
JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
얼룩말 피시 가스트룰라의 2광자 현미경을 사용하여 깊고 공간적으로 제어된 볼륨 절제
요약
배아 발달은 세포 운동의 대규모 조정을 요구합니다. 2광자 포동 중재 레이저 절제는 심층 세포의 큰 그룹의 공간 적으로 제어 된 3 차원 절제를 허용한다. 또한, 이 기술은 생체 내에서 집단적으로 이동하는 세포의 반응을 기계적 환경에서 의 동요로 조사할 수 있다.
초록
Morphogenesis는 조직과 기관으로 세포를 조직하기 위하여 많은 세포 운동을 관련시킵니다. 적절한 개발을 위해서는 이러한 모든 움직임을 엄격하게 조정해야 하며 증거를 축적하면 적어도 부분적으로는 기계적 상호 작용을 통해 이 것을 달성해야 합니다. 태아에서 이것을 시험하는 것은 직접적인 물리적 인 동요를 필요로 합니다. 레이저 절제는 기계적 제약을 완화하거나 두 세포 집단을 물리적으로 격리할 수 있는 점점 더 많이 사용되는 옵션입니다. 그러나, 많은 절제는 제한된 축 분해능 및 조직 침투를 제공하는 자외선 (UV) 레이저로 수행됩니다. 방법은 2광자 현미경을 사용하여 깊고, 중요하고, 공간적으로 잘 정의된 볼륨을 축하하기 위하여 여기에서 기술됩니다. 절제는 축 중음근에서 녹색 형광 단백질을 표현하는 형질 형질 제브라피 시라인에서 입증되고 과열 된 자궁 또는 기본 노른자 세포에 영향을 주지 않고 축 성 메센도더름을 절단하는 데 사용됩니다. 세포 행동은 절제 전후에 살아있는 화상 진찰에 의해 감시됩니다. 절제 프로토콜은 몇 미크론에서 백 미크론에 이르는 비늘에서 모든 세포 유형 또는 조직에 다른 발달 단계에서 사용할 수 있습니다.
서문
세포 세포 상호 작용은 개발에 중요한 역할을합니다. 세포는 그들의 직접적인 이웃, 또는 더 멀리 세포가, 그들의 운명 및/또는 행동에 영향을 미치는, 인식할 수 있는 신호를 제공합니다. 이러한 신호의 대부분은 본질적으로 화학 물질이다. 예를 들어, 잘 특징인 유도 이벤트에서, 한 세포 그룹은 다른 세포 집단의 운명에 영향을 미치는 확산 분자를 생성1. 그러나 다른 신호는 기계적입니다. 세포는 이웃이 인식하고 대응하는 이웃에게 힘과 제약을 행사합니다2.
생체 내에서 이러한 세포 세포 상호 작용의 중요성을 연구 하는 한 가지 방법은 일부 세포를 제거 하 고 후속 개발을 관찰 하는. 불행히도 세포를 제거하거나 파괴하는 데 사용할 수 있는 기술은 제한됩니다. 세포는 바늘 또는 작은 전선을 사용하여 외과적으로 제거 될 수 있지만, 이러한 치료는 침습적이며 매우 정확하지 않으며 일반적으로 현미경으로 즉시 이미징을 방지하여 스테레오 현미경으로 수행됩니다. 또한 깊은 세포를 대상으로 하는 것은 조직 과대 결막에 구멍을 뚫고 원치 않는 혼란을 일으킵니다. KillerRed와 같은 유전적으로 인코딩된 광세제는 빛 조명을 통해 세포 사멸을 유도하는 데 사용되어 왔다5. 광세제는 빛 조사 시 반응성 산소 종을 생성하는 크로모포라이트입니다. 그들의 주요 한계는 세포가 움직이는 경우에 달성하기 어려울 지도 모르다 긴 빛 일루미네이션 (약 15 분)를 필요로 하고, 즉각적인 것이 아닙니다 세포 사멸을 통해 세포 죽음을 유도한다는 것입니다.
마지막으로, 레이저 절제는 개발 되었으며 널리 지난 15 년 동안 사용 되었습니다6,7,8,9,10,11,12. 레이저 빔은 표적 세포/조직에 초점을 맞습니다. 그것은 가열을 통해 그것의 절제를 유도, 광절, 또는 플라즈마 유도 절제; 관련 프로세스는 전력 밀도 및 노출 시간13에 따라 달라집니다. 대부분의 절제 프로토콜은 높은 에너지에 자외선 레이저를 사용합니다. 그러나, UV 광은 모두 흡수되고 생물학 조직에 의해 산란됩니다. 따라서 깊은 세포를 대상으로하려면 높은 레이저 전력이 필요하며, 이는 더 피상적이고 평면 외 조직에서 손상을 유도합니다. 이것은 표면 구조에 UV 레이저의 사용을 제한하고 상대적으로 낮은 축 해상도를 설명합니다. 비선형 광학(소위 2광자 현미경 검사법)은 적외선 영역에서 약 반 에너지의 두 광자를 가진 형광을 자극하기 위해 빛의 비선형 특성을 사용합니다. 절제에 적용하면 세 가지 주요 장점이 있습니다. 첫째, 적외선은 생물학적 조직에 의해 UV 빛보다 덜 산발적이고 덜 흡수되어 필요한 레이저 전력을 증가하지 않고 더 깊은 구조물에 도달할 수 있습니다. 둘째, 펨토초 펄스 레이저의 사용은 매우 높은 전력 밀도를 제공하여 플라즈마 유도를 통해 절제를 생성하며, 이는 가열과는 달리 공간적으로 확산되지 않습니다15. 셋째, 플라즈마 형성을 유도하는 전력 밀도는 초점에서만 도달한다. 이러한 특성 덕분에, 2광자 레이저 절제는 주변 조직 환경에 영향을 주지 않고 깊은 세포를 정확하게 표적으로 하는 데 사용될 수 있다.
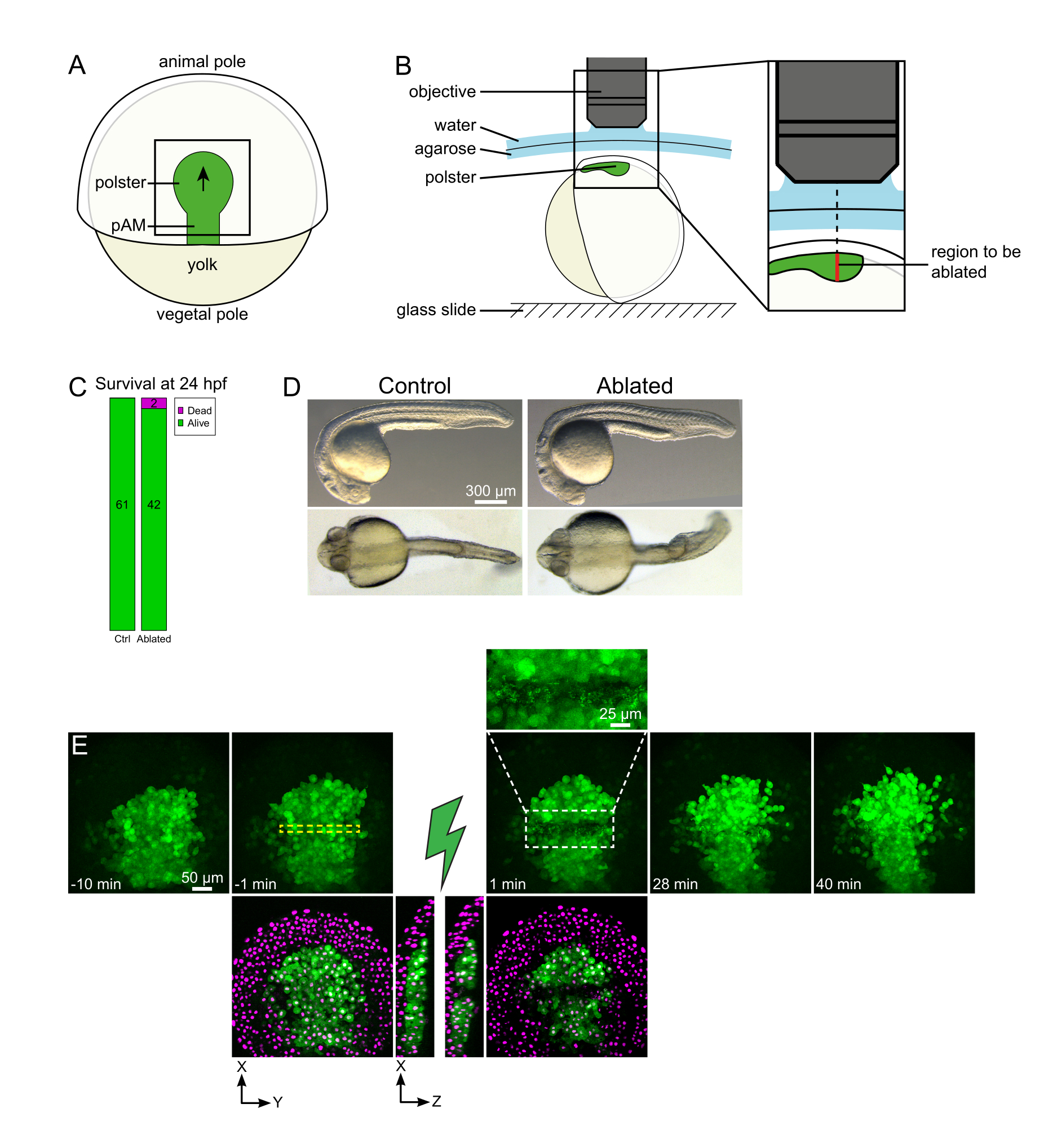
집단 이동은 세포 세포 상호 작용이 기본인 발달 과정의 훌륭한 예입니다. 집단 이동은 인접한 세포가 하나의 cell16의 동작에 영향을 미치는 세포 마이그레이션으로 정의됩니다. 이러한 상호 작용의 본질 (화학 또는 기계적) 그리고 세포 마이그레이션에 미치는 영향은 크게 다를 수 있으며 종종 완전히 이해되지 않습니다. 세포를 제거하고 이것이 다른 사람들에게 어떻게 영향을 미치는지 관찰하는 능력은 이러한 집단 적 과정을 더욱 해명하는 데 중요합니다. 몇 년 전, 우리는 제브라피쉬 위장 중 폴스터의 이주가 집단 이주라는 외과 적 접근법을 사용하여 설립했습니다17. polster는 배아18의 등쪽 측에 있는 첫번째 내재화 세포를 구성하는 세포의 단입니다. Tg(gsc:GFP) 형질전환선에서 녹색으로 표지된 이 세포는 태아 의 깊은 곳에 위치하며, 에피블라스트 세포의 여러 층 아래에 있습니다. 위질 도중, 이 단은 축 중구의 확장을 이끌고, 배아 조직자에서 동물 극으로 이동19,20,21,22,23 (도 1A). 우리는 세포가 동물 극의 방향으로 그들의 이주를 방향을 그들의 이웃과 접촉이 필요하다는 것을 설치했습니다. 그러나, 이 집단 이주의 세포 및 분자 기지를 더 잘 이해하는 것은 이것이 나머지 그들에 어떻게 영향을 미치는지 보기 위하여 몇몇 세포를 제거하는 관련시킵니다. 따라서 우리는 2광자 현미경 설정을 사용하여 크고 깊은 양의 절제를 개발했습니다. 여기에서, 우리는 Histone2B-mCherry로 표지된 핵을 추적하여 중앙에 있는 polster를 단절하고 세포 이동에 대한 결과를 관찰하기 위하여 이 프로토콜의 사용을 보여줍니다.
프로토콜
모든 동물 작업은 윤리위원회 N 59와 미니스티에르 드 l'Education Nationale, de l'Enseignement Supérieur et de la Recherche에 의해 승인되었다 파일 번호 APAFIS#15859-2018051710341011v3. 아래에 설명된 단계 중 일부는 장비 및 소프트웨어에 만지지만 다른 장비에 쉽게 적응할 수 있습니다.
1. 사출 준비
- 배아 매체(EM)에서 1% 아가로즈 용액의 75mL를 준비한다.
- 주입 금형을 90mm 페트리 접시에 넣고 약 50mL의 아가로즈를 부어 곰팡이가 떠있을 수 있습니다. 아가로즈가 고화되어 주입 금형을 제거합니다.
- 아가로즈 코팅 된 요리를 30mm 페트리 접시에 1 mL를 부어 준비하십시오.
- RNase가 없는 물에 스톡 용액을 희석하고 얼음을 유지하여 30 ng/μL Histone2B-mCherry mRNA 용액의 4 μL을 준비하십시오.
참고: RNase 매개 저하를 피하기 위해 mRNA를 조작하는 동안 장갑을 착용하십시오. - 마이크로 피펫 풀러를 사용하여 모세관에서 주사 바늘을 당깁니다.
2. 배아 준비
- 물고기가 계란을 낳으면, 수집, 헹지, EM에서 90mm 페트리 접시에 수확하십시오. 배아를 28.5°C 인큐베이터에 배치합니다.
- 첫 번째 셀이 표시될 때까지 20분 기다립니다.
- EM으로 채워진 주사 판에 30개의 배아를 전달합니다. 약간 무딘 집게를 사용하여 홈에 배아를 짜내고 동물 극으로 방향을 지정합니다.
- 마이크로 로더 팁을 사용하여 2 μL의 mRNA 용액으로 주사 바늘을 채웁니다. 폴리테트라플루오로틸렌(PTFE) 튜브와 연결된 마이크로 조작자에 놓인 모세관 홀더에 바늘을 삽입하여 공기 인젝터에 넣습니다.
- 스테레오 현미경에서 조심스럽게 바늘의 끝을 깰.
- 세포내 바늘을 삽입하여 1세포 단계 배아에 mRNA 용액을 주입한다.
참고: 주입된 부피는 세포 부피의 약 1/3입니다. - 28.5°C 인큐베이터에 다시 주입된 배아를 배치합니다.
3. 2광자 현미경의 준비
참고: 이 프로토콜에는 두 개의 레이저가 사용됩니다. 하나는 GFP (920 nm에서)를 이미지하고 절제 (820 nm에서)를 수행하는 데 사용됩니다. 그것은 녹색 / 절제 레이저라고합니다. 다른 하나는 이미지 mCherry에 1160 nm에서 사용된다. 그것은 빨간색 레이저로 언급 될 것입니다.
- 녹색/절제 레이저를 820nm(절제 파장)로 설정하고 빨간색 레이저를 1160nm(mCherry 여기)로 설정합니다.
- 광학 경로에 이동식 거울을 사용하여 스캔 헤드의 입구와 출구에서 녹색/절제 및 빨간색 레이저 빔을 정렬합니다.
참고: 이것은 레이저 빔 포커스를 증가시키고 흥분 및 절제에 대한 초점 볼륨을 최소화합니다. - 목표아래 820 nm에서 녹색/절제 레이저의 최대 출력을 측정합니다. 이렇게하려면, 목표 아래에 전원 미터를 배치, 검은 챔버를 닫습니다, 녹색 / 절제 레이저 전력을 설정 100%, 셔터를 엽니 다. 300mW에 도달하는 데 필요한 레이저 전력의 백분율을 계산합니다.
- 그린/절제 레이저를 920nm(GFP 여기)로 되돌리고 레이저 파워를 7%로 설정합니다. 빨간색 레이저 전원을 15%로 설정합니다.
- 녹색 및 빨간색 선에 대한 에피 PhotoMultiplier 튜브 (PMT) 검출기를 활성화; 녹색 및 빨간색 선 PMT 감도를 65로 설정합니다.
- 시야를 400 x 400 μm로 설정하고 이미지 해상도를 512 x 512픽셀로 설정하고 주파수를 800Hz로 스캔합니다.
- 3D 타임랩스 이미징 모드를 선택합니다. 그런 다음 폴더를 만들고 각 수집 후 데이터에 대해 Autosave를 활성화합니다.
- 가열 챔버를 조립하고 28 °C로 설정합니다. 챔버와 목표를 따뜻하게 하기 위해 적어도 10 분 기다립니다.
4. 배아 장착
- 형광 스테레오 현미경의 밑에, GFP를 표현하는 70%에 태아를 확인합니다.
참고: 축 중소에서 밝은 신호가 있는 배아를 선택하고 더 나은 이미징 품질을 위해 배경 형광이 없습니다. - 플라스틱 파스퇴르 파이펫을 사용하여 아가로즈 코팅 접시(1.3단계)에서 선택된 배아 3~4개(1.3단계)를 옮기고 미세포프를 사용하여 조심스럽게 비반향을 불러올 수 있습니다.
참고: 점진 배아는 매우 섬세하며 공기 또는 플라스틱과의 접촉시 파열됩니다. - 작은 유리 바이알에 1x 페니실린-연쇄절제술 EM으로 0.2% 아가로즈 1mL를 붓습니다. 유리병을 예열된 42°C 드라이 블록 히터에 놓습니다.
참고: 아가로즈가 설정되기 전에 배아 방향을 허용하려면 다음 단계를 신속하게 수행해야 합니다. - 불을 연마한 유리 파이펫을 사용하여 0.2% 아가로즈 유리 바이알에 비료배아를 전달한다. 그것을 희석하지 않도록 아가로즈에 너무 많은 EM을 추가하지 않도록주의하십시오. 파이펫에서 나머지 EM을 버리고 배아가 파이펫에서 떨어지기 전에 유리 바닥 접시의 슬라이드를 덮을 충분한 아가로즈와 함께 배아를 다시 흡인시합니다.
- 접시의 유리 슬라이드에 아가로즈와 배아를 날려. 배아가 접시의 공기 나 플라스틱 측면을 만지지 않도록주의하십시오. 다음으로, 아가로즈로 유리 슬라이드 주위에 챔버를 채웁니다.
- 표적 부위가 맨 위에 있도록 속눈썹을 사용하여 배아를 지향합니다(그림 1B).
참고: 배아를 지향할 때, 매우 깨지기 쉬운 노른자가 아니라 blastoderm을 만질 수 있습니다. 아가로즈는 실온에 따라 약 1분 만에 설정됩니다. - 아가로즈가 완전히 설정하려면 ~ 5 분 기다린 다음 페니실린 연쇄 절제술 EM 몇 방울을 추가하십시오.
5. 배아 및 절제 전 이미징 찾기
- 유리 바닥 접시를 가열 된 챔버에 목표 아래에 놓습니다. 페니실린 연쇄상 구균 EM에 목표를 몰입하고 가열 챔버를 닫습니다.
- 슬라이더를 이동하여 가벼운 경로를 안구로 설정합니다. 그런 다음, 안구, 형광 등등 및 단계 제어를 사용하여 배아를 찾아 배아의 표면에 초점을 설정합니다.
- 형광 램프를 끄고 PMT로 가는 광 경로를 설정하고 검은 색 챔버를 닫습니다.
참고: PMT가 손상될 수 있으므로 검은 색 챔버의 모든 광원을 끄십시오. - 라이브 이미징을 시작하고 축 중구를 찾습니다. 녹색/절제 및 빨간색 레이저 힘을 조정하여 좋은 신호(즉, GFP 표현 영역에 대한 픽셀당 1,000~20,000광자 사이)를 갖습니다. 빨간색 채널을 사용하여 스테이지를 배아의 맨 위로 이동하고 이 위치를 Z = 0으로 설정합니다.
- 1분의 시간 단계와 2 μm의 Z 단계를 선택합니다. 110 μm의 Z 코스는 전체 폴스터를 포괄하기에 충분하며 이러한 설정으로 1 분 이내에 획득됩니다. 축 중소(더 피상적인 자궁)의 위에 첫 번째 슬라이스 15 μm을 설정합니다.
참고: 폴스터는 곡선선을 따라 움직여 Z 스택의 하단 슬라이스를 시간 경과 이미징(그림 1E)에서 움직임을 수용하기 위해 폴스터 의 가장 깊은 위치보다 30 μm 더 깊게 설정되어야 합니다. - 사전 절제 영화의 10-15 분 기록.

그림 1: 레이저 절제의 성공적인 결과. (A) 등쪽 보기에서 70%의 진동 배아의 계획; pAM: 후방 축 중소. 검은 색 화살표는 폴스터 마이그레이션의 방향을 표시합니다. 검은 색 사각형은 폴스터의 절제에 대한 일반적인 시야 필드를 나타냅니다. (B) 폴스터 절제를 위한 배아 장착 방식. 측면 보기입니다. 배아는 폴스터의 평면이 광학 축에 수직이 될 정도로 장착된다. (C) 생존 및 (D) 24시간 후 수정에서 대조군 및 축약배의 형태. 스케일 바는 Histone2B-mCherry를 발현하는 Tg(gsc:GFP) 배아의 폴스터에서 레이저 절제로부터 300 μm.(E) 시간 시퀀스이다. 녹색 채널이 있는 뷰는 최대 투영값일 뿐입니다. 클로즈업은 셀 파편이 들어 있는 압하영역을 표시합니다. 녹색과 빨간색(마젠타로 표시) 채널이 있는 뷰는 절제 전후의 XY 및 XZ 슬라이스입니다(녹색 번개 볼트는 절제를 나타냅니다). XZ 슬라이스는 과대 조직 (GFP 발현이없는 마젠타 핵)이 기본 구조의 절제에 의해 영향을받지 않았음을 보여줍니다. 노란색 대시 박스는 레이저 절제 치료를 위해 선택한 ROI에 해당합니다. 스케일 바는 대형 뷰50 μm, 클로즈업 시 25 μm입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
6. 대상 위치 및 레이저 절제
- 라이브 이미징에서 폴스터 윤곽을 찾아, 관심의 전기 광학 변조기 영역 (EOM ROI) 도구를 사용하여, 폴스터의 폭에 걸쳐 20 픽셀 (15 μm) 큰 사각형을 그립니다. 이 사각형을 폴스터 의 중간에 놓습니다(그림 1E).
- polster 셀을 포함하는 가장 높고 낮은 평면의 축 위치를 유의하십시오. 절제는 이 두 비행기 사이에 있는 10 μm마다 수행됩니다. ROI가 이 비행기의 노른자 셀과 겹치지 않도록 주의하십시오.
- 스테이지를 간격의 가장 낮은 Z 위치에 배치합니다. 이물질이 빛을 흡수하기 때문에 절제는 상향식 수행해야 합니다.
- 녹색/절제 레이저 파장을 820nm로 설정하고 파워 백분율 을 설정하여 300mW(단계 3.3)의 출구 전력을 얻습니다.
- 이미징 주파수를 200Hz로 설정합니다.
- 녹색/절제 레이저 이미징 EOM을 0으로 설정하고 ROI-Treat 모드를 선택합니다.
- EOM을 켜고 치료를 즉시 시작하도록 설정합니다(0 프레임 후).
- 이미징 모드를 시간 경과로 설정하고 자동 저장을 해제합니다.
- 시간 단계를 빠른 모드로 설정합니다.
- 처리 프레임 수와 프레임 수를 대상 깊이에 해당하는 값으로 설정합니다(표 1).
| 깊이(μm) | 트리트먼트 프레임 |
| -30 | 1 |
| -35 | 1-2 |
| -40 | 1-2 |
| -45 | 2 |
| -50 | 2-3 |
| -55 | 3 |
| -60 | 3-4 |
| -65 | 4 |
| -70 | 4 |
| -75 | 4-5 |
| -80 | 4-5 |
| -85 | 5 |
| -90 | 5 |
| -95 | 5-6 |
| -100 | 6 |
| -105 | 6 |
표 1: 배아내 표적 세포 깊이의 함수로서 레이저 처리 프레임의 수를 건의하였다(0은 배아의 표면임).
- 이미징을 시작합니다. EOM 처리 중에 PMT셔터가 닫히면서 획득이 검은색입니다.
- 스테이지를 목록의 다음 Z 위치로 이동합니다(6.2단계).
- 폴스터 의 상단에 도달 할 때까지 6.10에서 6.12 단계를 반복합니다.
7. 절제 후 검증 및 이미징
- 녹색/절제 레이저를 920nm 및 5%의 전력으로 설정합니다. 녹색/절제 레이저 이미징 EOM을 100으로 설정하고 풀필드 모드를 선택합니다.
- 이미징 주파수를 800Hz로 설정합니다.
- 라이브 모드에서 전체 스택을 통과하여 모든 비행기가 절단되었는지 확인합니다. 그렇지 않은 경우 6.2단계로 돌아갑니다.
참고: 절제는 때때로 Z 스택을 다시 정의해야 할 수 있도록 인접한 조직의 수직 이동을 유도합니다. - 이미징 모드를 3D 타임랩스로 설정하고 자동 저장을 다시 활성화합니다. 절제 후 영화의 40-60 분 기록.
- 절제 후 영화에서 표적 세포가 효과적으로 절단되었는지 확인하십시오. 형광 회복, 또는 공간을 점유하고 추종자 세포가 통과하는 것을 방지하는 표적 세포는 표적 세포가 단지 광표백되고 abLATED되지 않았다는 것을 나타냅니다 (그림 1E 및 도 2A).

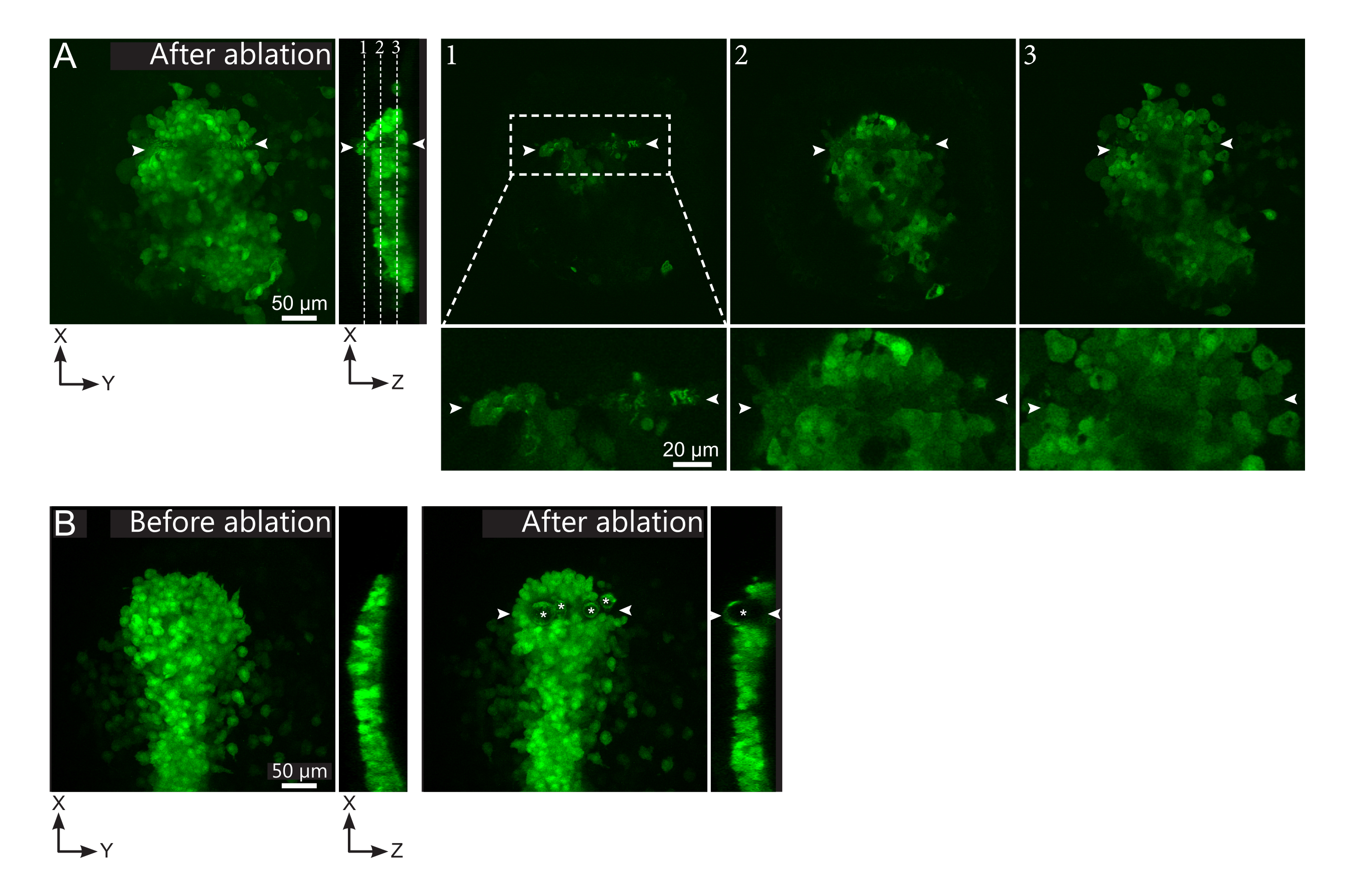
그림 2: 레이저 절제의 부정적인 결과. (A) 레이저 절제에 잠재적인 실패의 전형적인 예. 큰 XY 뷰는 최대 투영이며 XZ 뷰는 재구성된 섹션입니다. 레이저 처리 영역은 두 개의 흰색 화살촉 사이에 있습니다. 세 개의 초점 평면이 재구성된 섹션에서 강조 표시되고 오른쪽에 표시됩니다. 그들은 실패의 세 가지 종류에 해당합니다. 평면 1은 폴스터 위의 세포가 절단되었음을 보여줍니다. 이는 폴스터 위에 있는 이 초점 평면(클로즈업 참조)에 자동 형광 파편이 존재하여 식별할 수 있습니다(재구성된 섹션의 평면 1 위치 참조). 이렇게 하면 영역의 잘못된 정의가 제거될 수 있습니다. 평면 2는 표백되었지만 절단되지 않은 세포를 보여줍니다. 그(것)들은 낮은 형광 신호가 아직도 그대로 세포 윤곽을 드러내기 때문에 확인될 수 있습니다 (클로즈업 참조). 비행기 3는 레이저 치료에 의해 표백되지 않은 그대로 세포를 표시합니다. 이 영역의 잘못 된 정의에서 발생할 수 있습니다 또는 가난한 치료에서. 평면 2 및 3에 묘사된 상황에서, 비압 표적 세포에 절제 처리를 다시 적용할 수 있다. 스케일 바는 넓은 시야에서 50 μm, 클로즈업시 20 μm입니다. (B) 너무 강렬한 레이저 처리로 인해 캐비테이션에 의해 형성된 기포(백색 별표로 표시)의 전형적인 예. 이러한 기포는 Z 평면에 국한되지 않고 때로는 폴스터의 전체 높이에 걸쳐 이웃 조직을 변형시합니다. 스케일 표시줄은 50 μm입니다 .
{kind=link}
8. 데이터 분석
- 이미지 분석 소프트웨어가 있는 타임랩스 시리즈를 열고 올바른 픽셀 크기를 설정합니다.
- 스팟 함수에서, 이것은 위화 시 평균 핵 크기이기 때문에 오브젝트 크기를 10 μm로 설정합니다. 그런 다음 스팟 함수를 실행하여 핵을 감지하고 추적합니다.
참고: Z축을 따라 12μm 길이의 타원 형상을 장착하여 낮은 축 해상도를 고려하여 검출이 약간 향상될 수 있다. - 필터를 사용하여 거짓 긍정을 제거합니다. Tg (Gsc:GFP) 선에서 배아 축 및 일부 내피 세포에서 세포는 녹색으로 표시됩니다. 따라서 녹색 강도에 필터링하면 이러한 셀을 빠르게 선택할 수 있습니다(그림 3A).
- 연속 점 사이의 최대 거리를 셀 속도와 호환되는 값으로 설정합니다.
참고: 두 프레임 사이의 시간 간격을 고려하십시오. 폴스터 세포는 0.8 μm/분 에서 2.8 ± 이동합니다. 따라서 시간 단계 1분 동안 최대 변위 4μm을 허용하면 대부분의 관절 트랙이 제거됩니다. - 한두 시간 포인트에 걸쳐 간격을 허용하면 연속 트랙이 길어지지만 추적 오류가 발생할 수 있습니다. 핵이 일회성 지점에서 올바르게 감지되지 않으면 서로 다른 매개 변수/필터를 사용하는 스팟 감지를 다시 실행하는 것이 좋습니다.
- 시각적으로 트랙을 확인하고 필요한 경우 수정합니다.
- 결과를 .xlsx 파일로 내보냅니다. 게시된 스프레드시트 루틴24 (그림 3B) 및 데이터 분석 소프트웨어에 대한 사용자 지정 루틴(요청 시 사용 가능)을 사용하여 파일을 처리합니다.

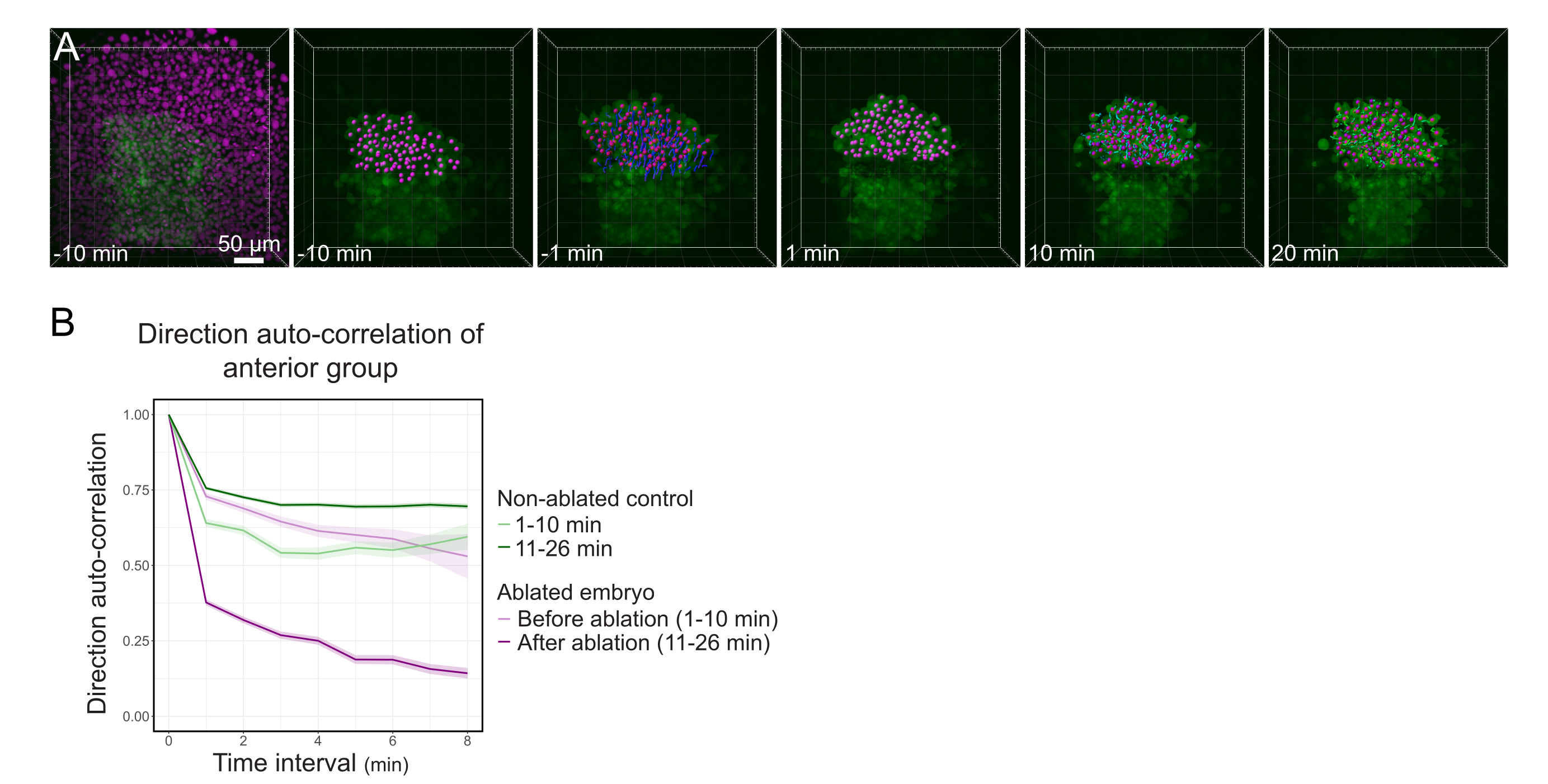
도 3: 폴스터의 전방 반의 격리는 세포 방향성에 영향을 미칩니다. (A) 3D 재구성은 Histone2B-mCherry (마젠타에 표시됨)를 표현하는 Tg (gsc:GFP) 배아를 중간에 있는 폴스터를 절단하는 레이저 절제 전후를 발현한다. 폴스터의 전방 절반에 속하는 핵은 마젠타 점으로 표시되고 절제 전후에 시간이 지남에 따라 추적됩니다( Movie S1 참조). 스케일 바는 50 μm. (B) 이력 지속성의 척도로서, 절제 전후의 폴스터의 전방 부분에 속하는 세포의 방향 자동 상관관계이다. 세포는 절제 전에 연속 모션을 표시하며, 이는 절제 후 크게 감소하여 집단 지향 마이그레이션의 손실을 나타냅니다. 방향 자동 상관관계는 또한 비ablated 태아의 폴스터의 전방 절반을 형성하는 세포에 측정되었다, 대조군으로. 그래프 봉투는 표준 오류를 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
결과
중간에 폴스터를 끊기 위해, Histone2B-mCherry mRNAs로 주입된 Tg(gsc:GFP) 배아는 4단계에서 설명된 바와 같이 70% 에피보리 스테이지에 장착되었다. 폴스터는 GFP 발현에 의해 확인되었고, 배아는 폴스터의 평면이 광학 축(도 1B)에 수직되도록 장착되었다. 이 위치에서 배아를 기울이면 절차가 복잡해집니다. 빛은 절제 비행기에 도달하기 위해 더 많은 조직을 통과해야하며, 절?...
토론
여기서는 비선형 광학을 사용하여 깊고 공간적으로 잘 정의된 볼륨 절제를 수행하는 프로토콜을 설명합니다. 프로토콜의 가장 중요한 단계는 과도한 파편이나 캐비테이션을 피하기 위해 절제를 허용하지만 너무 많은 에너지를 허용하는 충분한 에너지를 제공하는 치료 조건을 찾는 것입니다. 대상 부위에 전달되는 에너지의 양은 주로 레이저 출구 전력, (2) 레이저 정렬의 품질, (3) 빛이 절제 평면...
공개
저자는 경쟁 이익을 선언하지 않습니다.
감사의 말
우리는 물고기 관리에 대한 에밀리 메넌트, 폴리 테크닉 바이오 이미징 시설, 특히 피에르 마후, 부분적으로 레지온 일 드 프랑스 (interDIM)와 기관 국립 드 라 레체체 (ANR-11-EQPX-0029 Morphoscope2, ANR-10-INBS-04)에 의해 지원 자신의 장비에 라이브 이미징에 대한 지원을 제공합니다. 이 작품은 ANR 보조금 15-CE13-0016-1에 의해 지원되었다, 18-CE13-0024, 20-CE13-0016, 그리고 마리 Skłodowska-Curie 보조금 협정에 따라 유럽 연합의 호라이즌 2020 연구 및 혁신 프로그램 No 840201, 미니슈레 드 l'Enseignement Supérieur et de la Recherche 및 국립 센터 드 라 리셔.
자료
| Name | Company | Catalog Number | Comments |
| 25x water immersion objective | Olympus | XLPLN25XWMP2 | |
| Agarose | PanReac AppliChem | A8963,0500 | |
| Data analysis software : Matlab | Math Works | ||
| Electro-optic modulator (EOM) | ConOptics | 350-80LA | |
| Embryo Medium (EM) solution | Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 5th Edition. University of Oregon Press, Eugene (Book). (2000). | ||
| Environmental chamber chamber | Okolab | H201-T-UNIT-BL | |
| EOM driver | ConOptics | 302RM | |
| Fluorescence source | Lumencor | SOLA | |
| Glass bottom dishes | MatTek | P35G-0-10-C | |
| Glass capillaries | Harvard Apparatus | 300085 | Outside diameter 1.0 mm, inside diameter 0.58 mm |
| Glass pipettes | Volac | D810 | Tip should be fire polished |
| Green/ablation laser | Spectra Physics | Mai Tai HP DeepSee | |
| Histone2B-mCherry mRNA | Synthesized from pCS2-H2B-mCherry plasmid (Dumortier& al. 2012) | ||
| Image analysis software: IMARIS | Bitplane | ||
| ImSpector software | Abberior Instruments Development Team | ||
| Injection mold | Adapative Science Tools | I-34 | |
| Microloader tips | Eppendorf | 5242956003 | |
| Micromanipulator | Narishige | MN-151 | |
| Micropipette puller | Sutter | P-1000 | |
| mMESSAGE mMACHINE SP6 Transcription Kit | Invitrogen | AM1340 | |
| Penicillin-Streptomycin | Thermofisher | 15140-122 | 10 000 units penicillin and 10 mgstreptomycin per ml |
| Photomultiplier tube (PMT) | Hammamatsu | H7422-40 | |
| PicoPump (Air injector) | World Precision Instrument | PV820 | |
| Red laser | Spectra Physics | OPO/Insight DeepSee | |
| RNAse free water for injection | Sigma | W3500 | |
| Spreadsheet software: Excel | Microsoft | ||
| Stereomicroscope | Nikon | SMZ18 | |
| Tg(gsc:GFP) zebrafish line | Doitsidou, M. et al. Guidance of primordial germ cell migration by the chemokine SDF-1. Cell. 111 (5), 647–59, doi: doi.org/10.1016/S0092-8674(02)01135-2 (2002). | ||
| TriM Scope II microscope | La Vision Biotech |
참고문헌
- Slack, J. M. W. Embryonic induction. Mechanisms of Development. 41 (2-3), 91-107 (1993).
- Fernandez-Sanchez, M. -. E., Brunet, T., Röper, J. -. C., Farge, E. Mechanotransduction's impact on animal development, evolution, and tumorigenesis. Annual Review of Cell and Developmental Biology. 31, 373-397 (2015).
- Shih, J., Fraser, S. E. Characterizing the zebrafish organizer: microsurgical analysis at the early-shield stage. Development. 122 (4), 1313-1322 (1996).
- Selleck, M. A. J. Culture and microsurgical manipulation of the early avian embryo. Methods in Cell Biology. 51 (51), 1-21 (1996).
- Bulina, M. E., et al. A genetically encoded photosensitizer. Nature Biotechnology. 24 (1), 95-99 (2006).
- Fang-Yen, C., Gabel, C. V., Samuel, A. D. T., Bargmann, C. I., Avery, L. Laser microsurgery in Caenorhabditis elegans. Methods in Cell Biology. 107, 177-206 (2012).
- Colombelli, J., Grill, S. W., Stelzer, E. H. K. Ultraviolet diffraction limited nanosurgery of live biological tissues. Review of Scientific Instruments. 75 (2), 472-478 (2004).
- Smutny, M., Behrndt, M., Campinho, P., Ruprecht, V., Heisenberg, C. -. P. UV laser ablation to measure cell and tissue-generated forces in the zebrafish embryo in vivo and ex vivo. Methods in Molecular Biology. 1189, 219-235 (2015).
- Behrndt, M., et al. Forces driving epithelial spreading in zebrafish gastrulation. Science. 338 (6104), 257-260 (2012).
- Volpe, B. A., Fotino, T. H., Steiner, A. B. Confocal microscope-based laser ablation and regeneration assay in zebrafish interneuromast cells. Journal of Visualized Experiments: JoVE. (159), (2020).
- Bonnet, I., et al. Mechanical state, material properties and continuous description of an epithelial tissue. Journal of the Royal Society, Interface. 9 (75), 2614-2623 (2012).
- Rauzi, M., Lenne, P. F., Lecuit, T. Planar polarized actomyosin contractile flows control epithelial junction remodelling. Nature. 468 (7327), 1110-1115 (2010).
- Niemz, M. H. . Laser-Tissue Interactions. Encyclopedia of Biomaterials and Biomedical Engineering, Second Edition - Four Volume Set. , (2019).
- Smith, A. M., Mancini, M. C., Nie, S. Bioimaging: second window for in vivo imaging. Nature Nanotechnology. 4 (11), 710-711 (2009).
- Rauzi, M., Lenne, P. -. F. Cortical forces in cell shape changes and tissue morphogenesis. Current Topics in Developmental Biology. 95, 93-144 (2011).
- Theveneau, E., David, N. B. Migrations cellulaires collectives. Medecine/Sciences. 30 (8-9), 751-757 (2014).
- Dumortier, J. G., Martin, S., Meyer, D., Rosa, F. M., David, N. B. Collective mesendoderm migration relies on an intrinsic directionality signal transmitted through cell contacts. Proceedings of the National Academy of Sciences of the United States of America. 109 (42), 16945-16950 (2012).
- Solnica-Krezel, L., Stemple, D. L., Driever, W. Transparent things: cell fates and cell movements during early embryogenesis of zebrafish. BioEssays. 17 (11), 931-939 (1995).
- Montero, J. -. A., Kilian, B., Chan, J., Bayliss, P. E., Heisenberg, C. -. P. Phosphoinositide 3-kinase is required for process outgrowth and cell polarization of gastrulating mesendodermal cells. Current Biology. 13 (15), 1279-1289 (2003).
- Ulrich, F., et al. Slb/Wnt11 controls hypoblast cell migration and morphogenesis at the onset of zebrafish gastrulation. Development. 130 (22), 5375-5384 (2003).
- Kai, M., Heisenberg, C. -. P., Tada, M. Sphingosine-1-phosphate receptors regulate individual cell behaviours underlying the directed migration of prechordal plate progenitor cells during zebrafish gastrulation. Development. 135 (18), 3043-3051 (2008).
- Smutny, M., et al. Friction forces position the neural anlage. Nature Cell Biology. 19 (4), 306-317 (2017).
- Johansson, M., Giger, F. A., Fielding, T., Houart, C. Dkk1 controls cell-cell interaction through regulation of non-nuclear β-Catenin pools. Developmental Cell. 51 (6), 775-786 (2019).
- Gorelik, R., Gautreau, A. Quantitative and unbiased analysis of directional persistence in cell migration. Nature Protocols. 9 (8), 1931-1943 (2014).
- Grill, S. W., Howard, J., Schäffer, E., Stelzer, E. H. K., Hyman, A. A. The distribution of active force generators controls mitotic spindle position. Science. 301 (5632), 518-521 (2003).
- Desprat, N., Supatto, W., Pouille, P. -. A. A., Beaurepaire, E., Farge, E. Tissue deformation modulates twist expression to determine anterior midgut differentiation in Drosophila embryos. Developmental Cell. 15 (3), 470-477 (2008).
- Farhadifar, R., Röper, J. -. C., Aigouy, B., Eaton, S., Jülicher, F. The influence of cell mechanics, cell-cell interactions, and proliferation on epithelial packing. Current Biology. 17 (24), 2095-2104 (2007).
- Willier, B. H., Oppenheimer, J. M. . Foundations of Experimental Embryology. , (1964).
- Ashby, W. J., Zijlstra, A. Established and novel methods of interrogating two-dimensional cell migration. Integrative Biology: Quantitative Biosciences from Nano to Macro. 4 (11), 1338-1350 (2012).
- Bosze, B., et al. Pcdh18a regulates endocytosis of E-cadherin during axial mesoderm development in zebrafish. Histochemistry and Cell Biology. 154 (5), 463-480 (2020).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유