
Method Article
より厳格な特異性を有する制限エンドヌクレアーゼのインビトロ指向進化
要約
新しい配列特異性を有する制限エンドヌクレアーゼは、部分的に縮退配列を認識する酵素から開発することができる。ここでは、NlaIV酵素の配列特異性を変更するために使用した詳細なプロトコルを提供します。プロトコルの主な成分は、転写/翻訳反応のインビトロ区画化と新しい配列特異性を有するバリアントの選択である。
要約
制限エンドヌクレアーゼ(REase)特異度工学は極めて困難である。ここでは、親酵素よりも厳密な特異性を持つ REase バリアントを生成するのに役立つマルチステップ プロトコルについて説明します。このプロトコルでは、DNA結合に影響を及ぼす可能性のある位置の変動性を持つ、REaseのバリアントに対する発現選択カセット(ESC)のライブラリの作成が必要です。ESCは、所望の制限部位活性とビオチンタグの配列によって一方の側に並び、他方では望ましくない活性およびプライマーアニーリング部位に対する制限部位によって横たわっている。ESCは、液滴当たり複数のDNA分子が存在する可能性が低い条件で、油中水エマルジョンで転写および翻訳されます。したがって、各カセット分子中のDNAは、翻訳された、コードされた酵素の活性のみに供される。所望の特異性のREase変異体は、ビオチンタグを除去するが、プライマーアニール部位を除去しない。エマルジョンを破壊した後、DNA分子はビオチンプルダウンを受け、上清内のものだけが保持されます。このステップでは、目的のアクティビティを失っていないバリアントの ESC のみが保持されます。これらのDNA分子は、次いで、最初のPCR反応を受ける。望ましくない配列の切断は、プライマーの1つに対するプライマー結合部位を遮断する。したがって、PCRは望ましくない活性を持たない液滴からのESCのみを増幅する。次いで、第2のPCR反応を行い、所望の特異性およびビオチンタグに対する制限部位を再導入し、選択ステップを繰り返すことができるようにする。選択されたオープンリーディングフレームは、新たに進化したREaseがメチルトランスレーゼ標的部位のサブセットのみを標的とするため、親のREaseの同結合メチルトランスフェラーゼを発現する細菌細胞で過剰発現することができる。
概要
シーケンス特異性工学はクラスIIの安楽のための非常に挑戦的である。エンドヌクレアーゼのこのクラスでは、配列認識と触媒は密接に絡み合っており、おそらく宿主DNAに損傷を与える同卵性メチルトランスレセファーゼよりも広い特異性のエンドヌクレアーゼの作成に対する進化的保護手段として、細胞内の新しい特異性の指向進化は、新たに設計されたエンドヌクレアーゼ活性に対して宿主DNAを保護する必要性によってさらに複雑化する。したがって、報告されたREaseエンジニアリングの成功した試みはほんの少しあり、それらのすべては、特定の酵素11、2、3、4、5、6、72,3,4,5,6,7のユニークな機能を利用しています。
ここでは、NlaIVエンドヌクレアーゼ8の成功したエンジニアリングに基づく親酵素よりも狭い特異性を有するエンドヌクレアーゼ変異体を生成するために使用できる特異性工学のための詳細なプロトコルを提供する。任意の認識配列を有する任意の酵素に対して、側面の塩基に対して余分な特異性を導入することができる。部分的に縮退配列を認識する親酵素(GGNNCC標的を有するNlaIVなど)に対して、認識配列内に追加の特異性を導入することもできる。余分な特異性はタンパク質とDNAの接触を必要とする可能性が高いので、新たに認識された塩基はDNA上の親エンドヌクレアーゼの足跡内にあるべきである。原則として、選択スキームは、認識順序の任意の所望の特殊化のために設定することができます。しかし、パリンドロームおよびほぼパリンドロミック標的配列を認識するほとんどのREasesは、パリンドロームの半分の部位のみを認識する機能的なダイマーである。したがって、タンパク質核相互作用の対称性に違反する新しい特異性の選択は機能しそうにない。例えば、ダイマーNlaIVの場合、GGNNCC配列は理論的にはGGATCCに絞り込むことができますが、GGAACCに対する特異性の絞り込みはより困難であると予想されます。私たちのスキームは、正と負の両方の選択を伴います。
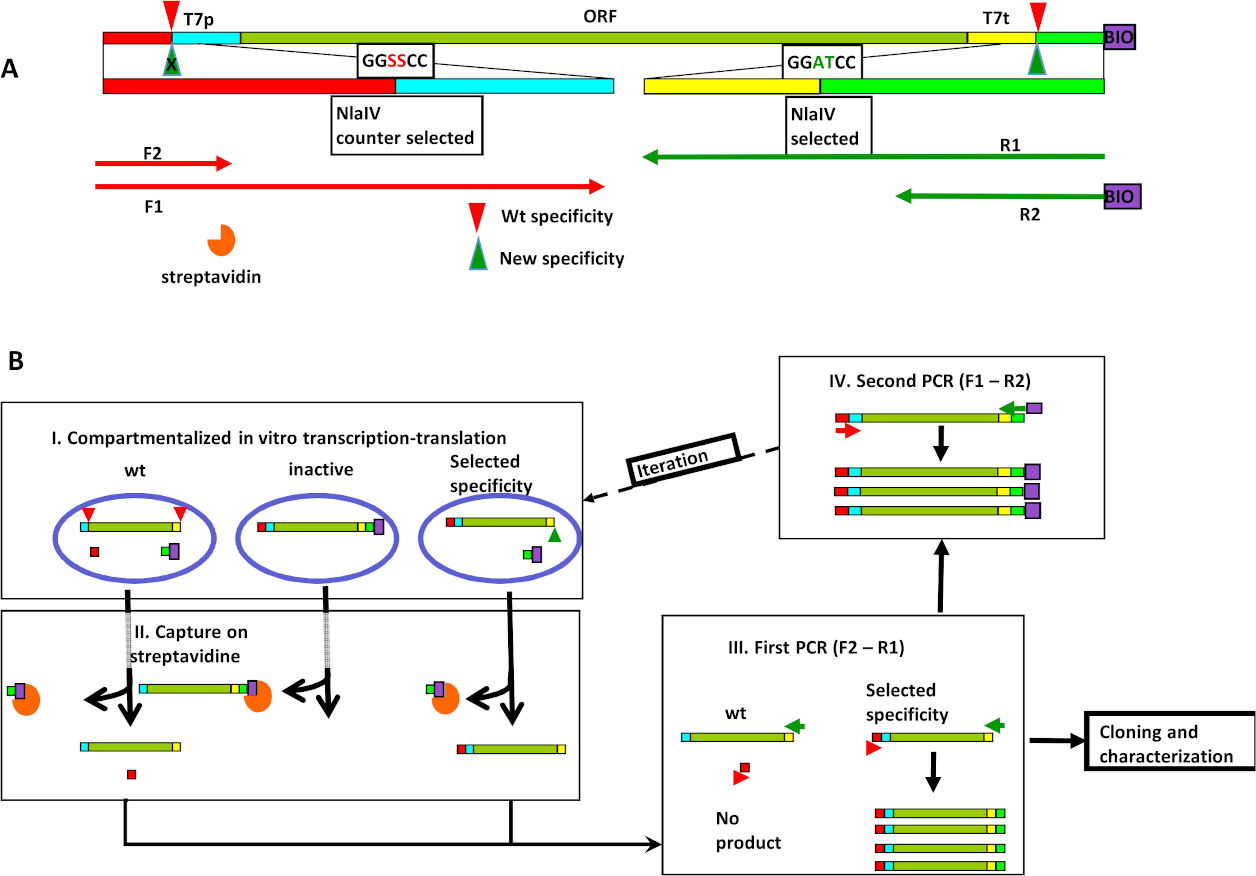
このプロセスは、ネガティブ選択が好ましい狭い特異性以外のすべての配列を切断することができる特異性を除去するためにも使用される場合に、より効率的である。例えば、GGATCC の選択は GGBVCC に対する選択の反対と組み合わせることができる (B は A 以外のベース、V は T 以外の任意のベース)。可能なターゲットシーケンスの一部がカバーされない場合、選択実験の結果は、正と負の選択の有効性に依存します。NlaIVの研究では、GGATCCに選択し、GGSSCC(SはGまたはCである)に対して選択し、対称破断ターゲットを無視してGGWWCC(WはAまたはT)と表現できる特異性を得たが、この特定のケースでは否定的な選択がより多かることを示唆している。肯定的な選択よりも重要です。
このアプローチは、式選択カセット (ESC) の作成から始まります。ESC はセクションで構成されています。内部コアセクションには、T7プロモーター制御下で、REaseのオープンリーディングフレーム(ORF)の変種があります。ESC のこのコアセクションには、設計された REase の同一のサイトを含めることはできません。コアは、野生型のREaseの2つの同卵部位の間に挟まれる:望ましくない活動のための切断部位(この例では選択された配列、GGSSCC)および所望の活動のための切断部位(選択された配列、GGATCC例)。PCRでESCを調製する最終ステップは、5'の終わりに所望の活性に近いビオチンを追加し、様々なカウンター選択配列(例ではGGSSCC)を作成します。選択戦略は、in vitro転写/翻訳/選択プロトコルの後にESC再増幅プロトコルで慎重に設計されたプライマーを使用することによるものである(図1A)。ESCライブラリは、インビトロ区画化転写変換水油エマルジョン99、10、1110,11で表現されています。各液滴内では、発現酵素の特異性はESCの状態に影響を及ぼす(図1B、ステップI)。記載された配列については、翻訳されたタンパク質の所望の切断活性は、DNAのビオチンタグを除去するが、カウンター選択配列を有する他のESC末端には影響を及ぼさない。エマルジョンが壊れると、ビオチン化フラグメントはストレプトアビジンアフィニティープルダウンによって除去され、所望の活性を有する液滴からの断片のみが残るようにする(図1B、ステップII)。この手順では、非アクティブな REase バリアントを削除します。プルダウンステップの上清分は、次いでPCRによって増幅されます。第1のPCR反応プライマーF2およびR1には、(図1A,B,ステップIII)が用いられる。プライマーF2は、選択されたカウンター配列と分子末端との間のESCセクションに結合する。したがって、選択されたカウンター配列を切断することができる変異体を発現するESC(したがって、プライマーF2とR1の結合部位を2つの異なるDNA分子に分離する)は増幅されず、したがってライブラリから除去される。プライマーR1は、選択された部位とESCのコアとの間に結合して、選択された部位の切断状態の影響を受けないようにし、所望の活性(GGATCC)の切断部位を復元する。サイクルは、選択した部位に近い5'末端にビオチンを追加し、ESCの反対側の端に近いカウンター選択部位で設計されたバリエーションを復元する第2のPCR(プライマーF1およびR2付き)によって閉じられる(図1B、ステップIV)。得られたDNA混合物は、別の選択ラウンドの準備ができています。
選択プロトコルの成功は、新しい、より厳格な標的認識シーケンスの適切な選択と変異生成戦略とその効果的な実装の慎重な設計に強く依存します。REaseのわずかな既存の好みを克服するよりも、はるかに簡単に改善できるため、既存の好みの運動学的研究から始めることをお勧めします。慎重な変異生成設計の必要性は、提示されたプロトコル(1回の実験で109クローン)で処理することができる変異型ライブラリの限られたサイズから生じる。したがって、20個のアミノ酸置換の可能性は、わずか数箇所で有効にテストすることができます(議論参照)。代わりの方法として提示されるエラーを起こしやすいPCR(EP-PCR)のようなランダムな変異誘発は、既存の複雑さの深いアンダーサンプリングにつながる。DNAとの接触に関与する潜在的なアミノ酸位置に関する情報(または同離配列中の縮退ヌクレオチドに近接している場合)が利用可能である場合、オリゴヌクレオチド誘導飽和変異体に対していくつかのアミノ酸を選択するために使用されるべきである(プロトコルステップ1.6-3.10)。
プロトコル
1. ESCの準備
- 低コピー数プラスミド(例えば、pACYC184またはpACYC174またはそれらの誘導体)で操作される制限修飾系のメチルトランスファーーゼのクローン。
注:細菌宿主株は、クローン酵素によって導入されたメチル化を許容し、T7 RNAポリメラーゼの誘導可能な発現を提供できる必要があります。ER2566株(McrA、McrBC、およびMrr突然変異を運ぶ)の使用が推奨されます。 - 組換えプラスミドDNAが、酵素供給業者が2時間推奨する10単位の共生ReEaseで0.5μgのプラスミドDNAを処理することにより、同結合エンドヌクレアーゼによる切断から保護されていることを確認します。
- この株の有能な細胞を準備します。
注:任意の方法を使用できます。NlaIVエンジニアリングプロジェクトは、単純な塩化カルシウム法12を使用しました。 - 異なる排除基から T7 プロモーターの制御下にある REase 用 ORF を用いて組換えプラスミドを構築し、ステップ 1.1 でメチルトラスフラーゼ遺伝子を含むものとは異なる選択マーカーを使用する。ベクトル pET28 および pET30 を使用できます。
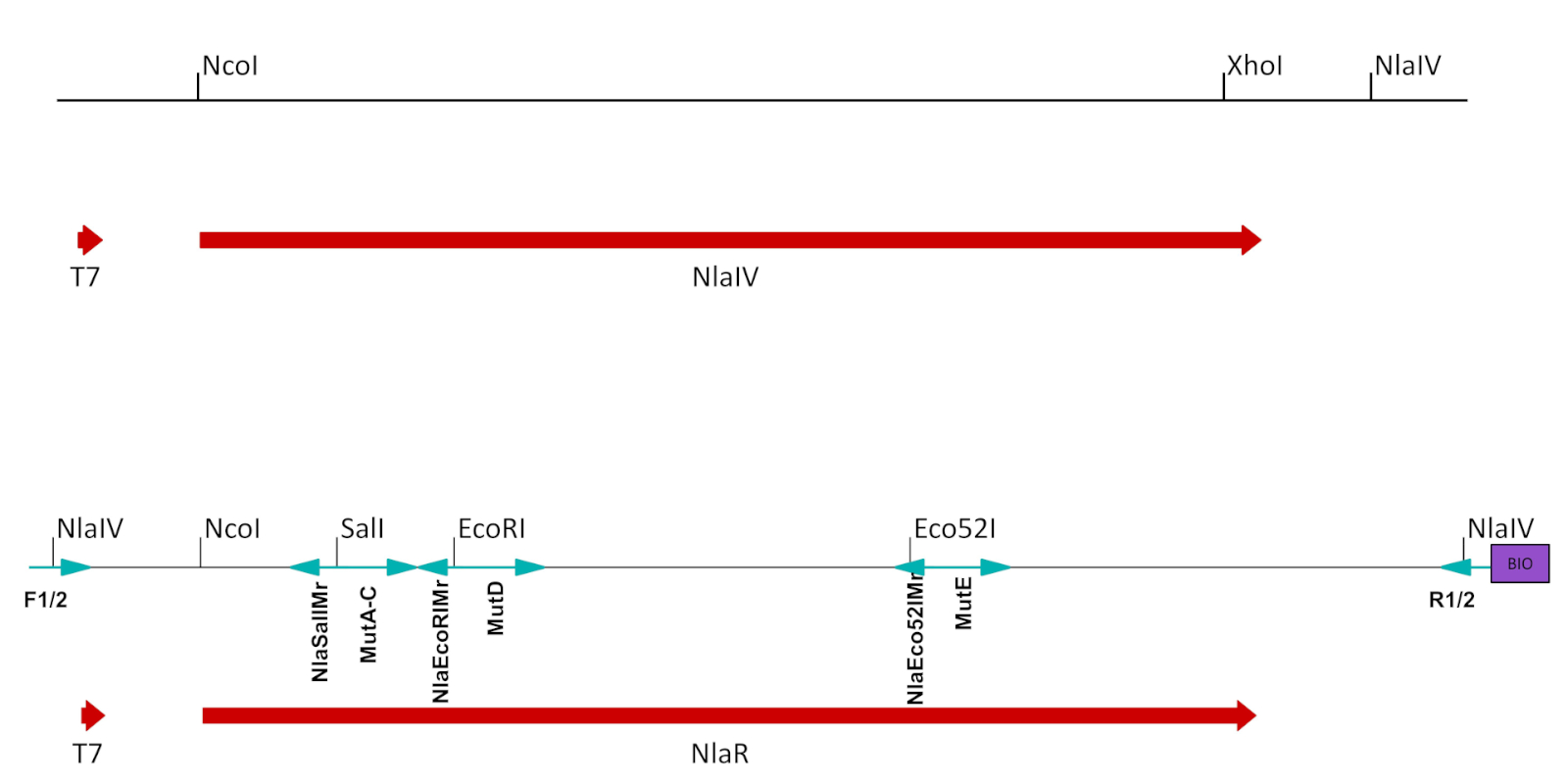
- T7プロモーターと酵素ORFの停止コドンとの間の組換えプラスミドのセクションから、無声変異を導入して、操作酵素の認識部位をすべて取り除く(図2、表1A)。
注:複数のそのようなサイトを削除する必要がある場合は、複数のミューテーションラウンドが必要です (ステップ 1.5.1 ~ 1.5.7)。- プライマーの5'の端部に導入された設計されたバリエーションで全長プラスミドを増幅するインサイドアウトPCR反応を使用する(表2A)。
- PCR反応の50 μLにDpnIエンドヌクレアーゼを10 U添加して鋳型DNAを除去し、37°Cで2時間インキュベートする。
- アガロースゲル電気泳動により製品を解決します。全長プラスミドに対応するバンドを切り取り、市販キットで精製します。
- 10xライゲーションバッファー(1倍の濃度)を加え、ATP(1 mMまで)で補います。T4ポリヌクレオチドキナーゼを10U添加し、37°Cで20分間インキュベートする。70°Cで10分間加熱して酵素を不活性化する。
- PEG 4000~5%を加え、ATP(1 mM)で再度補い、T4 DNAリガーゼを5 Uに加えます。室温(RT)で2時間インキュベートする。
- 同じメチルトランスファーーゼを運ぶ有能な細菌株に変身(ステップ1.1)。
- プラスミドDNAを小規模に分離し、ディデオキシシーケンシングによる配列変化の導入を確認する。
- オリゴヌクレオチド誘導突然変異誘発によって標的となる配列に近い固有の制限部位を導入する(図2、表1B)。各サイトの手順 1.5.1 ~ 1.5.7 に従います。
注:このステップは、標的性変異誘発が使用される場合にのみ実行されます。ランダム突然変異誘発を行う場合は、手順 2 ~ 3 をスキップし、代わりにセクション 3 に進みます。この例では、すべてのサイトがターゲット領域の上流に導入されましたが、下流にも導入できます。 - ESCの増幅用のプライマーを設計する(表1C)。
- 選択した認識部位(R1)とその短いバージョン(R2)を導入するエンドヌクレアスORFの逆プライマー結合を設計し、選択されたNlaIV配列の外側に結合し、5'末端にビオチンを含む(図1を参照)。
- T7プロモーターのESC上流に結合するフォワードプライマー(F1)を設計する。このプライマーはまた、元の認識シーケンスのカウンター選択されたバリアント(すなわち、選択された逆配列を除いて元の酵素によって認識される最大の配列変動)を導入する必要があります。
注:このプライマーの短いバージョン(F2)は、選択PCR(ステップ5.9)で後で使用される、副選択されたシーケンスに遠位のシーケンスをカバーする。
2. 変異原性プライマーのスプリットアンドミックス合成
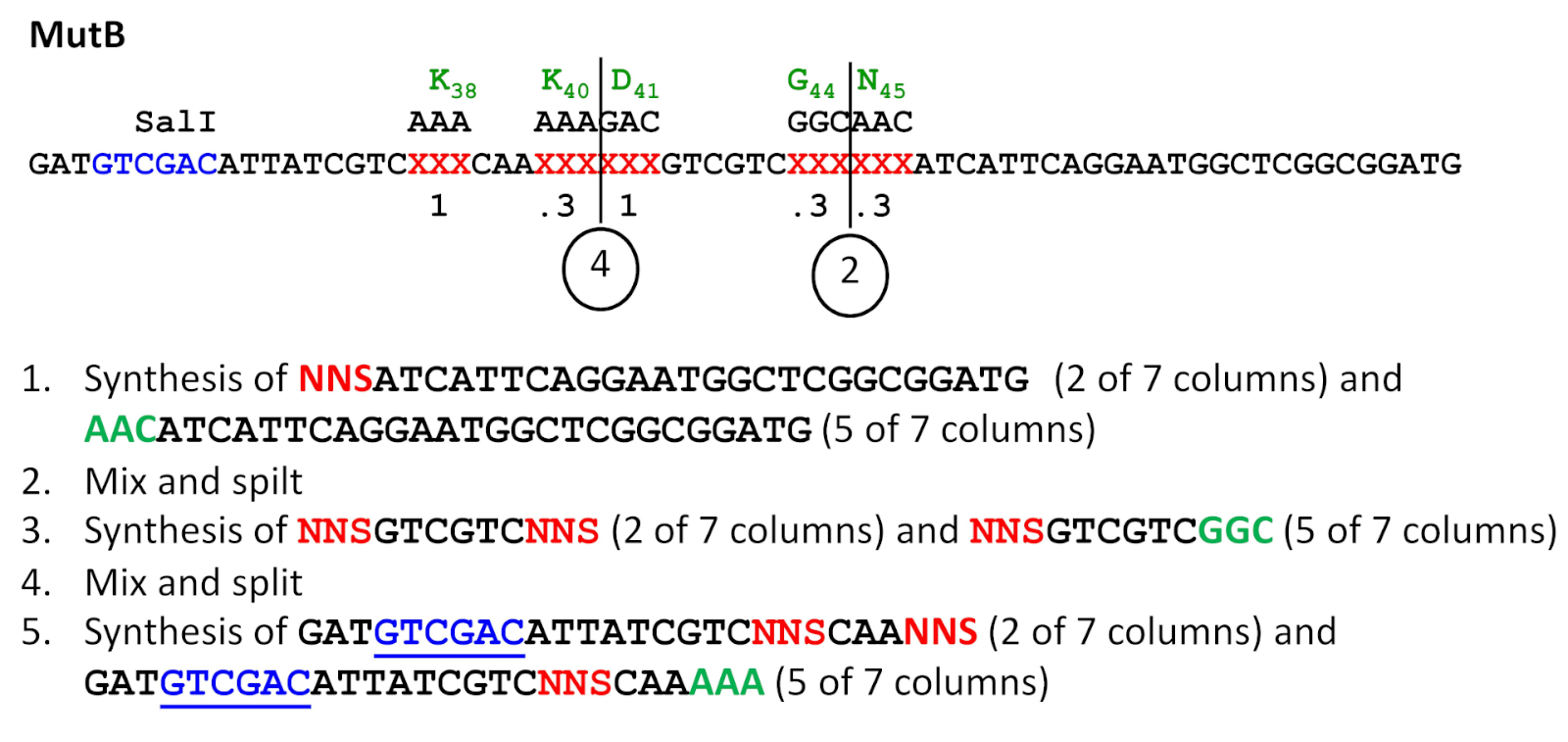
注:この手順は、複数のサイトで亜飽和変異生成を必要とするプロジェクトでのみ使用されます。複数の合成列を持つシンセサイザーが必要です。変異発生頻度に応じて、ランダム化NNSコドン三つ子と野生型コドン三つ子の合成のためのカラムを割り当てる。例えば、7つの等量合成カラムが利用可能で、特定の部位で0.3の変異生成率が望ましい場合、ランダム化NNSコドンを〜0.3 x 7または2列に、野生型コドンを〜0.7 x 7または5列に追加する(図3)。
- 亜飽和変異体の部位について決定します。.サイトの仮定の重要性に従って変異発生頻度を選択します(すなわち、サイトの重要性が高いほど頻度が高くなります)、ライブラリ全体の複雑さに関する制限を念頭に置いてください(説明を参照)。
- オリゴヌクレオチドを全てのカラムで合成し、3'末端から数える第2亜飽和変異形成部位の直前の三重項まで合成する。最後の合成サイクルで5'-トリチウム保護基を除去しないでください(シンセサイザーのトリチルオンオプションを使用してください)。保護基は、次の合成サイクルの開始時に除去されます (図 3のステップ 1)。
- 合成列を開きます。制御された細孔ガラス(CPG)合成サポートを乾燥した1.5 mLチューブに集め、渦を混ぜて混ぜます。混合CPG樹脂を新しい合成カラムに再パーティション化します。湿度の導入は、全体的な収量が低下するため避けてください(図 3のステップ 2 と 4)。
- 合成を継続し、亜飽和変異誘発部位トリプレットから始まる。カラムを、所望の突然変異発生頻度に従ってランダム化NNS三小または野生型三つ子に割り当てます(上記の注を参照)。追加の亜飽和部位が存在する場合は、次の亜飽和変異形成部位の前に三重項に進む。ここでも、5'-トリチウム基を最後に置いたままにします(シンセサイザーの5'-トリチルオンオプション)(図3のステップ3)。次に、ステップ 2.3 に進みます。
- 下流に亜飽和部位が存在しない場合は、合成を完了し、5'-トリチウム基を最後に残す(シンセサイザーの5'-トリチウムオンオプション)(図3のステップ5)。
- 精製カートリッジメーカーの指示に従ってオリゴヌクレオチドライブラリーを保護解除し、精製してください。
注:CPGから脱保護によって放出されるオリゴヌクレオチドは、逆相高速液体クロマトグラフィー(HPLC)でトリチウムイオン除去(RTで80%酢酸で1時間処理)および第2のHPLC精製を続けて精製することもできる。 - 尿素ページゲルのオリゴヌクレオチドライブラリーの品質を確認してください。
3. バリアントライブラリの生成
注:ステップ1.6の組換えプラスミドを使用してください。
-
オリゴヌクレオチド指向変異生成によりライブラリを生成する。
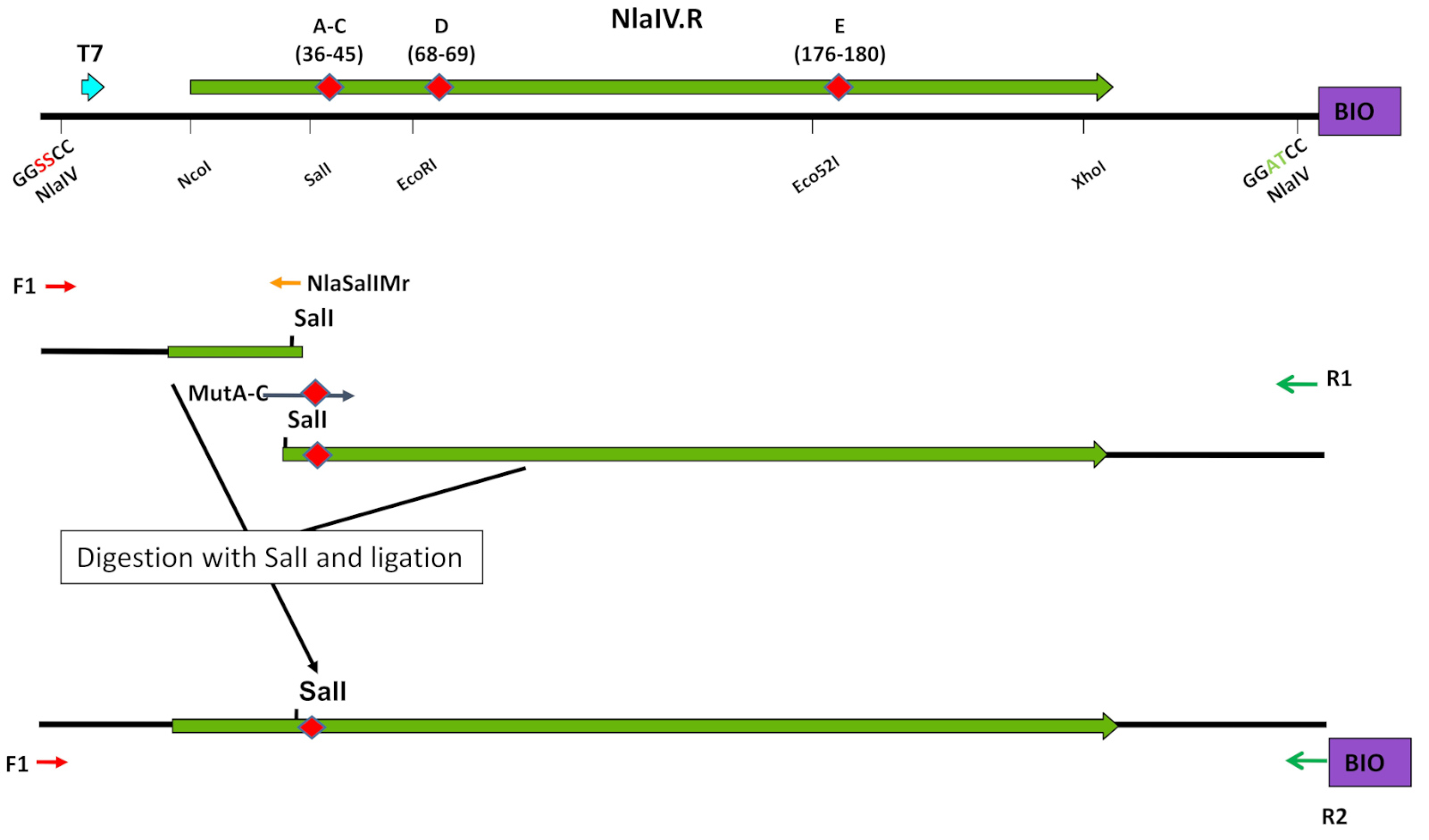
注: EP-PCR プロトコルを使用することもできます(ステップ 3.2)。- T7プロモーターから変異誘発を標的とする配列に隣接するユニークな制限酵素部位までのセクションを増幅する(NlaIVの場合:SalI、EcoRI、またはEco52I)(表1B-C、表2B、図4)。ユニークな制限酵素部位からESCの3'末端まで第2部を増幅します。
- 8 μLの水、10x制限酵素バッファーの1.5 μL、および適切な制限酵素(SalI、EcoRI、または Eco52I)の5単位を用いて、PCR反応を5 μL(ステップ3.1.1から)別々に混合し、適切な温度で2時間インキュベートします。
- アガロースゲル電気泳動を用いて、両反応の生成物を解決します。予想サイズのバンドを切り取り、市販キットで精製します。
- アガロースゲルで精製された製品の1/3まで実行し、濃度測定によって各精製バンドの濃度を測定します。
- 1xリガーゼバッファーとT4 DNAリガーゼの1 Uを用いた1:1モル比でESCの2つの部分のライゲーションを設定し、RTで2時間インキュベートします。
- アガロースゲル中の反応生成物を解決します。期待サイズの製品を切り取り、市販キットで浄化します。
- プライマーF1およびR2とのPCR反応で精製されたライゲーション生成物を増幅する(表1Cおよび表2A)。20以上の増幅サイクルを実行しないでください。
- アガロースゲル中のPCR反応を分画する。製品を切り取り、市販キットで浄化します。
- アガロースゲルの前のステップから精製されたライブラリーの5 μLアリコートを実行し、濃度測定法によって濃度を測定する。
- ライブラリの小さなサンプル(最大5μL)とシーケンス>15クローンをクローン化して、突然変異の頻度と分布を確認します(表3)。手順 4 に進みます。
注:あるいは、ESCの小サンプルのハイスループットシーケンシングを使用することもできます。
-
EP-PCR を実行します。
- ステップ1.5.7で得られたプラスミドからESCをプライマーF1およびR1で増幅する。Taq Iポリメラーゼ(表1B)で20サイクルを実行する。
- ゲルはPCR産物を精製する。
- 前のステップから精製 PCR 産物の 2 ng で EP-PCR を設定し、F1 および R1 プライマーを使用して 15 サイクルの EP-PCR (表 1C) を実行します。
- ゲルは製品を精製し、ゲル密度測定によって定量化します。
注:精製されたEP-PCR産物の低濃度のため、定量化に約1/3を使用した。 - ライブラリの小さなサンプル(最大1/5)とシーケンス>15クローンをクローン化して、突然変異の頻度と分布を確認します(表4)。
注:または、ESC の小さなサンプルのハイスループットシーケンスを実行します。
4. 区分化インビトロ転写翻訳反応の実施
-
インビトロ転写翻訳におけるエンドヌクレアーゼ発現および酵素活性を試験する。
- 分子の中心近くに位置するエンドヌクレアーゼの認識部位を1つ備えた短い(200~500 bp)基材を用意し、切断反応を容易に検出できるようにします。
注:基質を調製する最も簡単な方法は、任意のDNA分子の適切な断片のPCR増幅によってである。基質は、切断検出を単純化するために、放射標識または蛍光標識を行うことができる。 - メーカーの推奨に従って、0.5 μgの野生型ESCで50μLの転写変換反応を設定します。マグネシウム塩(MgCl2、MgSO4、および酢酸マグネシウムを試験可能)を1.5mMに加え、前工程から適切な量の基質(標識なしDNAの場合は0.5μg以上)を加えます。2
注:マグネシウムで活性化されたヌクレアーゼを含まない転写/翻訳キットを使用できます。一部のキットベンダーは、製造中のDNA汚染を除去し、ククレアーゼ阻害剤としてキレート剤を添加するためにヌクレアーゼを使用しています。このようなキットは、この方法と互換性がありません。 - 製造者の指示に従って転写翻訳反応をインキュベートします。その後、制限酵素に対する最適な温度に反応混合物を2時間移動させる。
- アガロースゲル中の基質の切断を分析し、その後適切な検出(例えば、DNA染色、蛍光ビジュアライゼーション、またはオートラジオグラフィー)を分析する(図5)。
注:区画化を進める前に、少なくとも基板の部分的な切断が必要である。これが達成されない場合は、マグネシウムの化学的形態またはその濃度のさらなる最適化が必要である。
- 分子の中心近くに位置するエンドヌクレアーゼの認識部位を1つ備えた短い(200~500 bp)基材を用意し、切断反応を容易に検出できるようにします。
- 15 mL円錐形チューブに225 μLのスパン80と25 μLのTween 80~5 mLの鉱油を加えて、油界面活性剤を調製します。管15倍を優しく反転して十分に混ぜます。
- 各ライブラリは、油界面活性剤混合物の950 μLを2mLの丸底極低温バイアルに移し、ライブラリ名でラベルを付け、氷に移します。小さな円筒形の攪拌棒(5 x 2 mm2)を各バイアルに入れます。
- メーカーの提案に従って、インビトロ転写変換反応混合物(ライブラリごとに50 μL)を用意します。塩化マグネシウムを加えた混合物を1.5 mMの最終濃度に補います(ステップ4.1.4を参照)。
- 氷上の1.5 mLチューブに50 μLのアリコートを分配します。
- 氷上の反応混合物にライブラリ(セクション3から)の1.7 fmoleを加えます。
注:選択効率を高くするために、より多くの式ライブラリを使用しないでください。複数のDNA分子を含む水性液滴の頻度を最小限に抑えることが重要です。 -
各ライブラリに連続して油中水エマルジョンを準備します。
- 1,150 rpmに設定された攪拌速度で、磁気スターラーに氷で満たされた小さなビーカー(または大きなボトルカップ)を置きます。
- 950 μLの油界面活性剤混合物と小さな攪拌棒を備えた極低温バイアルを、ステップ 4.3 から磁気攪拌機の氷冷ビーカーに移します。攪拌バーが回転していることを確認します。
- 30秒間隔で2分間にわたってin vitroライブラリ転写変換混合物の5つの10 μLアリコートを加え、さらに1分間攪拌を続けます。エマルジョンを使用してバイアルを氷容器に移します。ステップ 4.7.2 から始まる次のライブラリに進みます。
- すべてのライブラリが処理された後、キットメーカーの推奨事項に従って、すべてのライブラリのインキュベーションを開始します。
- バイアルを、さらに2時間、設計されたエンドヌクレアーゼに最適な温度に移し、少なくとも10分間氷の上に置きます。
5. ライブラリと選択の継続的な処理
- 低温バイアルから冷たい1.5 mLチューブにエマルジョンを移し、0.5 M EDTAの1μLを加え、室温で5分間13,000 x gで遠心分離します。
- ピペットで上部オイル相を取り除きます。油水相が見えない場合は、水相を凍結するために-20°Cで少なくとも5分間チューブをインキュベートし、すぐに液体油相をパイプアウトします。
- 10 mM Tris HCl pH 8.0の100 μLを加え、13,000 x gで30秒の遠心分離を経て、短い渦を流してフェノール:クロロホルム(1:1 v/v)の150 μLですぐに抽出を行います。上部水相を収集します。
- 3 M酢酸ナトリウム(pH = 5.2)、グリコーゲン2.5~5μg、エタノール2.5~5~5~2.5~2.5~2.5~2.5~2.5~エタノールを添加し、DNAを沈殿させます。-20°Cで1時間、遠心分離機を15分間13,000xg、4°Cでgインキュベートします。上清を捨て、1mLの冷たい70%エタノールでペレットを短時間洗います。
- DNA/グリコーゲンペレットをスピードバックまたはエアドライで5分間乾燥させます。
- ペレットを10 mMトリス-HCl(pH = 7.5)の50 μLに溶かします。メーカーの指示に従って調製したストレプトアビジン磁気ビーズを5 μL加え、できればカルーセルミキサーまたは穏やかな渦で1時間混合します。
- 磁気スタンドでビーズを分離し、ビオチンなしでDNAに富んだ液体を収集します。
- エタノール沈殿によってDNAを濃縮する(ステップ5.4~5.5)。
- 前工程の濃縮DNAを5μLの水に溶解し、F2およびR1プライマーとのPCR反応で鋳型として使用する(表1A)。
注:テンプレート汚染の問題を回避し、PCR アーティファクトを最小限に抑えるために、Taq ポリメラーゼ (Pfu または Phusion ではない) を使用し、テンプレート サイズ (1 kb = 1 分) に比例した延長時間で 18 ~ 20 サイクルを実行します (表 2Bを参照)。 - アガロースゲルでPCR産物を分画し、期待サイズの製品を切り取ります。一部のスミアは、異なるサイズの製品があることを示します(図6を参照)。ゲルスラブからDNAを市販キットで精製します。
- ステップ 5.10 から最大 50 ng の DNA と、ステップ 5.9 と同じプロトコルを使用してプライマー F1 および R2 を使用して、2 番目の PCR 反応を実行します。5.10に記載された製品の精製を進めます。アガロースゲルデンシソメトリー(UV分光法ではない)による定量後の精製DNAは、次のインビトロ選択のラウンドで使用することができる(ステップ4.6)。
6. 変更された順序特異性の画面バリアント
- 選択したバリアントを複製します。
- ORFのクローニングに適した制限酵素の10 Uで2時間のステップ5.10から製品を消化し、酵素ベンダーが推奨する温度とバッファーで発現ベクター(NlaIV:NcoIおよびXhoIの場合) にします。アガロースゲル電気泳動で製品を解決し、予想サイズの断片を分離します。
- プラスミドベクター(例えば、pET28)をステップ6.1.1と同じ酵素で二重切断して調製し、製品を市販のDNAゲル精製キットでゲル精製します。
- アガロースゲル電気泳動による濃度測定により、ベクターおよびインサートの濃度を推定します。
- T4 DNAリガーゼとベクターの1〜5 Uでライゲーションを設定:酵素ベンダーが推奨する1xリガーゼバッファーの1:3〜1:5のモル比を挿入します。RTで2時間インキュベートし、変換またはエレクトロポレーション12によって適切な宿主細菌(ステップ1.3から)に導入する。
- 適切な抗生物質(pET28またはpET30ベクターの場合は50μg/mLのカナマイシン)および1%グルコースを含むLBプレート上のエトランスタントを選択します。
- 発現タンパク質変異体。
- 単一コロニーをトランスフォーメーションから単一コロニー(1回の実行で簡単に処理できる)から、2mLのLBにカナマイシン(50μg/mL)、1%グルコースを加え、37°Cで一晩揺れで成長させます。
- 100 μg カナマイシンを含む 15 mL の暖かい (37 °C) LBを、一晩培養した 0.75 mL のブドウ糖を含み、激しく振るとともに 37 °C でインキュベートします。
注:50 mL遠心分離管または100 mLエルレンマイヤーフラスコを使用することができます。 - 176 μLのグリセロールを一晩培養した1 mLに加える(グリセロールの最終濃度= 15%)十分に混ぜ、-70°Cで凍結してください。
- 2–3 h の後、15 mL の培養物(ステップ 6.2.1 から)、IPTG を 1 mM に、培養するとさらに 5 時間になります。
- 細菌ペレットを遠心分離(10,000 x g、4°C、10分)で回収し、-70°Cで凍結します。 g
- タンパク質変異体を精製します。
- ニッケル親和性樹脂懸濁液20μLを、広いボアピペットチップを備えた1.5mLチューブ内のB1バッファの200μLに移し、穏やかに混ぜ、遠心g分離機(5,000 x g、30 s、4°C)を混合します。ピペットで上清を取り除き、チューブを氷の上に置いておく。
- B1のステップ6.2.5から細菌ペレットを激しい渦を起こし、B1の300 μLで再懸濁する。サスペンションを1.5mLチューブに移します。
- B1に100xプロテアーゼ阻害剤カクテルとリソソーム溶液を3μL加えます(最終濃度は1mg/mL)。先端装備のプローブと超音波処理によって細胞を崩壊させます。>15 s 先端冷却時間を間の氷で、サンプルごとに 6 つの 10 秒バーストを使用します。細胞懸濁液を常に氷の上に置いてください。
- 遠心分離(2分、12,000 x g、4°C)によるgペレット細胞デブリス、およびステップ6.3.1から樹脂アリコートに250μLの上清を移す。
- 冷たい部屋で15分間混ぜ、好ましくはカルーセルミキサーで、または穏やかな渦によって。
- 遠心分離機(5,000xg、30s、4°C)とピペットで上清を吸引する。 g
- 500 μL の W バッファーを加え、樹脂を緩やかに再懸濁します。遠心分離機(5,000xg、30s、4°C)とピペットで上清を吸引する。 g
- ステップ 6.3.7 を繰り返します。
- バッファーEを20μL加え、静かに樹脂を再懸濁し、試料を2~5分間氷上に置いて、遠心分離機(5,000 x g,30s,4°C)を残し、上清を回収します。
- ステップ 6.3.9 を繰り返します。プール上清。
- SDS-PAGE(5-10 μL)でタンパク質サンプルを分析する(図7)。
- 特殊性が変更されたバリアントの画面。
- バクテリオファージラムダDNAに対するアッセイ切断活性。タンパク質サンプルは、最終反応量の最大10%を構成することができます。0.5 μgのDNAおよび2時間の反応時間あたりの合計2 μLのタンパク質サンプルが良い出発点です。
- アガロースゲル電気泳動による反応生成物と、野生型酵素によって生成された生成物を分析します。更なる分析のために野生型酵素によって生成されたものと明確に区別できる切断パターンを生成するクローンを選択する(図8)。
結果
このプロトコルは、不活性酵素と不変の野生型配列特異性を持つエンドヌクレアーゼの2つの不要なクラスを枯渇させることによって、設計されたREaseの望ましい変異体の頻度を増加させるだけのツールです。一方、REase特異性の変化は極めて困難であるため、24クローンの単一スクリーニングにおいて野生型酵素とは異なる切断パターンを産生するそのような変異体を1つでも見つけることは成功と考えるべきである。私たちの手の中で最高の画面は有望な変種の最大20%を識別することができた(図8A)。
肯定的な結果は、ライブラリの品質(すなわち、置換の頻度とランダム分布の制限)と図書館メンバーのビオチン化された集団の効率的な捕獲(ステップ3.6-3.7)に強く依存する。両方の問題を検出できます。ライブラリの品質は、可能な限り多くのクローンをシーケンシング(>15)、または高スループットのシーケンス処理によるライブラリの直接シーケンスによって、選択前にチェックする必要があります(ステップ3.10、表3)。選択したクローンの大部分がアクティブでない場合、これはストレプトアビジンのキャプチャ選択の失敗を明確に示す。多くの選択サイクルを経るライブラリの場合も同様の効果が見られるが、そのようなライブラリはストレプトアビジン捕獲選択ステップを逃れた非アクティブな変異体によっておそらく支配されている可能性が高いからである(図8B)。したがって、選択サイクルのたびにスクリーニングを実行し、選択反復に依存するのではなく、手動で選択した有望なバリアントをさらに開発することをお勧めします。

図1:NlaIV工学に基づく新しい配列特異性のインビトロ選択。(A)式/選択カセット(ESC)の組織には、REaseの2つの認識部位、1)選択したシーケンス(GGATCC)が右端に近い、2)左側に近いカウンター選択シーケンス(GGSSCC)、T7pおよびT7t-T7プロモーターおよびT7ターミネーターが含まれる。プライマー結合部位を以下に示す。野生型および選択されたNlaIV変異体による切断は、それぞれ赤と緑の三角形として示される。(B) 選択サイクルステップ: I) ESCライブラリと転写翻訳-切断反応の乳化が混入する;II)すべてのビオチン化されたDNAは、ストレプトアビジンでコーティングされた磁性粒子上で捕捉され、除去され、非活性変異体のコードを除去する。III)ESCは野生型活性を有するREases(すなわち、GGSSCC配列を切断することができるもの)は、配列の切断が前方および逆プライマーの結合部位を分離するので排除される。したがって、これらのESCの増幅は起こりません。IV) 次の選択ラウンドの入力は、右端にビオチンを添加し、左端に選択されたカウンターシーケンスのバリエーションを再導入することによって作成されます。ツァピンスカら8世からエルゼビアの許可を得て転載。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図2:ESCの準備T7プロモーターの制御下でNlaIV ORFを含む発現ベクター中の元の構築物から導出された断片は、発現/選択に適するように改変した。NlaIV ORFから下流のNlaIVサイトが削除され、選択した位置を変異させるために使用されたユニークなサイト(SalI、EcoRIおよびEco52I)がサイレント突然変異としてNlaIV ORFに導入されました。最終的な構造は、2つの側面NlaIVサイトを導入した横向きプライマーで増幅された:左側に選択された配列(GGSSCC)と右側の選択されたシーケンス(GGATCC)のカウンター。逆プライマーはまた、ビオチンを導入しました。変異ECSの作成に使用されるプライマーは、青色の矢印で示され、下にラベルが付いています(表1B,Cを参照)。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図3:分割と混合合成のスキーム。この例は、4つの位置で0.8周波数でNNSシーケンスが導入されたMutBプライマー合成を指します(表3も参照)。化学合成は3'から5'まで行われるが、全ての配列は正規5'-3'の向きで示されている(すなわち、このスキームでは右から左へ進む)。変異原性の位置における野生型配列は緑色で示され、NNS変異原性配列は赤色である。後にESCに変異を導入するために使用されるSalI認識部位には下線が引かれている。混合および分割ステップのポイント(2および4)が示されている。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図4:オリゴヌクレオチド標的突然変異誘発における独特の制限酵素部位の使用。変異導入の戦略は、ライブラリA-Cの構築例に示されています(ステップ3.1~3.7を参照)。ツァピンスカら8世からエルゼビアの許可を得て転載。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

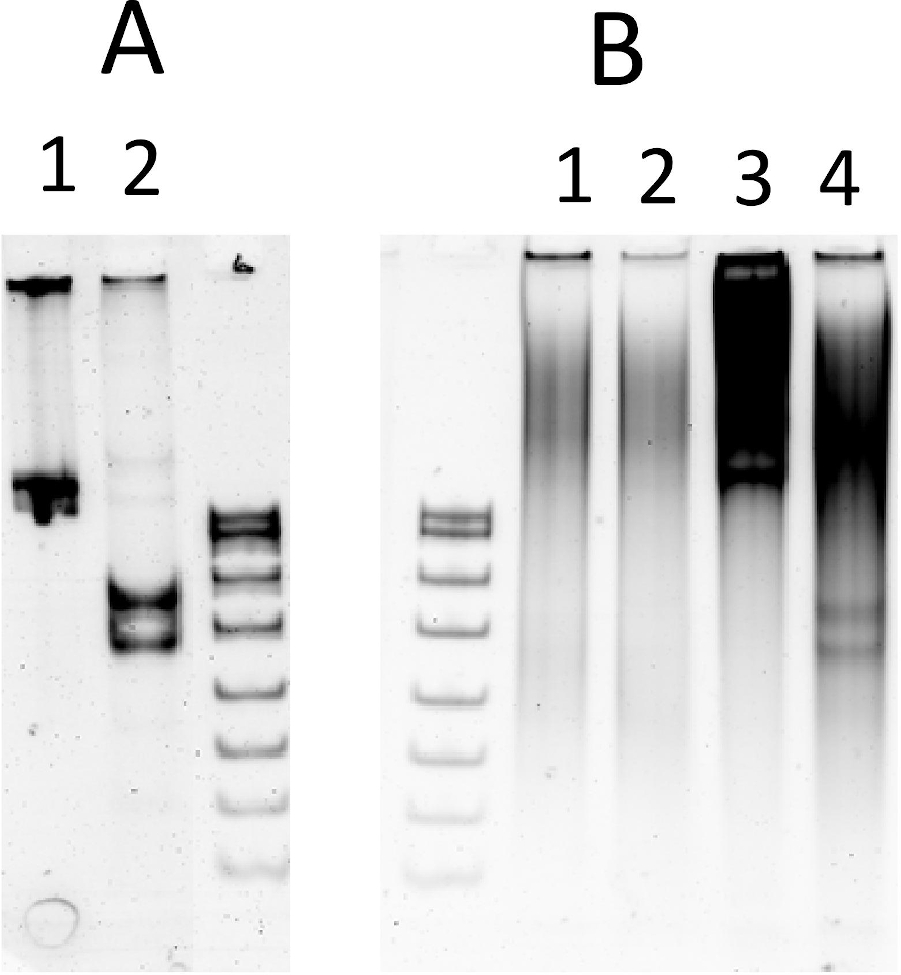
図5:インビトロ転写翻訳における内核分解性切断。(A)最適な REase バッファーにおけるテスト基質の切断: 1) 基質、単一の NlaIV 認識部位を持つ 612 bp PCR 産物;2) 切断生成物, 355 bp および 257 bp. (B) in vitro転写変換反応における切断 (ESCの0.5 μgを含む): 1-2) 15 μL のインビトロ転写翻訳のアリコート(ライン2:1.5 mM MgCl2を補った)3–4) 1μGの試験基板を用いたインビトロ転写翻訳の15 μLアリコート;(ライン 4: 1.5 mM MgCl2を補った反応。S-DNAサイズマーカー(mspIで消化されたpBR322)。サンプルは6%ネイティブPAGEで解決されました。DNAを臭化エチジウムで染色した。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

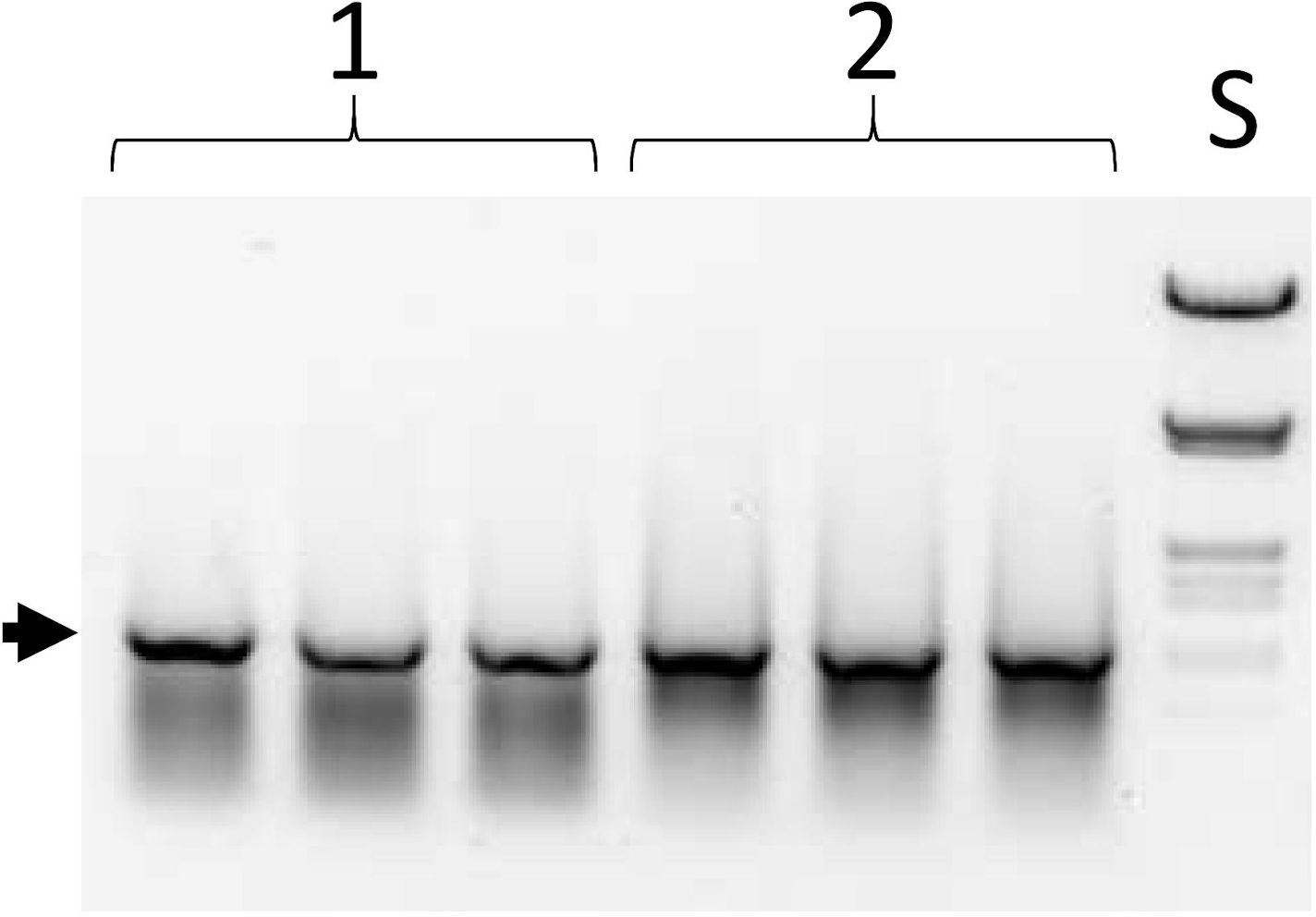
図6:選択サイクルにおける最初のPCRの製品。図 1B、ステップ III を参照してください。プロトコルステップ5.10。列セット 1 と 2 は、三重にロードされた 2 つの異なるライブラリのアリコートです。S-DNAサイズ標準(ヒンドIIIおよびエコリオンで消化されたラムダDNA)。矢印は、全長 ESC (1,050 bp) の位置を示します。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

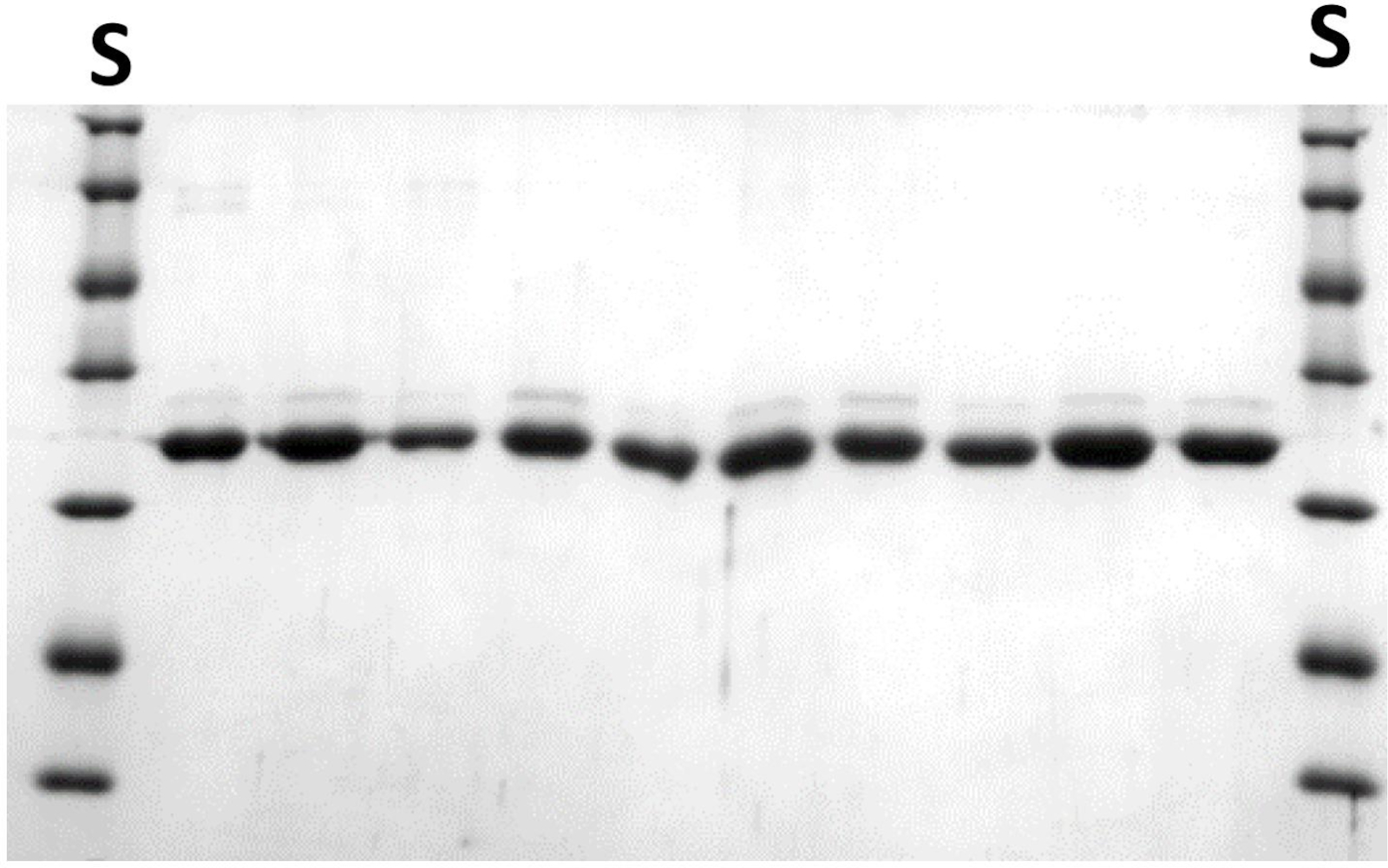
図7:ミニスケールでさらなるスクリーニングのために精製されたNlaIV変異体。ステップ 6.3.11 を参照してください。各行には、異なるバリアントの 10 μL アリコートが含まれています。Sタンパク質分子量標準。NaIV REaseサブユニットの分子質量は29.9 kDaである。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図8:配列特異性変化のためのNlaIVバリアントのスクリーニング例ステップ 6.4.2 を参照してください。(A) 有望な変異体の高頻度でのスクリーニングに成功した。S = DNA サイズ マーカー、ヒンダ III とエコRI と切断されたラムダ DNA;野生型 (wt) = ラムダ DNA は野生型 NlaIV で切断;λ=λDNA基質、切断されない;その他の列=非常に低いアクティビティを持つバリアント。バリアントにはラベルが付いています!=野生型酵素とは異なる切断パターンを生成する有望な変異体; ?= シーケンスの優先順位も変更された可能性があるバリアント。(B) 不活性の変異体の大部分と明らかに変化しない切断パターンを持つ 1 つのバリアントを伴う、スクリーニングに失敗した。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

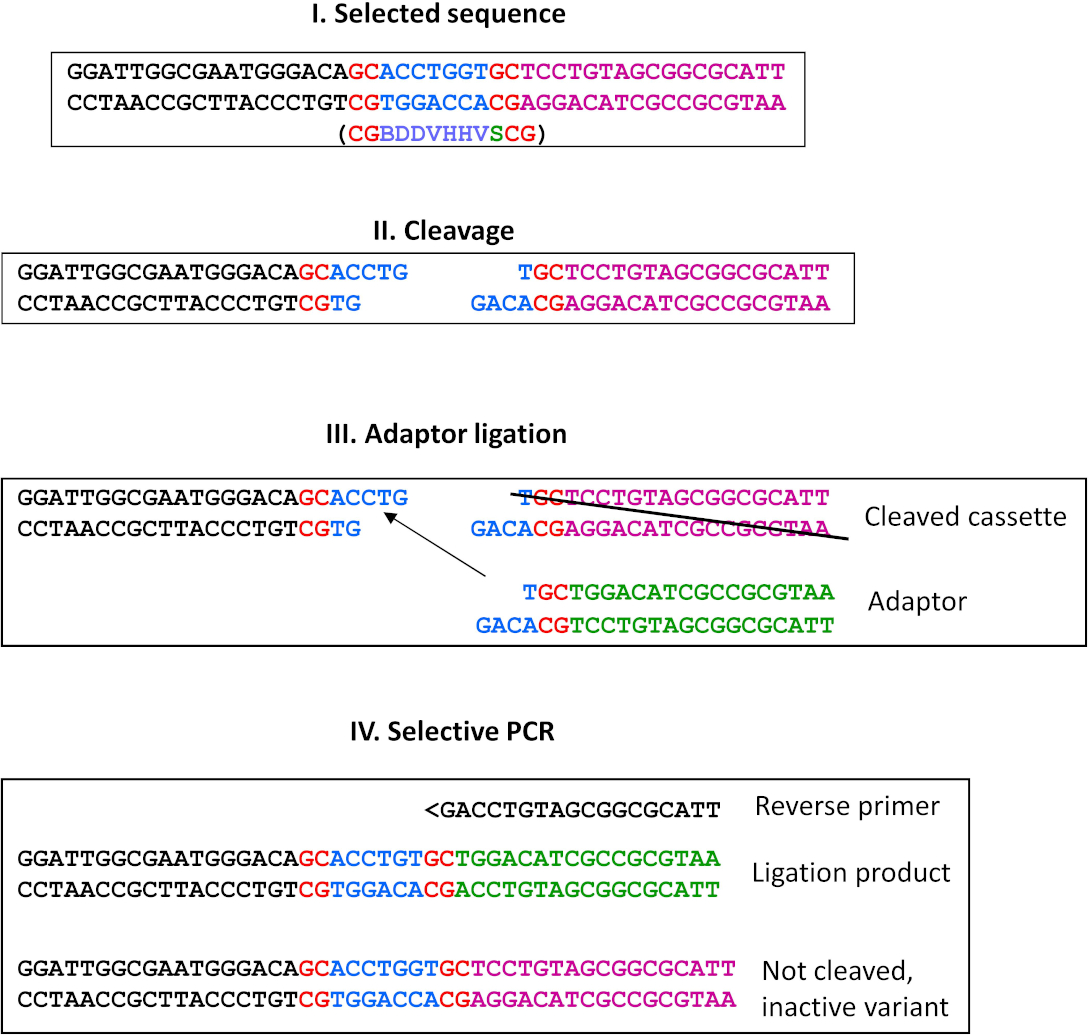
図9:ライゲーションによる代替選択。この代替は、スティッキーエンドを生成するすべてのREaseに使用できます。ここでは、MwoI酵素(未発表)の選択スキームのプロトコルの例を紹介します。I)選択された配列(ESCの右端に位置)で、定義された残基が赤で示され、青色で示された同結合配列のバリエーションが選択された。ESC の左端に配置するカウンタ選択シーケンスの下のかっこ内に表示されます。II) MwoI切断の産物;III) インビトロ転写/翻訳を終了した後、製品を精製し、過剰なアダプターでライゲーションを行います。選択された配列で切断された切断産物のみがライゲーションに関与することができる。したがって、非アクティブなバリアントは削除され、プルダウンステップは不要です。カウンター選択されたシーケンスの切断物 (ESC の左端に、図示せず) は、アダプタの突出先端が選択されたカウンターシーケンスと相補的ではないため、このライゲーションに参加できません。IV) 選択的 PCR は、メイン プロトコルと同じ戦略を使用して、野生型の退化配列特異性 (F1 プライマー結合遠位から選択された部位に遠位) を持つ変異体を除去するのに対し、非活性変異体は、非切断(したがって、アダプタ結紮によって修飾されない)に結合できない選択的リバースプライマーによって排除される。次のサイクルでは、前工程の切断産物と同一のアダプター(すなわち、パネルIIIの「切断カセット」)と、適切な選択的リバースプライマーを用いてプロセスを反復処理することができる。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
表1 NlaIVエンジニアリングで使用されるプライマーコメントに記載されている制限サイトのシーケンスには下線が付いています。小さな文字は、DNAテンプレートに補数を持たない配列を示します。こちらの表を表示するには、ここをクリックしてください (右クリックしてダウンロードしてください)。
表2:プロトコルに使用するPCR反応の条件。Tm = プライマー融解温度(プライマーに対して Tmが異なる場合は、低い Tmを使用する必要があります)。こちらの表を表示するには、ここをクリックしてください (右クリックしてダウンロードしてください)。
表3:スプリットアンドミックス戦略で合成した2つの変異原性プライマーの品質チェックの結果。変異原化コドンは[XXX]で示される。低いインデックス数は、コード化されたアミノ酸の位置を示します。ツァピンスカら8世からエルゼビアの許可を得て適応。こちらの表を表示するには、ここをクリックしてください (右クリックしてダウンロードしてください)。
表4:EP-PCRの結果ECSの22クローンの配列解析から導出された主要なパラメータ。こちらの表を表示するには、ここをクリックしてください (右クリックしてダウンロードしてください)。
ディスカッション
ここで説明した選択プロトコルは、NN基地の中央を有するパリンドローム標的部位を認識し、NN基地間の鈍い端部切り線を触媒する二量体PD-(D/E)XK折り返し認識シーケンスであるNlaIV8について試験された。Nn塩基間の切断は、これらの塩基が複合体中のタンパク質に近いということを示唆しているので、NlaIVが選ばれた。原則として、このプロトコルは、触媒ドメインと特異性ドメインが一致するか(例えば、FokI)であるかにかかわらず、任意の折りたたみグループの任意のシーケンス固有の制限エンドヌクレアーゼ、単量体または二量体に使用することができる。さらに、原則としてこのプロトコルは、新しい、より狭い酵素特異性の生成に有用であるだけでなく、星の活性を排除したり、高忠実度のエンドヌクレアーゼを作成するためにも使用することができる。しかし、このすべてはまだテストされていません。特に、同じアミノ酸残基が所望および望ましくない塩基への結合に関与する可能性があるため、スター活性の標的排除は複雑であり得る。このプロトコルで説明されているin vitroステップは、限定的に絞られた特異性の選択に限定されるものではなく、それ以外の変更された特異性を選択するためにも使用することができる。しかし、その後、変異型エンドヌクレアーゼに問題がある:基質のスペクトルが親エンドヌクレアーゼによって切断されない新しい標的を含む場合、一般的には、この活性の有害な影響から細胞を保護する良い方法はない。対照的に、エンドヌクレアーゼ特異性が絞り込まれるだけなら、標的は野生型の標的のサブセットであり、したがって既に利用可能な結合メチルトランスフェーゼは完全に保護されるべきである。
私たちのプロトコルは、多くの指向進化プロトコルとは異なります。オープンリーディングフレームの多様性は、実験の開始時に一度生成され、すべての反復処理では生成されません。さらに、EP-PCRではなく、スプリットアンドミックス合成によって作成されます。コドンのNNS置換については、本研究で用いられているように、6つの位置に対して(4 x 4 x 2)6~1.07 x 109の組み合わせがある。したがって、任意のバリアントは、ESCの1.7 fmolesに一度平均で存在します。この容量は、グレンリサーチが提供する20のトリヌクレオチド前駆体の混合物との合成を使用するか、またはスプリットアンドミックスオリゴヌクレオチド合成であまり有望な位置で突然変異頻度を減少させることによって、7つの位置に増加させることができます。可能であれば、変動の範囲を6桁に制限することをお勧めします。明らかに、このような変異誘発標的化は、基質結合に関与する少なくともREaseの領域に関するいくつかの既存の知識を必要とする。多様性を生成するスプリットアンドミックスプロトコルは、EP-PCRと比較して明らかな利点を有します。EP-PCRを用いて、同じEP-PCRでNlaIV ESCに対して8個の置換を行う不変の変異体と配列を得た(表4)。EP-PCRのライブラリには、避けるべきクローンのかなりの部分(野生のタイプ配列、複数の置換、フレームシフトとナンセンス突然変異、および配列特異性に影響を与える可能性が低い場所での突然変異)が含まれています。
また、我々のプロトコルは、2つの逐次選択ステップの存在によって、他の多くの指向進化プロトコルとは異なる。正の選択は、望ましい活性が保持されていることを確認し、そうでなければビオチンタグは除去されず、コード配列はプルダウンによって除去することができる。技術的には、新しい、重複しない特異性(GCATGCなど)の偶然の出現は、適切な切断部位が所望の切断の近くに存在するが、他の場所に存在しない場合、ビオチンタグの切断につながる可能性がある。しかし、これは可能性が非常に低いはずです。負の選択は、まだ望ましくない活性を持つ酵素をコードするオープンリーディングフレームを削除します。この手順は、選択シーケンスを切断できるが、ESCの他の場所で切断することができないバリアントを使用して出力ライブラリを強化するため、厳密には必須ではありません。しかし、元の配列特異性を持つ酵素は出力から除去されず、特異性が変化する有望な変異体より、酵素活性も低下するため、選択効果は低くなると予想されます。母集団レベルでは、望ましいターゲットシーケンスと望ましくないターゲットシーケンスの両方が退化する必要はない点に注意してください。NlaIVの例では、アンチターゲットは退化し、ターゲットは非退化した。人口レベルで縮退がある場合でも、単一の液滴では1つの(非退化)標的または抗標的のみが存在する。私たちのプロトコルでは、ターゲットとアンチターゲットシーケンスは、選択ステップの繰り返しごとに再導入されます。したがって、オープンリーディングフレームは、複数の選択ラウンドを生き残るために、可能なすべてのターゲットを切断し、抗標的のいずれを切断することができない酵素をコードする必要があります。プロトコルの各反復で、選択解除対象を再導入する必要がある場合は、2 つのシーケンシャルな PCR が適用されます。最初のPCRは、抗標的の外側にアニールするプライマーを使用し、抗標的の切断がPCR反応を防止するようにする。2 番目の PCR では、アンチ ターゲットを超えて到達するプライマーを必要とし、アンチ ターゲットを再導入し、複数の選択ラウンド中に、各オープンリーディング フレームがアンチ ターゲットのすべてのバリアントに対してテストされることを確認します。
粘着末端を生成する酵素の場合、REase ORF10の分離のための前述の方法に基づく関連代替プロトコルを使用することができる。我々の実験で使用されるビオチン捕捉による不活性変異体の枯渇は、選択的PCRにおいてプライマー結合部位として使用される配列を有する適合アダプタのライゲーションによって代替プロトコルに置き換えられる(図9)。選択した特異性を有する酵素を産生するESCのみがライゲーション可能な末端を生成し、選択されます。カウンターの末端の付着の配列は、アダプタとの結紮に関与できないように設計されなければならない。選択プロセスの反復は、2つの異なるアダプタと結果として選択的PCRにおける2つの異なるリバースプライマーを切り替えることによって容易に達成することができる。
新しいプロトコルがあっても、インビトロでの新しい特異性のエンジニアリングのタスクはまだ非常に困難です。典型的なII型のリーズでは、配列特異性および内核分解活性は同じタンパク質領域に依存する。したがって、他方に影響を与えずに1つを変更することは困難です。成功は、酵素のフットプリントを考慮に入れ、タンパク質とDNA相互作用の対称性を尊重し、NlaIV例8で行われたように、生化学的実験で事前に決定されるべき既存の酵素好みに基づいて構築される戦略によってより可能性が高い。
開示事項
著者らは開示するものは何もない。
謝辞
この研究は、ポーランド国立科学センター(NCN)(UMO-2011/02/A/NZ1/00052)から、科学高等教育省(0295/B/PO1/2008/34~MB、N301 100 31/3043からKS)からの助成金によって支えられ、 UMO-2014/13/B/NZ1/03991およびUMO-2014/14/M/NZ5/00558からMB)およびKSへの短期EMBOフェローシップ(ATSF 277.00-05)。
資料
| Name | Company | Catalog Number | Comments |
| 1000Å CPG Support (dA, dT, dC, dG) | Biosset | 45-1000-050 | Other vendors can be used as well |
| ASM-800 DNA/RNA | Biosset | 800-001-000 | |
| GeneJET Gel Extraction Kit | Thermo Scientific | K0691 | Any other kit can be used |
| Glen-Pak DNA purification cartridge | Glen Research | 60-5200 | |
| HIS-Select Nickel Affinity Gel | Sigma | P6611 | |
| pET 28a vector | Any other vector with T7 promoter upstream of plycloning site can be used instead | ||
| Phusion High-Fidelity DNA Polymerase | Thermo Scientific | F530S | Any other high fidelity and highly processive thermophilic polymearse can be used instead |
| Porous steel foil | Biosset | 40-063 | |
| Rapid Translation System RTS 100, E.coli HY Kit | Roche | 3 186 148 | |
| Restriction endonucleases | Thermo Scientific | Obviously other vendors, enzymes can be used | |
| Streptavidin Magnetic Beads | New England Biolabs | S1420S | Other vendors can be used as well. We have positively tested beds form Sigma |
| Synthesis chemicals including phosphoramidities | Carl Roth | Other vendors can be used as well | |
| Synthesis columns (different sizes) | Biosset | ||
| T4 DNA ligase | Thermo Scientific | EL0011 | Any other ligase can be used |
参考文献
- Schöttler, S., Wenz, C., Lanio, T., Jeltsch, A., Pingoud, A. Protein engineering of the restriction endonuclease EcoRV--structure-guided design of enzyme variants that recognize the base pairs flanking the recognition site. European Journal of Biochemistry. 258 (1), 184-191 (1998).
- Wenz, C., Hahn, M., Pingoud, A. Engineering of variants of the restriction endonuclease EcoRV that depend in their cleavage activity on the flexibility of sequences flanking the recognition site. Biochemistry. 37 (8), 2234-2242 (1998).
- Samuelson, J. C., Xu, S. Y. Directed evolution of restriction endonuclease BstYI to achieve increased substrate specificity. Journal of Molecular Biology. 319 (3), 673-683 (2002).
- Samuelson, J. C., et al. Engineering a rare-cutting restriction enzyme: genetic screening and selection of NotI variants. Nucleic Acids Research. 34 (3), 796-805 (2006).
- Rimseliene, R., Maneliene, Z., Lubys, A., Janulaitis, A. Engineering of restriction endonucleases: using methylation activity of the bifunctional endonuclease Eco57I to select the mutant with a novel sequence specificity. Journal of Molecular Biology. 327 (2), 383-391 (2003).
- Morgan, R. D., Luyten, Y. A. Rational engineering of type II restriction endonuclease DNA binding and cleavage specificity. Nucleic Acids Research. 37 (15), 5222-5233 (2009).
- Skowronek, K., Boniecki, M. J., Kluge, B., Bujnicki, J. M. Rational engineering of sequence specificity in R.MwoI restriction endonuclease. Nucleic Acids Research. 40 (17), 8579-8592 (2012).
- Czapinska, H., et al. Crystal Structure and Directed Evolution of Specificity of NlaIV Restriction Endonuclease. Journal of Molecular Biology. 431 (11), 2082-2094 (2019).
- Miller, O. J., et al. Directed evolution by in vitro compartmentalization. Nature Methods. 3 (7), 561-570 (2006).
- Zheng, Y., Roberts, R. J. Selection of restriction endonucleases using artificial cells. Nucleic Acids Research. 35 (11), e83(2007).
- Takeuchi, R., Choi, M., Stoddard, B. L. Redesign of extensive protein-DNA interfaces of meganucleases using iterative cycles of in vitro compartmentalization. Proceedings of the National Academy of Science U.S.A. 111 (11), 4061-4066 (2014).
- Howland, J. L. Short Protocols in Molecular Biology. Ausubel, F., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., Struhl, K. , Third edition, John Wiley & Sons. New York. (1995).
- Wilson, D. S., Keefe, A. D. Chapter 8 Unit 8.3: Random mutagenesis by PCR. Current Protocols in Molecular Biology. , John Wiley & Sons. New York. (2001).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved