
Method Article
決定の 3' 末端と HIV-1 の間に初期の一本鎖のウイルス DNA 分子のシーケンスを逆に感染細胞における転写
要約
初期 3' 末端の公平な測定だけでなく、単一座礁させた DNA の分子の変異プロファイルを提供するディープ シーケンス アプローチをご紹介します。メイン アプリケーションは、初期のレトロ ウイルス相補的 Dna (Cdna)、レトロ ウイルスの逆転写の過程で生成される中間体の特性です。
要約
ウイルスの複製中に核酸中間体の監視ウイルスの DNA 合成の効果および抗ウイルス化合物とホスト細胞タンパク質の作用メカニズムに洞察力を提供します。ここでウイルス感染症の生理学的なコンテキスト内でレトロ ウイルスの逆のトランスクリプション中間体を定義することができる細胞ベース、カバー力が高く、高解像度のアッセイの欠如に取り組みます。記載されているメソッドは、単一のヌクレオチドの解像度での HIV-1 感染細胞内で初期相補的 DNA (cDNA) 分子の 3' テルミニをキャプチャします。プロトコルを含む全細胞の DNA ハイブリッド キャプチャを介してウイルス DNA、アダプター結紮、ゲル精製、PCR 増幅、ディープ シーケンスとデータ解析によるサイズ分別の対象となる濃縮の収穫します。重要なステップは、3'-DNA テルミニを開くアダプター分子の効率的かつ公平な結紮です。記載されているメソッドのアプリケーションは、ある特定のサンプルのそれぞれの特定の長さの逆の転写の豊富さを決定します。それはまた任意の潜在的な突然変異により逆のコピーの (内部) シーケンス変化に関する情報を提供します。一般に、アッセイは、テンプレートの順序は知られている、DNA 3'-拡張に関する質問に適しています。
概要
分析し、ウイルスの複製に、ますます洗練されたレプリケーションをキャプチャ技術を理解するために、中間体が必要です。日付に多くのウイルスの複製機構を持っているので特に、ウイルス核酸種感染細胞のコンテキスト内での正確な定義は新しい洞察力を提供できるされて分離の in vitro反応を検討しました。典型的な例は、ひと免疫不全ウイルス (HIV-1) 1 など、レトロ ウイルスの逆転写の過程です。浄化された蛋白質とプライマー延長の試金を中心に学び、核酸酵素ウイルス逆転写酵素 (RT) が二本鎖 DNA に一本鎖 RNA ゲノムをコピー中に、HIV 1 逆のトランスクリプションの様々 なステップがされています。酸1,2,3,4,5。基本原則が確立された、このような試金すべてのウイルスと細胞成分を内蔵していないと関与因子の生物学的関連性の高い結果を反映していない可能性があります。したがって、正確な cDNA 3' 末端 (すなわち、彼らの正確な長さを決定する) と生活の伝染のコンテキストで塩基配列の逆のトランスクリプション中間体のスペクトルを決定するための強力な手法を考案しました。セル6。抗ウイルスの分子や蛋白質が効率性と DNA 合成のドメインに影響を与える可能性がありますの存在など、様々 な条件下での成績証明書のプロファイルを比較する実験・実習を利用できる時間からのデータ収集、蓄積。これにより標的薬剤設計および成功した治療的介入の基礎は、しばしば自然の病原体のライフ サイクルの詳細を理解します。
HIV 1 逆転写する一連のマイナス鎖の強力な停止と呼ばれる短い一本鎖 cDNA のトラン スクリプトを生成する RT 延長はゲノムの RNA テンプレートに tRNA プライマーのアニーリングによる連続イベント (-sss) (参照してください図 1)。その後、sss cDNA リピート (左から右) 5'-長いターミナルから、焼鈍、機能継続の RT のプライマーはマイナスの伸びを媒介として鎖 DNA ゲノムの RNA の 3' LTR に転送されます (逆のトランスクリプション1のレビューを参照してください。,2,3,4). この最初の鎖の転送は逆のトランスクリプションの率制限ステップの 1 つしたがって、蓄積する sss cDNA が知られています。感染細胞の逆のトランスクリプション製品をキャプチャする基本的なワークフローとライブラリの設計は、図 2 aに記載されています。特異的プライマーとプロトコルで使用され、表 1に記載されている設定はすべてを早くターゲット分析逆転写 cDNA 中間長さ範囲 23 ~ 650 の nt は、180-182 nt - sss DNA にはが含まれています。ただし、適切なマイナーな適応戦略は、後半の逆のトランスクリプション製品だけでなく、他のウイルスやシステムへの応用目的は 3' OH 含む DNA 終了を検出できます。考慮する重要な制限事項は、ライブラリの最終的な PCR の製品の長さの範囲特に、オープン 3'-末端のアダプターと上流プライマー サイト間の距離が 1000 nt を超えるテンプレートが可能性が高いより効率的にシーケンス、ライブラリの準備 (の間に技術的なバイアスを誤解を招く潜在的導入詳細と適応提案についての説明を参照)。
以前核酸鎖の 3'-テルミニの系統的定量のための報告の技術は、DNA ではなく RNA を分子に焦点を当てています。1 つの例は 3' レース (cDNA の端の急速な拡大)7mRNA の起こるに依存しています。さらに、RLM レース (RNA のリガーゼ仲介競争)8またはレース (cDNA の端の結紮による増幅)9含まれている RNA リガーゼを用いたアダプター結紮ベース戦略は開発されました。結紮ベースの拡大、ligation の反作用自体によって導入された任意のバイアスに敏感であることを強調することが重要です。たとえば、結紮は、3' の位置、シーケンス、合計の分子の長さ、または局所構造の特定の塩基配列によってもっとまたはより少なく効率的かもしれません。このようなリガーゼの好みは分子と私たちと他の人は9,10を観察している読み出しの不実表示の不完全なキャプチャに します。記載プロトコルでは、アダプターに追加の手順の中に、結紮バイアスを最小限に抑える、我々 は結紮戦略の数をテストし、T4 DNA リガーゼ ヘアピン一本鎖 DNA アダプターでの使用を発見した (國らによって記述します。11) ライゲーション効率制御オリゴヌクレオチド6の特別に選択したセットで評価するときに有意差には至らなかった定量的結紮の近くとプロシージャだけをします。この結紮戦略の選択は、したがって、このプロトコルの成功に重要な機能です。
日には、感染細胞内の HIV-1 RT 進行の監視主によって達成されている一意に測定短いまたは長いプライマー-プローブ セットを使用して量的な PCR (qPCR) と様々 な長さの逆のトランスクリプション製品を測定 (初期と後半、それぞれ) cDNA 製品12,13,14。この qPCR のアプローチはセルラ システムにおける逆のトランスクリプション プロセスの本質的な効率を決定する適切なのですが、出力は比較的低解像度の派生シーケンス情報がありません。我々 の新しいアプローチに基づいて最適化されたアダプター結紮、PCR を利用したライブラリを生成、およびアドレス ディープ シーケンス技術格差と定量的および単一のヌクレオチドの HIV-1 感染時に逆のトランスクリプションを監視する機会を提供しています解像度。
2 つ提案モデル間の HIV-1 制限因子 APOBEC3G の容量 (アテロームの mRNA 編集酵素触媒のポリペプチドのような 3 G) 干渉する識別における本法の有用性を説明している、ウイルスの逆のコピー6の生産。
プロトコル
注: 特定の試薬とこのプロトコルで使用する機器、材料表を参照してください。
1. ウイルス産生と細胞への感染
注意: 感染の HIV-1 は、承認されたバイオ封じ込め実験室でのみ処理必要があります。
注: ヒト胚性腎 (HEK) 293 t 細胞、に従って 1.1 をステップ、標準的な手順し、されているのトランスフェクションによる HIV 1 粒子の生産15,16前述。一般的な細胞培養プロシージャは、17をは前述。
- 1 HIV ウイルス産生。
- ダルベッコ 293 t 細胞変更 10% 牛胎児血清 (FBS) と 37 ° C、5% CO2 17を前述のように標準的な細胞文化のインキュベーターでペニシリン/ストレプトマイシン 1% (完全 DMEM) イーグル培 (DMEM) を維持します。
- 標準流ティッシュ文化フード成長媒体を削除し、293 t 細胞の合流付近 10 cm の細胞培養ディッシュ (~1.2 x 107セル) に予め温めておいた (37 ° C) のトリプシンの 3 mL を追加します。2-3 分のためのインキュベーターに皿を置きます。
- ティッシュ文化フードに戻ってインキュベーターから料理を取る、7 mL の完全培地を追加します。ピペット、上下皿内で数回に細胞を再懸濁します。新しい 10 cm 皿に細胞懸濁液 2.5 mL を追加してセル 1:4 を分割し、7.5 ml の完全培地のそれを埋めます。
- 次の日、無血清最小必須培地 1 mL でプロウイルス HIV-1 プラスミド DNA (pNL4.3) などの 10 μ g を混合し、1 μ g DNA あたり 4.5 μ L でポリエチレンイミン (PEI) ソリューション (25,000 mW、1 mg/mL pH 7)。室温で 10 分間インキュベートし、293 t 細胞に滴下し追加します。
- トランスフェクション後、24 h はメディアを取り出して、20 U/mL 中に RNase フリーの DNase を含む完全 DMEM の 6 mL を置き換えます。6 時間後、培地を完全 DMEM の 10 mL に置き換えます。
- トランスフェクション後、48 h 上清を収穫し、15 mL ポリプロピレン チューブに 10 mL の注射器を使用して、0.22 μ m フィルターをフィルター処理します。
- オープン トップ薄肉遠心チューブに 20% スクロースを滅菌リン酸緩衝生理食塩水 (PBS) x 1 の 2 mL を追加します。ゆっくりとフィルター処理された細胞培養上清にショ糖をオーバーレイします。
- 4 ° c、超遠心機を使用しての 134,000 x g で 1 時間 15 分間遠心します。
- 超遠心機からチューブを慎重に取り外します。徐々 に清とショ糖の吸引またはピペットを使用して電源を取る。小さいピペットを使用し、最後のショ糖液を取り出し時にチューブを傾斜します。チューブの底に小球形にされたウイルスを残します。
注: ペレットは表示されません。 - 1 × PBS を 200 μ l 添加、12 h 4 の冷蔵庫の中に残して、再懸濁します、-80 ° C で 20 μ L の因数でフリーズ
- P24ギャグコンテンツ使用を決定、p24 HIV 1 抗原検出 ELISA キット (次の製造元の指示)。
- T 細胞ラインの感染症。
- 不死化 T 細胞株を培養 (e.g。、CEM SS セル) 10% FBS と 1% ペニシリン/ストレプトマイシン (完全 RPMI) ロズウェル パーク記念研究所 (RPMI) 1640 培で。1 ml の 2 × 106セル/ml、12 ウェル フォーマット細胞培養プレートで完全 RPMI 検定18と種子 1 サンプルあたりもを使用してセルをカウントします。
- HIV-1 の粒子を追加 150 ng の p24 に相当ギャグと場所に揺れるプレート遠心分離機ベンチトップ遠心分離機 30 ° C で 2,000 x g で 2 時間の遠心分離でスピン感染する生物学的封じ込め蓋付きバケツ
- 遠心分離機からプレートを削除し、37 ° C、5% CO2で標準的な組織文化のインキュベーターで 1 時間休ませます。
- 入力ウイルスを洗い流すために細胞懸濁液を遠心管に転送して、2 分の 500 x g で RT (RT) で遠心機で遠心分離によって細胞を収集します。細胞ペレットを乱すことがなく上澄みを脱ぐ。
- 予め温めておいた (37 ° C)、1 mL の細胞ペレットを再懸濁します滅菌 1 × PBS。遠心、上清除去および再懸濁の手順を 2 回繰り返します。
- 再度遠心分離機、上澄みを除去し、完全 RPMI の 1 mL の細胞ペレットを再懸濁します。新しい 12 ウェル プレートの 1 つの井戸にそれぞれの懸濁液を追加します。
- ウイルス (4 h 後遠心分離) の 6 h 後最初に加えでステップ 1.2.4 で遠心分離によってセルを収穫します。削除し、上澄みを廃棄します。細胞ペレットを-80 ° C で凍結または DNA の抽出のために直接処理することができます。
2. DNA 抽出、HIV 1 DNA の定量化、およびハイブリッド キャプチャで濃縮
- ティッシュ文化セルのキット マニュアルに従うことによって、血液や組織総 DNA 抽出キットの全細胞の DNA を抽出します。唯一の変化は、ヌクレアーゼ フリー H の 200 μ L の溶出2O 提供溶出バッファーの代わりに。
注: カオトロ ピック換散バッファー (キットの"アル") とプロティナーゼの付加の後でサンプル バイオ セーフティ封じ込め実験室から削除できプロトコルの残りのための標準的な安全レベルの研究室で処理されます。 - QPCR による HIV 1 cDNA のコピーの数を決定します。
- ステップ 2.1 溶出の 17 μ L を取り出して DpnI 制限酵素の 1 μ L と 10 x 制限酵素バッファーの 2 μ L を追加します。任意の潜在的なの残留は、入力プラスミッド DNA をトランスフェクションから削除する 37 ° C で 1 時間インキュベートします。
- 次プライマー プローブ セットを使用してマイナス鎖強い停止 cdna qPCR 遂行: oHC64 (3 '5'-taactagggaacccactgc-) と oHC65 (3 '5'-gctagagattttccacactg-) と oHC66 をプローブ (5 '-ファム-acacaacagacgggcacacacta-タムラ-3 ')。qPCR セットアップと正確な条件は、参照6,13で見つけることが。CDNA 分子のコピー数を決定するための標準的な曲線として pNL4.3 プロウイルス プラスミドのシリアル希釈試料に沿って運ぶ。
注: 予想される量のための議論を参照してください。
- ハイブリッド キャプチャによって HIV 1 DNA 濃縮。
注: このステップからそれはエアロゾル フィルター ピペット チップと同様、低核酸結合特性と全ての DNA サンプルのマイクロ遠心チューブ用を使うことが望ましい。可能であれば、PCR のワークステーションで作業します。すべてのステップと試薬は、特に明記しない限り、RT (RT) でです。- 磁気ストレプトアビジン ビーズのマスター ミックスを準備するには、単一微量遠心チューブにサンプルあたり 100 μ L ビーズをピペットします。マイクロ遠心チューブ用に適した磁石にチューブを置きます。
- ビーズは、チューブ (~ 1 分) の磁石側に落ち着いて後、ストレージ バッファーを脱いで、磁石からチューブを削除しバインドを 500 μ l 添加でビーズを再懸濁します、洗浄バッファー (BW バッファー、5 mM トリス塩酸 pH 7.5、0.5 mM EDTA、1 M 塩化ナトリウム) を洗浄します。
- 背面マグネット チューブを置き、上澄みを除去し、500 μ L カゼイン溶液を追加します。磁石を取る、再懸濁します、常温では、10 分間インキュベートし、BW バッファーで洗浄します。
注意: 洗浄は、磁石にチューブを配置する、上澄みを取って、チューブを取って磁石、オフ、バッファーを追加して、再です。 - 背面磁石チューブを置き、上澄みを取る、BW バッファーを 500 μ l 添加のビードを再停止しなさい。サンプルあたり (この場合は表 1、3 oligos を参照) の各キャプチャのビオチン標識オリゴヌクレオチドの 50 pmol を追加します。(たとえば、5 の DNA のサンプルを処理する場合は、使用して 500 μ L 各オリゴヌクレオチドのステップ 2.3.1 と 250 pmol から磁気ビーズの)。
- エンド オーバー エンド ミキサーでロッキングしながら室温で 30 分間インキュベートします。
- 1 x 10 バッファー (10 mM トリス塩酸 pH 8.0、1 ミリメートルの EDTA、100 mM の NaCl) の 500 μ L で 2 回固定化オリゴヌクレオチドとビーズを洗浄します。
- 10 μ L のサンプルあたり 1 x 10 バッファーのビードを再懸濁します。
- 各サンプルのラベル 1 つ遠心チューブ、ビーズ懸濁液の 10 μ L、170 μ L の DNA の追加 (手順 2.1) から、3 x 10 のバッファーの 90 μ L。DNA を変性する 2 分 92 ° C で乾燥した熱ブロックで孵化させなさい。
- 52 ° C に設定すると、別の乾熱ブロックにチューブを移動し、1 h. 定期的にミックスする反転インキュベート (~ 10 分毎) この潜伏中。
- 500 μ L x 10 のバッファー 1 ので 1 回洗浄し、ヌクレアーゼ フリー 35 μ L で再懸濁します H2o.
- 溶出が、92 ° c を 2 分間の乾熱ブロックでチューブをインキュベートします。マグネット (一度に 1 つの管) にチューブをすばやく移動します。チューブの側面にビーズをバインドすると、新鮮なチューブに HIV 1 DNA を含む上清を転送します。
- オプション: 繰り返して qPCR (2.2.2 のステップで実行) 回復の HIV-1 cDNA を決定します。
注: 予想される量のための議論を参照してください。
3. アダプター結紮
- アダプターを準備します。
- 凍結乾燥のアダプターを再懸濁します (表 1 「完全國 + MiSeq」参照) ヌクレアーゼ フリー H で 100 μ m で2o.
- サンプル、コントロールは 1 つのサンプルあたり 0.45 μ L を組み合わせて T4 DNA リガーゼ バッファー、アダプターの 4 μ L および H のヌクレアーゼ フリーの 0.05 μ L x 10 の22 分の 92 ° c o. 熱せの各クールダウンゆっくり。
注: オプションが利用可能な場合は、調節可能な冷却率 (使用率 2%) と PCR 機械を使用します。これは 92 ° C から 16 ° C に約 30 分かかりますまた、92 ° C で乾燥した熱ブロックを使用しをオフにします。熱ブロックは右に戻るときアダプター マスター ミックスを取り出すこれはアダプターのヘアピン構造を形成させる (図 2bを参照)。
- 一連の細胞から抽出した DNA の代わりに合成オリゴヌクレオチド (表 2参照) と制御反応を準備します。
- 各オリゴヌクレオチドの 100 μ M の在庫を確認します。17 oligonucelotides のそれぞれの 1 μ L を混合し、25 μ L の最終巻で H2O のモル比の 8 μ L を追加します。
- ヌクレアーゼ フリー H でミックス 1:2,500 を希釈2O シリアル希釈で。ヌクレアーゼ フリー h 17.3 μ L とミックスの 1 μ L を組み合わせて各オリゴヌクレオチドは 1.6 fmol であるのでステップ 3.3.1 でコントロール サンプル結紮に使用する2O (0.026 に相当 60 μ L 反応 nM)。
- を設定
- PCR チューブに 60 μ L 最終巻反応結合 T4 DNA リガーゼ x 10 の 6 μ L のバッファー、40% の 24 μ L ペグ、5 M のベタイン、4.5 μ (400 pmol) アダプター (前 annelead ステップ 3.1.2 のように)、6 μ T4 DNA リガーゼ (2,000,000 単位/mL) の 1.2 μ L、(からの DNA の 18.3 μ L2.3.11 のステップ)
注: 粘性溶液の 40% など特別な注意を取る量を正確に維持するためにペグ。マスター ミックスをしないようにします。 - 3.3.1 のステップが準備した 3.2.2 制御オリゴヌクレオチド ミックスを実行、同じ反応を設定します。
- 反応をよく混ぜ、一晩 16 ° C で PCR 機で孵化させなさい。
- PCR チューブに 60 μ L 最終巻反応結合 T4 DNA リガーゼ x 10 の 6 μ L のバッファー、40% の 24 μ L ペグ、5 M のベタイン、4.5 μ (400 pmol) アダプター (前 annelead ステップ 3.1.2 のように)、6 μ T4 DNA リガーゼ (2,000,000 単位/mL) の 1.2 μ L、(からの DNA の 18.3 μ L2.3.11 のステップ)
4. アダプターの取り外しとサイズ分離
- ゲルの電気泳動の変化
- それぞれの ligation の反作用にローディングバッファー ホルムアミドを含む DNA ゲルの 30 μ L を追加します。ピペッティングでよく混ぜます。
- PCR マシンで 94 ° C で 2 分加熱し、すぐに氷の上に置きます。
- 適切なゲル タンクにプレキャスト 6% トリス/ホウ酸/EDTA (TBE) 変化尿素ポリアクリルアミドのゲル (10 よく櫛) を配置します。1 x TBE (89 mM 89 mM ホウ酸トリス ベース 2 ミリメートルの EDTA) 連続したバッファーを追加し、前 250 V/最大定数で 20 分のためのゲルを実行します。
- 21 G 針とシリンジを使用してバッファーを実行するとゲルのポケットを洗い流します。
- 負荷 20 分 (250 V/最大) 濃い青色色素前面まで 3 つの井戸 (30 μ L/ウェル) に各 90 μ L のサンプルはゲルを通って途中でです。
- 染色・加工核酸ゲルから
- 21 G 注射針 (注射針などでの作業中を取る注意) を使用して下部に穴を突っついて、サンプルあたり 3 小さなマイクロ遠心チューブ用 (0.5 mL) を準備します。準備されたチューブ 2.0 mL 遠心チューブに挿入し、「ミッド」または「高」サンプル名に加えて、「低」のラベルを付けます。
- 取り出して、ゲル カセットをこじ開けます。惜しみなくロードされたサンプルの 3 の井戸とストリップを消費税にかみそりの刃と垂直方向にゲルをカットします。1 x TBE (約 30 mL) とシアニン核酸染色の 5 μ L の容器にゲルのストリップを追加します。3-5 分間インキュベートします。
注: ゲル抽出手順は、交差汚染に特に敏感です。ゲルおよび各ゲルの汚損のため独立したきれいなコンテナーの使用につき 1 サンプルをのみ実行することをお勧めします。ゲル粒子が手袋をはめた指を連絡する場合、手袋を変更必要があります。 - DdH2o. で徹底的に青い光の transilluminator の表面をきれいに染色容器からゲル部分を取り、それをライト ボックスに追加します。
- ライト ボックスをオンにし、オレンジ色のフィルターを通してステンド グラスの核酸を検査します。
注: アダプターは、通常オーバー ロードされた表示され、連勝として実行する結紮の HIV-1 の DNA と大きな"blob"として実行されます。 - ないサンプルの区域がある場合は、ゲルの側面を切り取る新しいかみそりの刃を使用して今も読み込まれます。次に、アダプターを削除して、ゲルの部分の下にアダプター上だけカットします。最後に、切り取るゲルなどの最上部約 1 mm のゲルのポケットには、しばしば高分子量 DNA の鋭い強烈な信号があります。
- サンプルでは、これは通常 2 〜 3 倍を含む残りのゲル部分を分割 cm 三個でも水平方向にサイズ:「低」、「中」、「高」分子量領域。
注: 各部分は今は個別に処理 [i.e。、(低、中、および高) 3 つの管がある] 元のサンプルあたり。 - 小さな断片 (~ 2 × 2 mm 粒子) にそれぞれの 3 つのゲル断片をカットし、準備中の 0.5 mL 遠心チューブ (ステップ 4.2.1) にそれらを転送します。
- ゲルの秘密を作成する 2 mL チューブに穴をゲル部分を絞る 1 分のオープン蓋をトップ スピードでスピンします。任意のゲル粒子が 0.5 mL チューブの底に残っている場合は、針またはピペットの先端を使用して手動で 2 mL チューブに転送できます。
- DNA の抽出
- ゲル スラッシュに尿素ゲル抽出バッファー (0.5 M NH4CH3CO2、1 ミリメートルの EDTA、0.2 %sds) の 1 つの mL を追加します。3 h 以上のチューブを回転 (一晩は許容) エンド オーバー エンド ミキサー常温。
- セルロース アセテート膜フィルター (0.2 μ m)、膜の目詰まりを避けると遠心分離機の列に 1 つの小さな丸いガラス繊維フィルターを追加するのにピンセットのきれいなセットを使用します。倒立ピペット先端のある場所でフィルターをかけます。
- 簡単に遠心機にゲルの秘密と抽出を 2 mL 管をスピンし、上澄みの 700 μ L を準備されたフィルター列に転送します。ゲルの秘密と残りの上清を維持します。
- 1 分新しい 2.0 mL 遠心チューブに転送、透水のトップ スピードでの遠心フィルター列を遠心します。
- 残りの上清を持つ列を再読み込みします。秘密の抽出からできるだけ多くの液体としてを取得しようとします。ゲル部分の転送は、重要ではないです。再びスピンし、同じ抽出サンプルの flowthroughs を組み合わせます。
- DNA の沈殿物
- ポリア RNA の 3 μ L を追加 (1 μ g/μ L; キャリアとして)、グリコーゲンと 4.3.5 のステップから、吸収にイソプロパノール 0.7 mL の 1 μ L。簡単に渦と一晩-80 ° C で凍結。
- -80 ° C のフリーザーのサンプルを取るし、簡単に解凍させます。冷却 (4 ° C) 遠心と最高速で 30 分のスピンに入れてください。
- 削除し、上澄みを廃棄します。ペレットを削除しないように非常に注意します。そのペレットがそれ以外の場合削除されるが不明である場合は、液体の 30 に 50 μ L をままにします。
注: 通常すべての「高」のサンプル表示「ミッド」、「低」のサンプルよりももっと目に見えるペレットです。 - 80% エタノールの 800 μ L を追加します。チューブとトップ スピードで 1 分にもう一度スピンを反転します。ピペットとエタノールの大半を削除、簡単にチューブをもう一度、スピンし、小さいボリューム ピペットとより多くのエタノールを削除します。
- 55 ° C の乾燥した熱ブロックに開くふた付きチューブを配置することによって蒸発させる残りのエタノールができます。サンプルのドライ (2 〜 4 分) 追加 20 μ L ヌクレアーゼ フリー h2O および DNA の餌を確保するためチューブの下部全体に広がりましたが解散したとき。DNA サンプルは-20 ° C で保存できます。
5. PCR 増幅とライブラリの準備
- DNA ポリメラーゼ プレミックスの 20 μ L で 40 μ L の PCR 反応を設定、18 μ L の沈殿し、ステップ 4.4.5、前方のプライマー「MP1.0 + 22HIV」の 1 μ l から DNA を再溶解 (10 μ m) (表 1を参照)、および多重オリゴ プライマー (インデックス プライマー 1 に 24) の 1 μ l ( Ma の表を参照丈)。
注: 3 つの反作用 (低中、高) が同じ別の PCR の反作用で各サンプルのプライマーをインデックス付けを実行します。元の感染サンプルの各別のインデックスを使用します。- 以下の条件で PCR の反作用を実行: 変性 94 ° C で 2 分し、3 ステップ pcr 法の 18 サイクル15 秒 94 ° C の変性、15 s は、55 ° C、および 68 の ° C で 30 s 拡張子の焼鈍
- 品質管理オプションとして自動高感度ゲル電気泳動システム、PCR 反応を分析します。2 μ L の製造元の指示に従ってを実行する低・中・高のサンプルを取る。
注: 2 つのプライマーは、表示され、多くの場合 (実際の長さとは異なります) 45 と 95 nt についての計算された長さで実行する必要があります。150 に 500 の間 DNA を検出必要がありますさらに、nt。信号が存在しない場合は、追加の 2 と 10 のサイクルの間、追加の PCR のサイクルを追加することをお勧めします。3.3.2 の手順で作成したオリゴヌクレオチド コントロール サンプルの追加サイクルを追加しないでください。 - 削除プライマーは常磁性ビーズを用いた PCR クリーン アップ システムを使用します。
- 各 PCR 反応 20 μ L を取り、(この時点ですべてのサンプルをミックス) 一緒にサンプルをプールします。-20 ° C でバックアップとして残りの 20 μ L 反応を凍結します。
- RT に来て、ミックス ビーズ溶液の量 x 1.8 プールされた PCR の反作用の常磁性ビーズをしましょう。ピペッティングで混ぜるし、5 分間インキュベートします。
注: 例、4 サンプルが準備された場合、それぞれ低・中・高の反応がある、ボリュームになる 20 μ L x 3 x 4 = 432 μ L ビーズ ソリューションと 240 μ L PCR の反作用。 - 遠心チューブ マグネットにチューブを入れて、ビーズ 〜 1 分、バインドし、離陸、上清を破棄します。磁石をチューブのまま、80% エタノール 500 μ L を追加します。
- 30 のエタノールのまま s、徹底的に離陸し、ヌクレアーゼ フリー h ~ 5 分追加 40 μ L のビーズ センサー2O、磁石からチューブを取るし、上下に複数回ピペットします。
- 磁石に戻って 5 分入れて懸濁液管を残して、ビーズ側に解決し、上清を新しいチューブに転送。これは、ライブラリです。10 μ L 分注精度管理用と-20 ° C で残りの部分を凍結
6. ライブラリの評価
- ライブラリの品質、濃度、モル濃度を決定します。
- 蛍光定量法を使用します。1 μ L と製造元の指示に従って高感度 dsDNA アッセイ キットとライブラリの 3 μ L を測定します。
注: 一般的な濃度は 1 〜 10 ng/μ L の間です。 - 測定高感度によってライブラリ DNA 分子スペクトルは、(ステップ 5.2) 上記のようにゲル電気泳動を自動化しました。
- 自動化されたゲルの電気泳動解析ライブラリの平均分子量を決定するため、ヌクレアーゼ フリー H でライブラリを希釈する計算使用2O 4 nM。すべてのインデックスが一意である限り、いくつかのライブラリを結合できます。
- 蛍光定量法を使用します。1 μ L と製造元の指示に従って高感度 dsDNA アッセイ キットとライブラリの 3 μ L を測定します。
- 省略可能なスループットの品質管理
- 増幅のためのベクトルにライブラリ分子を挿入する TA クローニング19の DNA ライブラリを対象します。キットの指示に従って、10 〜 20 を成長のコロニーそして抽出 DNAを介してminiprep のプロトコル、20をここで説明。
- サービスをローカル配列と挿入を含む目的の HIV-1 シーケンス ベクトルの派生シーケンスおよびライブラリ固有のアダプター。
7. 高スループット シーケンス実行
- シーケンスのプラットフォームが提供する商用ソフトウェアにシーケンスのサンプル シートを作成します。
- 選択したシーケンス キットを示します。通常、150 サイクル キットを選択、他は必要な読み取り距離に応じて適切なです。
- アプリケーション ワークフローとして「Fastq のみ」を選択します。(キット マニュアルに示される) 多重オリゴヌクレオチド キットに 24 のインデックスを格納するテンプレートのいずれかを選択します。
- 選択"25 nt」Read1 の「125 nt"Read2 の。単一インデックスを読むにしてください 6 nt。
注: 自社分析 Read2 のみが分析で使用されています。少なくとも 25 の Read1 を保つ nt プラットフォーム アルゴリズム目的のシーケンス処理。
- 実行前のライブラリの準備とセットアップについて正確に製造元の指示に従ってください。最大 20 を選ぶ pM 濃度と非常に低い複雑さはライブラリとして 15 %phix スパイク使用。
8. データ分析
- パス フィルターの割合と平均 Q30 品質スコアがシーケンス処理プラットフォーム製造元のガイドラインに従って許容されるを確認します。
注: フィルターは通常 > 90%、Q30 スコアは > 80% で、通常。 - ダウンロードします。 fastq.gz ファイルを製造元のシーケンス ハブから。
- シーケンス スクリプトの設定
- "AnalysisXYZ"をという名前の新しいディレクトリ (フォルダー) を作成し、このディレクトリにすべてのソース コード ファイル (parse_sam.pl、rc_extract.pl、parse.sh) をダウンロードする https://github.com/malimlab/seqparse に移動します。
- 同じディレクトリに http://bowtie-bio.sourceforge.net/index.shtml から短鎖リード アライナ、ボウタイ、バージョン 1.1.2 をダウンロードしてください。
- ダウンロードは、"AnalysisXYZ"「ボウタイ 1.1.2"をという名前サブディレクトリを作成します。このディレクトリ内にあるサブディレクトリ「インデックス」を開き、.ebwt 拡張子を持つ六つのファイルから成る提供されるテンプレート配列をダウンロードします。
- 処理前の"AnalysisXYZ"ディレクトリにツールキット fastx - http://hannonlab.cshl.edu/fastx_toolkit/download.html から 0.0.13 FASTQ/A 短いリードをダウンロードします。
- 「ドキュメント」ディレクトリに Samtools (https://sourceforge.net/projects/samtools/files/) と bam readcount (https://github.com/genome/bam-readcount) をダウンロードします。
- 移動します。 すべての 8.2、手順でダウンロードした fastq.gz ファイルは"AnalysisXYZ"ディレクトリに期... _R2_001.fastq.gz 2 s を読みます。
- コマンド コンソール/ターミナルを開きます。Cd コマンドを使用して現在のディレクトリとして"AnalysisXYZ"に移動します。スクリプトを実行する"./parse.sh"の種類。
- 合計読み取り数のすべてのサンプルの概要を含む .csv ファイルを見つける、長さが調整数を読み取りと読み取り数を正規化し同様"分析 XYZ"ディレクトリ内の parse_results という名前のディレクトリで各サンプルに対して、基本バリエーションを持つファイルします。
注: 分析プロセスについてのディスカッションを参照してください。スクリプトは、ヌクレオチド 635 までとは、HIV-1NL4.3強力な停止シーケンスと鎖の最初の転送に沿って各ヌクレオチドのため合計読み取り csv ファイルを返します。、ガイダンスとして、50,000、100,000 のユニークな読み取りは通常示された細胞数, ウイルス接種と抗ウイルスのタンパク質や化合物なしの感染症からのサンプルにおいてください。オリゴヌクレオチド コントロールのサンプルは通常 100,000、200,000 の読み取りを生成します。
結果
この資料で説明する手法は HIV-1 抗レトロ ウイルス人間蛋白質 APOBEC3G による逆転写 (A3G) 抑制のメカニズムに対処するための幅広い研究に適用される6。図 3は、 vif感染 CEM SS T 細胞からのサンプルでプロトコルを採用した代表的な結果を示しています-欠乏の HIV-1 A3G の有無で。同じ 6 を持っている PCR 重複除外後に各サンプルから得られるユニークな読み取りの合計数 nt バーコードと同じ長さ (提供される分析ソフトウェアで実行されます) は図 3を順にプロットされます。A3G の増加レベルは、A3G の抑制効果を介した RT cDNA を合成する以前に説明した、qPCR6,13,21,22で測定に反映して合計読み取り数を減らします。図 3b、最初 182 内それぞれの可能な長さで分子の割合 nt が表示されます。A3G の不在の HIV-1 感染、最も豊富な種はメイン 180 nt 強い停止分子自体は、いくつかの蓄積と読み取りの短い範囲で (23 に 40 nt) (青いグラフ ヒストグラム上)。このプロファイルは短く、急激な増加は、いくつかの非常に特定、再現可能な位置で cDNA 分子を切り捨てられてので、A3G 変更の追加 (中間および低いグラフ) が検出されました。A3G はシチジン アデノシンデアミナーゼ、シトシン-ウリジン (C ・ T として識別される) ので、A3G が感染しているウイルス粒子21,23,24cDNA の突然変異が発生します。得られた配列情報を使用して、C-T の突然変異の割合は (赤点線) の同じのグラフにプロットしました。変異プロファイルはすべて一意の読み取りを組み合わせるから派生し、各ヌクレオチドの報道とは異なることに留意。ただし、シーケンス情報を各分子に関連し、特定 3' 末端と相関できるために必要な場合。提供されるデータは、Pollpeterらから採取しました。6および突然変異の間の相関と cDNA 長さプロファイルを示した DNA 修復する機械の検出と細胞によって脱 cDNA の胸の谷間に起因します。
3'-マッピングのアプローチの肯定的な制御は、知られていたシーケンス、長さ、および濃度の総合的なオリゴヌクレオチドのプールを処理することにより簡単に生成できます。このコントロールは、3.3.2 の手順でアダプター結紮で追加され、多重のすべてのライブラリに含めることをお勧め。コントロールのサンプルから得られたデータは、予期される入力比、唯一の非常にマイナーなバック グラウンドの読み取りですべてのオリゴヌクレオチドをはずです。図 4は、17 の化学的合成オリゴヌクレオチドの肯定的な制御セットの結果を示しています (シーケンスは、表 2を参照)、モル比でミックスされました。予想通り、すべての分子がだけ小さな変化 (上のグラフ) と同じ豊富な近くに表示されます。オリゴヌクレオチドによる表されなかった sss DNA シーケンス内のほとんどのポジションを返す 0 の読み取り数、実際のコントロールのオリゴヌクレオチドより短い 1 または 2 nt のマイナーな種を見ました。我々 はこれらのマイナーな種をさらに調査していないが劣化または不完全な製品購入時指定されたオリゴヌクレオチド株に潜在的に存在を表すことを前提として (オリゴヌクレオチドは、HPLC 精製を命じられた、メーカーを示す > 80% の高純度)。下のグラフは、別のライブラリを実行すると、変化はわずか 17 オリゴヌクレオチドとの間に高いし、長い分子の制御がより効率的に検出されるという点で、全体の長さと相関し、短いものからコントロールのサンプルを示しています。これは PCR の反作用または挿入サイズ最適のある MiSeq シーケンス処理中にクラスタ リングでマイナーなバイアスが原因である可能性があります、特に広い挿入範囲を運ぶライブラリで発生する可能性があります。このバイアスに対処する基本的な方法は、分子の長さ (ピンクの線) にバイアスを示す斜面に基づいて正規化係数のアプリケーションです。必要な計算の解析プログラムに含まれている (プロトコル 8.3 を参照)。

図 1: HIV-1 の最初のステップを逆転写を示す図。開始とウイルス cDNA (青、ステップ 2) の伸長を可能にするゲノム ウイルス rna (ステップ 1)、tRNA(Lys,3) (オレンジ) のプライマー結合部位 (PBS) に焼鈍をまずします。付随して、テンプレートのゲノム RNA は RT (ステップ 3) の RNaseH 活性が低下します。逆転写の過程で最初の完全の中間ですマイナス鎖強い-(-) sss cDNA RT 触媒重合 gRNA リピート (R) 領域 (ステップ 3) の 5' 末端に達したとき完全であります。中間の (-) sss は、相補的 3'-長いターミナル リピート (LTR) R 領域にアニールによるゲノム RNA テンプレートの 3' 末端に転送されます。ここからは、重合は、(手順 4) を続けています。この方法で逆のトランスクリプションの進行は新生ウイルス cDNA (青) の正確な長さをマッピングによって決定されます。PPT、polypurine 管;U5、ユニークな 5'-シーケンス;U3、ユニークな 3'-シーケンス。この図は、以前文書6から再発行されます。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 2: ワークフローのアウトラインとアダプターの結紮と PCR 増幅戦略の概略図。(3'-HIV-1 の終点を決定する記述手法の主な手順の概要ワークフロー a) 逆感染細胞の転写物です。図は、以前の文書6から適応。(b) アダプターの ligation および PCR 増幅戦略の概略図。前の手順で精製されている様々 な長さの初期 cDNA 分子が T4 DNA リガーゼを使って単一座礁させた DNA アダプターに組み合わされて。ヘアピン アダプター (名前付き「フル國 + MiSeq」、表 1を参照) のデザイン國らに触発されました。11. アダプターを運ぶランダム 6 nt バーコード シーケンス ベース ペアリング結紮を容易にすることができますし、同時に識別子として一意の読み取りを提供しています。アダプター 3' 末端は、スペーサー (SpC3) 自己 ligation を防ぐために運ぶ。結紮製品は、余分なアダプターから変性ポリアクリルアミドゲル電気泳動 (ページ) で区切られます。核酸ゲルで、ステンド グラスし、25で井戸の中にアダプターから地区別、等しいサイズのゲル 3枚にカットします。溶出・降水量・巻き上がり後、製品は、アダプターの知られていたシーケンスにアニールするプライマーで増幅した PCR (プライマー 1、多重オリゴヌクレオチド キット、材料表を参照してください)、最初の 22 を運ぶプライマー HIV-1 の nt5'-LTR シーケンス (プライマー 2、MP1.0 + 22HIV) tRNA の直後します。選択したプライマーの 5' テルミニは、同じライブラリで実行する個々 のサンプルを区別するためにインデックス シーケンスだけでなく、選択したシーケンスのプラットフォーム (P5 と P7) 用のアダプターを運ぶ。シーケンスの開始ポイントを読むプライマーが示されます。青いボックスは、オリジナル 3' 末端キャプチャされた分子の決定に関心の領域を示します。この図は、以前の文書6から適応。この図の拡大版を表示するのにはここをクリックしてください。
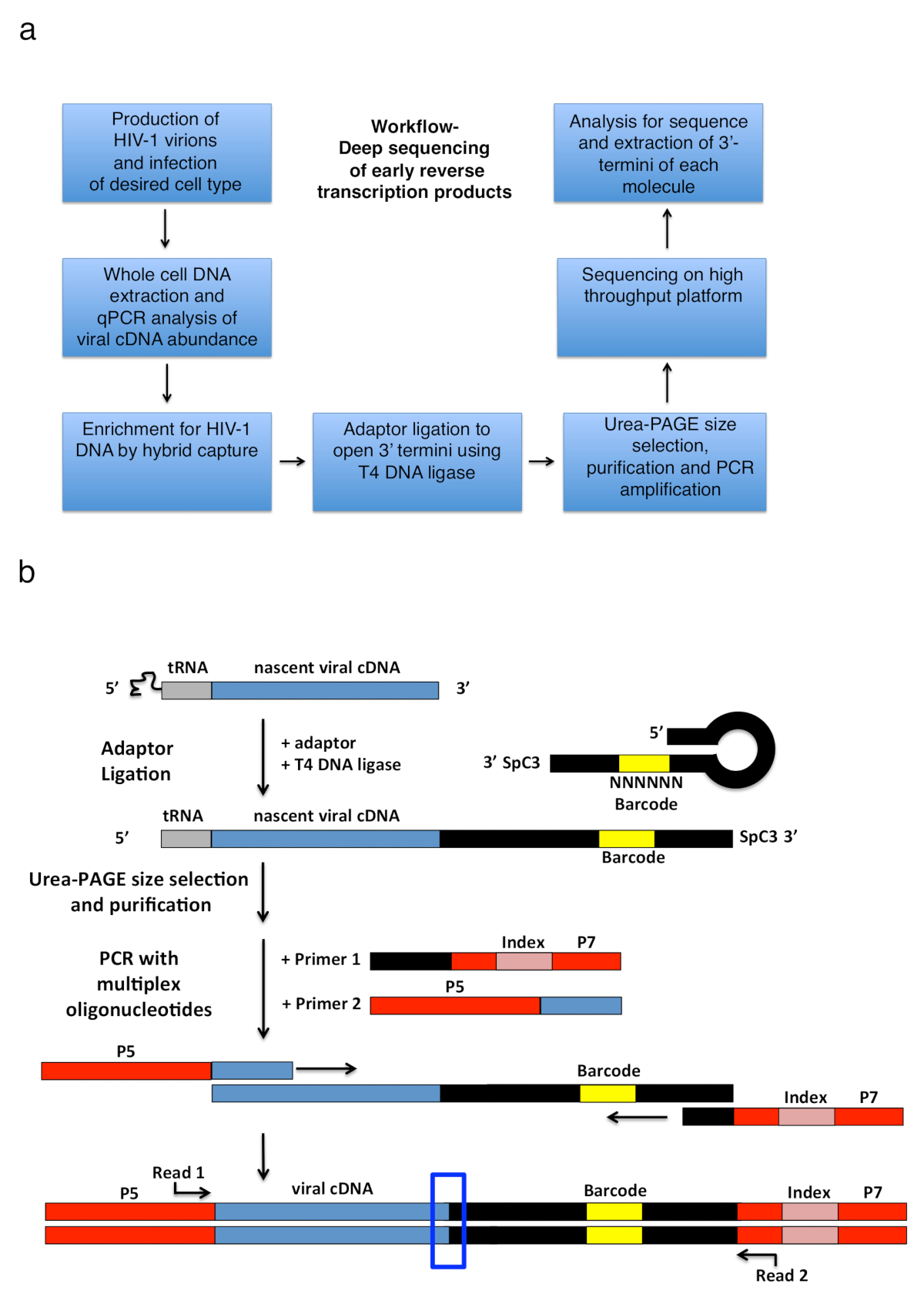{kind=link}

図 3: 代表結果。(a) 代表的なサンプルで説明されているプロトコル処理の合計読み取り数。これはその 3' 末端最初 635 内で HIV 1 分子のユニークな読み取りとして識別されたすべてのシーケンスが含まれていますマイナスの nt 鎖 cDNA (PPT、までは図 1を参照)。A3G cDNA 合成を阻害する、読み取りカウント総数が減少に対し、A3G を運んでいない HIV-1 感染は最高数のリードを生成します。総合的なオリゴヌクレオチドのセットは、肯定的な制御を提供しながら、感染していない細胞はネガティブ コントロールとして提供しています。b) nt 位置 23 と 182 の間長ごとの Cdna の相対的な豊かさ (sss 完全長 cDNA は 180 に 182 nt) HIV-1NL4.3のシーケンス (x 軸) が青のヒストグラム (左側の y 軸のスケール) で表示されます。CDNA の相対的な豊かさは、シーケンス 182nt の測定のすべての読み取りの合計で割った値 sss cDNA シーケンス内で特定の塩基配列の終端または以下の絶対数から算出しました。(-右の y 軸のスケール) のそれぞれの位置での C-T/U の突然変異を運ぶ読み取りのパーセンテージは、赤の破線で表示。図 3bは、以前文書6から再発行されます。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 4: コントロールのサンプルの代表の結果。17 の長さの異なる合成オリゴヌクレオチドの等モル量を含むプールの 2 つのプロファイルを示します。これらのオリゴヌクレオチド HIV-1NL4.3からのシーケンスがあるし、様々 な距離をカバーし、3'-ヌクレオチドとしてすべての 4 拠点が存在した (表 2参照)。上のグラフは、図 3から肯定的な制御のサンプルを示しています。分子の長さや、オープン 3' 末端に向かって重要なバイアスは検出されません。下のグラフは、実行すると、シーケンスでマイナーな長さバイアスを生成する別のライブラリを示しています。この場合、サイズのバイアスを表す (ピンクで表示) 斜面に由来する正規化係数を適用することをお勧めします。この図は、以前文書6から再発行されます。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}
| オリゴ名 | Nt の長さ | シーケンス | 目的 | メーカー (浄化) | ||||||||||||||
| フルの國 + MiSeq | 61 | 5'-フォー-tgaagagcctagtcgctgttcannnnnnctgcccatagagagatcggaagagcacacgtct-SpC3-3' | アダプター | IDT DNA 技術 (HPLC) | ||||||||||||||
| 2xBiotin SS 餌 | 40 | 5'-ビオチン-cagtgtggaaaatctctagcagtggcgcccgaacagggac-ビオチン-3' | ハイブリッド キャプチャ | MWG Eurofins (HPLC) | ||||||||||||||
| ビオチン 1 16 ss | 22 | 5'-cagtgtggaaaatctctagcag-顕微鏡-3' | ハイブリッド キャプチャ | MWG Eurofins (高速液体クロマトグラフィー | ||||||||||||||
| ビオチン tRNA + CTG | 16 | 5'-cagtggcgcccgaaca-顕微鏡-3' | ハイブリッド キャプチャ | MWG Eurofins (HPLC) | ||||||||||||||
| MP1.0 + 22HIV | 82 | 5'-aatgatacggcgaccaccgagatctacactctttccctacacgacgctcttccgatctcactgctagagattttccacactg-3' | PCR の拡大 | MWG Eurofins (高速液体クロマトグラフィー | ||||||||||||||
テーブル 1: テーブル記述プロトコルの長さ、シーケンス、および使用されている修正を含むオリゴヌクレオチドの。テーブルは、以前の文書6から適応。Excel ファイルとしてこの表をダウンロードするここをクリックしてください。
| オリゴ名 | Nt の長さ | シーケンス | メーカー (浄化) | |||||||||||||
| HTP con 長い C | 120 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcacacactactttgagcactcaaggcaagctttattgaggcttaagc-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP con 長い G | 119 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcacacactactttgagcactcaaggcaagctttattgaggcttaag-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP con 長い T | 116 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcacacactactttgagcactcaaggcaagctttattgaggctt-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP コン ロング A | 118 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcacacactactttgagcactcaaggcaagctttattgaggcttaa-3' | MWG Eurofins (HPLC) | |||||||||||||
| C 半ば HTP コン | 76 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcac-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP con 半ば G (、) | 71 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacagacg-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP con 半ば G (b) | 72 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacagacgg-3' | MWG Eurofins (HPLC) | |||||||||||||
| A 半ば HTP コン | 69 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacaga-3' | MWG Eurofins (HPLC) | |||||||||||||
| T 半ば HTP コン | 85 | 5'-ctgctagagattttccacactgactaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcacacactactt-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP コン ショート A | 40 | 5'-ctgctagagattttccacactgactaaaagggtctgaggga-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP con ショート T | 33 | 5'-ctgctagagattttccacactgactaaaagggt-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP con 短い G | 41 | 5'-ctgctagagattttccacactgactaaaagggtctgaggg-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP con 短い C | 34 | 5'-ctgctagagattttccacactgactaaaagggtc-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP コン 46 (T) | 46 | 5'-ctgctagagattttccacactg actaaaagggtctgagggatctct-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP Con 83 (C) | 83 | 5'-ctgctagagattttccacactg actaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcacacactac-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP Con 103 (C) | 103 | 5'-ctgctagagattttccacactg actaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcacacactactttgagcactcaaggcaagc-3' | MWG Eurofins (HPLC) | |||||||||||||
| HTP コン 107 (A) | 107 | 5'-ctgctagagattttccacactg actaaaagggtctgagggatctctagttaccagagtcacacaacagacgggcacacactactttgagcactcaaggcaagcttta-3' | MWG Eurofins (HPLC) | |||||||||||||
表 2: 肯定的な制御サンプルとして使用される 17 の合成制御オリゴヌクレオチドのテーブル。トップ 13 オリゴヌクレオチドは、サイズに基づいて選ばれた [長い (116 を 120 nt)、中期 (69 に 85 nt)、短い (33 に 41 nt)] その 3' 末端だけでなく。テーブルは、以前の文書6から適応。 Excel ファイルとしてこの表をダウンロードするここをクリックしてください。
ディスカッション
高速、信頼性、およびコスト効率の高いディープ シーケンスのシーケンスに基づく分析の偉大な深さを許可する生命科学の分野での多くの側面をもたらしました。残りの課題は、革新的なデザイン、代表者の作成シーケンス ライブラリをあります。ここで初期のウイルス cDNA 分子、特に HIV-1 の逆のトランスクリプション プロセスの中間体をキャプチャするプロトコルについて述べる。
この戦略の最も重要なステップは、定量的かつ公平な方法、オープン 3'-テルミニ駅までのアダプターの ligation。両方の間の 2 つの ssDNA テルミニ間および効率、- と分子内、調査し、様々 なアプリケーション11,26,27,28,29用に最適化されています。手順 3.3 で説明した条件の下で T4 DNA リガーゼとヘアピンのアダプターを使用しての選択は異なるリガーゼ、アダプターとを表す総合的なオリゴヌクレオチド ligation 用試薬を行った実証の最適化の結果HIV-1 シーケンス (データは示されていない) (表 2)。生体外のテスト、これらの反応を確認した T4 DNA リガーゼがヘアピンのアダプターの ligation を介した國らによる記述で.11、非常に低いバイアスがあり過剰でアダプターを使用する場合、アクセプター分子の完全な結紮近く実現します。ライゲーション効率が多重プライマー システムの互換性のあるアダプターをレンダリングする塩基の添加による影響を受ける (図 4を参照)。比較では、ことがわかった、耐熱性 5' DNA ・ RNA リガーゼ (「リガーゼ」、参照テーブルの材料に比べて正確なリガーゼの)、は、設計された RNA リガーゼの一部開発された受容体とライゲーション効率を改善するには27、RNA リガーゼ (「リガーゼ B」) がある重要なバイアスは、オリゴヌクレオチドを単一塩基長の違い [表 2 間もライゲーション効率の強い相違よりも、2 つの一本鎖 Dna 分子を縛ることで確かに効果的であった;G (a) と (b) 半ば詐欺 HTP]。さらに、"リガーゼ C"、無作為に選ばれた 5' 末端を運ぶアダプター併用で最小限のバイアスのみで反応である (戦略"リガーゼ C"の知られているヌクレオチド バイアスをオフセットするために使用; たとえば鼎らを参照してください。30). ただし、"リガーゼ C"-仲介された分子間を完了、T4 DNA リガーゼ システムに優れた選択をレンダリングしていなかった。
プロトコルのコースと正と負のコントロールを含めることにいくつかの品質管理手順アッセイ継続する前に潜在的な問題の検出および努力のトラブルシューティングのためのガイダンスを提供できます。QPCR 数量 2.2.2 と 2.3.12 の手順では、入力素材の量が十分であることを確認します。約 10,000 に 30万個 μ L から 200 μ L (ステップ 2.1) からの溶出範囲で典型的な cDNA コピー数。ハイブリッド キャプチャ ステップことができます全体的な HIV-1 のいくつかが失われる cDNA 数量 HIV 1 cDNA の強い濃縮によって濃縮の前後にゲノム DNA を定量化する適切なプライマーを使用して定めることができる細胞の DNA にありますが、qPCR または DNA 濃度の測定によって。回復の HIV-1 cDNA ハイブリッド キャプチャ手順を実行した後は、入力の少なくとも 10% をする必要があります。開始材料低可能性があります成功したオリゴヌクレオチド肯定的な制御を説明するそれ以外の場合 (手順 3.3.2 参照) しますが、限られたサンプルでの読み取り。低は、MiSeq アダプターなし無関係 DNA 種の存在のためのライブラリ濃度の過大評価で説明できるは、全体の数字もお読みください。これは低クラスター密度になるし、蛍光アッセイによって合計の DNA 量に加えて qPCR によるライブラリ内の HIV-1 シーケンスの濃度を決定することで改善できます。メソッドの非常に敏感な性質のため特別な注意が必要 (特に、高濃度コントロール オリゴヌクレオチド株式) からでなく、実験装置からの他のサンプルの両方からも低レベルの汚染を避けるために。PCR のワークステーションを殺菌紫外線での作業この点で有益です。最後のライブラリ (ステップ 6.1.2) の自動ゲル電気泳動は、さらに品質管理指標です。通常観察核酸サイズ範囲は 150 に 500 nt プライマー PCR 後浄化 (ステップ 5.2 の注を参照してください) 今欠席すべき前に、オプションのコントロールで検出可能です。代表的な結果でサンプル強度曲線のピークは約 160 に 170 nt と 2 番目のシャープ ピーク約 320 に 350 nt。これはおそらく比較的短い (1 に 20 nt 挿入長さ) 逆のコピーとフルレングスの強力な停止 (182 に 180 nt 挿入長さ) の両方でよく見かける高い豊かさを反映している (図 3b)。
提案するプロトコルと選択したプライマーが早期の HIV-1 逆のトランスクリプションの構成要素の特定、メソッドはオープン 3' 末端の DNA を決定することを目指して研究に一般的に適用されます。他のコンテキストに必要な主な変更は、ハイブリッド キャプチャ ・ プライマー デザイン戦略メソッドになります。たとえば、ターゲットを後半の HIV-1 の成績証明書に適応する場合は、多数の異なるキャプチャ ビオチン標識オリゴヌクレオチド Dna の長さにわたって焼鈍勧められる、ハイブリッド キャプチャ手順で損失を減少する可能性が。導入で述べたように、その 3' 末端がさまざまなソース バイアスを避けるために検出する範囲を設計するときの制限を考慮することが重要です。最初に場合、可能性がありますバイアス PCR の反作用でアダプターのテンプレートは、非常に様々 な長さの。第二に、(例えばMiSeq) シーケンス プラットフォーム使用ここでは、最適なクラスタ リングの最寄りの挿入長さの範囲と短く、長い製品が同じ効率でのシーケンスを処理しない可能性があります大幅。一部では、これは計算上、長さバイアスの補正係数を計算することによって行われていた対処できる (図 4下のグラフを参照してください)。ただしの 3' 末端のマッピングが必要な領域が長い場合 (> 1000 nt)、それはより結紮の転写産物が関与する反応を分割し、3' 末端のセクションでを評価するために複数の上流のプライマーを使用することをお勧めします。
解析プログラムは、任意の突然変異を識別するために全ての基地の基本バリエーションだけでなく、固定アダプター シーケンスに隣接する HIV-1 シーケンスの両方の最後のヌクレオチドの分析の特定の目的のため社内に書かれていた。個々 のステップは、以下を構成する: まず、アダプター シーケンスは fastx 0.0.13 ツールキット; を使用してトリムされますその後、(バーコードを含む同一のシーケンスを意味する) 重複したシーケンスが削除されます。残りのすべてのユニークな読み取りは、ボウタイ (http://bowtie-bio.sourceforge.net/index.shtml) を用いた 3 つの拠点に設定最大の不一致 HIV-1 シーケンスに配置されます。テンプレートのシーケンスで構成されています最初の 635 - sss シーケンスと polypurine トラック (U5 R U3 PPT; を参照してください図 1) までの最初の鎖転送製品を含む HIV 1 cDNA (NL4.3 株) の nt。これにより、指定されたソフトウェアとテンプレートも、直接適切なメソッドを使用して同じアプリケーション (HIV-1NL4.3の初期の逆のコピーの検出) の場合だけです。調整は、他のターゲット シーケンスのために作られる必要があります。各 read に対して 3' テルミニの位置は、配置位置によって決定されました。各位置のためのデータベース呼び出しが記録され、突然変異率は異なりますが、さまざまな長さの読み取りがあり、Read2 の 125 ベース シーケンスによって長い挿入は全く扱われない各拠点の全体のカバレッジから計算されます。
最後に、研究の多くの種類のための貴重なツールであることこの方法と考えています。明白なアプリケーションには、抗レトロ ウイルス薬や携帯電話の制限要因を逆転写抑制機構の調査が含まれます。ただし、唯一の比較的マイナーな調整は 3' 末端他一本鎖 DNA ウイルス中間体、パルボ ウイルス複製にたとえば、ある内でマッピングするシステムを適応する必要する必要があります。さらに、法、特にその最適化された結紮のステップの原理が伸び触媒による細胞の二本鎖 DNA を含むすべての 3' の DNA の拡張の特性のライブラリ作成デザインのコアな部分を提供できます。ポリメラーゼ。
開示事項
著者は、開示するものがあることを宣言します。
謝辞
著者・ マリム研究所、ルイス アポロニア、Jernej Ule とレベッカ Oakey のメンバーのサポートを認めます。著者に感謝キングス カレッジ ロンドンのゲノム センターでマット アルノとデビー ヒューズ大学大学ロンドン (UCL) で、研究所神経次世代シーケンス処理施設は、MiSeq シーケンスのヘルプが実行されます。付与契約の下での作業、英国医学研究評議会 (G1000196 ・氏/M001199/1 MM に)、Wellcome の信頼 (106223/Z、14、Z MM の ~)、欧州委員会の第 7 フレームワーク プログラム (FP7/2007-2013) 支えられないです。(懲戒パネル) に PIIF-ジョージア州 2012-329679 と、保健省経由で男のと聖トマス NHS 財団保健管理センター総合医研究賞、国立衛生研究所キングス ・ カレッジ ・ ロンドンとキングとの提携で信頼大学病院 NHS の基礎信頼。
資料
| Name | Company | Catalog Number | Comments |
| 293T cells | ATCC | CRL-3216 | |
| Dulbecco's Modified Eagle's Medium | Gibco | 31966-021 | |
| Penicillin/Streptomycin | Gibco | 15150-122 | |
| Fetal Bovine Serum | Gibco | 10270-106 | |
| HeraCell Vios 250i CO2 Incubator | Thermo Scientific | 51030966 | |
| Laminar flow hood - CAS BioMAT2 | Wolflabs | CAS001-C2R-1800 | |
| 10mm TC-treated culture dish | Corning | 430167 | |
| TrypLE™ Express (1x), Stable Trypsin Replacement Enzyme | Gibco | 12605-010 | |
| OptiMEM® (Minimal Essential Medium) | Gibco | 31985-047 | |
| HIV-1 NL4-3 Infectious Molecular Clone (pNL4-3) | NIH Aids reagent program | 114 | |
| Polyethylenimine (PEI) - MW:25000 | PolySciences Inc | 23966-2 | dissolved at 1mg/ml and adjusted to pH7 |
| RQ1- Rnase free Dnase | Promega | M6101 | |
| Filter 0.22 μm | Triple Red Limited | FPE404025 | |
| 15 mL polypropylene tubes | Corning | CLS430791 | |
| Sucrose | Calbiochem | 573113 | |
| Phosphate Buffered Saline (1x) | Gibco | 14190-094 | |
| Ultracentrifuge tubes | Beckman Coulter | 344060 | |
| Ultracentrifuge | Sorval | WX Ultra Series | Th-641 Rotor |
| Alliance HIV-1 p24 antigen ELISA kit | Perkin Elmer | NEK050001KT | |
| CEM-SS cells | NIH Aids reagent program | 776 | |
| Roswell Park Memorial Institute Medium | Gibco | 31870-025 | |
| CoStar® TC treated multiple well plates | Corning | CLS3513-50EA | |
| Benchtop centrifuge: Heraus™ Multifuge™ X3 FR | Thermo Scientific | 75004536 | |
| TX-1000 Swinging Bucket Rotor | Thermo Scientific | 75003017 | |
| Microcentrifuge: 5424R | Eppendorf | 5404000060 | |
| Total DNA extraction kit (DNeasy Blood and Tissue kit) | Qiagen | 69504 | |
| Nuclease free H2O | Ambion | AM9937 | |
| Cutsmart buffer | New England Biolabs (part of DpnI enzyme) | R0176S | |
| DpnI restriction enzyme | New England Biolabs | R0176S | |
| Oligonucleotides for qPCR | MWG Eurofins | N/A | HPSF purification |
| TaqMan PCR Universal Mastermix | Thermo | 4304437 | |
| LoBind Eppendorf® tubes | Eppendorf | 30108078 | |
| Axygen™ aerosol filter pipette tips, 1000 μL | Fisher Scientific | TF-000-R-S | |
| Axygen™ aerosol filter pipette tips, 200 μL | Fisher Scientific | TF-200-R-S | |
| Axygen™ aerosol filter pipette tips, 20 μL | Fisher Scientific | TF-20-R-S | |
| Axygen™ aerosol filter pipette tips, 10 μL | Fisher Scientific | TF-10-R-S | |
| PCR clean hood | LabCaire | Model PCR-62 | |
| DynaMag™2-magnet | Thermo | 12321D | |
| Streptavidin MagneSphere® paramagnetic particles | Promega | Z5481 | |
| Casein | Thermo Scientific | 37582 | |
| End over end rotator, Revolver™ 360° | Labnet | H5600 | |
| Tris-Base | Fisher Scientific | BP152-5 | |
| Hydrochloric Acid | Sigma | H1758-100ML | |
| EDTA disodium salt dihydrate | Electran (VWR) | 443885J | |
| Sodium Chloride | Sigma | S3014 | |
| Dri-Block® Analog Block Heater | Techne | UY-36620-13 | |
| PCR tubes and domed caps | Thermo Scientific | AB0266 | |
| PCR machine | Eppendorf | Mastercycler® series | |
| T4 DNA ligase | New England Biolabs | M0202M | |
| 40% Polyethylene glycol solution (PEG) in H2O, MW: 8000 | Sigma | P1458-25ML | |
| Betaine solution, 5M | Sigma | B0300-1VL | |
| Gel loading buffer II (formamide buffer) | Thermo Scientific | AM8546G | |
| Precast 6% TBE urea gels | Invitrogen | EC6865BOX | |
| Mini cell electrophoresis system | Invitrogen, Novex | XCell SureLock™ | |
| Tris/Borate/EDTA solution (10x) | Fisher Scientific | 10031223 | |
| Needle 21 G x1 1/2 | VWR | 613-2022 | |
| SYBR Gold nucleic acid stain (10000x) | Life Technologies | S11494 | |
| Dark Reader DR46B transilluminator | Fisher Scientific | NC9800797 | |
| Ammonium acetate | Merck | 101116 | |
| SDS solution 20% (w/v) | Biorad | 161-0418 | |
| Centrifuge tube filter | Appleton Woods | BC591 | |
| Filter Glass Fibre Gf/D 10mm | Whatman (VWR) | 512-0427 | |
| polyadenylic acid (polyA) RNA | Sigma | 10108626001 | |
| Glycogen, molecular biology grade | Thermo Scientific | R0561 | |
| Isopropanol (2-propanol) | Fisher Scientific | 15809665 | |
| Ethanol, molecular biology grade | Fisher Scientific | 10041814 | |
| Accuprime™ Supermix I (DNA polymerase premix) | Life Technologies | 12342-010 | |
| NEBNext® Multiplex Oligo for Illumina (Index Primer Set 1 and 2) | New England Biolabs | E7335S; E7500S | |
| Tapestation D1000 Screentape High sensitivity | Agilent Technologies | 5067- 5584 | |
| Tapestation D1000 Reagents | Agilent Technologies | 5067- 5585 | |
| 2200 Tapestation - automated gel electrophoresis system | Agilent Technologies | G2965AA | |
| Agencourt® AMPure® beads XP | Beckman Coulter | A63880 | |
| Qubit™ dsDNA HS Assay Kit | Invitrogen | Q32851 | |
| Qubit™ 2.0 Fluorometer | Invitrogen | Q32866 | |
| Topo™ TA cloning Kit | Invitrogen | 450071 | |
| Sequencing platform: MiSeq System | Illumina | ||
| Experiment Manager (Sample sheet software) | Illumina | Note: Use TruSeq LT as a template | |
| Miseq™ Reagent kit V3 (150 cycle) | Illumina | MS-102-3001 | |
| Sequencing hub: Basespace | Illumina | https://basespace.illumina.com | |
| Ligase A: Thermostable 5’ App DNA/RNA ligase | NEB | M0319S | Not used in this protocol, but tested in optimization process with results described in the discussion. |
| Ligase B: T4 RNA ligase 1 | NEB | M0204 | Not used in this protocol, but tested in optimization process with results described in the discussion. |
| Ligase C: CircLigase | Epicentre | CL4111K | Not used in this protocol, but tested in optimization process with results described in the discussion. |
参考文献
- Herschhorn, A., Hizi, A. Retroviral reverse transcriptases. Cellular and Molecular Life Sciences. 67 (16), 2717-2747 (2010).
- Hu, W. S., Hughes, S. H. HIV-1 reverse transcription. Cold Spring Harbor Perspectives in Medicine. 2 (10), (2012).
- Levin, J. G., Mitra, M., Mascarenhas, A., Musier-Forsyth, K. Role of HIV-1 nucleocapsid protein in HIV-1 reverse transcription. RNA Biology. 7 (6), 754-774 (2010).
- Menendez-Arias, L., Sebastian-Martin, A., Alvarez, M. Viral reverse transcriptases. Virus Research. , (2016).
- Telesnitsky, A., Goff, S. P., Coffin, J. M., Hughes, S. H., Varmus, H. E. . Retroviruses. , (1997).
- Pollpeter, D., et al. Deep sequencing of HIV-1 reverse transcripts reveals the multifaceted antiviral functions of APOBEC3G. Nature Microbiology. 3 (2), 220-233 (2018).
- Frohman, M. A., Dush, M. K., Martin, G. R. Rapid production of full-length cDNAs from rare transcripts: amplification using a single gene-specific oligonucleotide primer. Proceedings of the National Academy of Sciences of the United States of America. 85 (23), 8998-9002 (1988).
- Liu, X., Gorovsky, M. A. Mapping the 5' and 3' ends of Tetrahymena thermophila mRNAs using RNA ligase mediated amplification of cDNA ends (RLM-RACE). Nucleic Acids Research. 21 (21), 4954-4960 (1993).
- Ince, I. A., Ozcan, K., Vlak, J. M., van Oers, M. M. Temporal classification and mapping of non-polyadenylated transcripts of an invertebrate iridovirus. Journal of General Virology. 94, 187-192 (2013).
- Hafner, M., et al. RNA-ligase-dependent biases in miRNA representation in deep-sequenced small RNA cDNA libraries. RNA. 17 (9), 1697-1712 (2011).
- Kwok, C. K., Ding, Y., Sherlock, M. E., Assmann, S. M., Bevilacqua, P. C. A hybridization-based approach for quantitative and low-bias single-stranded DNA ligation. Analytical Biochemistry. 435 (2), 181-186 (2013).
- Abram, M. E., Tsiang, M., White, K. L., Callebaut, C., Miller, M. D. A cell-based strategy to assess intrinsic inhibition efficiencies of HIV-1 reverse transcriptase inhibitors. Antimicrobial Agents and Chemotherapy. 59 (2), 838-848 (2015).
- Bishop, K. N., Verma, M., Kim, E. Y., Wolinsky, S. M., Malim, M. H. APOBEC3G inhibits elongation of HIV-1 reverse transcripts. PLoS Pathogens. 4 (12), 1000231 (2008).
- Zack, J. A., Haislip, A. M., Krogstad, P., Chen, I. S. Incompletely reverse-transcribed human immunodeficiency virus type 1 genomes in quiescent cells can function as intermediates in the retroviral life cycle. Journal of Virology. 66 (3), 1717-1725 (1992).
- Adachi, A., et al. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. Journal of Virology. 59 (2), 284-291 (1986).
- Shah, V. B., Aiken, C. In vitro uncoating of HIV-1 cores. Journal of Visualized Experiments. (57), (2011).
- JoVE Science Education Database. Science Education Database: Basic Methods in Cellular and Molecular Biology: Passaging Cells. Journal of Visualized Experiments. , (2018).
- JoVE Science Education Database. JoVE Science Education Database: Basic Methods in Cellular and Molecular Biology: Using a Hemocytometer to Count Cells. Journal of Visualized Experiments. , (2018).
- Zhou, M. Y., Gomez-Sanchez, C. E. Universal TA cloning. Current Issues in Molecular Biology. 2 (1), 1-7 (2000).
- Zhang, S., Cahalan, M. D. Purifying plasmid DNA from bacterial colonies using the QIAGEN Miniprep Kit. Journal of Visualized Experiments. (6), 247 (2007).
- Mangeat, B., et al. Broad antiretroviral defence by human APOBEC3G through lethal editing of nascent reverse transcripts. Nature. 424 (6944), 99-103 (2003).
- Gillick, K., et al. Suppression of HIV-1 infection by APOBEC3 proteins in primary human CD4(+) T cells is associated with inhibition of processive reverse transcription as well as excessive cytidine deamination. Journal of Virology. 87 (3), 1508-1517 (2013).
- Harris, R. S., et al. DNA deamination mediates innate immunity to retroviral infection. Cell. 113 (6), 803-809 (2003).
- Zhang, H., et al. The cytidine deaminase CEM15 induces hypermutation in newly synthesized HIV-1 DNA. Nature. 424 (6944), 94-98 (2003).
- Konig, J., et al. iCLIP--transcriptome-wide mapping of protein-RNA interactions with individual nucleotide resolution. Journal of Visualized Experiments. (50), (2011).
- Troutt, A. B., McHeyzer-Williams, M. G., Pulendran, B., Nossal, G. J. Ligation-anchored PCR: a simple amplification technique with single-sided specificity. Proceedings of the National Academy of Sciences of the United States of America. 89 (20), 9823-9825 (1992).
- Zhelkovsky, A. M., McReynolds, L. A. Structure-function analysis of Methanobacterium thermoautotrophicum RNA ligase - engineering a thermostable ATP independent enzyme. BMC Molecular Biology. 13 (24), (2012).
- Li, T. W., Weeks, K. M. Structure-independent and quantitative ligation of single-stranded DNA. Analytical Biochemistry. 349 (2), 242-246 (2006).
- Gansauge, M. T., et al. Single-stranded DNA library preparation from highly degraded DNA using T4 DNA ligase. Nucleic Acids Research. 45 (10), 79 (2017).
- Ding, Y., Kwok, C. K., Tang, Y., Bevilacqua, P. C., Assmann, S. M. Genome-wide profiling of in vivo RNA structure at single-nucleotide resolution using structure-seq. Nature Protocols. 10 (7), 1050-1066 (2015).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved