
Method Article
Utilizando Genómica Funcional de triagem para identificar alvos de drogas potencialmente romance em Spheroid culturas de células de câncer
Neste Artigo
Resumo
Identifying novel drug targets that transition from pre-clinical testing to human trials is a scientific priority. To that end, here we describe a functional genomics approach for examining the impact of gene depletion on cancer cell line spheroids, which more appropriately model human cancers in vivo.
Resumo
A identificação de eventos excitador funcionais no câncer é fundamental para aprofundar nossa compreensão da biologia do câncer e indispensável para a descoberta da próxima geração de novos alvos de drogas. Está se tornando evidente que os modelos mais complexos de câncer são obrigados a apreciar plenamente os fatores que contribuem para que impulsionam tumorigênese in vivo e aumentar a eficácia de novas terapias que fazem a transição de modelos pré-clínicos para ensaios clínicos.
Aqui apresenta-se uma metodologia para a geração de esferóides tumorais uniforme e reprodutível, que pode ser submetido a triagem funcional de siRNA. Estes esferóides exibir muitas características que são encontradas em tumores sólidos que não estão presentes na cultura bidimensional tradicional. Mostramos que várias linhas celulares de cancro da mama comumente utilizados são passíveis de este protocolo. Além disso, nós fornecemos a prova de princípio de dados utilizando a linha de células de cancro da mama BT474, confirmando a suadependência de amplificação do receptor do factor de crescimento epidérmico e HER2 mutação de fosfatidilinositol-4,5-bifosfato-3-quinase (PIK3CA), quando crescidas como tumores esferóides. Finalmente, somos capazes de investigar e confirmar o impacto espacial destas dependências usando imuno-histoquímica.
Introdução
Os tumores sólidos exibir histológica significativa, a heterogeneidade genética intra-tumoral e micro-ambientais, que apresenta os clínicos com um desafio significativo na capacidade de tratar pacientes com sucesso. A maioria dos modelos utilizados para identificar novas terapias direcionadas não incorporam muitos desses recursos. De facto, as terapias correntes orientadas utilizados na clínica foram desenvolvidos na década passada utilizando abordagens de rastreio que se baseiam em linhas celulares de cancro cultivadas sob condições bidimensionais (2D) de cultura. Embora este trouxe vários sucessos, tais como inibidores do receptor tirosina quinase, está se tornando evidente que os modelos mais complexos de câncer são obrigados a apreciar plenamente os fatores que contribuem para que impulsionam tumorigênese in vivo e aumentar o número de novas terapias que fazem a transição de modelos pré-clínicos para ensaios clínicos. Além disso, é agora bem apreciado que os sistemas de cultura 2D não reflectem emvivo comportamento 1,2. Por exemplo, em tumores vascularizados mal, como a procura de oxigénio e nutrientes, no microambiente superar a oferta, as regiões de alta e baixa entregas desenvolver. A presença de baixos níveis de oxigênio (hipóxia) em tumores, como detectado pela coloração imuno-histoquímica de secções de tumor para os marcadores de hipóxia estabelecidos, como anidrase carbônica IX (CAIX), correlaciona-se com pior evolução clínica no cancro da mama 3,4 características Assim que incorporem tais como hipoxia em modelos de triagem pode melhorar a nossa capacidade de descobrir alvos de drogas novas que serão mais eficazes in vivo. Na verdade, dirigidas com sucesso tumores agressivos que contenham hipóxia é uma prioridade clínica 5.
A alteração de rastreio tradicional 2D siRNA, numa tentativa de recapitular com mais precisão os elementos de condições encontradas por células cancerosas no microambiente tumoral, levou à identificação de vários genes thno foram encontrados como sendo importantes para o crescimento do tumor in vivo. Estes incluem telas de genômica funcional realizadas em condições séricos 6, condições de hipóxia 7 e 8 em combinação. Por exemplo, o silenciamento de 6-Phosphofructo-2-Quinase / frutose-2,6-Biphosphatase 4 (PFKFB4), uma proteína responsável pela regulação da glicólise entrar carbono, somente induziu a apoptose em linhas de cancro da próstata derivadas de metástases, quando cultivada em baixo teor de soro. O silenciamento de PFKFB4 em linhas de células da próstata normais e sob as mesmas condições não teve nenhum efeito, ao passo que, a depleção de PFKFB4 ablated completamente o crescimento de linha celular de cancro da próstata de xenoenxertos 6.
Em um painel alargado de linhas celulares de cancro da mama, o silenciamento de mono-carboxilase transportador 4 (MCT4) preferencialmente conduzido para a redução do crescimento da linha de células sob condições de baixo teor de oxigénio. Esta vulnerabilidade foi validado in vivo no cancro da mama linha de células orthotropic xenografts. Talvez o mais impressionante, silenciando de acetil-CoA sintetase 2 (ACSS2), uma enzima responsável pela conversão de acetato em acetil-CoA, número reduzido de câncer de células sob condições de estresse de nutrientes (baixa de oxigênio e soro), mas teve pouco ou nenhum efeito em condições de cultura normais 8. A ablação de ACSS2 impacto no crescimento de xenoenxertos de cancro da próstata e da mama, sugerindo que os gradientes de nutrientes não existem no isolamento dentro do microambiente do tumor e que as células que residem nestas regiões são essenciais para a progressão do tumor 8. Além disso, também ACSS2 verificou-se ser importante no glioblastoma e carcinoma hepatocelular 9,10, sugerindo que o aumento da actividade ACSS2 em tumores pode ser um mecanismo fundamental que suporta o crescimento sob condições desfavoráveis.
Colectivamente, estes estudos demonstram que recapitulando as condições encontradas in vivo e realizar telas siRNA permite a identificação de genes essenciais para a sobrevivência de câncer. Como bem como tendo impacto sobre o crescimento de células de cancro 2D nutrientes sob condições de stress, o esgotamento dos genes alvo nestes estudos inibiu o crescimento do esferóide linha celular de cancro, à semelhança do que foi observado em xenoenxertos tumorais 6,8. Assim, esferóides linha celular de cancro conter várias das condições encontradas no microambiente tumoral que conferem sensibilidade ao silenciamento ACSS2. Com efeito, esferóides apresentar gradientes de nutrientes e oxigénio (soro), alterações no pH, tridimensional de contacto célula-célula (3D), mas também, alterações na celeridade proliferativa com as células submetidas a paragem do ciclo celular e apoptose. Isto é exemplificado pela presença de esferóides de cancro de regiões necróticas, uma característica não encontrada na cultura 2D tradicional.
esferóides de células de cancro já foram utilizados como modelos mais biologicamente relevantes para rastrear inibidores de moléculas pequenas, mas isto só permite a validação da eficácia do composto ou reaproveitamento de COMPOunds originalmente projetar para outras doenças 11. métodos de rastreio esferóide actuais não permitem a análise da depleção de gene específico de uma de alto rendimento de modo de alta conteúdo. Aqui, nós descrevemos, pela primeira vez, um oleoduto genómica funcional para a descoberta de genes específicos dependências utilizando a tecnologia de pequeno ARN interferente (siRNA) em esferóides linha celular de cancro. Nós projetamos uma biblioteca sob medida com siRNAs segmentação dos 200 genes mais frequentemente mutado em cancros da mama humanos e avaliou o impacto do esgotamento gene do tamanho do esferóide e da atividade metabólica em BT474 esferóides de câncer de mama. Fomos capazes de detectar de forma robusta e reproduzível do impacto da ErbB2 e PIK3CA silenciando em culturas 3D. Além disso, pode-se avaliar o impacto da depleção do gene na arquitectura espacial dos esferóides BT474 usando imuno-histoquímica.
Protocolo
1. Preparação de placas de 96 poços de siRNA
NOTA: As bordas exteriores de uma placa de 96 poços são mais propensas a evaporação em comparação com outros poços, portanto, limitar a quantidade de siRNAs de 60 por placa de 96 poços. Preencha poços exteriores com meio simples ou PBS para limitar isso. Além disso, um ecrã de visitas de validação é recomendado para aliviar polarizações placa.
- Diluir os siRNAs de 250-500 ng / mL em meio de soro reduzido (conforme as recomendações do fabricante) e uma alíquota de 10 ul às cavidades apropriados da placa de fixação ultra-baixa. Executar todas as telas em triplicado.
NOTA: Incorporar controles siRNA não-alvo (negativo) e matando (positivo, como UBB e PLK1) apropriadas no design da placa para garantir a eficiência de transfecção pode ser avaliado. A utilização de chapas de fixação para baixo o trabalho siARN é crítico, como células vão ser adicionados à mistura de transfecção. Não podemos descartar que alguns fabricantes de ultra-baixo apegoplacas podem ter efeitos sutis sobre crescimento de esferóides. Como tal, recomenda-se que estes são testados pelo utilizador final. - Cubra as placas com selos adesivos e armazenar a -20 ° C.
2. A transfecção reverso da Linha celular
- No dia da tela, descongelar as placas à temperatura ambiente e girar durante 5 minutos a 1000 x g utilizando uma centrífuga de bancada. Adicionar 10 ul de meio de soro reduzido contendo o reagente de transfecção optimizada para cada poço utilizando uma pipeta de canais múltiplos. Deixar as placas durante 15 min para os complexos de transfecção contendo siARN para formar.
NOTA: Este estudo utilizou a linha celular do cancro da mama BT474 para o rastreio genómico funcional (cultivadas em DMEM de glicose elevada (por Dulbecco Modified Eagle Médium) suplementados com 10% de soro fetal de bovino (FBS) e antibióticos). - Tripsinizar as células a serem rastreadas (tripsina a 0,05% e EDTA 0,53 mM) até que se destacam a partir do balão, neutralizar usando volume apropriado de linha celular especiIC médio e centrifugação durante 5 min a 1000 xg numa centrífuga de bancada para remover a tripsina. Ressuspender as células com um volume apropriado de meio e determinar o número de células precisa utilizando um contador de células.
NOTA: Normalmente 5.000 células por poço são suficientes para a formação esferóide na maioria das linhas celulares testadas. É importante contar com precisão as células como uma suspensão de células individuais para comparações de tamanho precisos. - Dilui-se as células a 5000 células por 180 uL de meio frio (4 ° C).
NOTA: A adição de componentes de matriz de membrana basal reconstituída irá impactar negativamente a eficiência de transfecção e recomenda-se que ele não é usado. linha de otimização de celular para confirmar a formação esferóide capacidade e eficácia knockdown deve ser realizada antes da tela. - Transferir a suspensão de células a um reservatório, pipeta para misturar e adicionar 180 uL de cada poço da placa de fixação 96 poços ultra-baixo, contendo o siRNA preparado anteriormente (2,1).
- Centrifuge a placa a 1000 x g numa solução pré-arrefecida de 4 ° C, centrifuga-se durante 10 min e, em seguida, voltar para uma incubadora de cultura de tecido de 37 ° C.
- NOTA: As células aparece como um mosaico no fundo dos poços. Durante o próximo 12-24 horas, as células irão agregar em conjunto para formar uma única esfera.
- Após 24 h, as células observar formar um único esferóide no centro do poço. Adicionar 100 uL de meio completo a cada poço para encorajar o crescimento.
- Reabastecer meio depois de três dias. Remova cuidadosamente 100 ul de meio de cada poço e adiciona-se 100 uL de meio fresco.
- No dia 7, quantificar o tamanho esferóide automatizado em um leitor de placas que pode monitorar quantitativamente crescimento de esferóides ao longo do tempo (ver Secção 3).
- Em seguida, determinar a viabilidade celular utilizando um corante de viabilidade celular luminescente (ver Secção 4).
3. Automated Image Acquisition
- Digitalizar as placas em uma bancada, imagiologia placa micro-bem citômetro no dia 7.
- Abra o software, selecione o seguinte: '96 -bem placa ', selecione o tipo de placa apropriado e insira um nome de experimento. NOTA: Todas as informações adicionais também podem ser adicionadas para o software.
- Selecione o aplicativo 'Tumorsphere'. Alterar o foco para que o esferóide está em foco e tem um contraste perfeito.
NOTA: Recomendamos 'foco baseado em imagem' como variações significativas nos tamanhos esferóides são esperados. - Selecione os poços que necessitam de digitalização e "Iniciar verificação".
- Usando o software do scanner placa, certifique-se de que a máscara objeto representa com precisão o tamanho esferóide. Para fazer isso, ajustar as configurações de diâmetro colônia, dilatação das fronteiras, de espessura mínima e precisão, específico para cada linha de células testadas 11,12.
Nota: A área esferóide será então calculada usando o algoritmo de software. Por exemplo, para calcular com precisão a área de esferóides BT474 cultivados por sete dias ajuste de precisão para 'alta' umd definir o diâmetro mínimo colônia para 200? M. Isso deve produzir um representante máscara esferóide adequada da área esferóide.
NOTA: Estes dados podem ser exportados a partir da máquina, usando a função de exportação, no sob a forma de um arquivo de dados anotada. Data é placa média normalizada, combinadas e analisadas tanto por z-score ou diferença média rigorosamente padronizado (SSMD) para identificar siRNAs que tiveram um efeito estatisticamente significativo na área de esferóide.
4. Determinar a viabilidade celular
- Após a área de esferóide foi calculada, determinar a viabilidade celular utilizando um corante de viabilidade luminescente. Prepara-se o reagente de acordo com as instruções do fabricante.
- remover cuidadosamente 100 ul de meio de cada poço e adiciona-se 100 ul de corante de viabilidade. Incubar as placas durante 15 min, em seguida, digitalizar utilizando um leitor de placas luminescente.
NOTA: Estes dados podem ser exportados a partir da máquina, usando a função de exportação, em noa forma de um ficheiro de dados anotados. Data é placa média normalizada, combinadas e analisadas tanto por z-score ou diferença média rigorosamente padronizado (SSMD) para identificar siRNAs que tiveram um efeito estatisticamente significativo sobre a viabilidade esferóide.
Resultados
Os ensaios de ligação em placas de esferóide ultra-baixas de 96 poços fornecer uma avaliação fenotipica de alta capacidade para o potencial de oncogenicidade em um contexto, que mais facilmente recapitula as condições fisiológicas encontradas em tumores in vivo. De fato, as linhas celulares de cancro MCF10DCIS.com e forma BT474 estruturas esferóides apertadas (Figura 1A) e investigação imuno das secções esferóides mostrou mudanças espaciais distintas na morfologia celular e nuclear. Ao longo do tempo, alguns, tais como os esferóides esferóides BT474 desenvolver regiões necróticas, uma característica comum de tumores sólidos agressivos (Figura 1B). Alguns esferóides não desenvolvem múltiplos centros necróticos, tais como a linha celular MDA-MB-231, mas exibem variação marcada na marcador proliferativa Ki67, que inversamente correlacionada com a expressão clivado caspsase-3, um marcador de apoptose (Figura 1C). Para estabelecer que várias linhas de células são, de facto receptiva para BT474 silenciamento de genes mediada por ARNsi, MCF10DCIS.com, células JIMT1 MDA-MB-231 e foram transfectadas com ARNsi inversa durante sete dias. A presença de reagente de transfecção (simulado) ou a transfecção de siRNAs de controlo não teve nenhum efeito sobre a viabilidade esferóide, enquanto o silenciamento do gene essencial Ubiquitina B (UBB) reduziu significativamente a viabilidade esferóide em todas as linhas celulares testadas (Figura 1D).
Nós projetamos uma biblioteca siRNA humana que englobava os genes mais frequentemente mutados em desmarcada, ER +, HER2 + e triplo cânceres de mama negativos. Essa biblioteca consiste em genes de função conhecida, tais como MYC, PIK3CA e TP53 e aqueles cuja contribuição para a carcinogénese não está estabelecida. A tela também contém vários siRNAs controle-voltados para os não (Controle # 1, Controle # 2) e siRNAs segmentação genes essenciais, tais como PLK1 e UBB, que atuam como matando controles (Tabela 1). Nós escolhemos usar o canc de mamaer linhas de células BT474 em que prontamente formam esferóides sem a adição de membrana basal reconstituída, são estabelecidas linhas de células laborioso e têm uma arquitectura genómica conhecido. Por exemplo, as células BT474 são positivos para o receptor de estrogénio (ER +), sobre-expressam o receptor humano de crescimento epidérmico fator 2 (HER2 +) e mutações do porto em TP53 (E285K) e PIK3CA (K111N) 13.
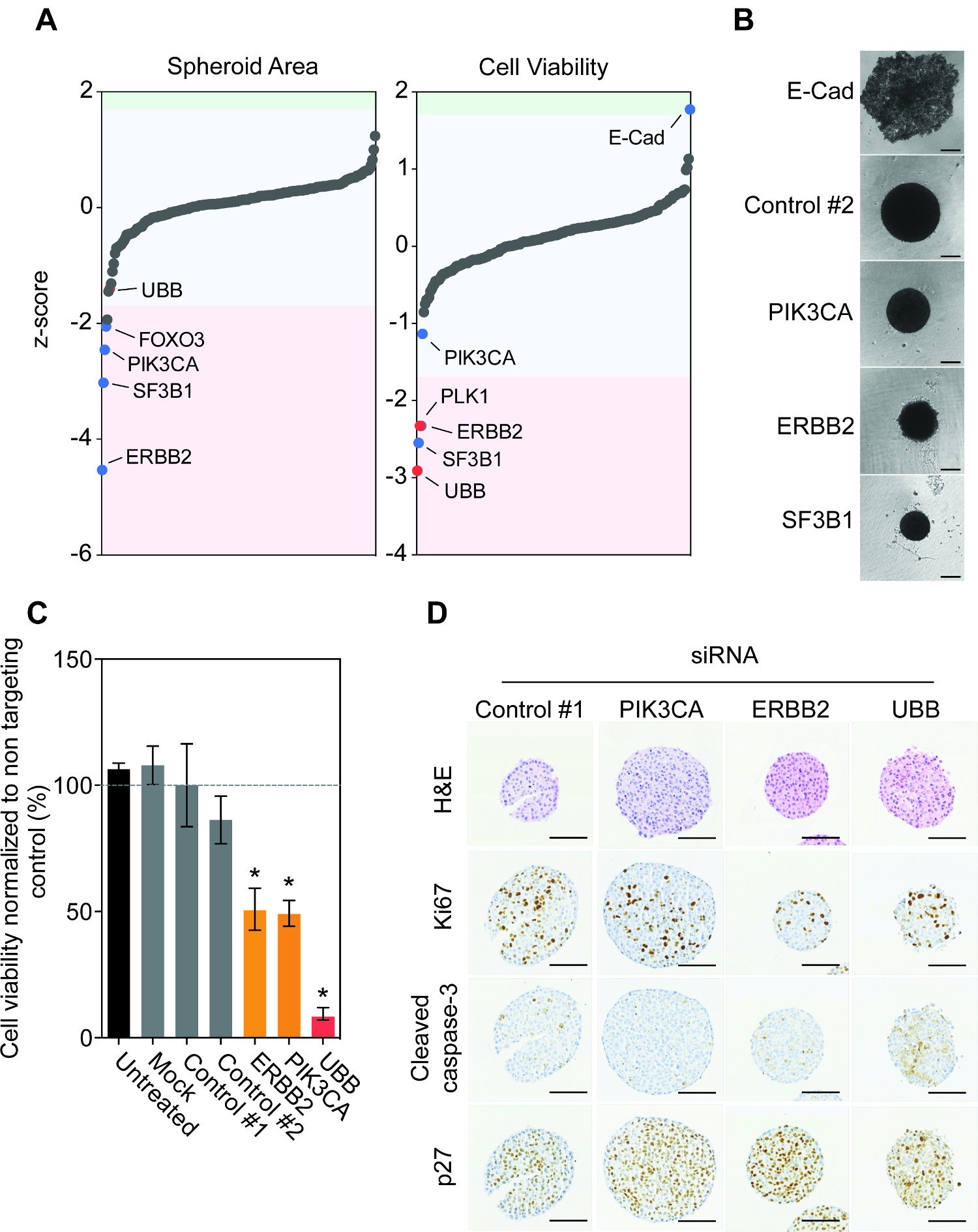
Empregando-se o protocolo descrito acima, que acompanhou a depleção teve impacto gene do tamanho do esferóide e a viabilidade depois de sete dias de siRNA transfecção inversa (Figura 1E). Curiosamente, a maioria dos genes não têm um efeito significativo sobre a área de esferóide ou viabilidade (Figura 2A). Silenciamento de FOXO3, PIK3CA, ErbB2 e SF3B1 resultou na redução reproduzível mais significativa no tamanho do esferóide. Esta redução também foi observada em esferóide viabilidade após ErbB2 e SF3B1 silenciamento. Encorajador, que confirmed o impacto de PIK3CA, ErbB2 e SF3B1 silenciamento do tamanho do esferóide utilizando microscopia de campo brilhante (Figura 2B). Nós previamente identificado SF3B1 como um gene essencial em numerosos modelos de linhas celulares e, portanto, siSF3B1 representa um bom controlo matança para além UBB 14. Curiosamente, todos os 200 genes de apenas o silenciamento de caderina-E, resultou num aumento significativo na viabilidade esferóide (Figura 2A). Investigação da morfologia esferóide mostrou que a caderina-E silenciamento resultou numa desagregação completa da arquitetura esferóide, com células viáveis que descansa na parte inferior do acessório bem baixo (Figura 2B). novo inquérito manual dos dados da tela do volume esferóide mostrou que este também tinha sido observada, mas tinha sido eliminado da área quantificação devido ao objeto que está sendo acima das restrições de tamanho definido. Como destacado anteriormente, as células BT474 sobre-expressam o receptor tirosina quinase HER2 e abrigar uma mutação oncogénica em PIK3CA (K111N). Confirmou-se que o silenciamento de ErbB2 e PIK3CA resultou em viabilidade esferóide reduzida, enquanto a transfecção com controlos não segmentação não teve nenhum efeito (Figura 2C).
Em seguida, investigamos o impacto do esgotamento siRNA na histologia esferóide. esferóides BT474 foram transfectadas reverso com não-alvo siRNA controle e siRNAs segmentação PIK3CA, ErbB2 e UBB. O silenciamento de ErbB2 e UBB resultou numa redução do marcador pró-proliferativa de Ki67 em comparação com ARNsi de controlo (Figura 2D). A activação do pró-apoptótica marcador clivada da caspase-3 foi apenas observada após UBB silenciamento, sugerindo que a depleção de HER2 e PIK3CA não resultaram em apoptose, mas eram em vez de citotóxico citostático. Com efeito, o silenciamento de HER2 e PIK3CA deu origem a um aumento da expressão da proteína p27 da proteína de paragem do ciclo celular em comparação com o controle esferóides transfectadas.
ENT "FO: manter-together.within-page =" 1 "> Tomados em conjunto, estes resultados mostram que as células BT474 são accionados por oncogénica HER2 e PIK3CA sinalização, quando cultivada como esferóides 3D Mais importante, estes resultados mostram que é possível. projetar e implementar uma biblioteca de triagem siRNA bespoke de centenas de genes em esferóides de linha de células de câncer robustamente e reprodutível.
Figura 1: Optimização celular Linha de crescimento 3-dimensional. A. As linhas celulares de cancro da mama, MCF10DCIS.com e BT474 foram cultivadas em placas de fixação baixas durante 7 dias. as imagens de campo claro representativos foram feitas usando um microscópio invertido. Barras de escala = 100 pm. B. esferóides MCF10DCIS.com e BT474 foram cultivadas durante 28 dias. 100 ul de meio fresco foi reabastecido a cada 3 - 4 dias. Esferóides foram fixados em 3,8% de formaldeído, embedded, seccionado e corado com hematoxilina e eosina (H & E). Imagens representativas são mostrados em baixo e alta ampliação. As barras de escala representam 33 e 100? M? M, respectivamente. C. MDA-MB-231 esferóides foram cultivadas durante 21 dias. 100 ul de meio fresco foi reabastecido a cada 3 - 4 dias. Os esferóides foram fixadas em 3,8% de formaldeído, embebida, seccionados e corados com Ki67 e caspase-3 clivada. Imagens representativas são mostrados. Barras de escala = 100 pm. D. BT474, MCF10DCIS.com, MDA-MB-231, e JIMT1 linhas de células foram transfectadas com reversa (apenas o reagente de transfecção) simulada e os siRNAs de controlo e UBB, placas ultra-baixas de fixação foram então centrifugada para formar esferóides. meio fresco (100 uL) adicionou-se nos dias 1 e 4. Após 7 dias, a viabilidade celular foi quantificada. Os dados representam a média ± DP de duas réplicas biológicas independentes realizadas em triplicado normalizadas para controlar # 1. A significância estatística foi calculada usando uma UNPAEstudantes IRED teste t (p <0,05). E. Um diagrama de fluxo resume o protocolo de transfecção inversa usado para interrogar funcionalmente dependência gene em esferóides linha de células cancerosas. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Investigação Genómica Funcional de BT474 Spheroids Descobre Oncogênicos Dependências. A. Células BT474 foram transfectadas com a biblioteca reversa siARN siGENOME humano 200-gene em triplicado. Observou tamanho esferoidal e viabilidade. Valores de dados em bruto foram normalizados placa e escores z médios foram calculados para identificar valores atípicos significativos superiores a 1,7 vez o desvio padrão da placa mediana de 15. Nota genes periféricas ERBB2, SF3B1, PLK1 e UBB umnd E-cad. siRNAs que significativamente aumentados ou reduzidos área esferóide e viabilidade são sombreadas em azul e vermelho, onde o vermelho representa os siRNA de controle de. B. As células foram transfectadas inversa com o controle não-alvo, a caderina-E (E-Cad), PIK3CA, ERBB2 ou UBB siRNA e, em seguida, girou em uma placa de ultra-baixo apego à forma esferóides. Após 7 dias, as imagens de campo claro esferóides representativos foram feitas usando um microscópio invertido. Barras de escala = 100 pm. C. As células foram transfectadas reverso com não-alvo de controle, PIK3CA, ERBB2 ou UBB siRNA e, em seguida, girou em uma placa de ultra-baixo apego à forma esferóides. Após 7 dias, a viabilidade celular foi quantificada. Os dados representam a média ± DP de duas réplicas biológicas independentes realizadas em triplicado normalizadas para controlar # 1. A significância estatística foi calculada usando um Estudantes teste t não emparelhado (p <0,05). D. Depois de 7 dias, esferóides foram fixados, embebidos, seccionados e coradospor H & E, Ki67, caspase-3 clivada, e p27. Imagens representativas são mostrados. Barras de escala = 100 pm. Note-se a H & E dos esferóides siControl parecer menor devido a um artefacto de esferóides de processamento e individuais intacta escolhidos para a coloração. No entanto, isto não diminui as mudanças observadas com as imagens digitalizadas e os resultados de viabilidade celular. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| Não-alvo 1 | TP53 | DST | KMT2C | não tratada 1 | FCGBP | ARID1B | FBXM7 | TTC40 | Não-alvo 2 | ||
| GATA3 | TTN | MUC12 | MUC4 | AHNAK | HUWEI1 | DNAH11 | itpr2 | ABCA13 | CREBBP | ||
| MAP2K4 | PIK3CA | F5 | APOB | ANKRD30A | MUC17 | DNAH17 | LAMA2 | ÁS | CSMD2 | ||
| STARD9 | USH2A | FAT3 | LPR2 | CSMD3 | MYO18B | DNAH5 | MDN1 | ARHGAP5 | DNAH9 | ||
| CXCR3 | MUC16 | RB1 | PKHD1L1 | DNAH2 | SYNE2 | DYNC1H1 | PCLO | CACNA1B | ERBB2 | ||
| PLK1 | SYNE1 | lyst | PTEN | SPTA1 | não tratada 2 | BVS | RYR1 | COL6A3 | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| Não-alvo 1 | ENAM | RYR2 | BRCA2 | não tratada 1 | FMN2 | HECW1 | LAMB4 | SI | Não-alvo 2 | ||
| FHOD3 | MACF1 | RYR3 | C2ORF16 | DMD | FRG1 | HERC2 | MYH11 | STAB1 | ZDBF2 | ||
| GOLGA6L2 | NEB | SMG1 | CACNA1E | DNA14 | GCC2 | HIVEP2 | NIPBL | TANC1 | ZNF536 | ||
| HMCN1 | NF1 | UBR5 | CACNA1F | DYNC2H1 | GON4L | HYDIN | PKD1L1 | TF | ANK3 | ||
| HRNR | OBSCN | USP34 | CYMA5 | FAM208B | GPR112 | ITSN2 | RNF213 | TPR | ASPM | ||
| PLK1 | PCDH15 | XIRP2 | COL7A1 | FLG2 | não tratada 2 | BVS | SAGE1 | UNC80 | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| Não-alvo 1 | DCHS2 | MUC5B | ZFHX4 | não tratada 1 | MYO9A | SPHKAP | CXORF22 | NCOR1 | VPS13D | ||
| ATR | DMXL2 | MXRA5 | ANK2 | KIAA1210 | PRUNE2 | TCHH | DANH6 | NOTCH2 | ANKRD12 | ||
| BIRC6 | DNAH10 | TENM1 | caixa eletrônico | LRP1 | SCN10A | VPS13C | DNAH7 | SPEN | C5ORF42 | ||
| CDH1 | DNAH3 | PEG3 | DIDO1 | MAP1A | ScN2a | IPTR3 | ErbB3 | SRRM2 | CCDC88A | ||
| CUBN | NOCK11 | RELN | DNAH8 | MED12 | SHROOM2 | CEP350 | FAT4 | SZT2 | CC4 | ||
| PLK1 | EYS | SACS | KIAA1109 | MED13 | não tratada 2 | BVS | KMT2A | VPS13A | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| Non-targeting | QSER1 | ARID1A | WDFY3 | não tratada 1 | sdk1 | TEX15 | LAMA1 | Não-alvo 2 | |||
| COL14A1 | SHROOM3 | ATRX | EFCAB5 | SF3B1 | CBFB | AHNAK2 | |||||
| CSMD1 | TBX3 | KIAA0947 | FOXA1 | itpr1 | DDX3X | KIF4A | |||||
| MEFV | UBR4 | MYCBP2 | INPPL1 | FLG | HECTD4 | FAT2 | |||||
| MGAM | VCAN | NBEAL1 | MAP3K1 | AKAP9 | GPR98 | FOXO3 | |||||
| PLK1 | ZNF462 | SETX | NRP1 | HERC1 | não tratada 2 | BVS | UBB | ||||
Tabela 1: layout da placa e siRNA Humano Biblioteca De-convolução. A tabela contém o conteúdo de cada uma das piscinas de siRNA e disposição para as placas baixas de fixação utilizados na tela.
Discussão
Modelos tridimensionais de cancro estão a ser cada vez mais utilizadas para avaliar a eficácia de compostos conhecidos e novos, que foram concebidos para matar selectivamente as células cancerosas. Esferóides de células cancerosas são estruturas que apresentam condições mais semelhantes às encontradas em tumores in vivo, assim, compostos que aumentaram a eficácia em 3D são mais propensos a ter um efeito in vivo. No entanto, essas modalidades não permitem a identificação de novos alvos potencialmente que não tenham sido objecto de desenho de drogas que poderia ter considerável eficácia no tratamento do cancro.
Desenvolvemos um siRNA abordagem genómica funcional que permitiu o silenciamento do gene durável por até sete dias em esferóides linha de células cancerosas. Existem vários passos críticos para o protocolo que requerem optimização antes de uma tela de siARN pode ser realizada. A capacidade para formar esferóides viáveis reprodutíveis em larga escala é essencial. Além disso, umacondições de transfecção ppropriate deve ser rigorosamente optimizada. Sugerimos testando vários reagentes de transfecção diferentes com não-segmentação adequada e matando os controles antes de tentar a tela. Fomos capazes de mostrar que várias linhas celulares de cancro da mama comumente utilizados, ou seja, BT474, MCF10DCIS.com, MDA-MB-231 e JIMT1 eram passíveis de siRNA transfecção. Além disso, nós fornecemos a prova de princípio de dados de rastreio dos 200 genes mais frequentemente mutado no cancro da mama em esferóides BT474, confirmaram a sua dependência de amplificação de HER2 e mutação oncogénica de PIK3CA. Curiosamente, o silenciamento do factor de transcrição FOXO3 resultou numa redução do tamanho do esferóide mas nenhum efeito significativo sobre a viabilidade. FOXO3 é conhecida para regular a resposta à hipóxia, alterando a capacidade metabólica das células cancerosas que lhes permite adaptar-se mais facilmente ao seu ambiente 16. Este papel poderia potencialmente interferir com a leitura viabilidade celular, uma vez que detecta ATP abundância, Um dos principais produtos do metabolismo celular.
Em apoio da observação de uma redução no tamanho do esferóide, foi demonstrado que o silenciamento de FOXO3 HeLa em xenoenxertos auditivos crescimento tumoral e induziu apoptose 17. É importante notar que alguns genes podem afectar a capacidade das células cancerosas para reter a sua arquitectura 3D, o que poderia resultar em falsos positivos. Por exemplo, knockdown da caderina-E resultou na dissolução da estrutura esferóide BT474. Esta havia sido relatado usando E-caderina alvo anticorpos 18. Tal como acontece com qualquer plataforma de triagem, alvos potenciais devem ser novamente testados para avaliar a reprodutibilidade do efeito observado. Existem limitações para a técnica, ou seja, a natureza transitória do silenciamento de genes mediada por siRNA. Sustentados silenciando mais de sete dias não era possível com siRNA.
A vantagem desta abordagem é que ele pode ser acoplado com vários outros biomacorantes tricas e não apenas aqueles que avaliar a viabilidade esferóide, por exemplo, dando informação espacial de hipóxia esferóide ou células monitoramento em apoptose. Além disso, porque os scans leitor de placas são relativamente rápida e não-invasiva, o impacto de ARNic em tamanho esferóide pode ser avaliado ao longo do tempo em vez de apenas no ponto final experimental. Na verdade, estamos a explorar várias dessas avenidas dentro do nosso pipeline de rastreio. Uma abordagem alternativa que utiliza culturas 3D para identificar novos dependências é o uso de bibliotecas químicas que inibem qualquer um de uma ampla gama de alvos particulares ou famílias de proteínas. Com efeito, Bitler et ai. utilizou essa abordagem orientada para identificar a interação letalidade sintética entre o estado ARID1A e inibidores EZH2 no ovário carcinoma de células claras 19. A descoberta da tecnologia de edição gene CRISPR-Cas9 também permitiu o desenvolvimento de telas genéticos em culturas organ�de e in vivo. No entanto, tsua abordagem depende da existência de instalações para animais apropriados e pode ser custo proibitivo 20.
Em conclusão, acreditamos que temos delineado um protocolo que mais precisamente modelos o oxigênio e nutrientes gradientes, que são características do microambiente do tumor in vivo, permitindo a identificação de alvos de câncer de romance ou de validação robusta de metas estabelecidas. Além disso, o protocolo pode ser aplicado a qualquer tipo de linha de células que se forma esferóides e, por conseguinte, pode ser utilizado rotineiramente na comunidade de investigação do cancro para telas de ARNsi de alto rendimento.
Divulgações
Open Access fees were supported by Nexcelom Bioscience, LLC.
Agradecimentos
The authors would like to thank Fredrik Walberg (FACS and imaging facility, ICR) for assistance with microscopy. This work was funded by Breast Cancer Now. RN is the recipient of a Breast Cancer Now Career Development Fellowship (2011MaySF01).
Materiais
| Name | Company | Catalog Number | Comments |
| Lullaby | Oz Biosciences | LL70500 | lipid-based transfection reagent |
| Viromer | Lipocalyx | VB-01LB-01 | virus-like polymer transfection reagent |
| Ultra-low attachment plate | Corning | CLS7007 | 96 well plate |
| Foil plate seals | ThermoFisher | AB-0626 | |
| Luminescent cell viability dye | Promega | G7570 | CellTitre-Glo |
| Pipette tips (200 μL) | Starlab | S1111-0806 | |
| Pipette tips (10 μL) | Starlab | S1111-3800 | |
| Pipette tips (1, 000 μL) | Starlab | S1122-1830 | |
| Serological pipettes (5 mL) | Sarstedt | 86.1253.025 | |
| Serological pipettes (10 mL) | Sarstedt | 86.1254.025 | |
| Serological pipettes (25 mL) | Sarstedt | 86.1685.020 | |
| RPMI Media | GIBCO | 11875-093 | |
| DMEM Media | GIBCO | 11965-084 | |
| Opti-MEM | GIBCO | 31985070 | |
| Feta bovine serum | GIBCO | 16140063 | |
| siRNA | Dharmacon | Cherry picked library | |
| Countess Cell Counter | ThermoFisher Scientific | AMQAX1000 | |
| Cell counting chamber slides | ThermoFisher Scientific | C10312 | |
| Celigo S | Nexcelom | contact company | |
| Victor X5 | Perkin Elmer | contact company | |
| Benchtop centrifuge | Various | ||
| Axiovert Inverted brightfield microscope | Zeiss | contact company | |
| Tissue culture CO2; Incubator | Various | ||
| Mulitichannel pipette | Various |
Referências
- Seton-Rogers, S. E., et al. Cooperation of the ErbB2 receptor and transforming growth factor beta in induction of migration and invasion in mammary epithelial cells. Proc Natl Acad Sci U S A. 101 (5), 1257-1262 (2004).
- Debnath, J., Muthuswamy, S. K., Brugge, J. S. Morphogenesis and oncogenesis of MCF-10A mammary epithelial acini grown in three-dimensional basement membrane cultures. Methods. 30 (3), 256-268 (2003).
- Chia, S. K., et al. Prognostic significance of a novel hypoxia-regulated marker, carbonic anhydrase IX, in invasive breast carcinoma. J Clin Oncol. 19 (16), 3660-3668 (2001).
- Trastour, C., et al. HIF-1alpha and CA IX staining in invasive breast carcinomas: prognosis and treatment outcome. Int J Cancer. 120 (7), 1451-1458 (2007).
- Wilson, W. R., Hay, M. P. Targeting hypoxia in cancer therapy. Nat Rev Cancer. 11 (6), 393-410 (2011).
- Ros, S., et al. Functional metabolic screen identifies 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4 as an important regulator of prostate cancer cell survival. Cancer Discov. 2 (4), 328-343 (2012).
- Baenke, F., et al. Functional screening identifies MCT4 as a key regulator of breast cancer cell metabolism and survival. J Pathol. 237 (2), 152-165 (2015).
- Schug, Z. T., et al. Acetyl-CoA synthetase 2 promotes acetate utilization and maintains cancer cell growth under metabolic stress. Cancer Cell. 27 (1), 57-71 (2015).
- Mashimo, T., et al. Acetate is a bioenergetic substrate for human glioblastoma and brain metastases. Cell. 159 (7), 1603-1614 (2014).
- Comerford, S. A., et al. Acetate dependence of tumors. Cell. 159 (7), 1591-1602 (2014).
- Vinci, M., Box, C., Eccles, S. A. Three-dimensional (3D) tumor spheroid invasion assay. J Vis Exp. (99), e52686 (2015).
- Vinci, M., Box, C., Zimmermann, M., Eccles, S. A. Tumor spheroid-based migration assays for evaluation of therapeutic agents. Methods Mol Biol. 986, 253-266 (2013).
- Barretina, J., et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature. 483 (7391), 603-607 (2012).
- Maguire, S. L., et al. SF3B1 mutations constitute a novel therapeutic target in breast cancer. J Pathol. 235 (4), 571-580 (2015).
- Brough, R., et al. Functional viability profiles of breast cancer. Cancer Discov. 1 (3), 260-273 (2011).
- Ferber, E. C., et al. FOXO3a regulates reactive oxygen metabolism by inhibiting mitochondrial gene expression. Cell Death Differ. 19 (6), 968-979 (2012).
- Jensen, K. S., et al. FoxO3A promotes metabolic adaptation to hypoxia by antagonizing Myc function. EMBO J. 30 (22), 4554-4570 (2011).
- Ivascu, A., Kubbies, M. Diversity of cell-mediated adhesions in breast cancer spheroids. Int J Oncol. 31 (6), 1403-1413 (2007).
- Bitler, B. G., et al. Synthetic lethality by targeting EZH2 methyltransferase activity in ARID1A-mutated cancers. Nat Med. 21 (3), 231-238 (2015).
- Dow, L. E., et al. Inducible in vivo genome editing with CRISPR-Cas9. Nat Biotechnol. 33 (4), 390-394 (2015).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados