
Method Article
利用功能基因组筛选识别在肿瘤细胞球体培养潜在的新药物靶标
摘要
Identifying novel drug targets that transition from pre-clinical testing to human trials is a scientific priority. To that end, here we describe a functional genomics approach for examining the impact of gene depletion on cancer cell line spheroids, which more appropriately model human cancers in vivo.
摘要
功能驱动程序的事件在癌症中的标识是中央对推进我们的癌症生物学的理解和不可缺少的下一代新型药物靶点的发现。它是日益明显的是,癌症更复杂的模型是必需的充分理解,驱动肿瘤发生在体内 ,并增加新疗法,使从临床前模型临床试验的过渡的功效的贡献因素。
这里,我们提出,用于产生均匀的和可再现的肿瘤球状体的方法,可以进行siRNA功能筛选。这些球状体显示了在实体瘤中不存在在传统的两维培养中发现许多特性。我们表明,几种常用的乳腺癌细胞系适合于该协议。此外,我们提供验证的原则,利用乳腺癌细胞系BT474的数据,确认其对表皮生长因子受体的磷脂酰肌醇-4,5-二磷酸3-激酶(PIK3CA)的HER2和突变的扩增依赖性时生长为肿瘤球状体。最后,我们可以进一步调查并确认采用免疫组化这些依赖的空间影响。
引言
实体瘤显示显著组织学,基因和微环境内肿瘤的异质性,这呈现临床医生在能够成功地治疗患者显著挑战。大多数用于鉴定新型模型针对性治疗不掺入许多这些特征。事实上,在临床上使用的当前靶向疗法已经开发了在过去十年中采用依赖于二维(2D)培养条件下生长的肿瘤细胞系的筛选方法。虽然这带来了各种成功,例如受体酪氨酸激酶抑制剂,它是日益明显的是,癌症更复杂的模型是必需的充分理解,驱动肿瘤发生在体内和增加的新疗法,使从过渡的数目的影响因素临床前模型的临床试验。另外,现在是很好理解的是,二维培养系统不能反映在体内行为1,2。例如,在不良血管瘤,因为微环境中的氧气和营养物质的需求超过供应,高,低交付的地区发展。在肿瘤低氧(缺氧)的存在下,由肿瘤切片的用于建立缺氧标记物,如碳酸酐酶IX(CAIX)的免疫组织化学染色检测到,关联与乳腺癌较差的临床结果,如3,4-因此掺入特征缺氧为筛选模型可以提高我们的能力,发现新的药物靶标,将在体内更有效。事实上,包含缺氧定位成功侵略性的肿瘤是临床重点5。
传统的2D的siRNA筛选的改变,在试图以更准确地概括的癌细胞在肿瘤微环境中遇到的条件的元素,已经导致几种基因的鉴定第在已被发现是在体内肿瘤生长重要的。这些措施包括低血清条件6,缺氧条件下,7和8的组合进行功能基因组屏幕。例如,6-磷酸-2-激酶/果糖-2,6- Biphosphatase 4(PFKFB4),负责调节碳进入糖酵解的蛋白质的沉默,低血清中生长时仅诱导从转移衍生的前列腺癌系的细胞凋亡。在相同的条件下正常前列腺细胞系PFKFB4沉默没有效果,而,PFKFB4耗竭完全消融前列腺癌细胞系的生长异种移植物6。
在乳腺癌细胞系的延长面板,单羧转运蛋白4(MCT4)的沉默优先导致细胞生长的低氧气的条件下的减少。此漏洞在体内被证实在乳腺癌细胞系各向异性xenograFTS。养分胁迫条件下也许是最引人注目的是,沉默乙酰-CoA合成酶2(ACSS2),负责转换乙酸转化为乙酰-CoA的酶,减少癌细胞数(低氧和血清),但在正常培养条件下很少或没有影响8。 ACSS2消融影响乳腺癌和前列腺癌异种移植物表明养分梯度不是孤立肿瘤微环境内,并且驻留在这些区域的细胞是肿瘤进展8必需存在的生长。此外,ACSS2也被发现在成胶质细胞瘤和肝细胞癌9,10-重要,这表明在肿瘤中增加ACSS2活性可能是支持不利条件下生长的基本机理。
总的来说,这些研究证明,扼要重述在体内遇到的条件并执行的siRNA屏幕允许克的识别埃内斯癌症生存至关重要。以及影响养分胁迫条件下的2D癌细胞生长,在这些研究中的靶基因的耗尽抑制癌细胞系球体生长,镜像在肿瘤异种移植物6,8-观察什么。因此,癌细胞系球状体包含若干的那赋予ACSS2沉默灵敏度在肿瘤微环境中遇到的条件。事实上,球状体显示养分梯度(血清和氧气),pH值的变化,三维(3D)细胞 - 细胞接触,而且,在与经历细胞周期阻滞和细胞凋亡增殖波速改变。这是通过在坏死区的肿瘤球状体的存在例举,特征未在传统二维培养中。
癌细胞球状体已被用作多种生物相关模型来筛选小分子抑制剂,但这仅允许化合物功效的验证或COMPO的再利用unds原本设计为其他疾病11。当前球体筛选方法不允许特定基因耗竭在高吞吐量的高含量的方式进行分析。在这里,我们描述了在第一时间,功能基因组学管道用于发现利用小干扰RNA(siRNA)技术在癌症细胞系球体特异性基因的依赖关系。我们设计的siRNA在人类乳腺癌针对200最频繁突变的基因一个定制库,评估基因消耗对球体的大小和代谢活动在BT474乳腺癌球体的影响。我们能够鲁棒地和再现地检测的ERBB2和PIK3CA在3D培养沉默的影响。此外,我们可以评估基因消耗对采用免疫组化BT474球体的空间结构的影响。
研究方案
1. 96孔的siRNA板制备
注:在96孔板的外边缘更容易蒸发相比其他井,从而限制的siRNA的量至60%的96孔板。在普通培养基或PBS外井填充限制这一点。此外,命中验证屏建议,以减轻板的偏见。
- 稀释的siRNA到250 - 500毫微克/微升血清培养基减少(根据制造商的建议)和等分试样10μL到超低附着板的适当的孔中。一式三份执行的所有屏幕。
注:在板设计纳入适当的非靶向(负)和杀死(正,如UBB和PLK1)的siRNA控制,以确保转染效率进行评估。为所述siRNA工作中使用的低装接板是重要的,因为细胞将被添加到转染混合物。我们不能排除一些厂家超低附件板块可能会对球体增长微妙的影响。因此,我们建议,这些都是由最终用户进行测试。 - 盖板用胶粘剂密封和储存在-20℃。
2.细胞系的反向转
- 在屏幕的当天,解冻在室温下将板和使用台式离心机在1000 xg离心旋转5分钟。添加血清培养基降低10μL的含有优化转染试剂至各孔用多道移液器。离开板为含有siRNA的形成转染复合15分钟。
注意:此研究中使用(补充有10%胎牛血清(FBS)和抗生素的高糖DMEM中培养(Dulbecco氏改良的Eagle培养基)),用于功能基因组筛查乳腺癌细胞系BT474。 - Trypsinize待筛选的细胞(0.05%胰蛋白酶和0.53 1mM EDTA)中,直到它们从烧瓶分离,中和使用的细胞系特异性的适当体积IC介质和在1000 xg离心自旋5分钟在台式离心机以除去胰蛋白酶。重悬的细胞用培养基的适当体积,并用细胞计数器确定准确的细胞数。
注意:通常,每孔5,000个细胞足以跨越测试大多数细胞系球体的形成。它准确地计数细胞作为单细胞悬浮液用于精确大小的比较是重要的。 - 稀释细胞至每180微升5,000个细胞在冷的介质(4℃)。
注意:除了重组基底膜基质成分会产生负面影响的转染效率和建议不使用它。细胞系的优化,以确认球体形成能力和击倒的功效应前屏幕进行。 - 细胞悬液转移到一个储存器,移液管以混合,并添加180微升含siRNA的预先制备(2.1)的96孔超低附着板的各孔中。
- CentrifugE在预冷4℃离心10分钟的板在1000 xg离心,然后返回到37℃组织培养孵化。
- 注:细胞将显示为在孔的底部上的镶嵌。在接下来的12 - 24小时的细胞将聚集在一起,形成一个单一的球面。
- 24小时后,观察细胞形成在井的中心的单一球体。加入100μl完全培养基,以每孔以鼓励增长。
- 三天后补充网上平台。轻轻地从每个删除100μL介质的很好,加上新媒体的100微升。
- 在第7天,量化上读板器能定量监测随时间球体生长自动化球体大小(见第3)。
- 之后,使用发光细胞生存力染料(参见第4节)确定细胞生存力。
3.自动图像采集
- 扫描仪在第7天对台式,微孔板成像板。
- 打开软件,选择以下内容:'96 -Well板",选择合适的板型和输入实验名称。注意:任何额外的信息也可以被添加到软件。
- 选择"Tumorsphere"的应用程序。改变重点,使球体处于焦点,具有最佳的对比度。
注:我们建议"基于图像的焦点"在球体的大小为显著变化的预期。 - 选择需要扫描"开始扫描"的井。
- 使用扫描板软件,确保对象准确面具代表了球体的大小。通过调整菌落直径,边界扩张,最小厚度和精度设定为此,具体到每一个细胞系测试11,12。
注:球体区域将然后使用软件算法来计算。例如,为了准确地计算生长七天BT474球状体的面积调整精度为"高"的D中的最小菌落直径为200微米。这将产生一个适当的代表球体面积的球体遮罩。
注意:这些数据随后可以从机器中的注释的数据文件的形式输出,使用导出函数,英寸日期是板块中位数标准化,组合和分析无论是Z值或严格标准平均差(SSMD)来标识的siRNA是对球体面积统计显著的效果。
4.确定细胞活力
- 球体区域已经计算之后,确定使用的发光活力细胞染料的生存能力。制备试剂按照制造商的说明。
- 小心地从每个删除100μL介质的很好,并添加活力染料100μL。孵育15分钟,板然后扫描使用的发光板读数器。
注意:这些数据随后可以从机器导出的,使用导出函数,在带注释的数据文件的形式。日期是板块中位数标准化,组合和分析无论是Z值或严格标准平均差(SSMD)来标识的siRNA是对球体的可行性有统计显著的效果。
结果
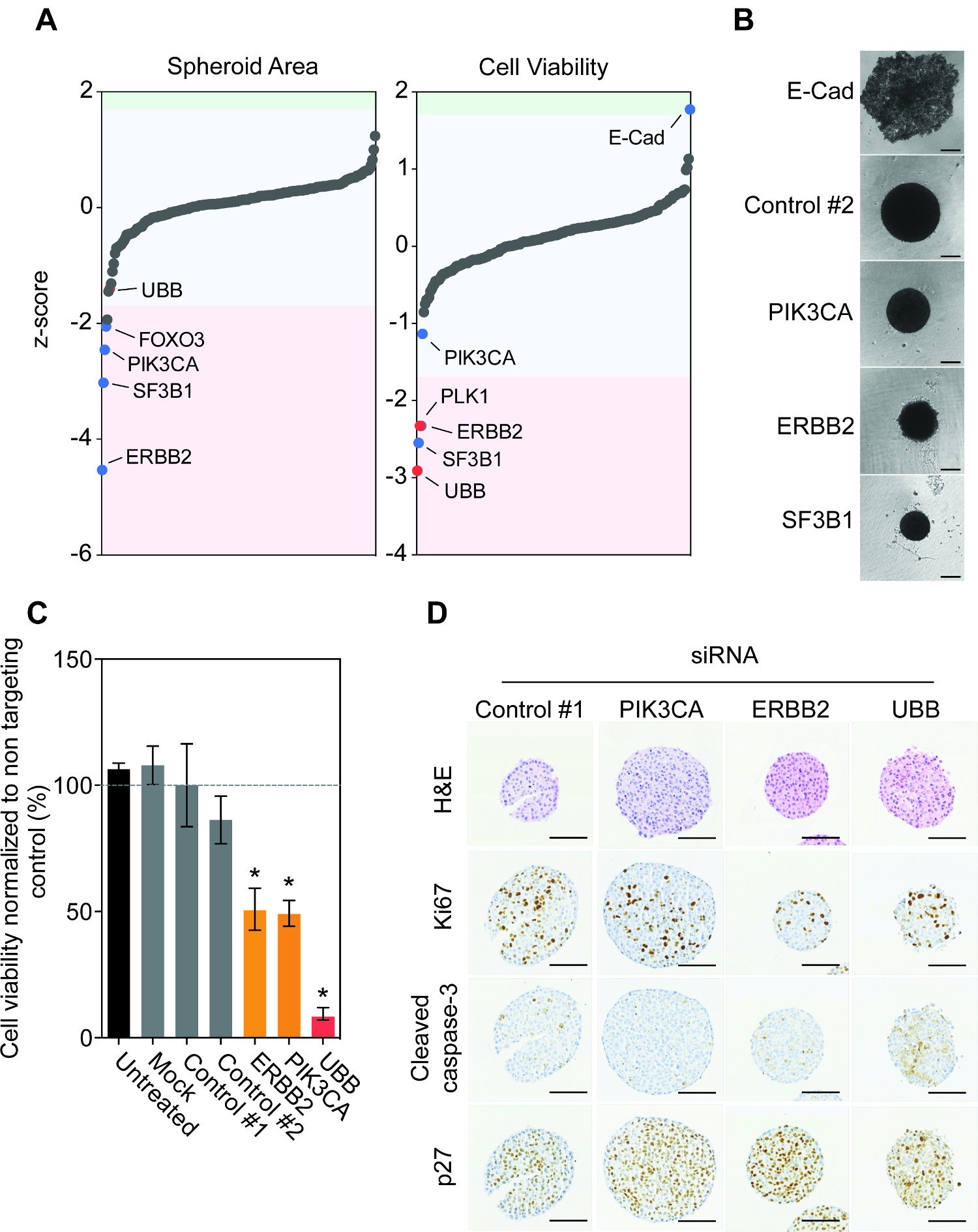
在超低附着96-孔板球体测定提供在该更容易地概括在体内的肿瘤中发现的生理条件的上下文潜在致癌高通量的表型评估。事实上,癌细胞系MCF10DCIS.com和BT474形式紧球体结构( 图1A)和球体部分的免疫组织学调查表明在细胞和核形态截然不同的空间变化。随着时间的推移,有些球体如BT474球体开发坏死区,攻击实体肿瘤( 图1B)的一个共同特点。一些球状体不发展坏死核心,如MDA-MB-231细胞系,但这样做在增殖标记Ki67的,它反比裂解caspsase-3表达,细胞凋亡的标志物( 图1C)相关的显示标记的变化。以建立多个细胞系是确实雷杰普略去对的siRNA介导的基因沉默的BT474,MCF10DCIS.com,MDA-MB-231和JIMT1细胞反向用siRNA转染的七天。转染试剂(模拟)或对照siRNA转染的存在下对球状体存活力没有效果,而在测试的所有细胞系( 图1D)沉默必需基因泛素B(UBB)显著减少球体生存能力。
我们设计了包含在未选择的最常突变的基因,ER +,HER2 +和三阴性乳腺癌人类的siRNA库。这个库由已知功能的基因,如MYC,PIK3CA和TP53和那些贡献致癌是不成立的。屏幕还包含几个非靶向对照siRNA(对照#1,对照#2)和siRNA的靶向必需基因,例如PLK1和UBB,它作为杀对照( 表1)。我们选择使用乳房CANC儿细胞系BT474,因为它们容易形成不加入重组基底膜的球状体,建立主力细胞系和具有已知的基因组结构。例如,BT474细胞是阳性对雌激素受体(ER +),过表达人类表皮生长因子受体2(HER2 +)和海港突变TP53(E285K)和PIK3CA(K111N)13。
采用以上概述的协议中,我们监测了冲击基因耗竭对球体的尺寸和生存能力七天后的siRNA的反向转染的( 图1E)。有趣的是,大多数的基因并没有对球体区域或活力( 图2A)一个显著效果。 FOXO3,PIK3CA,ERBB2和SF3B1的沉默导致球体大小的最显著重现性减少。这种减少也有人在球体的可行性后观察ErbB2和SF3B1沉默。令人鼓舞的是,我们的confirmed的PIK3CA,ERBB2和SF3B1使用明场显微镜( 图2B)中球体大小沉默的影响。我们先前确定SF3B1在众多细胞系模型必需基因,从而siSF3B1代表除了UBB 14良好的杀灭控制。有趣的是,所有的200个基因的仅E-钙粘蛋白的沉默导致球体活力( 图2A)一个显著增加。球体形态的调查显示,E-钙粘蛋白沉默导致球体结构的彻底破裂,用活细胞停留在低附着孔的底部( 图2B)。球体体积屏幕数据的手动重新调查表明,这也已经观察到,但已经从区域量化淘汰,由于其目标是高于设定大小限制。正如前面所强调的,BT474细胞过度表达受体酪氨酸激酶HER2和在港的致癌基因突变 PIK3CA(K111N)。我们证实ERBB2和PIK3CA的那沉默导致降低的球体可行性,同时用非靶向对照转染均无影响( 图2C)。
接下来我们调查的siRNA消耗对球体组织学的影响。 BT474球状体反向与非靶向对照siRNA和siRNA的靶向PIK3CA,ERBB2和UBB转。 ErbB2和UBB的沉默导致减少亲增殖标记Ki67的比较对照siRNA( 图2D)的。促凋亡标记物的活化裂解的caspase-3 UBB沉默之后才观察到,这表明HER2和PIK3CA的该耗尽不导致细胞凋亡,但均抑制细胞生长的,而非细胞毒性的。的确,HER2和PIK3CA的沉默也导致增加相比,控制转球体的细胞周期阻滞的蛋白的p27蛋白的表达。
ENT"FO:保持-together.within页="1">总之,这些结果表明BT474细胞由致癌HER2和PIK3CA当作为三维球状体生长信令从动更重要的是,这些结果表明,这是可能的。设计和实施数百个基因在癌细胞系球体稳健和可重复的定制的siRNA筛选库。
图1:3维生长细胞系优化。 答乳腺癌细胞系,MCF10DCIS.com和BT474均低装接板培养7天。使用倒置显微镜拍摄明代表性图像。比例尺= 100微米。 B. MCF10DCIS.com和BT474球状体28天培养。 4天 - 新鲜培养基100μL的每3补充。球状体固定在3.8%的甲醛,Embedded,切片,用苏木精和伊红(H&E)染色。代表图像在低和高放大倍数示出。比例尺分别代表100微米和33微米。 C. MDA-MB-231球状体21天培养。 4天 - 新鲜培养基100μL的每3补充。球状体被固定在3.8%的甲醛,包埋,切片,并与Ki67染色和裂解的caspase-3。代表图像被示出。比例尺= 100微米。 D. BT474,MCF10DCIS.com,MDA-MB-231,和JIMT1细胞系反向故作(仅转染试剂)和对照siRNA和UBB转染,超低附着板然后纺成球状体。天1和4 7天后,加入新鲜培养基(100微升),细胞存活率进行定量。数据代表平均值±一式三份标准化控制#1中执行两个独立的生物学重复的标准差。统计显着性使用UNPA计算IRED学生t检验(P <0.05)。 E.的流程图总结了用于在功能上审问癌症细胞系的基因球体的依赖反向转协议。 请点击此处查看该图的放大版本。
{kind=link}

图2:BT474 球体揭示致癌依赖的功能基因组研究。 A. BT474细胞与反一式三份200人的基因的siRNA siGENOME转库。观察球体的大小和生存能力。原始数据值分别为板中位数标准化和z值分别计算以确定显著离群大于1.7倍大于该板中位数15的标准偏差。注意边远基因ERBB2,SF3B1,PLK1和UBB一ND E-cad的。 siRNA来显著增加或减少球体面积和生存能力都在阴影蓝色和红色,其中红色描绘了对照siRNA的。 B.细胞反向与非靶向对照,E-钙粘蛋白(E-CAD),PIK3CA,ERBB2或UBB siRNA转,然后在超低附着板纺成球状体。 7天之后,明代表球体图像使用倒置显微镜拍摄。比例尺= 100微米。 C.细胞反向与非靶向对照,PIK3CA,ERBB2或UBB siRNA转,然后在超低附着板纺成球状体。 7天之后,细胞生存力进行定量。数据代表平均值±一式三份标准化控制#1中执行两个独立的生物学重复的标准差。统计显着性使用非配对学生t检验(P <0.05)来计算。 D. 7天后,球状体固定,包埋,切片,染色为H&E,Ki67的,裂解的caspase-3和p27。代表图像被示出。比例尺= 100微米。注意siControl球体的H&E由于选择了染色加工和个人完整的球体的假象显得更小。然而,这并没有从扫描图像和细胞活力结果观察到的变化得出。 请点击此处查看该图的放大版本。
{kind=link}
| 1 | 2 | 3 | 4 | 五 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 非定向1 | TP53 | DST | KMT2C | 未处理1 | FCGBP | ARID1B | FBXM7 | TTC40 | 非瞄准2 | ||
| GATA3 | TTN | MUC12 | MUC4 | AHNAK | HUWEI1 | DNAH11 | ITPR2 | ABCA13 | CREBBP | ||
| MAP2K4 | PIK3CA | F5 | 载脂蛋白B | ANKRD30A | MUC17 | DNAH17 | LAMA2 | 高手 | CSMD2 | ||
| STARD9 | USH2A | FAT3 | LPR2 | CSMD3 | MYO18B | DNAH5 | MDN1 | ARHGAP5 | DNAH9 | ||
| CXCR3 | MUC16 | RB1 | PKHD1L1 | DNAH2 | SYNE2 | DYNC1H1 | PCLO | CACNA1B | ERBB2 | ||
| PLK1 | SYNE1 | LYST | PTEN | SPTA1 | 未处理2 | VHL | RYR1 | COL6A3 | UBB | ||
| 1 | 2 | 3 | 4 | 五 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 非定向1 | ENAM | RYR2 | BRCA2 | 未处理1 | FMN2 | HECW1 | LAMB4 | SI | 非瞄准2 | ||
| FHOD3 | MACF1 | RYR3 | C2ORF16 | DMD | FRG1 | HERC2 | MYH11 | STAB1 | ZDBF2 | ||
| GOLGA6L2 | 鼻 | SMG1 | CACNA1E | DNA14 | GCC2 | HIVEP2 | NIPBL | TANC1 | ZNF536 | ||
| HMCN1 | NF1 | UBR5 | CACNA1F | DYNC2H1 | GON4L | HYDIN | PKD1L1 | TF | ANK3 | ||
| HRNR | OBSCN | USP34 | CYMA5 | FAM208B | GPR112 | ITSN2 | RNF213 | TPR | ASPM | ||
| PLK1 | PCDH15 | XIRP2 | COL7A1 | FLG2 | 未处理2 | VHL | SAGE1 | UNC80 | UBB | ||
| 1 | 2 | 3 | 4 | 五 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 非定向1 | DCHS2 | MUC5B | ZFHX4 | 未处理1 | MYO9A | SPHKAP | CXORF22 | NCOR1 | VPS13D | ||
| ATR | DMXL2 | MXRA5 | ANK2 | KIAA1210 | PRUNE2 | TCHH | DANH6 | NOTCH2 | ANKRD12 | ||
| BIRC6 | DNAH10 | TENM1 | 自动取款机 | LRP1 | SCN10A | VPS13C | DNAH7 | SPEN | C5ORF42 | ||
| CDH1 | DNAH3 | PEG3 | DIDO1 | MAP1A | SCN2A | IPTR3 | ERBB3 | SRRM2 | CCDC88A | ||
| CUBN | NOCK11 | RELN | DNAH8 | MED12 | SHROOM2 | CEP350 | FAT4 | SZT2 | CHD4 | ||
| PLK1 | 余仁生 | SACS | KIAA1109 | MED13 | 未处理2 | VHL | KMT2A | VPS13A | UBB | ||
| 1 | 2 | 3 | 4 | 五 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 非定向 | QSER1 | ARID1A | WDFY3 | 未处理1 | SDK1 | TEX15 | LAMA1 | 非瞄准2 | |||
| COL14A1 | SHROOM3 | ATRX | EFCAB5 | SF3B1 | CBFB | AHNAK2 | |||||
| CSMD1 | TBX3 | KIAA0947 | FOXA1 | ITPR1 | DDX3X | KIF4A | |||||
| MEFV | UBR4 | MYCBP2 | INPPL1 | FLG | HECTD4 | FAT2 | |||||
| MGAM | VCAN | NBEAL1 | MAP3K1 | AKAP9 | GPR98 | FOXO3 | |||||
| PLK1 | ZNF462 | SETX | NRP1 | HERC1 | 未处理2 | VHL | UBB | ||||
表1:平板布局和人类的siRNA库去卷积。表中包含的每个所述siRNA池和布局在屏幕中使用的低附着板的内容。
讨论
癌症三维模型正在越来越多地用于评价已设计成选择性地杀死癌细胞已知和新的化合物的功效。癌细胞球体是显示更类似于在体内肿瘤遇到的条件的结构,因此,在三维增加效力的化合物更可能在体内的效果。然而,这些方式不允许的,但没有药物设计的主题,可以在治疗癌症相当功效潜在的新的目标的识别。
我们开发了允许持久的基因沉默在癌症细胞系球体多达七天的siRNA功能基因组学的方法。存在需要优化可以执行的siRNA屏幕之前到协议几个关键步骤。以形成大规模可再现可行球状体的能力是必不可少的。而且,一个ppropriate转染条件应严格优化。我们建议试运行几个不同的转染试剂与适当的非定位和之前试图屏幕杀人控制。我们能够显示,几种常用的乳腺癌细胞系,即BT474,MCF10DCIS.com,MDA-MB-231和JIMT1分别适合于siRNA转染。此外,我们提供证明的原则数据筛查乳腺癌200最频繁的突变基因在BT474球体,确认了他们对HER2的扩增和PIK3CA致癌基因突变的依赖性。有趣的是,转录因子FOXO3的沉默导致球体尺寸的减小,但是对存活率没有显著效果。 FOXO3已知调节缺氧的响应,改变肿瘤细胞的代谢能力使它们更容易地适应环境16。这种作用可能与细胞活力的读数,因为它检测到大量的ATP,细胞代谢的主要产品之一。
为了支持在球体尺寸的减小的观察,它已被证明FOXO3在HeLa该沉默异种移植物受损肿瘤生长和诱导细胞凋亡17。要注意的是一些基因可能影响癌细胞保留其三维结构,这可能导致假阳性的能力是很重要的。例如,E-钙粘蛋白的敲低导致BT474球体结构的溶解。这不得不使用E-钙粘蛋白抗体靶向18先前的报道。与任何筛选平台,潜在的目标应当再次筛选,以评估观察到的效果的再现性。有该技术,即该siRNA介导的基因敲低的瞬时性质的限制。持续更长的沉默超过七天没有用siRNA实现的。
这种方法的优点是,它可以与各种其它生物群落被耦合TRIC染料而不仅仅是那些评估球体的可行性,例如,给球体缺氧的空间信息或监测细胞发生凋亡。此外,由于板读取器的扫描是相对快速和非侵入性的,siRNA对球体大小的影响可以在一段时间,而不是仅仅在实验的终点评估。事实上,我们目前正在探索多种途径,这对我们的筛选管道中。一种利用3D培养,以确定新的相关性的另一种方法是使用抑制或广泛的目标或蛋白质的特定家庭化学库。事实上,Bitler 等人 。利用这种有针对性的方式来确定卵巢透明细胞癌19 ARID1A地位和EZH2抑制剂的合成致死互动。 CRISPR-Cas9基因编辑技术的发现还允许在类器官培养物和体内遗传筛选的发展。然而,T他的方法是依赖于具有适当的动物设施,并且可以成本高昂20。
总之,我们认为,我们已经提出了一个协议,更准确的模型中的氧和营养物梯度,它们是在体内肿瘤微环境的特征,允许的新型癌症靶或既定目标的健壮验证的鉴别。此外,我们的协议可以被应用到任何类型的形成球状体,因此可以在癌症研究界为高通量的siRNA屏幕常规使用细胞系。
披露声明
Open Access fees were supported by Nexcelom Bioscience, LLC.
致谢
The authors would like to thank Fredrik Walberg (FACS and imaging facility, ICR) for assistance with microscopy. This work was funded by Breast Cancer Now. RN is the recipient of a Breast Cancer Now Career Development Fellowship (2011MaySF01).
材料
| Name | Company | Catalog Number | Comments |
| Lullaby | Oz Biosciences | LL70500 | lipid-based transfection reagent |
| Viromer | Lipocalyx | VB-01LB-01 | virus-like polymer transfection reagent |
| Ultra-low attachment plate | Corning | CLS7007 | 96 well plate |
| Foil plate seals | ThermoFisher | AB-0626 | |
| Luminescent cell viability dye | Promega | G7570 | CellTitre-Glo |
| Pipette tips (200 μL) | Starlab | S1111-0806 | |
| Pipette tips (10 μL) | Starlab | S1111-3800 | |
| Pipette tips (1, 000 μL) | Starlab | S1122-1830 | |
| Serological pipettes (5 mL) | Sarstedt | 86.1253.025 | |
| Serological pipettes (10 mL) | Sarstedt | 86.1254.025 | |
| Serological pipettes (25 mL) | Sarstedt | 86.1685.020 | |
| RPMI Media | GIBCO | 11875-093 | |
| DMEM Media | GIBCO | 11965-084 | |
| Opti-MEM | GIBCO | 31985070 | |
| Feta bovine serum | GIBCO | 16140063 | |
| siRNA | Dharmacon | Cherry picked library | |
| Countess Cell Counter | ThermoFisher Scientific | AMQAX1000 | |
| Cell counting chamber slides | ThermoFisher Scientific | C10312 | |
| Celigo S | Nexcelom | contact company | |
| Victor X5 | Perkin Elmer | contact company | |
| Benchtop centrifuge | Various | ||
| Axiovert Inverted brightfield microscope | Zeiss | contact company | |
| Tissue culture CO2; Incubator | Various | ||
| Mulitichannel pipette | Various |
参考文献
- Seton-Rogers, S. E., et al. Cooperation of the ErbB2 receptor and transforming growth factor beta in induction of migration and invasion in mammary epithelial cells. Proc Natl Acad Sci U S A. 101 (5), 1257-1262 (2004).
- Debnath, J., Muthuswamy, S. K., Brugge, J. S. Morphogenesis and oncogenesis of MCF-10A mammary epithelial acini grown in three-dimensional basement membrane cultures. Methods. 30 (3), 256-268 (2003).
- Chia, S. K., et al. Prognostic significance of a novel hypoxia-regulated marker, carbonic anhydrase IX, in invasive breast carcinoma. J Clin Oncol. 19 (16), 3660-3668 (2001).
- Trastour, C., et al. HIF-1alpha and CA IX staining in invasive breast carcinomas: prognosis and treatment outcome. Int J Cancer. 120 (7), 1451-1458 (2007).
- Wilson, W. R., Hay, M. P. Targeting hypoxia in cancer therapy. Nat Rev Cancer. 11 (6), 393-410 (2011).
- Ros, S., et al. Functional metabolic screen identifies 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4 as an important regulator of prostate cancer cell survival. Cancer Discov. 2 (4), 328-343 (2012).
- Baenke, F., et al. Functional screening identifies MCT4 as a key regulator of breast cancer cell metabolism and survival. J Pathol. 237 (2), 152-165 (2015).
- Schug, Z. T., et al. Acetyl-CoA synthetase 2 promotes acetate utilization and maintains cancer cell growth under metabolic stress. Cancer Cell. 27 (1), 57-71 (2015).
- Mashimo, T., et al. Acetate is a bioenergetic substrate for human glioblastoma and brain metastases. Cell. 159 (7), 1603-1614 (2014).
- Comerford, S. A., et al. Acetate dependence of tumors. Cell. 159 (7), 1591-1602 (2014).
- Vinci, M., Box, C., Eccles, S. A. Three-dimensional (3D) tumor spheroid invasion assay. J Vis Exp. (99), e52686 (2015).
- Vinci, M., Box, C., Zimmermann, M., Eccles, S. A. Tumor spheroid-based migration assays for evaluation of therapeutic agents. Methods Mol Biol. 986, 253-266 (2013).
- Barretina, J., et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature. 483 (7391), 603-607 (2012).
- Maguire, S. L., et al. SF3B1 mutations constitute a novel therapeutic target in breast cancer. J Pathol. 235 (4), 571-580 (2015).
- Brough, R., et al. Functional viability profiles of breast cancer. Cancer Discov. 1 (3), 260-273 (2011).
- Ferber, E. C., et al. FOXO3a regulates reactive oxygen metabolism by inhibiting mitochondrial gene expression. Cell Death Differ. 19 (6), 968-979 (2012).
- Jensen, K. S., et al. FoxO3A promotes metabolic adaptation to hypoxia by antagonizing Myc function. EMBO J. 30 (22), 4554-4570 (2011).
- Ivascu, A., Kubbies, M. Diversity of cell-mediated adhesions in breast cancer spheroids. Int J Oncol. 31 (6), 1403-1413 (2007).
- Bitler, B. G., et al. Synthetic lethality by targeting EZH2 methyltransferase activity in ARID1A-mutated cancers. Nat Med. 21 (3), 231-238 (2015).
- Dow, L. E., et al. Inducible in vivo genome editing with CRISPR-Cas9. Nat Biotechnol. 33 (4), 390-394 (2015).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。