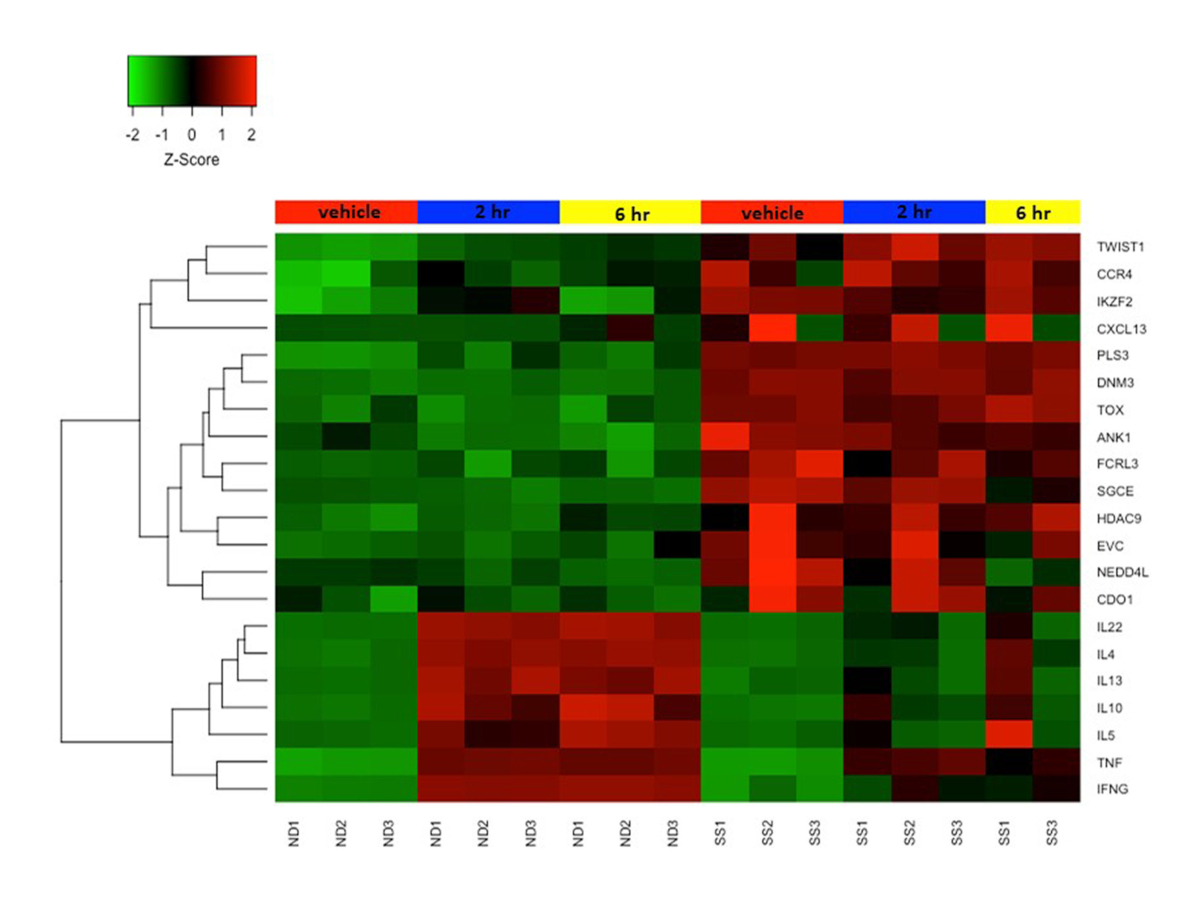{kind=link}
| Reagenti | |
| Densità Media | Mezzo di separazione dei linfociti, Ficoll-Hypaque o mezzo a densità equivalente con densità = 1.077-1.080g/ml a 20oC. |
| HBSS · | 1x soluzione salina bilanciata di Hank, 4,2 mM NaHCO3, 10 mM HEPES, pH 7,2 |
| RP10F · | RPMI 1640 medio, siero bovino fetale inattivato al 10 % di calore (FBS), 1x soluzione di penicillina-streptomicina, pH 7,2 |
| Tampone di lisi ACK | 155 mM NH4Cl, 10 mM KHCO3, 0,1 mM Na2EDTA, Non è necessario regolare il pH. Dovrebbe essere ~ 7.3. |
| Buffer di selezione | 1x HBSS, 2% FBS, 2 mM EDTA |
| forbol12-miristato13-acetato (PMA) | 50 μg/ml in DMSO |
| A23187 ionoforo | 500 μg/ml in DMSO |
Tabella 1: Reagenti.
Discussione
Sono stati sviluppati diversi modi per isolare i PBMC e ognuno ha i propri vantaggi e limiti28. Raccogliamo abitualmente fino a 50 ml di sangue in cinque tubi da 10 ml contenenti anticoagulanti. Il volume del sangue per l'isolamento PBMC dipende da diversi fattori come la salute e l'età del soggetto di ricerca e anche dall'esperienza flebotomista. Un passaggio procedurale critico nel protocollo è la formazione del gradiente di passo. Una scarsa stratificazione può causare il fallimento parziale o completo dei PBMC nel sedimentare l'interfaccia. Preferiamo il metodo di sottostrato descritto qui, in quanto è facile avviare lo strato inferiore. Per erogare completamente tutto il mezzo di densità sotto il sangue, è fondamentale utilizzare un ausilio per pipetta senza perdite d'aria. La contaminazione della frazione PBMC da parte di tipi di cellule indesiderate può essere ridotta al minimo mediante una raccolta attenta e coerente del buffy coat, che dovrebbe essere eseguita allo stesso modo per ogni isolamento. Se i PBMC non saranno ulteriormente frazionati, si dovrebbe evitare di raccogliere quantità diverse del gradiente di densità e degli strati plasmatici tra gli isolamenti. La lisi dei globuli rossi viene eseguita per ridurre al minimo il potenziale impatto della contaminazione dell'RNA derivato da RBC e reticolociti sulle analisi di espressione genica a valle. La lisi ipotonica sarà inibita dall'eccesso di tampone isotonico.
Un ulteriore isolamento del sottoinsieme di cellule T è importante per gli studi molecolari. Qui abbiamo descritto la successiva selezione delle cellule T CD4 + CD45RO + mediante selezione negativa per rimuovere i tipi di cellule indesiderate. La selezione negativa si basa sugli anticorpi che riconoscono specifici marcatori della superficie cellulare per tutte le cellule indesiderate. Le cellule rivestite di anticorpi vengono quindi rimosse da perline magnetiche. Questo protocollo di selezione rimuove le cellule indesiderate consentendo alle cellule bersaglio intatte e non stimolate di rimanere libere di fluttuare, il che è essenziale nello studio dell'attivazione genica. Tuttavia, è necessario prestare attenzione per evitare grumi cellulari, che riducono la purezza finale delle cellule T CD4 + CD45RO + selezionate. L'acido etilendiamminotetraacetico (EDTA) presente nel tampone di selezione riduce al minimo l'aggregazione cellulare. La resa delle cellule T dipende da fattori quali il volume iniziale del sangue, le variabili del paziente come il trattamento somministrato al paziente e lo stadio della malattia al momento della raccolta del campione. Il trattamento somministrato ai pazienti può anche influenzare la vitalità cellulare. Inoltre, la raccolta di campioni prima di qualsiasi procedura come la fotoferesi ha anche un impatto positivo sulla purezza delle cellule T CD4 + CD45RO +. Abbiamo osservato che la raccolta di campioni dopo la procedura di trattamento con fotoferesi ha un impatto negativo sulla resa delle cellule T CD4 + CD45RO +.
I cloni neoplastici di cellule T di pazienti con SS esprimono più frequentemente marcatori di superficie coerenti con un fenotipomaturo di cellule T CD4 di memoria 29,30. Tuttavia, la plasticità fenotipica è stata occasionalmente osservata rispetto ai marcatori di superficie tra cui CD4, CD45RO, CD45RA, CD7 e/o CD2631. Studi precedenti hanno anche dimostrato l'eterogeneità nell'espressione di CD45RO e CD45RA tra i pazienti con SS29, mentre la maggior parte dei casi di SS sono ancora CD45RO+. Roelens et al.31 hanno anche dimostrato che la SS può presentare eterogeneità interindividuale e intraindividuale con popolazione mista di sottoinsiemi naïve (TN), central memory (TCM), transitional memory (TTM), effector memory (TEM) e terminal effector memory (TEMRA). Tuttavia, i loro risultati mostrano chiaramente che la maggior parte delle cellule SS ha fenotipo TCM. Abbiamo focalizzato il nostro studio sull'immunofenotipo di superficie CD45RO+ più comune nei pazienti con SS e confermato il fenotipo mediante citometria a flusso. Negli studi di pianificazione dei sottoinsiemi di cellule T nei pazienti, è importante considerare l'eterogeneità fenotipica della malattia studiata e la strategia di purificazione può quindi essere regolata secondo necessità per ottenere la popolazione di cellule T desiderata per l'analisi.
Esistono diversi modi per stimolare le cellule T e le PBMC a esaminare l'espressione genica funzionale. Preferiamo l'attivazione chimica (PMA + A23187 ionoforo), poiché siamo interessati alla regolazione genica nel nucleo. L'attivazione chimica è l'opzione migliore per questo scopo perché agisce come un ampio attivatore ed è più uniforme rispetto alla stimolazione antigene specifica. La PMA è un piccolo composto organico che si diffonde attraverso la membrana cellulare nel citoplasma e attiva direttamente la proteina chinasi C. A23187 consente al calcio di passare attraverso le membrane. Questi composti bypassano i recettori di superficie e insieme imitano gli effetti della legatura del recettore delle cellule T con la co-stimolazione mediata da CD28. Le sostanze chimiche attivano diverse vie di segnalazione intracellulare, con conseguente attivazione del fattore di trascrizione nucleare e upregulation dei geni delle citochine che sono accessibili all'attivazione della trascrizione. Sebbene l'attivazione chimica e la legatura CD3CD28 producano profili di espressione genica globale sorprendentemente simili nelle cellule normali32, l'attivazione chimica con PMA + A23187 è una buona scelta poiché le cellule T SS possono perdere l'espressione dei recettori di superficie, compresi i componenti TCR33. Chong et al.22 hanno confrontato l'attivazione dei geni delle citochine tra PMA/A23187 e gli anticorpi anti-CD3 e anti-CD28 nelle PBMC di pazienti normali, precoci MF/CTCL e tardivi con MF/CTCL. Hanno riferito che PMA / A23187 ha causato un'attivazione più rapida e intensa del gene IL-2 rispetto alla stimolazione anti-CD3 / CD28. Inoltre, hanno dimostrato che la cinetica di attivazione più lenta con anticorpi anti-CD3 / CD28 è potenzialmente da cross-linking e segnalazione di membrana necessaria per la stimolazione. Inoltre, le tendenze nell'espressione delle citochine tra le diverse popolazioni cellulari studiate sono state preservate con PMA/A23187. Poiché siamo interessati all'attivazione dell'espressione genica, la stimolazione chimica è un approccio ideale perché agisce come un ampio attivatore ed è più coerente rispetto alla stimolazione antigene specifica. La legatura CD3/CD28 è ideale per studiare percorsi importanti nella trasduzione del segnale basata sulla membrana. Inoltre, l'attivazione chimica è meno costosa e non richiede attrezzature speciali. Nel presente studio, PMA + A23187 ha attivato in modo significativo i geni delle citochine nelle cellule T ND ma non SS, suggerendo che le cellule T SS hanno carenze funzionali a valle del TCR.
In sintesi, questo protocollo fornisce cellule T fenotipicamente pure da prezioso sangue derivato dal paziente e un metodo per valutare i cambiamenti a livello di genoma nell'espressione genica funzionale. Dimostriamo che il profilo trascrittomico delle cellule T SS rispetto alle normali cellule T CD45RO+ rivela profonde differenze nell'attivazione genica nelle cellule T umane fresche da pazienti con CTCL. Questi studi aiuteranno lo sviluppo di biomarcatori diagnostici e strategie terapeutiche mirate a nuovi marcatori in CTCL. Inoltre, questa strategia e protocollo nello studio delle cellule T umane primarie può essere utile per adattarsi agli studi di altre malattie mediate dalle cellule T.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Divulgazione etica:
Questo protocollo di ricerca è stato approvato dall'Institutional Review Board (IRB) dell'Università dell'Arkansas per le scienze mediche (UAMS, Little Rock, AR) I dati dei microarray presentati in questo studio sono stati effettuati sui campioni reclutati nell'ambito di un protocollo di ricerca approvato dall'IRB dell'Henry Ford Hospital (Detroit, MI).
Riconoscimenti
Ringraziamo i pazienti e i volontari che hanno partecipato alla nostra ricerca.
Materiali
| Name | Company | Catalog Number | Comments |
| 1.5 ml microcentrifuge tubes | Fisher Scientific | 02-681-320 | |
| 10 ml Disposable Plastic Pipette | Thermo Scientific | 170356 | |
| 1000 ul Pipet tips | VWR | 10017-038 | |
| 15 ml Conical Tubes | Corning | 352196 | |
| 25 ml Disposable Plastic Pipette | Thermo Scientific | 170357 | |
| 5 ml Disposable Plastic Pipette | Thermo Scientific | 170355 | |
| 50 ml Conical Tubes | Thermo Scientific | 339652 | |
| A23187 ionophore | Fisher Scientific | BP595 | |
| Centrifuge | Thermo Scientific | 75004381 | |
| DMSO | Sigma | D2650-5x5ml | |
| EasySep Human Memory CD4+ T cell Enrichment Kit | StemCell | 19157 | |
| FBS | GIBCO | 16140-071 | |
| HBSS | GIBCO | 14185-052 | |
| HEPES | Fisher Bioreagents | BP310-500 | |
| KHCO3 | Fisher Bioreagents | P184-500 | |
| Lymphocyte Separation Medium | Corning | 25-072-CV | |
| Na2EDTA | ACROS | 10378-23-1 | |
| NaHCO3 | Fisher Bioreagents | S233-500 | |
| NH4Cl | Fisher Bioreagents | A661-500 | |
| Penicillin-streptomycin solution | GIBCO | 15140122 | |
| phorbol12-myristate13-acetate (PMA) | Sigma | P-8139 | |
| Pipet-Lite LTS Pipette L-1000XLS+ | RAININ | 17014382 | |
| Pipet-Lite LTS Pipette L-10XLS+ | RAININ | 17014388 | |
| Pipet-Lite LTS Pipette L-200XLS+ | RAININ | 17014391 | |
| Pipet-Lite LTS Pipette L-20XLS+ | RAININ | 17014392 | |
| RNA Clean & Concentrator-5 | Zymo Research | R1013 | |
| Rneasy Plus Mini Kit | Qiagen | 74136 | |
| RPMI 1640 | GIBCO | 31800-022 | |
| T-25 Flask | Thermo Scientific | 2024-10 |
Riferimenti
- Kim, E. J., et al. Immunopathogenesis and therapy of cutaneous T cell lymphoma. Journal of Clinical Investigation. 115, 798-812 (2005).
- Wong, H. K., et al. Evolving Insights in the Pathogenesis and Therapy of Cutaneous T-cell lymphoma (Mycosis Fungoides and Sezary Syndrome). British Journal of Haematology. 155 (2), 150-166 (2011).
- Whittaker, S. Biological insights into the pathogenesis of cutaneous T-cell lymphomas (CTCL). Seminars in oncology. 33 (3), 3-6 (2006).
- Kallinich, T., et al. Chemokine receptor expression on neoplastic and reactive T cells in the skin at different stages of Mucosis Fungoides. Journal of Investigative Dermatology. 121 (5), 1045-1052 (2003).
- Rodd, A. L., et al. Current and Emerging Therapeutics for Cutaneous T-Cell Lymphoma: Histone Deacetylase Inhibitors. Lymphoma. 2012, 1-10 (2012).
- Olsen, E., et al. Revisions to the staging and classification of mycosis fungoides and Sezary syndrome: a proposal of the International Society for Cutaneous Lymphomas (ISCL) and the cutaneous lymphoma task force of the European Organization of Research and Treatment of Cancer (EORTC). Blood. 110, 1713-1722 (2007).
- Hameetman, L., et al. EPHA4 is overexpressed but not functionally active in Sézary syndrome. Oncotarget. 6 (31), 31868-31876 (2015).
- Willemze, R., et al. WHO-EORTC classification for cutaneous lymphomas. Blood. 105, 3768-3785 (2005).
- Bradford, P. T., et al. Cutaneous lymphoma incidence patterns in the United States: a population-based study of 3884 cases. Blood. 113, 5064-5073 (2009).
- Wilson, L. D., et al. Age, race, sex, stage, and incidence of cutaneous lymphoma. Clinical Lymphoma Myeloma and Leukemia. 12, 291-296 (2012).
- Li, Y., et al. Management of cutaneous T cell lymphoma: new and emerging targets and treatment options. Cancer Management and Research. 4, 75-89 (2012).
- Korgavkar, K., et al. Changing incidence trends of cutaneous T-cell lymphoma. Jama Dertmatology. 149 (11), 1295-1299 (2013).
- Dulmage, B. O., Geskin, L. J. Lessons learned from gene expression profiling of cutaneous T-cell lymphoma. British Journal of Dermatology. 169 (6), 1188-1197 (2013).
- Boonk, S. E., et al. Evaluation of Immunophenotypic and Molecular Biomarkers for Sézary Syndrome Using Standard Operating Procedures: A Multicenter Study of 59 Patients. Journal of Investigative Dermatology. 136 (7), 1364-1372 (2016).
- Scarisbrick, J. J., et al. Cutaneous Lymphoma International Consortium Study of Outcome in Advanced Stages of Mycosis Fungoides and Sézary Syndrome: Effect of Specific Prognostic Markers on Survival and Development of a Prognostic Model. Journal of Clinical Oncology. 10 (33), 3766-3773 (2015).
- Benoit, B. M., et al. CD164 identifies CD4+ T cells highly expressing genes associated with malignancy in Sézary syndrome: the Sézary signature genes, FCRL3, Tox, and miR-214. Archives of Dermatological Research. 309, 11-19 (2017).
- Dulmage, B., et al. The biomarker landscape in mycosis fungoides and Sézary syndrome. Experimental Dermatology. 26 (8), 668-676 (2017).
- Pick, E., et al. Intracellular Mediation of Lymphokine Action: Mimicry of Migration Inhibitory Factor (MIF) Action by Phorbol Myristate Acetate (PMA) and the Ionophore A23187. Annals of the New York Academy of Sciences. 94 (332), 378-394 (1979).
- Chong, B. F., et al. Induced Sézary syndrome PBMCs poorly express immune response genes up-regulated in stimulated memory T cells. Journal of Dermatological Science. 60 (1), 8-20 (2010).
- Fargnoli, M. C., et al. Diminished TCR signaling in cutaneous T cell lymphoma is associated with decreased activities of Zap70, Syk and membrane-associated Csk. Leukemia. 11, 1338-1346 (1997).
- Hansen, E. R., et al. Leukemic T cells from patients with cutaneous T-cell lymphoma demonstrate enhanced activation through CDw60, CD2, and CD28 relative to activation through the T-cell antigen receptor complex. Journal of Investigative Dermatology. , 667-673 (1993).
- Chong, B. F., et al. Immune Function Abnormalities in Peripheral Blood Mononuclear Cell Cytokine Expression Differentiates Stages of Cutaneous T-Cell Lymphoma/Mycosis Fungoides. Clinical Cancer Research. 14 (3), 646-653 (2008).
- Moerman-Herzog, A. M., et al. Transcriptome analysis of Sézary syndrome and lymphocytic-variant hypereosinophilic syndrome T cells reveals common and divergent genes. Oncotarget. 10, 5052-5069 (2019).
- Fanok, M. H., et al. Role of Dysregulated Cytokine Signaling and Bacterial Triggers in the Pathogenesis of Cutaneous T-Cell Lymphoma. Journal of Investigative Dermatology. 138, 1116-1125 (2018).
- Su, M. W., et al. Aberrant expression of T-plastin in Sezary cells. Cancer Research. 63, 7122-7127 (2003).
- van Doorn, R., et al. Aberrant expression of the tyrosine kinase receptor EphA4 and the transcription factor twist in Sezary syndrome identified by gene expression analysis. Cancer Research. 64, 5578-5586 (2004).
- Booken, N., et al. Sezary syndrome is a unique cutaneous T-cell lymphoma as identified by an expanded gene signature including diagnostic marker molecules CDO1 and DNM3. Leukemia. 22, 393-399 (2008).
- Dagur, P. K., McCoy, J. P. Collection , storage, and preparation of human blood cells. Current Protocol Cytometry. 73, 1-16 (2016).
- Fierro, M. T., et al. Heterogeneity of Circulating CD4+ Memory T-cell Subsets in Erythrodermic Patients: CD27 Analysis Can Help to Distinguish Cutaneous T-cell Lymphomas From Inflammatory Erythroderma. Dermatology. 216 (3), 213-221 (2008).
- Campbell, J. J., et al. Sézary syndrome and mycosis fungoides arise from distinct T-cell subsets: a biologic rationale for their distinct clinical behaviors. Blood. 116 (5), 767-771 (2010).
- Roelens, M., et al. Circulating and skin-derived Sezary cells: clonal but with phenotypic plasticity. Blood. 130 (12), 1468-1471 (2017).
- Diehn, M., et al. Genomic expression programs and the integration of the CD28 costimulatory signal in T cell activation. Proceedings of the National Academy of Sciences of the United States of America. 99 (18), 11796-11801 (2002).
- Fuji, K. New therapies and immunological findings in cutaneous T-cell lymphoma. Frontiers in Oncology. 8, 12-28 (2018).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati