
Method Article
더 엄격한 특이성을 가진 제한 endonuclease의 시험관 진화
요약
새로운 서열 특이성을 가진 제한 endonucleases는 부분적으로 퇴화한 서열을 인식하는 효소로부터 개발될 수 있다. 여기에서 우리는 우리가 성공적으로 NlaIV 효소의 순서 특이성을 바꾸기 위하여 이용된 상세한 프로토콜을 제공합니다. 프로토콜의 주요 성분은 전사/번역 반응의 생체외 구획화 및 새로운 서열 특이성을 가진 변이체의 선택이다.
초록
제한 엔도너첼리스(REase) 특이성 엔지니어링은 매우 어렵습니다. 여기에서 우리는 부모 효소 보다는 더 엄격한 특이성을 가진 REase 이체를 생성하는 것을 돕는 다단계 프로토콜을 기술합니다. 이 프로토콜은 DNA 결합에 영향을 미칠 가능성이 있는 위치의 가변성을 가진 이상적으로 REase의 변이체에 대한 발현 선택 카세트(ESC)의 라이브러리를 생성해야 합니다. ESC는 바람직한 제한 부위 활성 및 비오틴 태그에 대한 서열에 의해 한쪽에 측면되고, 다른 쪽에는 바람직하지 않은 활성 및 프라이머 어닐링 부위에 대한 제한 부위에 의해 서열이 있다. ESCs는 액적 당 하나 이상의 DNA 분자의 존재를 가능성 있게 하는 조건에서 물-인-오일 에멀젼으로 전사되고 번역됩니다. 따라서, 각 카세트 분자의 DNA는 번역된, 인코딩된 효소의 활성에만 적용된다. 원하는 특이성의 REase 변이체는 프라이머 어닐링 부위가 아닌 비오틴 태그를 제거한다. 에멀젼을 파괴 한 후 DNA 분자는 비오틴 풀다운을 받게되며 상류에 있는 분자만 유지됩니다. 이 단계에서는 원하는 활성을 잃지 않은 변형에 대한 ESC만 유지됩니다. 이 DNA 분자는 그 때 첫번째 PCR 반응을 복종됩니다. 바람직하지 않은 시퀀스에서의 절단은 프라이머 중 하나에 대한 프라이머 결합 부위를 차단한다. 따라서 PCR은 원치 않는 활성없이 액적에서 만 ESC를 증폭시다. 제2 PCR 반응은 선택 단계가 반복될 수 있도록 원하는 특이성 및 비오틴 태그에 대한 제한 부위를 재도입하기 위해 수행된다. 선택된 개방 판독 프레임은 또한 부모 REase의 코그네이트 메틸 트랜스퍼라제를 발현하는 세균 세포에서 과발현될 수 있으며, 이는 새로 진화된 REase가 메틸트랜스퍼라제 표적 부위의 서브세트만을 대상으로 하기 때문이다.
서문
시퀀스 특이성 엔지니어링은 클래스 II REases에 매우 까다롭습니다. endonucleases의이 클래스에서, 서열 인식 및 촉매는 밀접하게 얽혀, 가장 아마 그것의 동체 메틸 트랜스퍼 라제보다 더 넓은 특이성의 엔도누글레스의 생성에 대한 진화적 보호로, 이는 호스트 DNA를 손상시킬 것이다. 세포에 있는 새로운 특이성의 지시된 진화는 새로 설계된 endonuclease 활동에 대하여 호스트 DNA를 보호하는 필요에 의해 더 복잡합니다. 따라서 REase 엔지니어링의 몇 가지 성공적인 시도가보고되고 그들 모두는 특정 효소1,2,,3,3,4,,5,,6,,7의독특한 특징을 이용한다.
여기에서 우리는 NlaIV endonuclease8의우리의 성공적인 공학에 근거를 둔 부모 효소 보다는 더 좁은 특이성을 가진 endonuclease 이체를 생성하는 것을 이용될 수 있는 특이성 공학을 위한 상세한 프로토콜을 제공합니다. 임의 인식 서열을 가진 임의의 효소의 경우, 측면에 있는 염기들을 위해 추가 특이성을 도입할 수 있다. 부분적으로 퇴화된 서열을 인식하는 부모 효소(예: GGNNCC 표적을 가진 NlaIV)의 경우, 추가특이성은 또한 인식 서열 내에서 도입될 수 있다. 추가 특이성은 가능성이 단백질 DNA 접촉을 요구할 것이기 때문에, 새로 인식된 기지는 DNA에 부모 endonuclease의 발자국 안에 놓여야 합니다. 원칙적으로, 선택 방식은 인식 시퀀스의 임의의 원하는 전문화에 대해 설정될 수 있다. 그러나, 팔린드로믹 및 거의 팔린드로믹 표적 서열을 인식하는 대부분의 REases는 팔린드롬의 반부위만을 인식하는 기능적 이량체이다. 그러므로, 단백질 핵 상호 작용의 대칭을 위반하는 새로운 특이성의 선택은 작동하기 위하여 확률이 낮습니다. 디메릭 NlaIV의 경우, 예를 들어, GGNNCC 서열은 이론적으로 GGATCC로 좁힐 수 있지만 GGAACC로의 특이성을 좁히는 것은 더 어려울 것으로 예상된다. 우리의 계획은 긍정적이고 부정적인 선택을 모두 포함한다.
이 공정은 음의 선택이 바람직한 좁은 특이성 이외의 모든 서열을 분리할 수 있는 특이성을 제거하는 데 사용될 때 더욱 효율적입니다. 예를 들어 GGATCC에 대한 선택은 GGBVCC에 대한 선택 방지와 결합될 수 있습니다(B는 A 이외의 베이스이고 V는 T 이외의 베이스입니다). 가능한 대상 서열 중 일부가 다루지 않는 경우 선택 실험의 결과는 양수 및 음수 선택의 효과에 따라 달라집니다. NlaIV 작업에서 GGATCC및 GGSSCC(S가 G 또는 C인 경우)를 대상으로 선택하고 대칭 파괴 대상을 무시하고 GGWWCC(W가 A 또는 T인 경우)로 설명할 수 있는 특이성을 얻었으며, 이 경우 부정적인 선택이 더 많다는 것을 시사합니다. 긍정적 인 선택보다 중요합니다.
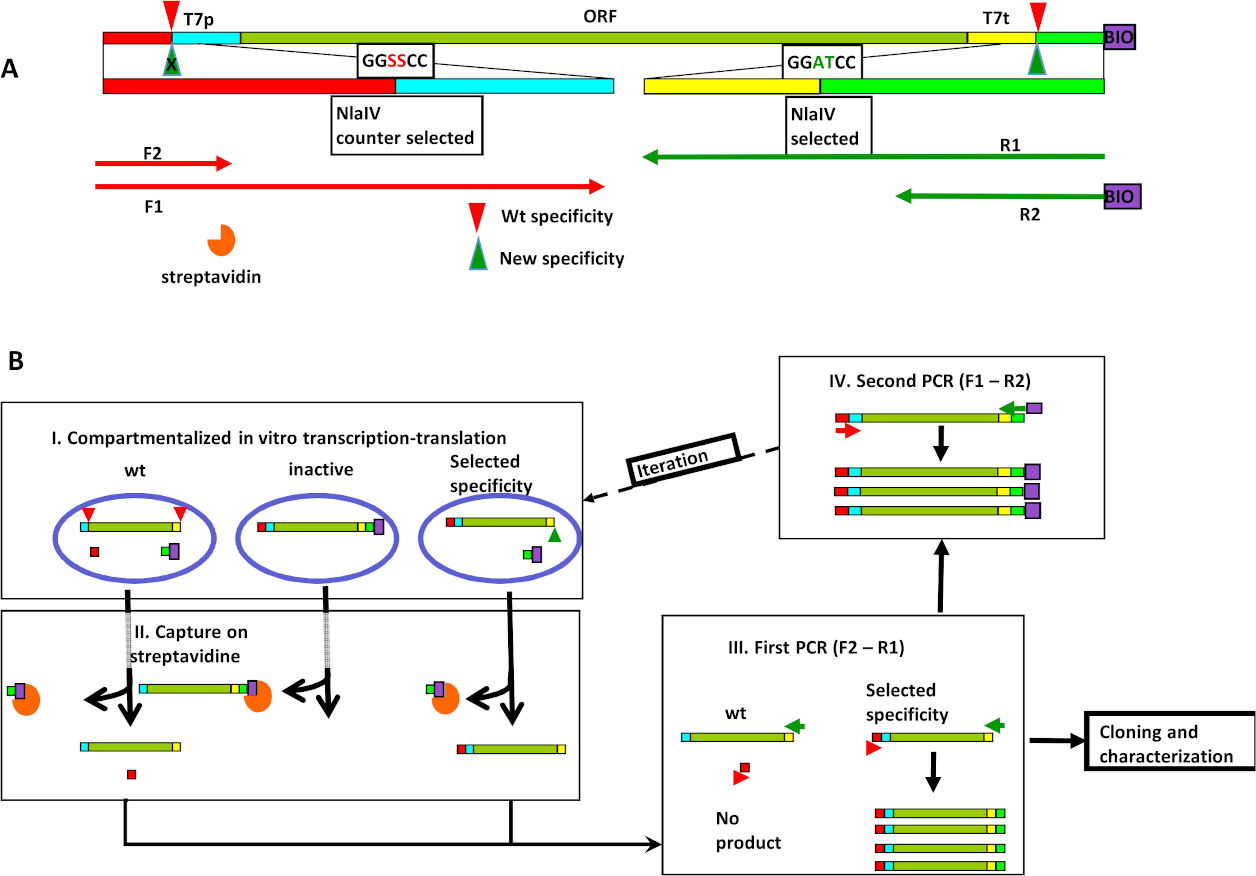
우리의 접근 방식은 식 선택 카세트 (ESC)의 생성으로 시작됩니다. ESC는 섹션으로 구성됩니다. 내부 코어 섹션에는 T7 프로모터 제어 하에 REase의 개방 판독 프레임(ORF)의 변이체가 있습니다. ESC의 이 핵심 섹션에는 엔지니어링된 REase에 대한 코그네이트 사이트가 포함되어 있지 않습니다. 코어는 야생형 REase에 대한 두 개의 동조 부위 사이에 끼어있다: 바람직하지 않은S활성을 위한 절단 부위(이 예에서 선택된 시퀀스, GGSCC)와 원하는 활성을 위한 절단 부위(예에서 선택된 시퀀스, GGATCC). PCR에서 ESC의 제조의 최종 단계는 5' 말단에서 원하는 활성에 가까운 비오틴을 추가하고 다양한 카운터 선택 서열을 생성한다(예에서 GGSSCC). 선택 전략은 시험관 내 전사/번역/선택프로토콜(그림 1A)을사용한 후 ESC 재증폭 프로토콜에서 신중하게 설계된 프라이머의 사용에 의존한다. ESC 라이브러리는 체외 구획화된 전사 번역 물-인-오일 에멀젼99,10,,11로표현된다. 각 액적 내에서, 발현된 효소의 특이성은 ESC의 상태에 영향을미친다(도 1B,단계 I). 기재된 배열을 위해, 번역된 단백질의 바람직한 절단 활성은 DNA의 비오틴 태그를 제거하지만, 카운터 선택서열로 다른 ESC 말단에 영향을 미치지 않는다. 에멀젼이 파손되면, 생체 티니글리드 단편은 스트렙타비딘 친화도 풀다운에 의해 제거되어 원하는 활성을 가진 액적으로부터의 단편만 남아 있게된다(그림 1B,단계 II). 이 단계는 비활성 REase 변형을 제거합니다. 풀다운 단계의 상급 분획은 PCR에 의해 증폭됩니다. 제1 PCR 반응 프라이머F2 및 R1이 사용된다(도1A, B,단계 III). 프라이머 F2는 카운터 선택서열과 분자 말단 사이의 ESC 섹션에 결합한다. 따라서, 카운터 선택 서열을 절단할 수 있는 변이체를 발현하는 ESCs(따라서, 프라이머 F2 및 R1에 대한 결합 부위를 두 개의 상이한 DNA 분자로 분리)는 증폭되지 않고 따라서 라이브러리로부터 제거된다. 프라이머 R1은 선택된 부위와 ESC의 코어 사이에 결합하여 선택된 부위의 절단 상태에 영향을 받지 않도록 하고 원하는 활성(GGATCC)에 대한 절단 부위를 복원한다. 사이클은 선택된 부위에 가까운 5' 끝에 비오틴을 추가하고 ESC의 반대쪽 끝에 가까운 카운터 선택 부위에서 설계 변형을 복원하는 제2 PCR(프라이머 F1 및 R2)에 의해 폐쇄된다(그림1B,단계 IV). 생성된 DNA 혼합물은 또 다른 선택을 위해 준비됩니다.
선택 프로토콜의 성공은 새롭고 더 엄격한 표적 인식 시퀀스의 적절한 선택과 돌연변이 유발 전략및 효과적인 구현의 신중한 설계에 크게 좌우됩니다. REase를 극복하는 것보다 REase의 기존 환경 설정을 약간 개선하는 것이 훨씬 쉽기 때문에 기존의 환경 설정에 대한 역학 연구로 시작하는 것이 좋습니다. 신중한 돌연변이 생성 설계의 필요성은 제시된 프로토콜에 의해 처리될 수 있는 돌연변이 라이브러리의9 제한된 크기로부터 발생한다(단일 실험에서 109클론). 따라서 모든 20 개의 가능한 아미노산 치환은 몇 가지 위치에서효과적으로 테스트 할 수 있습니다 (토론 참조). 대안으로 제시된 오류가 발생하기 쉬운 PCR(EP-PCR)과 같은 무작위 돌연변이 발생은 기존 복잡성의 심오한 하소 샘플링으로 이어질 것입니다. DNA와의 접촉에 관여하는 잠재적 아미노산 위치(또는 코그네이트 서열에서 퇴화된 뉴클레오티드에 근접한 위치)에 관한 정보가 있는 경우, 올리고뉴클레오티드 유도 포화 돌연변이 발생에 대한 몇 가지 아미노산을 선택하는 데 확실히 사용되어야 합니다(프로토콜 단계 1.6-3.10).
프로토콜
1. ESC 준비
- 낮은 카피 수 플라스미드(예를 들어, pACYC184 또는 pACYC174 또는 이들의 유도체)로 설계되는 제한-수정 시스템의 클론 메틸트랜스퍼라제.
참고: 세균성 숙주 균주는 복제 된 효소에 의해 도입 된 메틸화를 견딜 수 있어야하며 T7 RNA 폴리머 라제의 유도 성 발현을 제공해야합니다. ER2566 균주 (McrA, McrBC 및 Mrr 돌연변이를 운반하는)를 사용하는 것이 좋습니다. - 재조합 플라스미드 DNA가 0.5 μg의 플라스미드 DNA를 완충제 및 온도에서 10 단위의 완충제 및 온도로 처리하여 코그네이트 엔도누클로즈에 의한 절단으로부터 보호되는지 확인합니다.
- 이 균주의 유능한 세포를 준비하십시오.
참고: 모든 방법을 사용할 수 있습니다. NlaIV 엔지니어링 프로젝트는 간단한 염화칼슘 방법12를사용했습니다. - 1.1단계에서 메틸트라스래erase 유전자를 함유하는 것과는 다른 선택 마커로부터 다른 배제기로부터 T7 프로모터의 제어하에 REase에 대한 ORF와 재조합 플라스미드를 구성한다. 벡터 pET28 및 pET30을 사용할 수 있다.
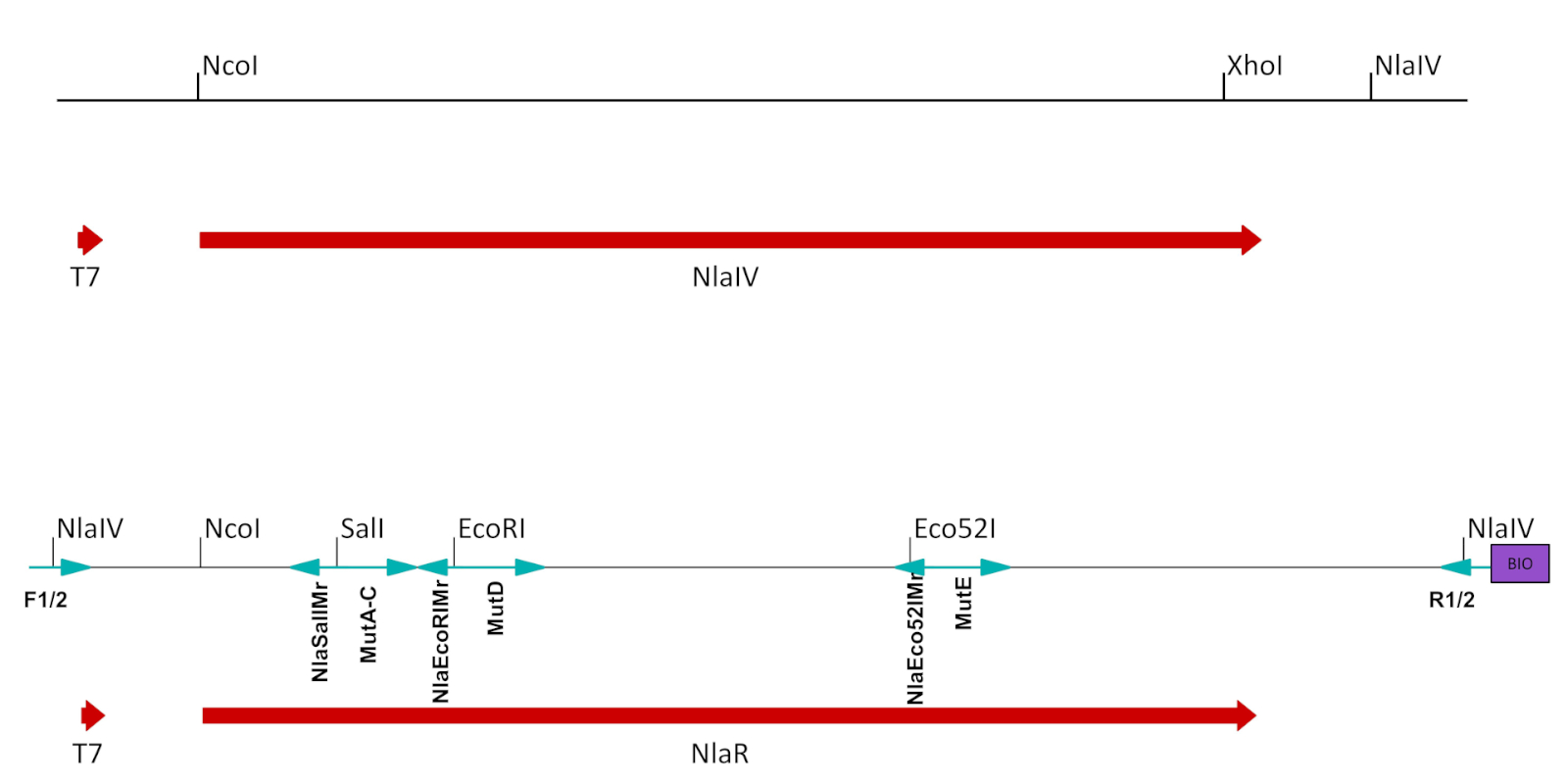
- T7 프로모터와 ORF 효소의 스톱 코돈 사이의 재조합 플라스미드의 단면도로부터 엔지니어링된 효소에 대한 모든 인식 부위를 제거하여 침묵하는 돌연변이를 도입하였다(도2,표 1A).
참고: 이러한 사이트를 두 개 이상 제거해야 하는 경우 여러 돌연변이 라운드가 필요합니다(단계 1.5.1-1.5.7).- 프라이머의 5' 끝에 도입된 설계 변형으로 전체 길이 플라스미드를 증폭시키는 내부 PCR 반응을 사용하십시오(표2A).
- PCR 반응의 50 μL에 DpnI 엔도뉴클레아제의 10 U를 첨가하여 템플릿 DNA를 제거하고, 37°C에서 2시간 동안 배양한다.
- 아가로즈 젤 전기 동에 의해 제품을 해결합니다. 전체 길이 플라스미드에 해당하는 밴드를 잘라 내고 상용 키트로 정화합니다.
- 10x 결찰 완충액(1x 농도)을 추가하고 ATP(1mMM)로 보충하십시오. T4 폴리뉴클레오티드 키나아제 10U를 넣고 37°C에서 20분 동안 배양합니다. 70°C에서 10분 동안 가열하여 효소를 비활성화합니다.
- PEG 4000을 5%까지 추가하고, ATP(1 mM)로 다시 보충하고, T4 DNA 리가제 5U를 추가합니다. 실온(RT)에서 2시간 동안 배양합니다.
- 코그네이트 메틸트랜스퍼라제를 운반하는 유능한 세균성 균주로 변신(1.1단계).
- 플라스미드 DNA를 소규모로 분리하고 디도옥시 시퀀싱에 의한 서열 변화의 도입을 확인한다.
- 올리고뉴클레오티드 유도 돌연변이 발생에 의해 표적화된 서열에 가까운 독특한 제한 부위를 소개한다(도2, 표 1B). 각 사이트에 대해 1.5.1-1.5.7 단계를 따릅니다.
참고: 이 단계는 표적 돌연변이 발생이 사용되는 경우에만 수행됩니다. 무작위 돌연변이 발생을 수행하는 경우, 단계를 건너 뛰고 섹션 3 대신 진행. 제시된 예제에서는 모든 사이트가 대상 영역의 업스트림에 도입되었지만 다운스트림에서도 도입될 수 있습니다. - ESC의 증폭을 위한 프라이머 설계(표 1C).
- 선택된 NlaIV 서열 외부에 결합하고 5' 끝에 바이오틴을 포함하는 선택된 인식 사이트(R1) 및 그 짧은 버전(R2)을 소개하는 엔도누클러 ORF의 역방향 프라이머 바인딩 다운스트림을 설계합니다(그림 1참조).
- T7 프로모터의 ESC 상류에 결합하는 전방 프라이머(F1)를 설계한다. 이 프라이머는 또한 본래 인식 서열의 역선택 된 변이체 (즉, 선택된 역열을 제외하고 본래 효소에 의해 인식된 최대 서열 변이)를 도입해야 한다.
참고: 이 프라이머(F2)의 더 짧은 버전은 선택적 PCR(단계 5.9)의 후반부에 사용될 것이다.
2. 뮤타게닉 프라이머의 분할 및 혼합 합성
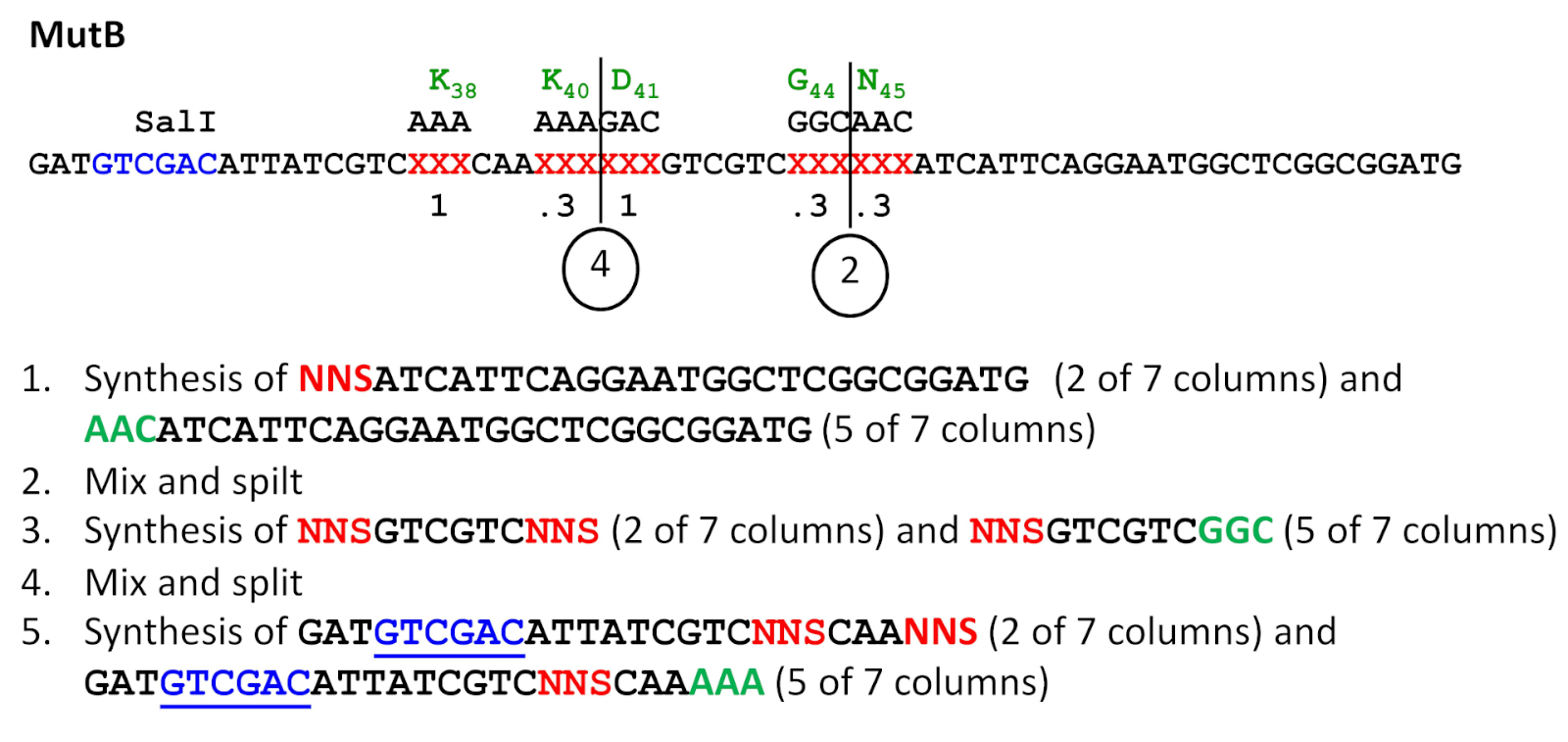
참고: 이 단계는 하나 이상의 사이트에서 하재 돌연변이 발생이 필요한 프로젝트에만 사용됩니다. 여러 합성 컬럼이 있는 신디사이저가 필요합니다. 돌연변이 발생 주파수에 따라 무작위 NNS 코돈 삼중항 및 야생형 코돈 삼중항의 합성을 위한 컬럼을 할당한다. 예를 들어, 7개의 동일한 부피 합성 컬럼을 사용할 수 있고, 0.3의 돌연변이 발생률이 주어진 부위에서 바람직한 경우, ~0.3 x 7 또는 2개의 컬럼에 무작위 NNS 코돈, 및 야생형 코돈은 ~0.7 x 7 또는 5개의 컬럼(그림3)에추가한다.
- 하부 포화 돌연변이 발생에 대한 사이트에 대해 결정합니다. 사이트의 가상적 중요성에 따라 돌연변이 발생 주파수를 선택하십시오(즉, 사이트가 더 중요할수록 빈도가 높을수록) 전체 라이브러리 복잡성에 대한 제한을 염두에 두어야 합니다(토론 참조).
- 모든 컬럼에서 올리고뉴클레오티드를 합성하고, 3'-말단으로부터 2차 하부 포화 돌연변이 발생 부위 바로 앞에 있는 삼중항까지. 마지막 합성 주기에서 5'-trityl 보호 그룹을 제거하지 마십시오 (신디사이저에 트리틸 온 옵션을 사용). 보호 기는 다음 합성 주기의 시작 부분에서 제거될 것이다(도 3의1단계).
- 합성 열을 엽니다. 제어 된 모공 유리 (CPG) 합성 지지서를 건조한 1.5 mL 튜브로 수집하고 소용돌이에 의해 혼합합니다. 혼합 된 CPG 수지를 새로운 합성 컬럼으로 재분할합니다. 전체 수율을 감소시킬 수 있으므로 습도를 도입하지 마십시오(그림 3의2단계 및 4단계).
- 합성을 계속, 하수 화 돌연변이 부위 삼중에서 시작. 원하는 돌연변이 발생 빈도에 따라 무작위 NNS 트리플렛 또는 야생 형 삼중항에 컬럼을 할당합니다(위 참고 참조). 추가의 하부 포화 부위가 존재하는 경우, 다음 하부 포화 돌연변이 발생 부위 이전의 삼중항으로만 진행한다. 다시 말하지만, 끝에 5'-trityl 그룹을 둡니다 (합성기에 5'-trityl-on 옵션) (그림 3의단계 3). 그런 다음 2.3 단계를 계속합니다.
- 더 이상 하층 사이트가 다운스트림에 존재하지 않는 경우, 합성을 완료하고, 5'-트리틸 그룹을 끝에 남겨두고(합성기상 5'-트리틸-온 옵션) (그림 3의5단계).
- 정제 카트리지 제조업체의 지침에 따라 올리고뉴클레오티드 라이브러리를 보호하고 정화합니다.
참고: CPG로부터의 탈보호에 의해 방출된 올리고뉴클레오티드는 또한 역상 고성능 액체 크로마토그래피(HPLC)에서 트리틸-온(manual trityl-on)과 수동 트리틸 그룹 제거(RT에서 80% 아세트산을 함유한 1시간 치료) 및 두 번째 HPLC 정제로 정제될 수 있다. - 우레아-PAGE 겔에서 올리고뉴클레오티드 라이브러리 품질을 확인한다.
3. 변형 라이브러리 생성
참고: 1.6단계에서 재조합 플라스미드를 사용한다.
-
올리고뉴클레오티드 지시 돌연변이 발생에 의해 라이브러리를 생성한다.
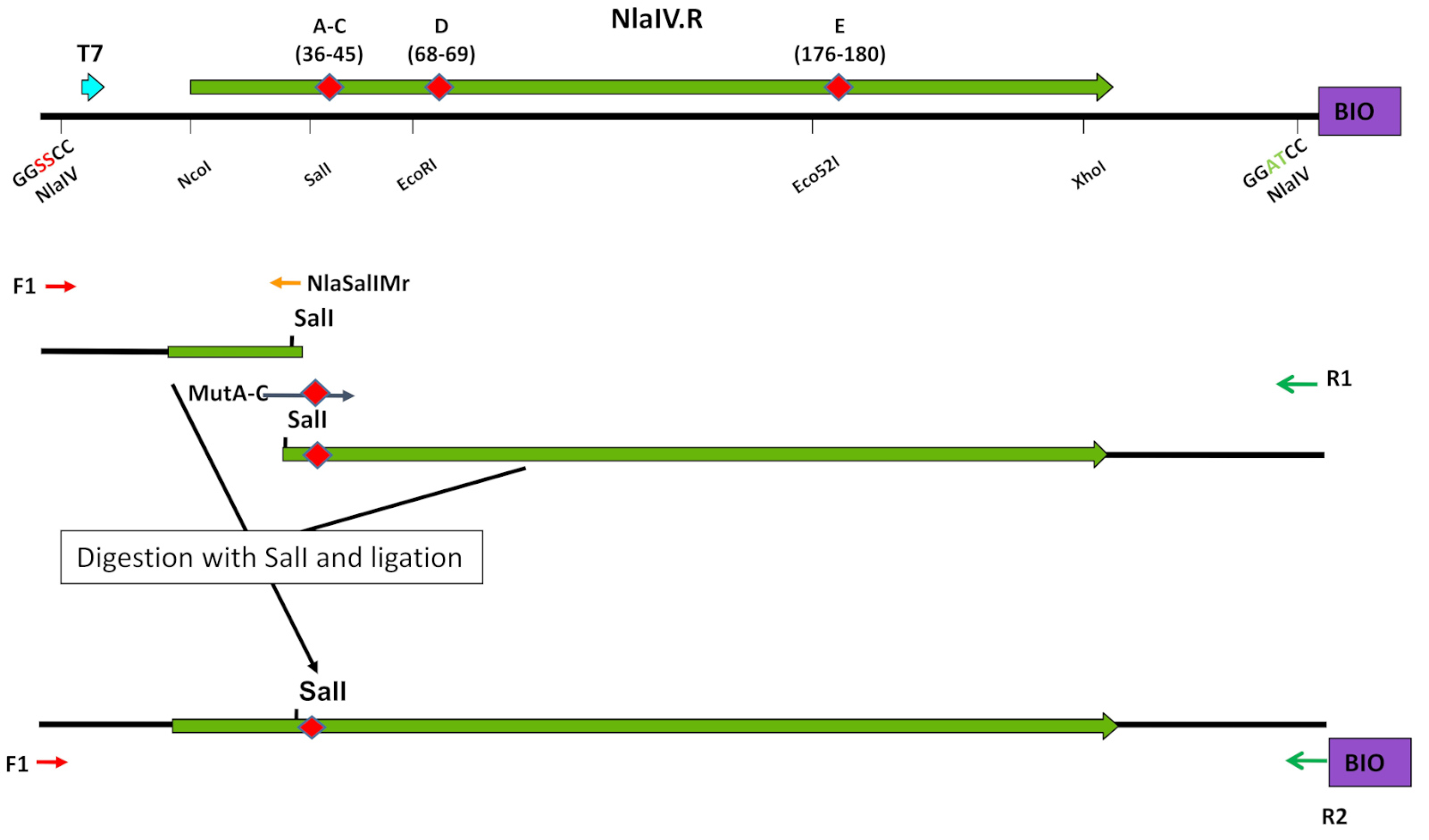
참고: 또는 EP-PCR 프로토콜(3.2단계)을 사용합니다.- T7 프로모터로부터 돌연변이 발생을 표적으로 하는 서열을 측면으로 하는 독특한 제한 효소 부위로 단면도(NlaIV: SalI, EcoRI, 또는 Eco52I)(표1B-C, 표 2B, 도 4). 독특한 제한 효소 부위로부터 ESC의 3' 말단까지 두 번째 부분을 증폭시다.
- PCR 반응의 5 μL(단계 3.1.1)을 8 μL의 물, 10x 제한 효소 완충액의 1.5 μL, 및 5 단위의 적절한 제한 효소(SalI, EcoRI, 또는 Eco52I)와 별도로 혼합하고 2시간 동안 적절한 온도에서 배양합니다.
- 아가로즈 겔 전기 동동을 사용하여 두 반응의 제품을 해결하십시오. 예상 크기 밴드를 잘라 상용 키트로 정화합니다.
- 아가로즈 젤에 정제 된 제품의 1/3까지 실행하고 밀도 측정에 의해 각 정제 밴드의 농도를 측정합니다.
- 1x 리가제 완충액과 1U의 T4 DNA 리가제와 함께 1:1 몰 비로 ESC의 두 부분의 결찰을 설정하고 RT에서 2시간 동안 배양하였다.
- 아가로즈 젤의 반응 제품을 해결하십시오. 예상 되는 크기의 제품을 잘라 하 고 상용 키트와 함께 정화.
- 프라이머 F1 및 R2(표 1C 및 표 2A)로PCR 반응에서 정제 된 결찰 제품을 증폭시다. 20회 이상의 증폭 주기를 실행하지 마십시오.
- 아가로즈 겔에서 PCR 반응을 분별. 제품을 잘라 상용 키트로 정화하십시오.
- 아가로즈 겔의 이전 단계에서 정제된 라이브러리의 5 μL aliquot를 실행하고 밀도 측정에 의해 농도를 측정하였다.
- 라이브러리의 작은 샘플을 복제(최대 5 μL) 및 서열 >15 클론돌연변이 빈도 및 분포를 확인한다(표3). 4단계로 진행합니다.
참고: 또는 ESC의 작은 샘플의 높은 처리량 시퀀싱을 사용할 수 있습니다.
-
EP-PCR을 수행합니다.
- 프라이머 F1 및 R1로 1.5.7 단계에서 얻은 플라스미드로부터 ESC를 증폭시다. Taq I 폴리머라제(표1B)로20사이클을 실행합니다.
- 젤은 PCR 제품을 정화합니다.
- 이전 단계에서 정제 된 PCR 제품의 2 ng로 EP-PCR을 설정하고 F1 및 R1 프라이머로 EP-PCR(표 1C)의15 사이클을 실행합니다.
- 젤은 제품을 정화하고 젤 밀도 측정에 의해 정량화.
참고: 정제 된 EP-PCR 제품의 낮은 농도로 인해 정량화를 위해 약 1/3을 사용합니다. - 라이브러리의 작은 샘플을 복제 (최대 1/5) 및 시퀀스 >15 클론 돌연변이 주파수 및 분포를 확인하려면(표 4).
참고: 또는 ESC의 작은 샘플에 대한 고처리량 시퀀싱을 수행합니다.
4. 생체외 전사-번역 반응 구획화 수행
-
시험 엔도누아제 발현 및 체외 전사-번역에서의 효소 활성.
- 분자의 중심에 위치한 엔도누클로즈에 대한 단일 인식 부위를 사용하여 짧은(200-500 bp) 기판을 준비하여 절단 반응을 쉽게 검출할 수 있도록 한다.
참고: 기판을 준비하는 가장 쉬운 방법은 임의의 DNA 분자의 적절한 단편의 PCR 증폭이다. 기판은 절단 검출을 단순화하기 위해 방사성 표지 또는 형광 표지될 수 있습니다. - 제조업체의 권고에 따라 0.5 μg의 야생 형 ESC로 전사 번역 반응 50 μL을 설정합니다. 마그네슘 염 (MgCl2,MgSO4및 마그네슘 아세테이트를 테스트 할 수 있음)을 1.5 mM에 추가하고 이전 단계에서 적절한 양의 기판을 추가하십시오 (라벨이 없는 DNA의 경우 적어도 0.5 μg).
참고: 마그네슘에 의해 활성화된 뉴클레아제가 함유되어 있지 않은 전사/번역 키트를 사용할 수 있습니다. 일부 키트 공급 업체는 생산 중 DNA 오염을 제거 한 다음 핵 억제제로 킬레이터를 추가하기 위해 뉴클레아제를 사용합니다. 이러한 키트는 이 방법과 호환되지 않습니다. - 제조업체의 지침에 따라 전사-번역 반응을 배양합니다. 이어서 반응 혼합물을 2시간 동안 제한 효소에 대한 최적 온도로 이월한다.
- 아가로즈 겔에서 기판의 절단을 분석하고 적절한 검출(예를 들어, DNA 염색, 형광 시각화, 또는 자가방사선 촬영)(그림5).
참고: 구획화를 진행하기 전에 기판의 적어도 부분절단이 필요하다. 이것이 달성되지 않으면 마그네슘 화학 적 형태 또는 농도를 추가로 최적화해야합니다.
- 분자의 중심에 위치한 엔도누클로즈에 대한 단일 인식 부위를 사용하여 짧은(200-500 bp) 기판을 준비하여 절단 반응을 쉽게 검출할 수 있도록 한다.
- 15 mL 원뿔 형 튜브에 225 μL의 스팬 80 및 25 μL의 트웬 80 ~ 5 mL의 미네랄 오일을 추가하여 오일 계면 활성제 혼합물을 준비합니다. 튜브를 15배 부드럽게 뒤집어 완전히 섞으세요.
- 각 라이브러리에 대해 오일 계면활성제 혼합물의 950 μL을 2 mL 둥근 바닥 극저온 바이알로 옮기고, 라이브러리 명칭을 가진 라벨을 얼음으로 옮김을 전송한다. 작은 원통형 교반 막대 1개(5 x 2mm2)를각 바이알에 넣습니다.
- 제조업체의 제안에 따라 시험관 내 전사-번역 반응 혼합물(각 라이브러리에 대해 50 μL)을 준비한다. 염화마그네슘으로 혼합물을 1.5 mM의 최종 농도로 보충하십시오(4.1.4단계 참조).
- 얼음에 1.5 mL 튜브에 50 μL 알리쿼트분배.
- 얼음에 반응 혼합물에 (섹션 3에서) 라이브러리의 1.7 fmole을 추가합니다.
참고: 선택 효율성을 위해 더 많은 양의 식 라이브러리를 사용하지 마십시오. 하나 이상의 DNA 분자를 포함하는 수성 액적의 빈도를 최소화하는 것이 중요합니다. -
각 라이브러리에 대해 연속적으로 물-오일 에멀젼을 준비합니다.
- 1,150rpm으로 설정된 교반 속도로 자석 교반기위에 얼음으로 채워진 작은 비커(또는 큰 병 컵)를 놓습니다.
- 950 μL의 오일 계면활성제 혼합물과 작은 교반 막대로 극저온 바이알을 4.3 단계에서 자기 교반기의 얼음 차가운 비커로 옮니다. 교반 막대가 회전하고 있는지 확인합니다.
- 시험관 내 라이브러리-전사-번역 혼합물의 5개의 10 μL aliquots를 30 s 간격으로 2분 동안 추가하고 추가1분 동안 계속 교반한다. 에멀젼으로 바이알을 얼음 용기에 옮김을 옮김. 4.7.2단계부터 다음 라이브러리를 진행합니다.
- 모든 라이브러리가 처리된 후 키트 제조업체의 권장 사항에 따라 모든 라이브러리의 인큐베이션을 시작합니다.
- 바이알을 추가로 2시간 동안 엔지니어링된 엔도누펄스에 최적 온도로 옮은 다음 적어도 10분 동안 얼음에 넣습니다.
5. 도서관 및 선택의 지속적인 처리
- 극저온 바이알에서 차가운 1.5 mL 튜브로 에멀젼을 옮기고 0.5 M EDTA 의 1 μL을 추가하고 실온에서 5 분 동안 13,000 x g에서 원심 분리합니다.
- 파이펫으로 상부 오일 위상을 제거합니다. 오일-물 중간 위상이 보이지 않는 경우, 수성 상을 동결하기 위해 -20 °C에서 적어도 5 분 동안 튜브를 인큐베이션 한 다음 즉시 액체 오일 상을 피펫 아웃시합니다.
- 100 μL의 100 μL을 추가 10mMTris HCl pH 8.0 즉시 페놀의 150 μL로 추출을 수행 : 클로로 포름 (1 : 1 v / v) 짧은 소용돌이에 의해 다음 13,000 x g에서30 s 원심 분리에 의해 위상 분리. 상부 수성 단계를 수집합니다.
- 3 M 아세테이트 나트륨 (pH = 5.2), 글리코겐 2.5-5 μg 및 에탄올 2.5 vol (375 μL)의 0.1 vol (15 μL)을 첨가하여 DNA를 침전시. -20°C에서 1시간 동안 배양하고 원심분리기는 13,000 x g,4°C에서 15분 동안 배양한다. 상급제를 버리고 차가운 70 % 에탄올 1 mL로 펠릿을 간략하게 씻으십시오.
- DNA/글리코겐 펠릿을 스피드백으로 건조시키거나 공기 건조시 >5분 동안 건조시키세요.
- 펠릿을 10 mM Tris-HCl (pH = 7.5)의 50 μL에 용해시고. 제조업체의 지침에 따라 제조된 스트렙타비딘 자기 비드 5 μL을 추가하고 RT에서 1시간 동안 혼합하고, 바람직하게는 회전목마 믹서또는 부드러운 소용돌이에 의해 혼합합니다.
- 마그네틱 스탠드에 구슬을 분리하고 비오틴없이 DNA에 농축 된 액체를 수집합니다.
- 에탄올 강수량에 의해 DNA를 집중시다(단계 5.4-5.5).
- 이전 단계에서 농축된 DNA를 5 μL의 물에 용해시키고 F2 및 R1 프라이머(표1A)로PCR 반응에서 템플릿으로 사용한다.
참고: 템플릿 오염 문제를 방지하고 PCR 아티팩트를 최소화하려면 Taq 폴리머라제(Pfu 또는 Phusion 아님)를 사용하고 템플릿 크기에 비례하는 연장 시간으로 18-20사이클을 실행합니다(1kb = 1분)(표 2B참조). - 아가로즈 젤에 PCR 제품을 분별하고 예상 크기 제품을 잘라냅니다. 일부 번짐은 크기가 다른 제품이 있음을 나타냅니다(그림 6참조). 상용 키트로 젤 슬래브에서 DNA를 정화합니다.
- 단계 5.10 및 프라이머 F1 및 R2로부터 5.9 단계에서와 동일한 프로토콜을 사용하여 최대 50 ng의 DNA로 두 번째 PCR 반응을 실행합니다. 5.10에 기재된 바와 같이 제품 정제를 진행한다. 아가로즈 겔 치석 측정법(UV 분광법 아님)에 의해 정량화한 후 정제된 DNA는 시험관내 선택의 다음 라운드(단계 4.6)에서 사용될 수 있다.
6. 변경 된 시퀀스 특이성에 대한 화면 변형
- 선택한 변형을 복제합니다.
- 5.10 단계에서 2 시간 동안 10 U의 제한 효소를 발현 벡터 (NlaIV : NcoI 및 XhoI의 경우)로 복제하는 데 적합한 제한 효소를 효소 공급 업체가 권장하는 온도 및 완충액으로 소화합니다. 아가로즈 겔 전기 동동으로 제품을 해결하고 예상 크기 조각을 분리합니다.
- 6.1.1단계에서와 동일한 효소로 이중 절단에 의해 플라스미드 벡터(예를 들어, pET28)를 준비하고, 겔은 상용 DNA 겔 정제 키트로 제품을 정제한다.
- 아가로즈 겔 전기 동동을 가진 고밀도 측정에 의하여 벡터 및 삽입의 농도를 추정한다.
- T4 DNA 리가제 및 벡터의 1-5 U로 결찰을 설정:효소 공급 업체가 권장하는 1x 리가아완충에 몰 비 1:3-1:5를 삽입합니다. RT에서 2시간 동안 배양하고 형형파 또는 전기포공(12)에12의해 적절한 숙주 박테리아(단계 1.3으로부터)를 도입한다.
- 적절한 항생제 (pET28 또는 pET30 벡터에 대한 카나마이신의 50 μg / mL)와 1 % 포도당을 포함하는 LB 플레이트에서 분변제를 선택하십시오.
- 단백질 변이체를 표현합니다.
- 카나마이신 (50 μg / mL)과 1 % 포도당과 LB의 2 mL로 변환 (최대 24 클론이 한 번의 실행으로 쉽게 처리 될 수 있음)에서 단일 식민지를 접종하고 흔들림과 함께 37 °C에서 하룻밤 성장합니다.
- 15 mL의 따뜻한 (37°C) LB를 100 μg카나마이신을 함유하고 0.75 mL의 포도당을 함유하고 격렬한 흔들림으로 37°C에서 배양한다.
참고: 50 mL 원심 분리튜브 또는 100 mL 에를렌마이어 플라스크를 사용할 수 있습니다. - 1mL의 글리세롤을 하룻밤 배양액에 176 μL 추가 (글리세롤의 최종 농도 = 15%) 완전히 혼합하고 -70 °C에서 동결.
- 후 2-3 h 보충 15 배양 (단계 6.2.1) IPTG와 1 mM 및 추가에 대 한 배양 5 시간.
- 원심분리(10,000 x g,4°C, 10분)로 세균 펠릿을 수집하고 -70°C에서 동결한다.
- 단백질 변이체를 정화합니다.
- 20 μL의 니켈 친화성 수지 현탁액을 넓은 보어 피펫 팁으로 1.5 mL 튜브에 B1 버퍼의 200 μL로 옮기고 부드럽게 혼합하고 원심 분리기 (5,000 x g,30 s, 4 °C). 피펫팅으로 상급체를 제거하고 튜브를 얼음 위에 둡니다.
- 활발한 소용돌이에 의해 B1의 300 μL에서 6.2.5 단계에서 세균 펠릿을 다시 중단합니다. 현탁액을 1.5mL 튜브로 옮김.
- B1에 100x 프로테아제 억제제 칵테일과 리소좀 용액 3 μL을 추가합니다(최종 농도 1 mg/mL). 팁 장착 프로브로 초음파 처리하여 세포를 분해합니다. 샘플당 6개의 10초 버스트를 사용하며 그 사이에 얼음에서 >15s 팁 냉각 시간을 사용하십시오. 항상 얼음에 세포 현탁액을 유지합니다.
- 펠렛 세포 파편은 원심분리(2분, 12,000 x g,4°C)에 의해 상급의 250 μL을 단계 6.3.1로부터 수지 알리쿼트로 이송한다.
- 차가운 방에서 15 분 동안 혼합, 바람직회전식 또는 부드러운 소용돌이에 의해.
- 원심분리기 (5,000 x g,30 s, 4 °C)를 피펫으로 상수부 흡입한다.
- 500 μL의 W 버퍼를 추가하고 수지의 부드럽게 다시 일시 중단합니다. 원심분리기 (5,000 x g,30 s, 4 °C)를 피펫으로 상수부 흡입한다.
- 6.3.7 단계를 반복합니다.
- 완충제 E 20 μL을 추가하고 수지를 부드럽게 다시 일시 중단하고 샘플을 2-5 분 동안 얼음에 두십시오. 원심 분리기 (5,000 x g,30 s, 4 °C)를 초월체를 수집합니다.
- 6.3.9단계를 반복합니다. 수영장 슈퍼 나타.
- SDS-PAGE(5-10 μL)로 단백질 샘플을분석합니다(그림 7).
- 변경된 특이성을 가진 변형을 위해 화면.
- 박테리오파지 람다 DNA에 대한 분석 분열 활동. 단백질 샘플은 최종 반응 량의 최대 10%를 구성할 수 있다. DNA의 0.5 μg 당 단백질 샘플의 총 2 μL 및 2 시간 반응 시간은 좋은 출발점이다.
- 야생형 효소에 의해 생성된 생성물과 함께 아가로즈 겔 전기동공에 의한 반응 생성물을 분석한다. 추가 분석을 위해 야생형 효소에 의해 생성된 것과 명확하게 구별할 수 있는 분열 패턴을 생성하는 클론을 선택한다(도8).
결과
이 프로토콜은 두 가지 원치 않는 클래스를 고갈시켜 엔지니어링 된 REase의 원하는 변이체의 빈도를 증가시키는 도구일 뿐입니다: 변하지 않는 야생 형 서열 특이성을 가진 비활성 효소 및 endonucleases. 한편, REase 특이성을 변경하는 것은 매우 어렵기 때문에, 24개의 클론의 단일 스크리닝에서 야생형 효소와 는 다른 분열 패턴을 생성하는 하나의 변이체를 찾는 것이 성공으로 간주되어야 한다. 우리의 손에 최고의 화면은 유망한 변종의 최대 20 %를 식별 할 수 있습니다(그림 8A).
긍정적 인 결과는 라이브러리 품질 (즉, 대체 및 임의 분포의 제한된 빈도) 및 라이브러리 구성원의 생체 생물 집단의 효율적인 캡처 (단계 3.6-3.7)에 크게 의존합니다. 두 가지 문제를 모두 감지할 수 있습니다. 라이브러리 품질은 가능한 한 많은 클론(>15)을 시퀀싱하거나 높은 처리량 시퀀싱을 통해 라이브러리의 직접 시퀀싱을 통해 선택하기 전에 확인해야 합니다(단계 3.10, 표 3). 선택한 클론의 대다수가 활성화되지 않은 경우, 이것은 스트렙타비딘 캡처 선택의 실패의 명확한 표시이다. 이러한 라이브러리는 스트렙타비딘 캡처 선택 단계를 벗어난 비활성 변이체에 의해 대부분 지배되기 때문에 많은 선택 주기를 거치는 라이브러리의 경우 유사한 효과가 관찰됩니다(그림8B). 따라서 모든 선택 주기 후에 스크리닝을 실행하고 선택 반복에 의존하지 않고 수동으로 선택한 유망한 변형을 개발하는 것이 좋습니다.

도 1: NlaIV 엔지니어링에 기초한 새로운 서열 특이성의 시험관 내 선택. (a)발현/선택 카세트(ESC)의 조직은 REase에 대한 2개의 인식 사이트, 1) 우측 끝에 가까운 선택된 서열(GGATCC)과 2) 카운터 선택서열(GGSSCC)을 왼쪽 끝에 가까운, 뿐만 아니라 T7p 및 T7t-T7 프로모터 및 T7 터미네이터를 포함한다. 프라이머 바인딩 사이트는 다음과 같습니다. 야생 유형및 선택된 NlaIV 변이체에 의한 분열은 각각 빨간색및 녹색 삼각형으로 도시된다. (B)선택 주기 단계: I) 전사-번역-분열 반응의 유화는 ESC 라이브러리와 혼합; II) 모든 생체 활성 DNA는 스트렙타비딘으로 코팅된 자기 입자에 포착되어 제거되어 비활성 변이체를 인코딩하지 않습니다. III) 야생형 활성(즉, GGSSCC 서열을 절단할 수 있는 것)을 가진 REase를 인코딩하는 ESCs는 순방향 및 역방향 프라이머에 대한 결합 부위를 분리하기 때문에 제거된다. 따라서 이러한 ESC의 증폭이 발생하지 않습니다. IV) 다음 선택 라운드에 대한 입력은 오른쪽 끝에 비오틴을 첨가하고 왼쪽 끝에 있는 카운터 선택 서열의 변형을 재도입하여 생성된다. 엘스비어의 허가를 받아 Czapinska 등8에서 전재. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 2: ESC의 준비. T7 프로모터의 제어하에 NlaIV ORF를 포함하는 발현 벡터에서 본래 구문으로부터 유래된 프래그먼트는 발현/선택에 적합하도록 수정되었다. NlaIV ORF로부터의 NlaIV 사이트 다운스트림은 제거되었고 선택된 위치를 돌연변이화하는 데 사용된 고유 사이트(SalI, EcoRI 및 Eco52I)는 NlaIV ORF에서 침묵하는 돌연변이로 도입되었다. 최종 구문은 왼쪽에 있는 카운터 선택 시퀀스(GGSSCC)와 오른쪽에 선택된 시퀀스(GGATCC)의 두 개의 측면 NlaIV 사이트를 도입한 측면 프라이머로 증폭되었습니다. 역프라이머는 또한 비오틴을 도입했다. 돌연변이 된 ECS의 생성에 사용되는 프라이머는 파란색 화살표로 표시되고 아래에 레이블이 지정됩니다 (표 1B, C참조). 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3: 분할 및 혼합 합성 방식. 이 예는 NNS 서열이 4위치에서 0.8 주파수로 도입된 MutB 프라이머 합성을 참조한다(표 3참조). 화학 합성은 3'에서 5'까지 수행되지만 모든 서열은 정식 5'-3' 방향으로 표시됩니다(즉, 이 방식에서 오른쪽에서 왼쪽으로 진행됨). NNS 돌연변이 성 서열이 빨간색인 동안 돌연변이 위치에서 야생 형 서열은 녹색으로 표시됩니다. 나중에 ESC에서 돌연변이를 소개하는 데 사용되는 SalI 인식 사이트는 밑줄이 그어져 있습니다. 혼합 및 분할 단계(2 및 4)의 점이 표시됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 4: 올리고뉴클레오티드 표적 돌연변이 발생에서 독특한 제한 효소 부위의 사용. 돌연변이 도입의 전략은 라이브러리 A-C의 구성의 예에 도시되어 있다(단계 3.1-3.7 참조). 엘스비어의 허가를 받아 Czapinska 등8에서 전재. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

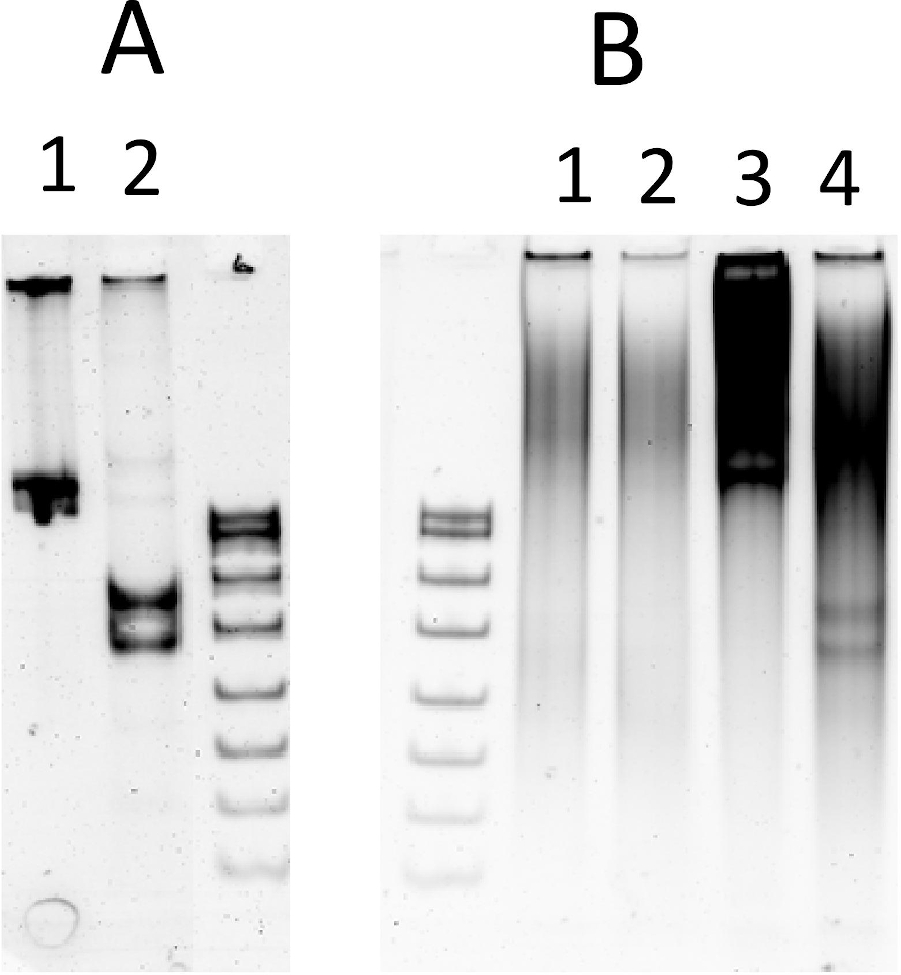
도 5: 체외 전사-번역에서 내핵성 절단. (a)최적의 REase 버퍼에서 시험 기판의 절단: 1) 기판, 단일 NlaIV 인식 사이트를 가진 612 bp PCR 제품; 2) 분열 제품, 355 bp 및 257 bp.(B)체외 전사 번역 반응에서 분열 (ESC의 0.5 μg을 포함): 1-2) 15 μL 기질 없이 시험관 내 전사 번역의 (라인 2: 1.5 mM MgCl2로보충 된 반응); 3-4) 시험기기의 1μg을 가진 시험관내 전사-번역의 15 μL aliquots; (라인 4: 반응 1.5 mM MgCl2로보충). S-DNA 크기 마커 (pBR322 는 MSPI로 소화). 샘플은 6% 네이티브 PAGE에서 해결되었습니다. DNA를 에티듐 브로마이드로 염색시켰다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

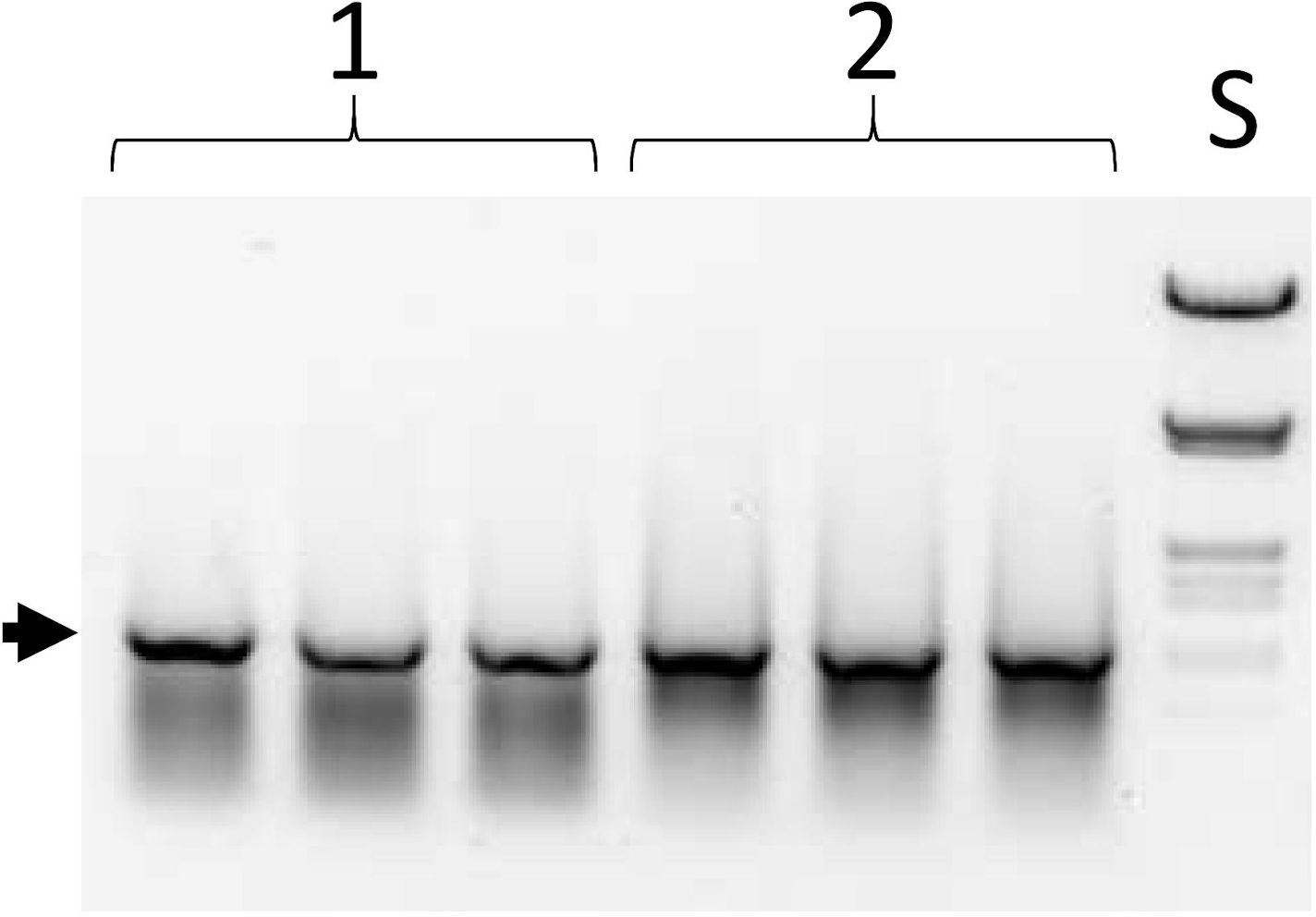
그림 6: 선택 주기에서 첫 번째 PCR의 제품입니다. 도 1B,단계 III; 프로토콜 단계 5.10. 열 집합 1과 2는 세 배로 로드된 두 개의 서로 다른 라이브러리의 aliquot입니다. S-DNA 크기 표준 (람다 DNA힌드III 및 EcoRI로 소화). 화살표는 전체 길이 ESC(1,050bp)의 위치를 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

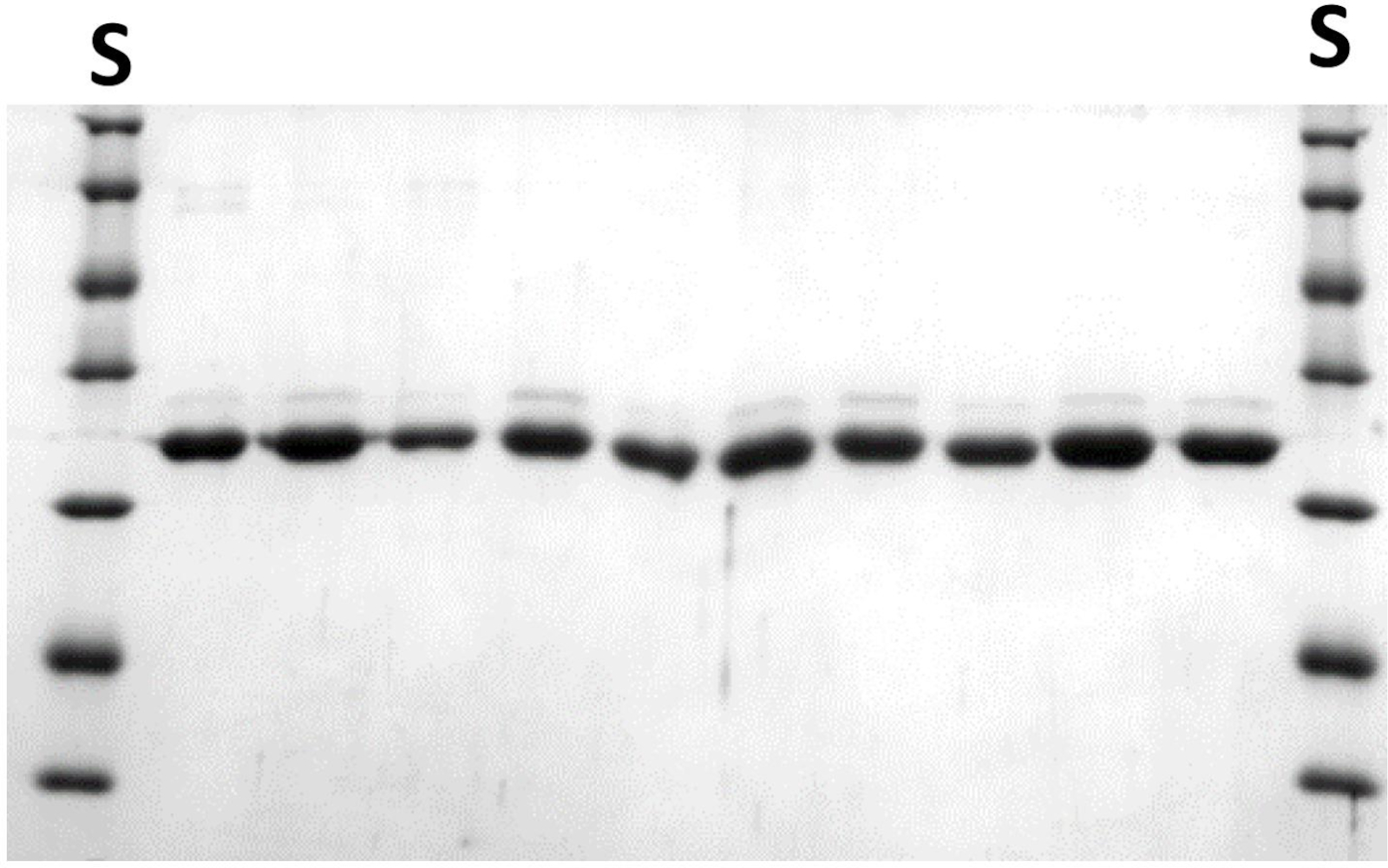
도 7: 미니 스케일에서 추가 스크리닝을 위해 정제된 NlaIV 변이체. 6.3.11 단계를 참조하십시오. 각 줄에는 다른 변이체의 10 μL aliquot가 포함되어 있습니다. S-단백질 분자량 표준. NaIV REase 소단위의 분자 질량은 29.9 kDa입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

도 8: 서열 특이성 변경을 위한 NlaIV 변이체의 스크리닝의 예. 6.4.2 단계를 참조하십시오. (A)유망한 변종의 높은 빈도로 성공적인 선별. S=DNA 크기 마커, 람다 DNA가 힌드III 및 에코리(EcoRI)로 갈라진; 야생형(wt) = 람다 DNA가 야생형 NlaIV로 갈라진; λ = 람다 DNA 기판, 갈라지지 않음; 활동이 매우 낮은 다른 열=변형. 변형은 레이블이 지정됩니다! = 야생형 효소와 구별되는 분열 패턴을 생성하는 유망한 변이체; ? = 시퀀스 기본 설정을 변경했을 수도 있는 변형입니다. (B)실패 한 선별, 변이체의 대다수비활성 및 명백하게 변경되지 않은 절단 패턴을 가진 1개의 변종. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

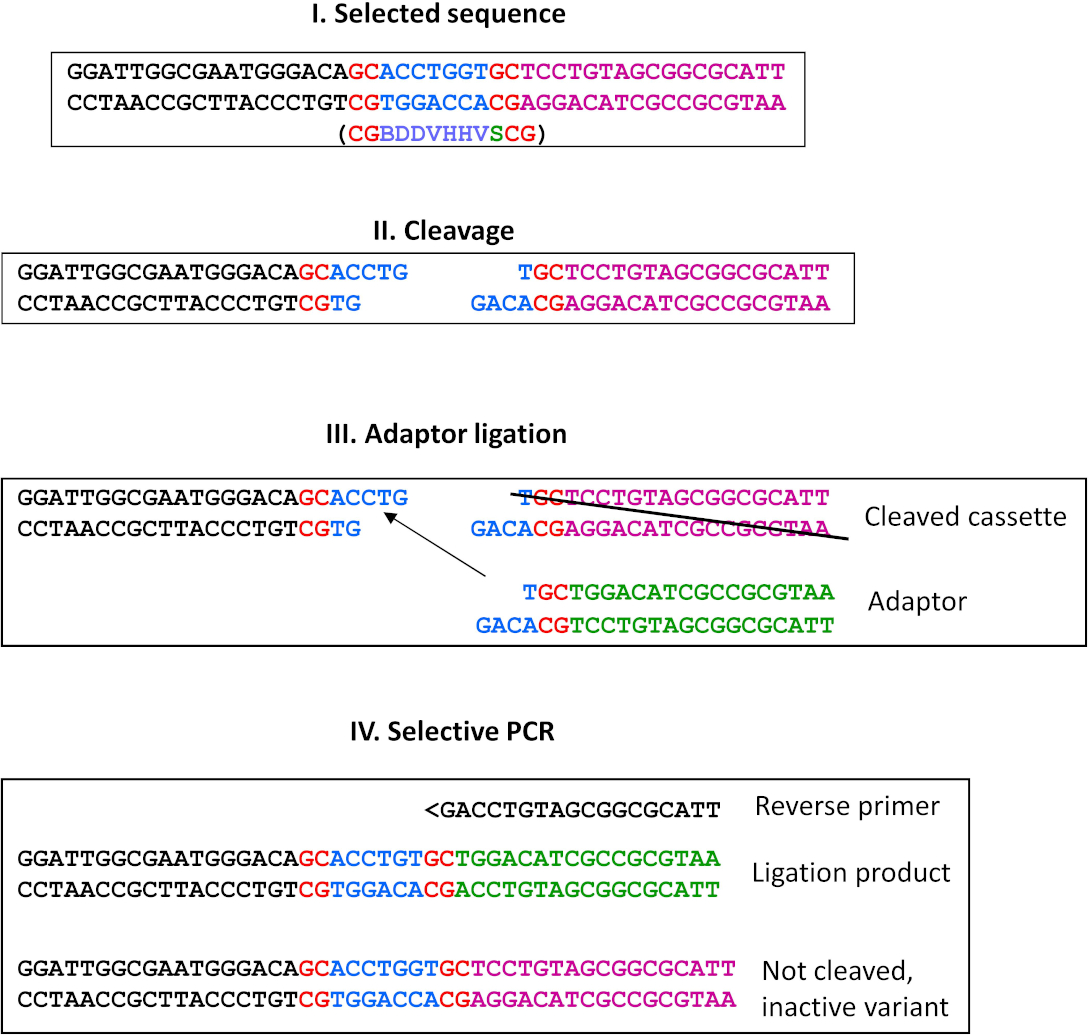
그림 9: 결찰에 의한 대체 선택. 이 대안은 끈적끈적한 끝을 생성하는 모든 REases에 사용할 수 있습니다. 여기서 우리는 MwoI 효소(미공개)에 대한 선택 계획에 대한 예시 프로토콜을 제시한다. I) 선택된 시퀀스(ESC의 오른쪽 끝에 위치)와 정의된 잔기가 빨간색으로 표시되고 청색으로 표시된 코그네이트 서열의 선택된 변형. ESC의 왼쪽 끝에 배치할 카운터 선택 시퀀스 아래의 괄호안에 도시된; II) MwoI 분열의 제품; III) 시험관 내 전사/번역을 종료한 후, 제품이 정제되고 과잉 어댑터로 결찰이 수행됩니다. 선택된 순서로 절단된 분열 산물만이 결찰에 참여할 수 있다. 따라서 비활성 변이체가 제거되고 풀다운 단계가 필요하지 않습니다. 카운터 선택 서열의 절단 생성물(ESC의 왼쪽 끝에, 도시되지 않음)은 어댑터의 돌출된 단부가 카운터 선택 서열에 상보적이지 않기 때문에 이 결찰에 참여할 수 없다; IV) 선택적 PCR은 메인 프로토콜에서와 동일한 전략을 사용하여 야생형 퇴화 서열 특이성(F1 프라이머 결합이 카운터 선택부위에 대한 말단)을 가진 변이체를 제거하는 반면, 비활성 변이체는 삼촌에 결합할 수 없는 선택적 역프라이머에 의해 제거된다(따라서 어댑터 결찰에 의해 수정되지 않음) 오른쪽 끝. 다음 사이클에서 공정은 이전 단계의 분열 생성물(즉, 패널 III의 "절단 카세트") 및 적절한 선택적 역프라이머와 동일한 어댑터를 사용하여 반복될 수 있다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
표 1: NlaIV 엔지니어링에 사용되는 프라이머. 주석에 언급 된 제한 사이트의 시퀀스는 밑줄이 그어져 있습니다. 작은 글자들은 DNA 템플릿에 보완이 없는 서열을 나타냅니다. 이 표를 보려면 여기를 클릭하십시오 (다운로드 오른쪽 단추로 클릭하십시오).
표 2: 프로토콜에서 사용되는 PCR 반응의 조건. Tm = 프라이머 용융 온도(프라이머에 대해 Tm이 다른 경우, 낮은 Tm을 사용해야 한다). 이 표를 보려면 여기를 클릭하십시오 (다운로드 오른쪽 단추로 클릭하십시오).
표 3: 분할 및 혼합 전략으로 합성된 두 개의 돌연변이원 프라이머의 품질 검사 결과. 돌연변이 코돈은 [XXX]로 표시됩니다. 인덱스 수가 낮을수록 인코딩된 아미노산의 위치를 나타냅니다. 엘스비어의 허가를 받아 Czapinska 등8에서 적응. 이 표를 보려면 여기를 클릭하십시오 (다운로드 오른쪽 단추로 클릭하십시오).
표 4: EP-PCR의 결과. ECS의 22개 클론의 서열 분석에서 파생된 주요 파라미터. 이 표를 보려면 여기를 클릭하십시오 (다운로드 오른쪽 단추로 클릭하십시오).
토론
여기서 설명된 선택 프로토콜은 NlaIV8,중앙 NN 염기를 가진 팔린드로믹 표적 부위를 인식하고 NN 염기 들 사이의 무딘 단부 절단을 촉매하는 디메릭 PD-(D/E)XK 접힌 인식 서열에 대해 시험되었다. NlaIV는 NN 기지 사이 분열이 이 기지가 복합체에 있는 단백질에 가깝다는 것을 건의하기 때문에 선택되었습니다. 원칙적으로, 프로토콜은 임의의 배그룹 중 임의의 임의의 계열 특이적 제한 엔도누클리스, 단일머티즘 또는 디메릭, 촉매 및 특이성 도메인이 일치하는지 여부(NlaIV 예에서와 같이) 또는 별도의(예를 들어, FokI)에 관계없이 임의의 스태거의 이중 가닥 브레이크를 촉매하는 임의의 서열에 사용될 수 있다. 더욱이, 원칙적으로 프로토콜은 새롭고 더 좁은 효소 특이성의 생성뿐만 아니라, 스타 활동을 제거하거나 높은 충실도 endonucleases를 만드는 데 사용될 수 있습니다. 그러나 이 모든 것은 아직 테스트되지 않았습니다. 특히, 스타 활성의 표적 제거는 동일한 아미노산 잔기가 원하는 및 바람직하지 않은 염기와 결합하는 데 관여할 수 있기 때문에 복잡할 수 있다. 이 프로토콜에 기술된 시험관내 단계는 좁아진 특이성의 선택에 한정되지 않고 달리 변경된 특이성을 선택하는 데 사용될 수 있다. 그러나, 변이체 endonucleases에 문제가 있습니다: 기판의 스펙트럼이 부모 endonuclease에 의해 절제되지 않은 새로운 표적을 포함하는 경우에, 일반적으로 이 활동의 유해한 효력에서 세포를 보호하는 좋은 방법은 없습니다. 대조적으로, endonuclease 특이성이 단지 좁혀지면, 표적은 야생형 표적의 서브세트이고, 따라서 이미 이용 가능한 코그네이트 메틸트랜스퍼라제는 완전히 보호되어야 한다.
우리의 프로토콜은 여러 가지 지향 진화 프로토콜에서 여러 가지 면에서 다릅니다. 개방형 판독 프레임 다양성은 모든 반복이 아니라 실험 시작 시 한 번 생성됩니다. 또한, 그것은 분할 및 혼합 합성에 의해 생성, 보다는 EP-PCR에 의해. 이 작업에서 사용되는 바와 같이, 코돈의 NNS 치환의 경우, 6개의6 위치에 대해 (4 x 4 x 2) 6~1.07 x 109조합이 있다. 따라서, 임의의 주어진 변이체는 ESC의 1.7 fmoles에서 평균적으로 한 번 존재한다. 이 용량은 Glen Research에서 제공하는 20개의 삼중뉴클레오티드 전구체의 혼합물과 합성을 사용하거나 분할 및 혼합 올리고뉴클레오티드 합성을 가진 덜 유망한 위치에서 돌연변이 빈도를 감소시킴으로써 7개의 위치로 증가될 수 있습니다. 가능하면 변형 범위를 6위치로 제한하는 것이 좋습니다. 명백하게, 이러한 돌연변이 유발 표적화는 기질 결합에 관여하는 REase의 적어도 영역에 대한 일부 기존 지식을 필요로 한다. 다양성을 생성하는 분할 및 혼합 프로토콜은 EP-PCR에 비해 분명한 장점이 있습니다. EP-PCR을 사용하여, 우리는 동일한 EP-PCR에서 NlaIV ESC에 대한 8개의 치환을 운반하는 변경되지 않은 변이체 및 시퀀스를얻었다(표 4). EP-PCR의 라이브러리는 피해야 할 클론의 실질적인 분획을 포함합니다(야생 형 서열, 다중 치환, 프레임 시프트 및 넌센스 돌연변이, 및 서열 특이성에 영향을 미치지 않는 장소에서의 돌연변이).
또한 당사의 프로토콜은 두 가지 순차적 선택 단계가 있음에 따라 다른 많은 지향 진화 프로토콜과도 다릅니다. 양수 선택은 원하는 활성이 유지되고, 그렇지 않으면 비오틴 태그가 제거되지 않고, 코딩 시퀀스를 풀다운하여 제거할 수 있는지 확인합니다. 기술적으로 는 소설, 비 중첩 특이성 (예를 들어, GCATGC)의 운이 출현은 적절한 분열 부위가 원하는 분열 근처에 존재하는 경우, 뿐만 아니라 비오틴 태그의 절단으로 이어질 수 있지만, 다른 곳에서는. 그러나, 이것은 매우 가능성이 있을 것 이라고. 음수 선택은 여전히 바람직하지 않은 활성이 있는 효소에 대해 코딩하는 열린 판독 프레임을 제거합니다. 프로토콜은 선택 시퀀스를 절단할 수 있지만 ESC의 다른 곳에서는 절단할 수 없는 변형으로 출력 라이브러리를 보강하므로 PCR 증폭에 적합하지 않기 때문에 이 단계는 엄격하게 필수가 아닙니다. 그러나, 원래 서열 특이성을 가진 효소가 출력에서 제거되지 않고 변경된 특이성을 가진 유망한 변이체를 능가할 것이기 때문에 선택 효과는 더 낮을 것으로 예상되며 또한 효소 활성이 감소할 것이다. 채우기 수준에서 원하는 대상 시퀀스와 원하지 않는 대상 시퀀스는 모두 퇴화될 수 있지만 그렇지 않을 수도 있습니다. NlaIV 예에서, 안티-타겟은 퇴화되고 표적은 퇴화되었다. 인구 수준에서 퇴화가 있더라도, 단일 액적에서 단 하나의 (비퇴화) 표적 또는 반표적이 존재한다. 프로토콜에서 대상 및 안티 대상 시퀀스는 선택 단계의 모든 반복에서 다시 도입됩니다. 따라서 개방 된 판독 프레임은 가능한 모든 대상을 갈라 놓을 수있는 효소를 인코딩해야하며, 여러 선택 라운드에서 살아남기 위해 항 표적 중 어느 것을 갈라 놓을 수 없습니다. 프로토콜의 각 반복에서 선택 취소 대상을 다시 도입해야 할 필요성은 두 개의 순차적 PcR을 적용합니다. 제1 PCR은 안티-타겟 외부의 어닐리마를 사용하는 프라이머를 사용하므로, 반대로 표적의 분열이 PCR 반응을 방지한다. 두 번째 PCR은 안티 타겟을 넘어 도달 프라이머를 필요로하고, 안티 대상을 다시 도입, 선택의 여러 라운드 동안 있는지 확인하기 위해, 각 열린 독서 프레임은 안티 대상의 모든 변종에 대해 테스트됩니다.
끈적끈적한 말단을 생성하는 효소의 경우, REase ORF10의 분리를 위한 앞에서 설명한 방법에 기초한 관련 대체 프로토콜이 사용될 수 있다. 실험에서 사용되는 비오틴 포획에 의한 비활성 변이체의 고갈은 선택적 PCR에서 프라이머 결합 부위로 사용되는 서열과 호환어댑터의 결찰에 의해 대체되는 프로토콜로 대체된다(도9). 선택된 특이성을 가진 효소를 생성하는 ESC만이 결찰 가능 말단을 생성하고 따라서 선택될 것이다. 카운터 e선택 시퀀스의 끈적끈적한 말단의 시퀀스는 어댑터와 결찰에 참여할 수 없는 방식으로 설계되어야 한다. 선택 프로세스의 반복은 선택적 PCR에서 두 개의 서로 다른 어댑터와 결과적으로 두 개의 서로 다른 역프라이머 사이를 전환하여 쉽게 달성할 수 있습니다.
새로운 프로토콜이 있더라도 시험관 내 새로운 특이성을 엔지니어링하는 작업은 여전히 매우 까다롭습니다. 전형적인 타입 II REases를 위해, 서열 특이성 및 내분핵용해 활성은 동일한 단백질 지구에 의존한다. 따라서 다른 하나에 영향을 주지 않고 하나를 변경하기가 어렵습니다. 성공은 효소의 발자국을 고려하고, 단백질-DNA 상호 작용의 대칭을 존중하며, NlaIV 예8에서와 같이 생화학 실험에서 선행으로 결정되어야 하는 기존의 효소 선호도를 기반으로 하는 전략에 의해 더 가능성이 높아집니다.
공개
저자는 공개 할 것이 없다.
감사의 말
이 작품은 폴란드 국립 과학 센터 (NCN)(UMO-2011/02/A/NZ1/00052)에서 과학 고등 교육부(0295/B/PO1/2008/34~ MB 및 N301 100 31/3043~KS)의 보조금으로 지원되었습니다. UMO-2014/13/B/NZ1/03991 및 UMO-2014/14/M/NZ5/00558- MB) 및 KS에 대한 단기 EMBO 펠로우십(ATSF 277.00-05).
자료
| Name | Company | Catalog Number | Comments |
| 1000Å CPG Support (dA, dT, dC, dG) | Biosset | 45-1000-050 | Other vendors can be used as well |
| ASM-800 DNA/RNA | Biosset | 800-001-000 | |
| GeneJET Gel Extraction Kit | Thermo Scientific | K0691 | Any other kit can be used |
| Glen-Pak DNA purification cartridge | Glen Research | 60-5200 | |
| HIS-Select Nickel Affinity Gel | Sigma | P6611 | |
| pET 28a vector | Any other vector with T7 promoter upstream of plycloning site can be used instead | ||
| Phusion High-Fidelity DNA Polymerase | Thermo Scientific | F530S | Any other high fidelity and highly processive thermophilic polymearse can be used instead |
| Porous steel foil | Biosset | 40-063 | |
| Rapid Translation System RTS 100, E.coli HY Kit | Roche | 3 186 148 | |
| Restriction endonucleases | Thermo Scientific | Obviously other vendors, enzymes can be used | |
| Streptavidin Magnetic Beads | New England Biolabs | S1420S | Other vendors can be used as well. We have positively tested beds form Sigma |
| Synthesis chemicals including phosphoramidities | Carl Roth | Other vendors can be used as well | |
| Synthesis columns (different sizes) | Biosset | ||
| T4 DNA ligase | Thermo Scientific | EL0011 | Any other ligase can be used |
참고문헌
- Schöttler, S., Wenz, C., Lanio, T., Jeltsch, A., Pingoud, A. Protein engineering of the restriction endonuclease EcoRV--structure-guided design of enzyme variants that recognize the base pairs flanking the recognition site. European Journal of Biochemistry. 258 (1), 184-191 (1998).
- Wenz, C., Hahn, M., Pingoud, A. Engineering of variants of the restriction endonuclease EcoRV that depend in their cleavage activity on the flexibility of sequences flanking the recognition site. Biochemistry. 37 (8), 2234-2242 (1998).
- Samuelson, J. C., Xu, S. Y. Directed evolution of restriction endonuclease BstYI to achieve increased substrate specificity. Journal of Molecular Biology. 319 (3), 673-683 (2002).
- Samuelson, J. C., et al. Engineering a rare-cutting restriction enzyme: genetic screening and selection of NotI variants. Nucleic Acids Research. 34 (3), 796-805 (2006).
- Rimseliene, R., Maneliene, Z., Lubys, A., Janulaitis, A. Engineering of restriction endonucleases: using methylation activity of the bifunctional endonuclease Eco57I to select the mutant with a novel sequence specificity. Journal of Molecular Biology. 327 (2), 383-391 (2003).
- Morgan, R. D., Luyten, Y. A. Rational engineering of type II restriction endonuclease DNA binding and cleavage specificity. Nucleic Acids Research. 37 (15), 5222-5233 (2009).
- Skowronek, K., Boniecki, M. J., Kluge, B., Bujnicki, J. M. Rational engineering of sequence specificity in R.MwoI restriction endonuclease. Nucleic Acids Research. 40 (17), 8579-8592 (2012).
- Czapinska, H., et al. Crystal Structure and Directed Evolution of Specificity of NlaIV Restriction Endonuclease. Journal of Molecular Biology. 431 (11), 2082-2094 (2019).
- Miller, O. J., et al. Directed evolution by in vitro compartmentalization. Nature Methods. 3 (7), 561-570 (2006).
- Zheng, Y., Roberts, R. J. Selection of restriction endonucleases using artificial cells. Nucleic Acids Research. 35 (11), e83(2007).
- Takeuchi, R., Choi, M., Stoddard, B. L. Redesign of extensive protein-DNA interfaces of meganucleases using iterative cycles of in vitro compartmentalization. Proceedings of the National Academy of Science U.S.A. 111 (11), 4061-4066 (2014).
- Howland, J. L. Short Protocols in Molecular Biology. Ausubel, F., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., Struhl, K. , Third edition, John Wiley & Sons. New York. (1995).
- Wilson, D. S., Keefe, A. D. Chapter 8 Unit 8.3: Random mutagenesis by PCR. Current Protocols in Molecular Biology. , John Wiley & Sons. New York. (2001).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유