
Method Article
Proteína de filme infravermelho eletroquímica demonstrada por estudo de oxidação de2 H pela hidrogenase [NiFe]
* Estes autores contribuíram igualmente
Neste Artigo
Resumo
Aqui, descrevemos uma técnica, eletroquímica infravermelho da filme de proteína, que permite que as proteínas redox imobilizado ser estudado espectroscopicamente sob o controle direto de eletroquímica de um eletrodo de carbono. Espectros de infravermelhos de uma amostra única proteína podem ser gravados em uma variedade de potenciais aplicadas e sob uma variedade de condições de solução.
Resumo
Noções básicas sobre a química dos métodos de demandas de proteínas redox que fornecem controle preciso sobre redox centros dentro da proteína. A técnica de eletroquímica de filme de proteína, em que uma proteína é imobilizada em uma superfície de eletrodo, tal que o eletrodo substitui doadores de elétron fisiológicos ou aceitadores, forneceu insights funcional sobre as reações redox de uma gama de diferentes proteínas. Compreensão plena química requer controle eletroquímico para ser combinado com outras técnicas que podem adicionar uma visão mecanicista e estrutural adicional. Aqui vamos demonstrar uma técnica, eletroquímica infravermelho da película de proteína, que combina eletroquímica de filme de proteína com amostragem de espectroscopia infravermelha de proteínas redox. A técnica utiliza uma geometria de reflectância total atenuada do múltiplo-reflexão para sondar uma proteína redox imobilizada em um eletrodo de carbono preto alta área de superfície. Incorporação deste eletrodo em uma célula de fluxo permite que o pH da solução ou concentrações de soluto a ser alterado durante as medições. Isto é particularmente forte no endereçamento de enzimas redox, onde ritmo acelerado catalítico pode ser sustentado e controlado no eléctrodo que permite a observação espectroscópica de long-lived espécies intermediárias no mecanismo catalítico. Demonstramos a técnica com experiências em Escherichia coli hidrogenase 1 em condições de não-volume de negócios e volume de negócios (oxidação de2 H).
Introdução
Um desafio fundamental no estudo da função da proteína envolve o desenvolvimento de métodos em situ que permitem a observação direta de proteínas realizando suas funções fisiológicas, in vivo ou usando isolado amostras da proteína. Isto requer a integração de controle ou desencadear processos em procedimentos experimentais e a utilização de técnicas combinadas que permitem tanto a reatividade ser avaliada e etapas químicas individuais durante a função da proteína a ser medido, simultaneamente. No caso de proteínas redox isso muitas vezes equivale a combinando técnicas de eletroquímicas, que precisamente controlam o potencial aplicado, mas não fornecem nenhuma informação química direta, com técnicas espectroscópicas que são sensíveis a quimicamente específicos mudanças associadas com a função da proteína. 1 , 2 , 3 Spectroelectrochemistry é um termo geral para uma variedade de métodos eletroquímicos e espectroscópicos em acoplado que abrangem uma variedade de técnicas espectroscópicas e níveis de controle eletroquímica. Muitas proteínas podem trocar elétrons com artificial, solúvel de elétrons doadores e aceitadores e isto tem sido explorado em estudos que utilizam pequenas moléculas para mediar a transferência de elétrons, incluindo o acoplamento com UV-visível,4,5 , 6 , 7 Dicroísmo circular magnético8 e infravermelho5,9,10,11,12,13,14 (IR) espectroscopia. Em um número limitado de casos revelou-se possível explorar a troca de elétrons sem intermediação, controlado por difusão entre proteínas e eletrodos. 15 , 16
Para reações catalíticas realizado por enzimas redox, abordagens de eletroquímica solução presente uma clara desvantagem. Difusão-controlado elétron transferência via redox mediadores em solução é provável tornar-se limitar a taxa. Cinéticas e mecanicistas informações sobre a enzima podem ser perdido, ou pelo menos tornar-se difícil para deconvolute de artefatos de difusão resultante do método experimental. Controle de eletroquímico direto, sem intermediação, portanto, é uma importante ferramenta para o estudo de enzimas e proteínas redox. A técnica de eletroquímica de filme de proteína (PFE) emprega proteínas imobilizadas de eletrodo redox, de tal forma que os electrões são transferidos diretamente para ou a partir de cofactores redox dentro a proteína como o eletrodo é polarizado em uma série de potenciais. 17 , 18 , 19 PFE é de particular valor para o estudo de reações de oxidação ou redução catalisada por enzimas redox, como transferência de elétrons interfacial pode ser alcançada em uma taxa muito elevada. Por exemplo, a taxa de rotatividade de electrocatalytic da hidrogenase ([NiFe]) de níquel-ferro de Allochromatium vinosum foi medida pela PFE como ca 1.000-10.000 s− 1 para oxidação de2 H. 20 o potencial de eletrodo age como um gatilho para transformar a catálise 'on' ou 'off' e os relatórios de electrocatalytic atual na atividade da enzima. PFE é, portanto, um valioso método para analisar a reatividade de enzimas complexas que dependem intimamente potencial, tais como reações de sítio ativo di-Ferro de [FeFe]-hidrogenases com CO e O2,21 ou induzida em potencial reações de inactivação de hidrogenases, desidrogenase de monóxido de carbono de22 ,23 e outras enzimas redox do complexo. 24
A principal barreira ao combinar técnicas espectroscópicas, com o controle de eletroquímico direto oferecido pela PFE decorre da baixa cobertura de superfície de enzimas redox, na ordem de 1-2 pmol cm-2 para a hidrogenase [NiFe] do a. vinosum, 20 em relação em situ estudos de ciência de superfície de adsorbates pequena molécula em eletrodos de metal em massa. Isso apresenta um desafio para a sensibilidade da medição espectroscópica. Vários métodos de spectroelectrochemical têm sido relatados para estudar imobilizada proteínas redox em uma variedade de eletrodos diferentes: espectroscopia UV-visível em eletrodos transparentes de óxido metálico; 25 , 26 , espectroscopia de fluorescência 27 em eletrodos de ouro; 28 , espectroscopia de absorção de infravermelho superfície reforçada (SEIRA) 29 em eletrodos de ouro; 30 , 31 , 32 , 33 e superfície reforçada técnicas de Espectroscopia Raman, principalmente em eletrodos de prata. 34 , 35
Aqui nós descrevemos um método para acoplamento PFE com espectroscopia de IR, em uma técnica conhecida como eletroquímica infravermelho do filme proteína (PFIRE). 36 o método the PFIRE estuda enzimas redox imobilizadas em um eletrodo de trabalho de carbono de alta área de superfície em conjunto com uma geometria de IR (ATR-IR) de reflectância total atenuada, explorando a facilidade de adsorção de uma gama de proteínas em superfícies de carbono. Espectroscopia de IR é útil em estudos de proteínas e enzimas redox como muitas moléculas pequenas, ligantes e cofactores têm absorvâncias de diagnósticos que podem ser usadas para avaliar o estado de reatividade, vinculação, inibição e redox. Exemplos de ligação de n de centros ferro-enxofre,37 estudo de flavoproteínas,38,39,40 pequena molécula vinculando a hemo centros etc. 41 o ATR-IR geometria permite a construção de uma célula eletroquímica de três-elétrodo otimizado (spectro)42 e, portanto, fornece controle excelente da eletroquímica. Resistência de solução e potencial deriva são minimizadas pela colocação de um eletrodo de referência perto do eletrodo de trabalho. Eletrodos de contador de alta área de superfície são usados que são compatíveis com as correntes de alta electrocatalytic produzidas pela rotatividade de enzima rápido para o eletrodo de trabalho. Fluxo de solução através da célula de spectroelectrochemical permite fácil controle sobre a concentração de substratos, inibidores e pH. 36 , 43 , 44 PFIRE o método, portanto, permite que a espectros de IR ser gravado em situ durante electrocatalysis enzima sustentado. 36 , 44 PFIRE também é capaz de fornecer informações químicas na ausência de catalisador atual,43 em contraste com PFE onde pode ser difícil extrair informações de processos não-catalítica de enzimas redox. 45 , 46
Nós demonstramos o método PFIRE para estudo de oxidação de2 electrocatalytic H por hidrogenases [NiFe], que contêm intrínseca CO e CN ligantes coordenados a fé a um sítio activo bimetálico. 36 , 43 , 44 [NiFe] hidrogenases, portanto, são particularmente adequados para estudar por PFIRE. O método PFIRE fornece informações sobre as espécies que estão presentes durante o volume de negócios de estado estacionário e, portanto, fornece uma visão mecanicista crucial, além da riqueza da literatura em espectroscopia de IR de hidrogenases sem controle experimental sobre o volume de negócios. 47 , 48 Dyer e cooperadores têm empregado tempo-resolvido métodos de IR estudar de NiFe hidrogenases,49,50,51 usando um disparador de luz também aplicar um pequeno passo de potencial negativo (através do uso de mediadores de solução e uma fonte de elétrons 'engaiolado') ou photolyse um hidreto acoplado. Embora o método PFIRE não pode, neste momento, fornecer resolução de tempo para combinar com as medições,40 , que permitem o estudo dos processos catalíticos redutoras e oxidativos, acessado em uma gama de potenciais bem definidos e livre de transporte em massa limitações.
O método PFIRE é distinto de estudos SEIRA de proteínas redox, que também usam uma geometria ATR-IR e empregam um eletrodo de metal nanoescala-áspera para melhorar a absorção de IR de moléculas adsorvidas na superfície do eletrodo. 30 SEIRA é uma técnica extremamente valiosa para o estudo de proteínas de membrana, em especial adsorvidas em ou dentro da membrana mimética arquiteturas,32 , mas a necessidade de um eletrodo de metal pode limitar o escopo de substrato e inibidor devido a reatividade o suporte de eletrodo para moléculas pequenas tais como CO, CN–, CO2 etc. Redução de próton e dessorção monocamada auto montado podem ser problemáticos em superfícies de metal em potenciais muito negativos,1,52 embora enzima electrocatalysis na desprotegido eletrodos de metal tem sido relatado. 53 , 54 uma desvantagem de PFIRE em relação a SEIRA é a relativa dificuldade de incorporação de proteínas de membrana em arquiteturas de membrana nativo ou mimética. No entanto, a relativa inércia química dos eléctrodos de carbono para reacções de ativação pequena molécula concorrentes faz PFIRE uma excelente técnica para o estudo da enzima electrocatalysis, particularmente no domínio de baixo potencial redox biológica relevante processos, tais como redução de prótons por hidrogenases. 1 , 43
O objetivo deste artigo é apresentar o método PFIRE como uma técnica para o estudo de proteínas redox eletrodo-imobilizado, usando NiFe hidrogenase 1 (Hyd1) de Escherichia coli como um exemplo. Considerações da preparação da amostra, a exigência de substrato bom fluxo e manipulação de dados são discutidas. PFIRE é uma técnica amplamente aplicável, well-suited ao estudar qualquer proteína redox (com características absorvâncias IR) que pode ser adsorvida em eletrodos de carbono, ou diretamente ou usando modificação da superfície, de tal forma que ele pode trocar elétrons com o eletrodo.
Protocolo
1. recriando o compartimento interno de amostra de um espectrômetro FTIR dentro de uma caixa de luvas anaeróbica, seca e requisitosgerais Experimental
- Use um espectrômetro FTIR comercial, equipado com um detector de Telureto (MCT) externo de mercúrio cádmio e acessório ATR. Limpar o corpo do aparelho com ar seco ou dinitrogênio, ou alternativamente, usar um espectrômetro com um banco óptico evacuado.
- Coloque o espectrômetro sobre uma mesa estável, livre de vibração à mesma altura como um anaeróbio (< 1 ppm O2), secar o porta-luvas (ponto de orvalho <-75 º C). Desvie o feixe do espectrómetro de IR para o porta-luvas, através de uma janela transparente de IR suficientemente grande para acomodar o feixe desfocado.
Nota: É importante que o espaço entre o porta-luvas e espectrômetro também é purgado ou evacuado, especialmente se um material higroscópico janela, tais como NaCl ou KBr, por exemplo, é usado. -
Replica o compartimento de amostra interna do aparelho dentro do porta-luvas.
- Direcionar o feixe dentro do porta-luvas para um espelho elipsoidal fora do eixo, idealmente com o mesmo comprimento focal como o espelho de focagem interno do espectrómetro.
Nota: É útil ter dois espelhos de avião adicional colocados antes este espelho de focagem, a fim de permitir que tanto a altura e a direção do feixe de IR ser ajustadas dentro do porta-luvas; Isto irá compensar qualquer imprecisão na colocação inicial do espectrómetro. - Colocar um detector MCT externo, completo com um apropriado focagem óptica (como uma lente de distância focal curta ZnSe ou espelho parabólico fora do eixo, com uma distância focal curta) dentro do porta-luvas, posicionado como replicar as dimensões da amostra interna compartimento do espectrómetro.
- Ajuste o 'foco' do feixe IR para ser aproximadamente equidistante entre o espelho de focalização e o detector do MCT.
- Direcionar o feixe dentro do porta-luvas para um espelho elipsoidal fora do eixo, idealmente com o mesmo comprimento focal como o espelho de focagem interno do espectrómetro.
- Legal o dewar do detector MCT externo com nitrogênio líquido.
- Monte o acessório ATR no compartimento de amostra do porta-luvas. Alinhe a entrada e saída de focagem óptica e acessório ATR para atingir a taxa de transferência máxima para o detector MCT. Se necessário, utilize uma abertura ou outra atenuação apropriada (por exemplo, uma grade de arame) para evitar supersaturação do sinal do detector.
Nota: para os dados apresentados aqui, um acessório ATR de cinco-reflexão com todos os sistema ótico reflexivo e um Si trapezoidal removível, elemento de reflexão interna (IRE) é usado (Crystal GmbH, dimensões ca 5 × 8 × 1 mm3, ângulo rosto 39,5 °). A ira é montada em uma placa removível base usinado a partir de poliéter éter cetona (PEEK). - Use uma caixa de luvas para abrigar o compartimento de amostra recriada com feedthroughs adequados para permitir a conexão de um potentiostat alojados externamente (conexões BNC apertadas de gás são úteis para essa finalidade), acesso a gás e cabos para transferir o sinal do MCT detector. É importante reter qualquer blindagem do cabo do detector para evitar a introdução de ruído ou interferência para as medições.
- Coloque uma bomba peristáltica, capaz de taxas de fluxo maiores que 60 mL/min dentro do porta-luvas. Uma visão esquemática do espectrómetro, ATR acessória, peristáltica bomba e potentiostat de configuração é mostrada na Figura 1.
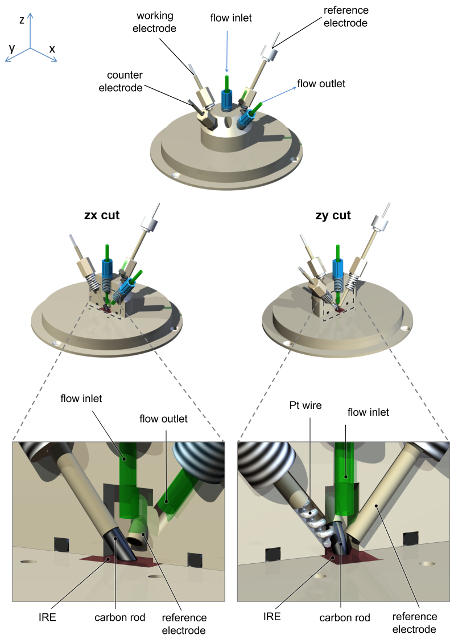
- Construa uma célula de spectroelectrochemical, como mostrado esquematicamente na Figura 2, que sela para a ATR acessório. A célula deve conter um eletrodo de contador de alta área de superfície (Pt fio ou gaze), uma conexão para o eletrodo de trabalho (haste de carbono) e um eletrodo de referência em miniatura. A célula deve conter pelo menos duas portas adicionais como entrada e saída para o fluxo rápido de soluções através da célula.
Nota: Um eléctrodo de referência de calomelano saturado em miniatura é ideal para esta finalidade. 36
2. preparação de partículas de negro de carbono modificado com Escherichia coli Hydrogenase 1
- Em uma caixa de luvas anaeróbica 'molhada', misture 20 mg de alta área de superfície partículas de negro de carbono (> 1.000 m3/g) com 1 mL de água de ultra alta pureza (resistividade > 18 cm MΩ) em um tubo de microcentrifugadora. Dispersar as partículas por sonication de baixa potência (< 100 W) pelo menos 15 min, ou até que a dispersão é uniforme e faz não sedimentos dentro de 1h, para dar uma dispersão de negro de carbono com uma carga de cerca de 20 mg/mL.
-
Tomar uma alíquota de Escherichia coli hidrogenase 1 (Hyd1, aproximadamente 50 µ l, preparada de acordo com um procedimento publicado55) em uma concentração de ~ 7 mg proteína/mL e trocá-lo em um buffer de baixa força iônica em um pH próximo ao ponto isoelétrico (para exemplo fosfato de potássio, 15 mM, pH 5.8, sem sal adicional). Para obter melhores resultados, use uma preparação enzimática é tão ativa quanto possível. Realize a troca do amortecedor por concentração e diluição, em um dispositivo de filtragem centrífuga com um peso molecular apropriado corte (50 kDa funciona bem para Hyd1).
- Adicione o Hyd1 para o dispositivo de filtro.
- Dilua com cerca de 450 µ l de tampão de troca.
- Reconcentrar a um volume de 50 µ l usando centrifugação suave (< ~ 2700 × g) para evitar a precipitação irreversível.
- Repita os passos 2.2.2 e 2.2.3 até troca de amortecedor é completa (normalmente dentro de ~ 5 ciclos).
- Misture um volume de 5 µ l de 20 mg/mL de dispersão de negro de carbono (preparado em 2.1) com a alíquota de 50 µ l de tampão-trocadas Hyd1. Armazene a mistura de enzima-partícula no frigorífico (a 0 ° C) durante a noite para permitir que a enzima adsorver. Pode ser necessário agitar a mistura ocasionalmente para garantir que as partículas permanecem dispersas.
- Lave as partículas Hyd1 modificado com tampão de baixa força iônica (fosfato de potássio, 15 mM, pH 5.8, sem sal adicional) por sucessivos ciclos de sedimentação e re-dispersão microcentrifuga (~ 2.700 × g). Repita este processo 3 - 5 vezes para remover a enzima não-adsorvido.
Nota: Antes da primeira lavagem o sobrenadante deve ser praticamente incolor, indicando boa adsorção da enzima. - Concentre as partículas até um volume final de ~ 5 µ l, para dar um carregamento final ~ 20 mg/ml de partículas enzima modificada.
Nota: As partículas Hyd1 modificados podem ser armazenadas a 4 ° C por até duas semanas se permanecer hidratados; Isto é melhor conseguido armazenando partículas diluídas com ~ 50 µ l de água ultra alta pureza.
3. preparação para medições de PFIRE em e. coli Hydrogenase 1
- Limpar a ira de Si por baixa potência sonication (< 100 W), primeiro em H2então4 (< 90% w/w) para ~ 15 min e depois em HNO3 (70% w/w) para até 1 hora. Este procedimento de limpeza deve conduzir a uma superfície hidrofílica de IRE. Se mais limpeza é necessária a ira pode ser colocada em solução de piranha (uma proporção de 1:3 de H2O2: H2SO4) para oxidar qualquer material orgânico remanescente.
Nota: Métodos de limpeza ácidos duros podem não ser adequados para uso com todos os materiais IRE. Solução de piranha é altamente corrosiva e um oxidante forte, as superfícies devem estar razoavelmente limpas, antes de uso da solução de piranha e cuidados deve ser tomado durante a preparação e uso de solução de piranha, devido à natureza exotérmica do processo - sempre adicionar H2O 2 lentamente a H2SO4 e referem-se aos procedimentos de segurança adequados para preparação, utilização e eliminação. - Enxágue a ira limpa em água ultra alta pureza e secar sob um fluxo de gás nitrogênio seco. Realize qualquer manuseio de prisma, usando uma pinça limpa para evitar a contaminação.
- Sele a ira para a ATR acessória placa de base usando uma tira fina de selante de silicone de elétrico-classe. Tome cuidado para restringir o selante nas bordas da ira. Permitir que o selante secar totalmente.
- Transferir a placa de base acessória ATR para o espectrómetro de luvas e montá-lo para o acessório ATR. Medida um espectro de referência (fundo) entre 4000-1.000 cm-1 , com resolução de 4 cm-1 , usando o padrão rápido scan mode do espectrómetro e 1024 interferograms em média (medição tempo ~ 3-5 min). Use este espectro como uma entrada para permitir o cálculo de espectros de absorvância mais tarde no experimento.
Nota: Devido a pequena absorbância esperada para o sítio ativo Hyd1 é necessário coletar espectros com sinal elevado para rácios de ruído. - Transferi a placa de base acessória ATR para o glovebox 'molhado', que contém a dispersão de partícula pré-preparadas Hyd1-modificado (secção 2). Gota-elenco uma alíquota de 1 µ l da dispersão de partículas para o grande rosto da ira e espalhá-los uniformemente em toda a superfície. Nota: Não permitir que as partículas se tornar completamente seco sobre a ira.
- Corte um pedaço de papel de carbono, como que usado como material de camada de difusão de gás nas células de combustível, para um tamanho de ~0.1 mm menor do que as dimensões de superfície da ira, (ou seja, ca. 8.2 × 4,9 mm2). A área da superfície do papel carbono deve ser grande o suficiente para cobrir as gota-elenco modificado Hyd1 partículas, mas não tão grande quanto se sobrepõem o vedador do silicone usado para fixar a ira. Mergulhe o papel de carbono em água e delicadamente, coloque-o sobre o topo do filme partícula. O papel de carbono irá fornecer uma boa conexão elétrica em todo o filme toda partícula.
Nota: É benéfico para preparar vários pedaços de papel carbono com antecedência e armazená-los previamente embebido em água de pureza ultra alta dentro do porta-luvas anaeróbico 'molhado'. - Montar a célula spectroelectrochemical (descrito em 1.7 e mostrado esquematicamente na Figura 2) sobre a ira, fixá-lo na placa de base com parafusos. Adicione ~ 200 µ l de buffer experimental para manter a enzima hidratada. Um sistema de buffer misto, capaz de buferização sobre uma ampla faixa de pH, é útil para estudos sobre NiFe hidrogenases: acetato de sódio56 , 2-[N'-Morpholinos] etano-sulfônico (MES), N'-[2-hidroxietil] piperazina -N' -[2-etano-sulfônico] (HEPES), N'-[hidroximetil] ácido metil-3-amino-propano-sulfônico (torneiras) e 2-[N'-cyclohexylamino] etano-sulfônico (CHES), com cada componente em uma concentração final de 15 mM e contendo 0,1 M NaCl como suporte eletrólito, com pH ajustado a pH 6 utilizando concentrada de NaOH e HCl.
Nota: A conexão do eletrodo de trabalho deve projetar-se ligeiramente abaixo do plano do topo da célula de spectroelectrochemical (~0.1 mm) para garantir boa conexão eletrônica para o papel de carbono (Figura 2). - Conectar-se a solução de entrada e saída da célula de spectroelectrochemical para um sistema de fluxo que contém um frasco do buffer experimental e tubo de bomba peristáltica. Transferi a célula montada para o porta-luvas 'seco' contendo o espectrômetro.
- Monte o conjunto de células de spectroelectrochemical sobre o acessório ATR. Conectar-se a trabalhar, contador e eletrodos de referência para o potentiostat. Conecte o tubo da bomba peristáltica para a bomba.
- Grave um espectro de absorbância com 1024 em média interferograms na resolução de 4 cm-1 em uma faixa espectral de 4.000-1.000 cm-1, usando o espectro coletado em 3.4 como referência/fundo. Neste ponto o espectro deve conter significativa (> 100 mO.D...) amida II bandas no ~ 1.540 cm-1 e bandas de sítio ativo de Hyd1 devem ser evidentes na região espectral 1.850-2.150 cm-1, em grande parte nos Estados oxidados, inativos (Figura 3 ).
4. activação de Escherichia coli Hydrogenase 1 e teste o celular Spectroelectrochemical
- Aplica um potencial de redução (−0.8 V vs SCE) para o filme de partícula Hyd1-modificado. Sature o buffer experimental com buffer de fluxo e2 H lentamente através da célula de spectroelectrochemical. Deixe a amostra durante a noite para ativar completamente o Hyd1.
Nota: É importante usar anaeróbica H2, N2 etc.e então todos os gases anaeróbicos deve ser passado através de um filtro de2 O. - Grave um espectro de absorbância da amostra após a ativação. O νCO e vCN bandas do sítio activo devem agora mostrar uma distribuição de Estados reduzidos, 'ativos'. Este é mais facilmente observado através do uso de um espectro de diferença, em relação ao espectro gravado em 3.10 (Figura 4).
- Teste a conexão elétrica da célula spectroelectrochemical. Para fazer isso, sature o buffer experimental com gás de2 N. Aplicar uma sequência de oxidação (0 V vs SCE) e redução (−0.8 V vs SCE) potenciais (de duração de 30 min de ca ) para o filme de partícula Hyd1 modificado e gravar um espectro de absorbância em cada um. O Hyd1 deve tornar-se rapidamente oxidado e reduzido, e se o filme de partícula está bem ligado 100% da amostra deve responder ao potencial aplicado.
- Defina uma taxa de fluxo apropriado do buffer experimental. Para fazer isso, aplique um potencial de redução (−0.8 V vs SCE) à amostra e saturar o buffer experimental com H2. Gravar uma série de voltammograms cíclicas entre-0.707 - 0.039 V vs SCE a uma taxa de varredura de 10 mV/s. gradualmente aumente a taxa de fluxo de H2-saturada buffer entre voltammograms até o protector catalítico que assemelha-se em uma rotação planar disco de eletrodo55 e a corrente máxima é independente da taxa de fluxo (Figura 5).
Nota: O limite de solubilidade de H2 em água é ~0.8 mM a 293 K e 1 bar. - Como a amostra está agora pronta para medições de PFIRE, colete espectros em uma variedade de potenciais, sob uma variedade de condições de solução (pH, temperatura, concentração de2 H etc.). Grave todos os dados eletroquímicos utilizando o software potentiostat, como é importante ser capaz de correlacionar os dados espectroscópicos e eletroquímicos, especialmente quando estudar processos electrocatalytic como oxidação de2 H por Hyd1.
5. manipulação de dados espectroscópicos
- Confirme que o sítio ativo não foi permanentemente alterado no decurso das medições por gravação espectros em 0 V e −0.8 V vs SCE no final do experimento. Estes devem ser idênticos para os espectros no ponto 4.3, e sem perda de sítio ativo deve ser observada durante a medição (Figura 6).
- Espectros de absorvância absoluto de exportação a partir do software de espectrômetro em um formato adequado (CSV, ASCII, 'matlab', Jcamp etc) para processamento usando o software como origem ou Matlab.
-
Linha de base corrigir os dados, usando o processo ilustrado na Figura 7.
- Tomar a segunda derivada de cada espectro de absorbância no intervalo 1.800-2.150 cm-1, a fim de identificar pequenas (< 1 mO.D...) faixas de sítio ativo no contexto de água altamente curvado.
- Coloque pontos de marcador de linha de base no espectro de absorção do original e definir os pontos de 'encaixe' para o espectro experimental.
- Cabe uma linha de base através de pontos usando uma função spline cúbica interpolados ou uma função polinomial. Subtrai a essa função de linha de base de dados experimentais.
Resultados
A Figura 1 mostra uma representação esquemática do arranjo experimental do sistema de fluxo espectrômetro, porta-luvas, acessório ATR, potentiostat e gás utilizado para medições de PFIRE. A Figura 2 mostra um representante desenho da célula spectroelectrochemical.
A Figura 3 mostra os espectros de absorvância de gota-elenco partículas Hyd1-modificado, com a reserva experimental (um sistema de buffer misto, descrito em 3,7, pH 6.0) flui através da célula de spectroelectrochemical. A cobertura de superfície de Hyd1 é particularmente elevada no exemplo mostrado na Figura 3, com uma intensidade de banda Amida II de mO.D ~ 235. e mínima 'em massa' água como evidenciado pela magnitude da região de estiramento O-H (~ 3.000-3.600 cm-1) em relação a banda no ~ 1.640 cm-1, uma convolução de Amida eu banda de Hyd1 e a curva de água H-O-H. Faixas adicionais devido a proteína podem ser vistas na região de estiramento C-H (ca 2900 cm-1). A banda larga, centralizada em torno de 2.100 cm-1 é uma banda de combinação da vibração dobra H-O-H com um conjunto de baixas bandas de libração de energia, que são restritas as rotações das moléculas de H2O devido a rede de ligação de hidrogênio em água líquida. A banda deCO νdo estado oxidado, inactivo, Ni-B de sítio ativo é claramente evidente no 1.943 cm-1, mesmo sem correção de linha de base, e νCN características são claramente visíveis entre 2.050-2.100 cm-1 . Em coberturas de Hyd1 altas, muito da estrutura microporosa do preto de carbono filme57 bloqueado pela enzima e, por conseguinte, a concentração de água 'em massa' é abaixada durante as medições PFIRE.
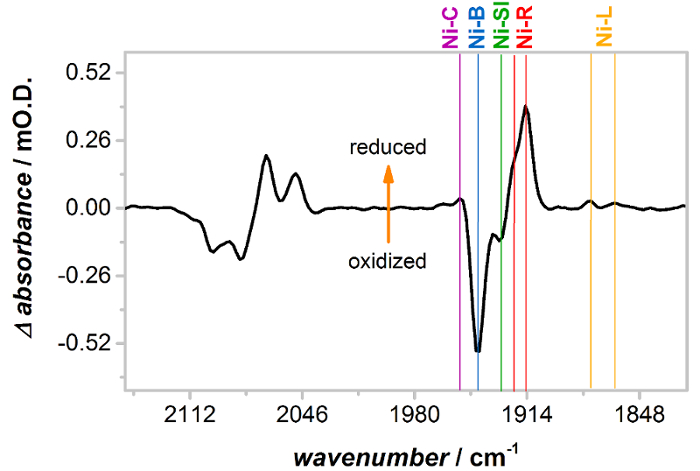
Os espectros na Figura 3 mostram que, antes da ativação, Hyd1 filmes contêm Hyd1 nos Estados oxidados, inativos. Ativação durante a noite no −0.8 V vs SCE sob uma atmosfera de2 H leva à formação de reduzida, Estados cataliticamente ativos conforme demonstrado na Figura 4 , que mostra um espectro ativado (reduzida) menos preparado como diferença (oxidado) de Hyd1. Espectros de diferença de Hyd1 podem ser mais claramente interpretados usando regiãoCO ν. Cada Estado único de sítio ativo tem somente uma faixa de CO, em comparação com duas bandas de NC e, portanto, a regiãoCN νé intrinsecamente mais complicada, com muitas bandas sobrepostas. O espectro de diferença na Figura 4 mostra que a ativação leva a perda (bandas de absorção negativa) de oxidado, inativo Ni-B e uma pequena quantidade de Ni-SI (o estado mais oxidado 'ativo') que estava presente no filme como preparado Hyd1. Estas são substituídas por 'ativos' Estados de Hyd1; Ni-C, R-Ni e Ni-L. nota que existem duas formas do Ni-R e Estados de Ni-L, como evidenciado pelas bandasCO dois νobservaram para essas espécies na Figura 4. A observação de vários Estados R-Ni e Ni-L está de acordo com outros NiFe hidrogenases. 48 , 58 , 59
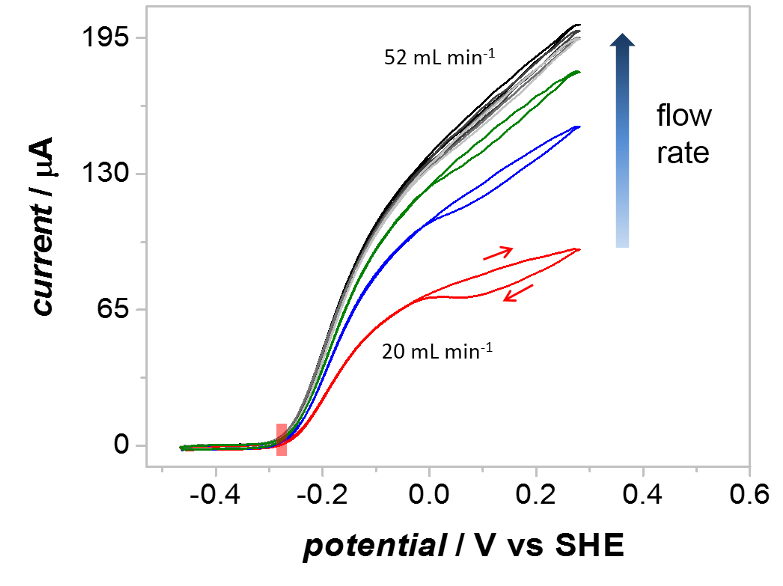
Uma verificação da chave do método PFIRE é que o voltammograms cíclico gravados de Hyd1 dentro da TV de célula de fluxo spectroelectrochemical waveshapes catalítica similar àquelas gravadas em um eletrodo de disco rotativo planar. 55 na prática isto significa que o transporte em massa de substrato (H2) e produto (H+) para o de Hyd1 imobilizado na célula spectroelectrochemical é eficiente para as taxas de fluxo usadas durante as medições PFIRE. O efeito da taxa de fluxo sobre o protector catalítico é mostrado na Figura 5, que mostra voltammograms sucessivas registadas sob uma atmosfera de2 H (1 bar), como a taxa de fluxo da solução através da célula spectroelectrochemical é aumentada. Em todos os casos a overpotential para oxidação de2 H por Hyd1 é idêntico (retângulo sombreado vermelho), mas a extensão da inactivação oxidativa (histerese entre a corrente durante o oxidativo e redutora varreduras em potenciais acima ca 0 V vs SCE) e a oxidação de2 H máxima atual dependem da taxa de fluxo da solução. Com vazões acima de 52 mL/min (luz cinzenta voltammogram) o protector catalítico é insensível a mais aumenta a taxa de fluxo.
Figura 6 compara as intensidades relativas da banda νCO do estado Ni-B após a ativação inicial e re-oxidação anaeróbica em 0 V vs SCE (sob uma atmosfera de Ar, conforme descrito em 4.3) e inativação oxidativa anaeróbia sob uma atmosfera de Ar em 0 V vs SCE após 48 horas de experimentos contínuos (como descrito no ponto 5.1). Sem perda de intensidade de banda local ativo é observada durante a medição, e todos a amostra Hyd1 responde para os potenciais aplicados.
A Figura 7 demonstra o procedimento de correção de base utilizado ao longo deste trabalho. O espectro absoluto da amostra Hyd1 na região sítio ativo (Figura 7uma) contém curvatura significativa devido à água. Na verdade, a exigência de usar a água como um solvente é um problema para a maioria das aplicações da espectroscopia de IR nas ciências da vida. A segunda derivada do espectro absoluta (Figura 7b), calculado usando o software de origem com Savitsky-Golay suavização sobre uma janela de 9 pontos, pode ser usada para identificar bandas afiadas do site Hyd1 ativo contra o fundo curvo. A identificação das posições de pico aproximado usando um espectro derivado segundo permite pontos de ancoragem da linha de base a ser colocados em regiões do espectro absoluta que estão livres de bandas sítio ativo (círculos na Figura 7um). Uma função spline cúbica é então montada através destes pontos de ancoragem para criar uma função de linha de base que pode então ser subtraída do espectro absoluto para dar um espectro de linha de base-corrigido contendo apenas picos provenientes do sítio activo Hyd1 (Figura 7 c).
A Figura 8 mostra os resultados de uma medição de PFIRE em Hyd1, sob condições de (atmosfera de2 H) volume de negócios em uma gama de potenciais e não rotatividade (atmosfera de Ar). 36 o hora atual traços (Figura 8um) relatório sobre o atual catalítico em cada aplicado potencial e permanecer perto de zero corrente sob condições não-volume de negócios (atmosfera de Ar). A titulação redox spectroelectrochemical parcial no Figura 8b , portanto, informa sobre o comportamento de redox de equilíbrio do sítio activo, mostrando a distribuição dos Estados esperados em cada potencial na ausência de volume de negócios catalítico. Os espectros na Figura 8c foram gravados sob condições de volume de negócios (em uma atmosfera de2 H) e, portanto, representam a distribuição de estado estacionário dos Estados de sítio ativo presentes durante a oxidação catalítica de2 H por Hyd1. Que condições de estado estacionário foram alcançadas é confirmada pelo facto de que o H2 oxidação catalítica atual (Figura 8um, H2) permanece constante em função do tempo no −0.199 V e −0.074 V vs que; o decaimento monotônico em corrente na +0.356 V vs ela é devido à inactivação oxidativa anaeróbica conhecida do Hyd1. 55 a distribuição dos Estados de sítio ativo é claramente diferente sob Ar e H2 em todos os potenciais onde Hyd1 executa a catálise (Figura 8b, espectros no −0.199, −0.074 e +0.356 V vs ). Os espectros no Ar e H2 são praticamente idênticos em −0.594 V vs que, no entanto, e isto representa um importante teste de consistência experimental; Hyd1 não reduz H+ a uma taxa significativa em pH 6,0 (a corrente na Figura 8um sob tanto Ar como H2 é próximo de zero), e os espectros na −0.594 V, portanto, são esperados para ser o mesmo.
A Figura 9 demonstra a inactivação oxidativa anaeróbia de Hyd1 através da formação de Ni-B de Ni-SI durante a oxidação de2 H em +0.356 V.36 , espectros foram gravados durante os intervalos de tempo cinza observados sobre o rastreamento de hora atual na Figura 9um. −0.074 V Hyd1 não sofre inativação oxidativa, e a distribuição dos Estados de sítio ativo permanece constante durante toda a etapa toda potencial. Isto é demonstrado pelos espectros na Figura 9beu, que se reporta o espectro de linha de base-corrigido absoluto no início da etapa potencial −0.074 e Figura 9bii , que foi gravado em uma posterior durante a etapa de potencial e é relatado como um espectro de diferença em relação à Figura 9beu. Os espectros na Figura 9biii e biv também foram descritos como espectros de diferença em relação à Figura 9beue mostrar a conversão gradual de Ni-SI Ni-B durante alto potencial inactivação, consistente com a diminuição monotonic corrente mostrada em +0.356 V na Figura 9um.
Espectros em uma faixa de pH da solução dar insights sobre a transferência de próton passos durante o Hyd1 catalítica do ciclo. 43 Figura 10 uma mostra espectros PFIRE sobre o mesmo filme Hyd1 a pH 3.0 e pH 9.0, usando o fluxo de solução na célula spectroelectrochemical para trocar o buffer experimental. As concentrações relativas dos Estados C-Ni e Ni-L são claramente diferentes destes dois espectros. Ao variar o potencial aplicado sob condições não-volume de negócios a dependência potencial da NiC e Ni-L Estados pode ser determinado rapidamente em um intervalo de valores de pH (Figura 10b), e ambos os Estados são encontrados para ser isopotential sobre uma ampla faixa de pH. (Note que as absorvâncias de pico na Figura 10b mostram apenas os Estados C-Ni e Ni-L para maior clareza, redox completo titulações de Hyd1 foram relatadas por Hidalgo et al) 36 uma titulação de pH a concentração de C-Ni e Ni-L pode então ser extraída, levando a absorvância de pico em potenciais onde a concentração total de C-Ni e Ni-L está no seu máximo em cadaFigura 10(c) de pH. Desta forma e em conjunto com os dados do EPR, um equilíbrio de pH entre o Ni-C e Estados Ni-L foi identificado. 43

Figura 1: Esquemático do arranjo de espectrômetro de IR glovebox anaeróbica, acessório ATR, detector de MCT, gás sistema de fluxo e potentiostat usado para medições de PFIRE. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Diagrama esquemático da célula de spectroelectrochemical usado para medições de PFIRE, mostrando o arranjo dos eletrodos e a solução de conexões de entrada/saída. A célula e a placa de base são usinadas de poliéter éter cetona (PEEK), com furos de parafuso para uma vareta de carbono trabalhando conexão eletrodo, elétrodo contrário de arame, de Pt, eletrodo de referência de calomelano saturado e solução de entrada e saída. A construção de eletrodos de referência de calomelano saturado é como anteriormente relatada. 36 Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: espectro de absorbância de preto de carbono modificado Hyd1 partículas, depositado sobre a ira e reidratado com buffer. As posições do Amida que banda, banda de Amida II e da região de sítio ativo Hyd1 são mostrados, juntamente com características adicionais devido as vibrações de estiramento C-H e solvente água. A inserção mostra uma visão ampliada da região do sítio activo, com as vCO vCN bandas e rotuladas. 'Como preparado' partículas contêm Hyd1 principalmente no estado oxidado, inactivo Ni-B. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4 : Ativação de Hyd1 em −0.8 V vs SCE sob uma atmosfera de2 H, apresentada-se como um espectro reduzido menos diferença oxidado. Após a ativação potencial baixa oxidada, Ni-B inativo (e uma pequena concentração de Ni-SI) converte para mais reduzidos, ativos Estados Ni-C, R-Ni e Ni-L. nota que Hyd1 tem dois distintos Estados sub de Ni-L e Ni-R. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5 : O efeito da taxa de fluxo da solução sobre o protector de catalítico cíclica voltammograms gravado na célula spectroelectrochemical. Voltammograms foram gravadas no aumento das taxas de fluxo de H2-saturado reserva conforme indicado. Com um caudal de 20 mL/min (vermelho) o voltammogram mostra significativa inativação acima de 0 V vs no exame para a frente. Com vazões acima de 52 mL/min, a extensão da inactivação é vastamente mais baixa e a corrente é independente da taxa de fluxo em todas as potencialidades. Outros parâmetros: 1 barra H2, taxa de varredura de 10 mV/s. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6 : Linha de base corrigida espectros de IR na região de CO νsítio ativo, inativo, oxidado Ni-B adjunto do 0 V vs SCE. Não há nenhuma perda mensurável de intensidade do sítio activo durante 48 h de medições contínuas de PFIRE, e, portanto, Hyd1 é adsorvido robustamente das partículas de negro de carbono. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 7: Detalhes dos procedimentos de correção de linha de base utilizados para manipulação de dados. Pontos de ancoragem da linha de base são colocados sobre o espectro de absorbância absoluto da região de sítio ativo (um), tendo o cuidado de evitar qualquer νCO νCN picos e identificados de uma segunda análise de derivativos (b). O espectro de corrigida de base resultante é mostrado em (c). Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 8: PFIRE das medições de Hyd1 não-volume de negócios (Ar) e volume de negócios (H2) condições. (uma) hora atual vestígios de Hyd1 na célula spectroelectrochemical em Ar saturado (cinza) e H2-tampão (preto) saturada; (b), (c) PFIRE espectros mostrando a região doCO νcada potencial sob Ar (b) e H2 (c). Potenciais citado em V contra ela. Reproduzido com a permissão de Hidalgo et al . 35 Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 9: Anaeróbia inactivação de Hyd1 através de formação de Ni-B de Ni-SI. (um) rastreamento de hora atual sob uma atmosfera de2 H, mostrando uma electrocatalytic estável atual no −0.074 V e inativação anaeróbica lenta (monotónica diminuição da corrente) na +0.356 V vs . (b) espectros gravado durante as regiões sombreadas cinza marcadas no (a). Espectro b é um espectro de linha de base corrigida gravado no início da etapa de potenciais −0.074 V. Espectro bii, gravado em um momento posterior, durante a etapa de −0.074 V, é relatado como um espectro de diferença em relação ao beu e mostra que não há alterações na distribuição dos Estados de sítio ativo ocorre, consistente com a estabilidade do potencial no −0.074 V. Espectros biii e biv também são relatados como espectros de diferença em relação a b,eu e mostrar a conversão gradual de Ni-SI Ni-B durante inactivação anaeróbia em +0.356 V vs . Reproduzido com a permissão de Hidalgo et al . 35 Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 10 : Espectros em uma faixa de pH da solução dar insights sobre a transferência de próton etapas durante o ciclo catalítico Hyd1. (um) IR espectros mostrando νCO região de Hyd1, gravado no pH 3.0 (-54 mV vs ) e pH 9.0 (-334 mV vs ). (b) Spectroelectrochemical titulações foram realizadas para determinar o potencial em que as concentrações de C-Ni e Ni-L são no máximo um intervalo de valores de pH da solução. Apenas as concentrações C-Ni e Ni-L são mostradas para maior clareza, para uma titulação spectroelectrochemical completo da Sé Hyd1 Hidalgo et al . dependência de pH 36 (c) da concentração relativa de C-Ni e Ni-L, como determinado a partir de uma série de experiências tais como as mostradas em (b). Espectros foram gravados a 20 ° C. Adaptado com permissão de Murphy et al . 42 Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
PFIRE é uma técnica de espectroscopia de IR amplamente aplicável para endereçamento de proteínas redox eletrodo-imobilizado. Em particular, electrocatalytic reações de enzimas redox podem ser analisadas sob condições de rápida rotatividade. O método PFIRE baseia-se o controle de eletroquímico direto fornecido pela técnica de PFE, que não dá nenhuma informação estrutural direta e casais a espectroscopia de IR em um eletrodo de carbono. A abordagem PFIRE assim adiciona uma visão química com as informações disponíveis da eletroquímica sozinha e é altamente adequada para o estudo de enzimas e proteínas redox envolvidas na ligação de pequenas moléculas e ativação. Além disso, PFIRE podem fornecer informações sobre potencial dependente de mudanças estruturais em proteínas na ausência de volume de negócios catalítico. Tais eventos de transferência de elétrons não-volume de negócios são frequentemente difíceis de detectar usando aplicativos 'padrão' de PFE, embora a extensão do PFE a transformada de Fourier AC voltametria tem sido usada com grande sucesso. 45 , 46
O método PFIRE é, em princípio, adequado para o estudo de qualquer proteína redox que pode ser estudada usando PFE. Portanto, como com PFE, adsorção de proteínas é um passo crítico para uma experiência bem sucedida de PFIRE. Neste protocolo, descrevemos uma aplicação da técnica PFIRE usando Hyd1 de Escherichia coli como um estudo de caso. 36 , 43 no entanto, também aplicámos a técnica PFIRE para a hidrogenase citoplasmática reguladora de r. eutropha,44 e o mononucleótido de flavina adsorvido em carbono preto. 40 em todos estes casos, a adsorção física simples ao carvão sem modificações alta área de superfície (conforme descrito neste protocolo) fornece uma cobertura de superfície da proteína que é alta o suficiente para espectros de IR gravar de boa qualidade com uma alta relação sinal-ruído. Em casos onde não é possível obter níveis tão altos de adsorção pode ser necessário modificar a superfície das partículas de carbono, por exemplo, para permitir a fixação covalente de proteína para a superfície do eletrodo. 60 , 61 , 62 a utilização de uma caixa de luvas para medições de PFIRE só é estritamente necessária quando estudar amostras que devem ser tratadas por via anaeróbia. No entanto, na prática o extremamente constante e baixo (ponto de orvalho < 80 ° C) níveis de vapor de água fornecida pela atmosfera glovebox dar altos níveis de sinal-ruído que permitem a extração das absorvâncias muito pequenas. 44 em muitos casos um ambiente anaeróbio, como o fornecido pelo porta-luvas, é também desejável para a medição de eletroquímica (integral para a técnica PFIRE) para evitar atual devido a redução de2 O para o eletrodo de trabalho.
Absorvâncias de IR devido ao volume de água, buffers de experimentais e as partículas de carbono no qual a amostra é adsorvida contribuem significativamente para os espectros experimentais e poderia sobrepor-se bandas de interesse, nomeadamente a amida, regiões II e III do espectro. 63 a região de Amida também contém informações de espécies orgânicas como flavinas ou nicotinamida cofactores, bem como os substratos e produtos de muitas reações de oxidação e redução. No caso de NiFe hidrogenases, bandasCN CO e ν νdo sítio ativo cair em uma região relativamente clara do espectro e então a técnica PFIRE é muito bem adaptada para o estudo destas enzimas. Em outros casos, no entanto, espectros de diferença juntamente com abordagens isotópicas de rotulagem podem ser necessário para isolar as mudanças devido a proteína imobilizada. Abordagens similares têm sido utilizadas para identificar, por exemplo, alterações de protonação, rearranjos estruturais e estudar complexos de Michaelis-Menten usando espectroscopia de IR. 64 , 65 , 66 PFIRE não é, portanto, limitado ao estudo de hidrogenases, mas pode ser aplicado a qualquer proteína redox que contém (ou cujos substratos, produtos ou inibidores contêm) grupos com diagnósticos vibrações IR-ativo; dehydrogeanses de monóxido de carbono,67 nitrogenases,68,14 flavoproteínas,40 e formiato de desidrogenases, por exemplo.
A técnica relacionada, SEIRA, está muito bem adaptada para o estudo de proteínas de membrana-associado em um ambiente biomimético. 32 SEIRA é uma adaptação da espectroscopia de IR que também usa uma configuração de ATR-IR e faz uso de um efeito de superfície do realce que amplifica a absorvância de IR de moléculas situadas perto de (alguns nm) a superfície do prisma ATR (IRE). SEIRA é, portanto, requintadamente sensível a alterações espectrais que ocorrem dentro da arquitetura de proteínas e membrana adsorvida e é relativamente livre de concorrentes sinais de solvente e substratos/inibidores presentes na solução. Isso é um pouco em contraste com a técnica PFIRE descrita aqui que se baseia em uma profundidade de penetração significativamente maior acima da superfície da ira (~ 1 µm), significando que PFIRE é mais sensível a substratos, produtos ou inibidores presentes na solução. Este aumento da sensibilidade à «a granel» solvente pode ser vantajoso; Se o substrato ou o produto pode ser observado diretamente por IR, espectros PFIRE relatam sobre ambos cinética do estado estacionário da longeva espécie ativa e formação de produto associado durante a electrocatalysis. 69 a capacidade de observar as concentrações de estado estacionário de substrato e o produto será particularmente valiosa para enzimas como a desidrogenase de monóxido de carbono (que catalisa a oxidação reversível de CO de CO2, um forte absorvente de IR) ou desidrogenase de formiato (que catalisa a oxidação reversível de formiato de CO2).
Atualmente, PFIRE é limitado a estudos cinéticos de estado estacionário de enzima electrocatalysis devido os eletrodos de carbono macroscópica costumávamos 'concentrado' a enzima para a superfície de uma ira para coleta de dados em e geometria ATR-IR. A este respeito estudos PFIRE de NiFe hidrogenases são complementares ao trabalho de Dyer e cooperadores,49,50 , quem usar a espectroscopia de absorção de transiente acionada por luz de IR estudar a cinética de sub volume de negócios. Trabalho está em andamento miniaturizar a célula spectroelectrochemical,40 , e com o uso de microeletrodos resolução de tempo da ordem de microssegundos deve ser alcançável. Isto permitirá o estudo da cinética sub volume de negócios para as enzimas com frequências de volume de negócios até ca 100-500 s-1e permitirá o estudo de processos redutoras e oxidativos.
Em geral, PFIRE é uma técnica espectroscópica que permite a caracterização química de electrocatalytic reações de enzimas redox sob condições de estado estacionário. A abordagem PFIRE permite múltiplas titulações químicas e eletroquímicas a efectuar sobre a mesma amostra de enzima, como os eletrodos de alta área de superfície usados fornecem uma robusta adsorção de proteínas e a geometria de ATR-IR permite troca de fácil solução. A capacidade de coletar tais informações estruturais em situ durante a função de enzima é uma ferramenta inestimável para a Comunidade bioelectrochemistry.
Divulgações
Os autores não declaram nenhum interesse financeiro concorrente.
Agradecimentos
O trabalho de K.A.V. e P.A.A. foi apoiado pelo Conselho Europeu de investigação (EnergyBioCatalysis-ERC-2010-StG-258600), engenharia e ciências físicas pesquisa Conselho IB Catalyst award EP/N013514/1 e pesquisas de ciências biológicas e biotecnologia Conselho (BB/L009722/1 e BB/N006321/1). R.H. foi apoiada pelo Ministerio de Ciencia y Tecnologìa, Universidad de Costa Rica e Lincoln College, Oxford. Os autores reconhecem Mr. Charlie Jones, Sr. Charlie Evans e pessoal da oficina mecânica (departamento de química) para assistência na concepção e fabrico de spectroelectrochemical células utilizadas neste trabalho.
Materiais
| Name | Company | Catalog Number | Comments |
| Spectrometer | Agilent | 680-IR | with an external MCT detector |
| ATR accessory | Pike Technologies | GladiATR | Customised for use with a 5-reflection Si IRE |
| Glovebox | Glove Box Technology Ltd. | N/A | Custom designed 'wet' and 'dry' box for anaerobic sample handling and measurement |
| KBr window | Crystran | Custom | To allow coupling of the glovebox with the external beam of the FTIR spectrometer |
| Additional optics | Agilent | N/A | Components from a PM-IRRAS accessory |
| Silicon IRE | Crystal GmbH | Custom | Trapezoidal: 8.4 mm x 5 mm (large face), 1 mm thickness, ca 39 degree face angle |
| Potentiostat | Metrohm | Autolab PGSTAT 128N | |
| Nova 10.1 | Metrohm | Software for controlling the potentiostat | |
| Peristaltic pump | Williamson Manufacturing Company Ltd | QL-1000-024-300 | |
| Pt wire | Surepure Chemetals | 3272 | 99.95% Pure Platinum Wire, 0.014 inch Diameter |
| Carbon rod | WH Smith | 30729209 | 0.7 mm HB pencil lead |
| Carbon black | Cabot Corporation | Black Pearls 2000 | |
| Ultrasonic bath | Ultrawave | U100 | 35 W |
| Centrifugal filter | Merck Millipore | UFC5050BK | Amicon Ultra, 50 KDa MW cutoff |
| Microcentrifuge | Eppendorf | 5452000018 | MiniSpin |
| NaCl | Sigma | 71376 | |
| H2SO4 | Fisher | S/9240/PB17 | |
| HNO3 | Fisher | N/2300/PB17 | |
| silicone sealant | Cow Corning Toray Co., Ltd. | SE 4486 | |
| Carbon paper | Toray | TGP-H-030 | |
| H2 gas | BOC | ||
| N2 gas | BOC | ||
| OriginPro 2015 | OriginLab | Data analysis/graphing software | |
| Resolutions Pro 4.0 | Varian | Software for controlling FTIR spectrometer |
Referências
- Ash, P. A., Vincent, K. A. Spectroscopic analysis of immobilised redox enzymes under direct electrochemical control. Chem. Commun. 48 (10), 1400-1409 (2012).
- Sezer, M., Millo, D., Weidinger, I. M., Zebger, I., Hildebrandt, P. Analyzing the catalytic processes of immobilized redox enzymes by vibrational spectroscopies. IUBMB Life. 64 (6), 455-464 (2012).
- Melin, F., Hellwig, P. Recent advances in the electrochemistry and spectroelectrochemistry of membrane proteins. Biol. Chem. 394 (5), 593-609 (2013).
- Dong, S., Niu, J., Cotton, T. M. Ultraviolet/visible spectroelectrochemistry of redox proteins. Methods Enzymol. 246, 701-732 (1995).
- Hellwig, P., et al. Electrochemical and Ultraviolet/Visible/Infrared Spectroscopic Analysis of Heme a and a3 Redox Reactions in the Cytochrome c Oxidase from Paracoccus denitrificans Separation of Heme a and a3 Contributions and Assignment of Vibrational Modes. Biochemistry. 38 (6), 1685-1694 (1999).
- Wu, S., et al. Redox chemistry of the Schizosaccharomyces pombe ferredoxin electron-transfer domain and influence of Cys to Ser substitutions. J. Inorg. Biochem. 105 (6), 806-811 (2011).
- Whitehouse, C. J. C., et al. A Highly Active Single-Mutation Variant of P450BM3 (CYP102A1). ChemBioChem. 10 (10), 1654-1656 (2009).
- Marritt, S. J., van Wonderen, J. H., Cheesman, M. R., Butt, J. N. Magnetic circular dichroism of hemoproteins with in situ control of electrochemical potential: "MOTTLE". Anal. Biochem. 359 (1), 79-83 (2006).
- Moss, D., Nabedryk, E., Breton, J., Mäntele, W. Redox-linked conformational changes in proteins detected by a combination of infrared spectroscopy and protein electrochemistry. Eur. J. Biochem. 187 (3), 565-572 (1990).
- Iwaki, M., et al. Direct Observation of Redox-Linked Histidine Protonation Changes in the Iron-Sulfur Protein of the Cytochrome bc1 Complex by ATR-FTIR Spectroscopy. Biochemistry. 44 (11), 4230-4237 (2005).
- Marshall, D., et al. ATR-FTIR Redox Difference Spectroscopy of Yarrowia lipolytica and Bovine Complex I. Biochemistry. 45 (17), 5458-5467 (2006).
- Lacey, A. L. D., Gutiérrez-Sánchez, C., Fernández, V. M., Pacheco, I., Pereira, I. A. C. FTIR spectroelectrochemical characterization of the Ni-Fe-Se hydrogenase from Desulfovibrio vulgaris Hildenborough. J. Biol. Inorg. Chem. 13 (8), 1315-1320 (2008).
- Millo, D., et al. Spectroelectrochemical Study of the [NiFe] Hydrogenase from Desulfovibrio vulgaris Miyazaki F in Solution and Immobilized on Biocompatible Gold Surfaces. J. Phys. Chem. B. 113 (46), 15344-15351 (2009).
- Paengnakorn, P., et al. Infrared spectroscopy of the nitrogenase MoFe protein under electrochemical control: potential-triggered CO binding. Chem. Sci. 8 (2), 1500-1505 (2017).
- Male, L., et al. Protein voltammetry and spectroscopy: integrating approaches. Theor. Chem. Acc. 119 (1-3), 107-111 (2008).
- Healy, A. J., Ash, P. A., Lenz, O., Vincent, K. A. Attenuated total reflectance infrared spectroelectrochemistry at a carbon particle electrode; unmediated redox control of a [NiFe]-hydrogenase solution. Phys. Chem. Chem. Phys. 15 (19), 7055-7059 (2013).
- Léger, C., Bertrand, P. Direct Electrochemistry of Redox Enzymes as a Tool for Mechanistic Studies. Chem. Rev. 108 (7), 2379-2438 (2008).
- Armstrong, F. A., Hirst, J. Reversibility and efficiency in electrocatalytic energy conversion and lessons from enzymes. Proc. Natl. Acad. Sci. U.S.A. 108 (34), 14049-14054 (2011).
- Gates, A. J., et al. The relationship between redox enzyme activity and electrochemical potential-cellular and mechanistic implications from protein film electrochemistry. Phys. Chem. Chem. Phys. 13 (17), 7720-7731 (2011).
- Pershad, H. R., et al. Catalytic Electron Transport in Chromatium vinosum [NiFe]-Hydrogenase: Application of Voltammetry in Detecting Redox-Active Centers and Establishing That Hydrogen Oxidation Is Very Fast Even at Potentials Close to the Reversible H+/H2 Value. Biochemistry. 38 (28), 8992-8999 (1999).
- Goldet, G., et al. Electrochemical Kinetic Investigations of the Reactions of [FeFe]-Hydrogenases with Carbon Monoxide and Oxygen: Comparing the Importance of Gas Tunnels and Active-Site Electronic/Redox Effects. J. Am. Chem. Soc. 131 (41), 14979-14989 (2009).
- Vincent, K. A., Parkin, A., Armstrong, F. A. Investigating and Exploiting the Electrocatalytic Properties of Hydrogenases. Chem. Rev. 107 (10), 4366-4413 (2007).
- Parkin, A., Seravalli, J., Vincent, K. A., Ragsdale, S. W., Armstrong, F. A. Rapid and Efficient Electrocatalytic CO2/CO Interconversions by Carboxydothermus hydrogenoformans CO Dehydrogenase I on an Electrode. J. Am. Chem. Soc. 129 (34), 10328-10329 (2007).
- van Wonderen, J. H., Burlat, B., Richardson, D. J., Cheesman, M. R., Butt, J. N. The Nitric Oxide Reductase Activity of Cytochrome c Nitrite Reductase from Escherichia coli. J. Biol. Chem. 283 (15), 9587-9594 (2008).
- Schaming, D., et al. Spectroelectrochemical Characterization of Small Hemoproteins Adsorbed within Nanostructured Mesoporous ITO Electrodes. Langmuir. 28 (39), 14065-14072 (2012).
- Kemp, G. L., et al. Opportunities for mesoporous nanocrystalline SnO2 electrodes in kinetic and catalytic analyses of redox proteins. Biochem. Soc. Trans. 37 (2), 368-372 (2009).
- Renault, C., et al. Unraveling the Mechanism of Catalytic Reduction of O2 by Microperoxidase-11 Adsorbed within a Transparent 3D-Nanoporous ITO Film. J. Am. Chem. Soc. 134 (15), 6834-6845 (2012).
- Krzemiński, Ł, et al. Spectroelectrochemical Investigation of Intramolecular and Interfacial Electron-Transfer Rates Reveals Differences Between Nitrite Reductase at Rest and During Turnover. J. Am. Chem. Soc. 133 (38), 15085-15093 (2011).
- Akkilic, N., Kamran, M., Stan, R., Sanghamitra, N. J. M. Voltage-controlled fluorescence switching of a single redox protein. Biosens. Bioelectron. 67, 747-751 (2015).
- Ataka, K., Heberle, J. Electrochemically Induced Surface-Enhanced Infrared Difference Absorption (SEIDA) Spectroscopy of a Protein Monolayer. J. Am. Chem. Soc. 125 (17), 4986-4987 (2003).
- Millo, D., Hildebrandt, P., Pandelia, M. -E., Lubitz, W., Zebger, I. SEIRA Spectroscopy of the Electrochemical Activation of an Immobilized [NiFe] Hydrogenase under Turnover and Non-Turnover Conditions. Angew. Chem. Int. Ed. 50 (11), 2632-2634 (2011).
- Ataka, K., Stripp, S. T., Heberle, J. Surface-enhanced infrared absorption spectroscopy (SEIRAS) to probe monolayers of membrane proteins. Biochim. Biophys. Acta, Biomembr. 1828 (10), 2283-2293 (2013).
- Kozuch, J., et al. Voltage-dependent structural changes of the membrane-bound anion channel hVDAC1 probed by SEIRA and electrochemical impedance spectroscopy. Phys. Chem. Chem. Phys. 16 (20), 9546-9555 (2014).
- Silveira, C. M., et al. SERR Spectroelectrochemical Study of Cytochrome cd1 Nitrite Reductase Co-Immobilized with Physiological Redox Partner Cytochrome c552 on Biocompatible Metal Electrodes. PLoS One. 10 (6), 0129940(2015).
- Todorovic, S., Hildebrandt, P., Martins, L. Surface enhanced resonance Raman detection of a catalytic intermediate of DyP-type peroxidase. Phys. Chem. Chem. Phys. 17 (18), 11954-11957 (2015).
- Hidalgo, R., Ash, P. A., Healy, A. J., Vincent, K. A. Infrared Spectroscopy During Electrocatalytic Turnover Reveals the Ni-L Active Site State During H2 Oxidation by a NiFe Hydrogenase. Angew. Chem. Int. Ed. 54 (24), 7110-7113 (2015).
- Grabarczyk, D. B., Ash, P. A., Vincent, K. A. Infrared Spectroscopy Provides Insight into the Role of Dioxygen in the Nitrosylation Pathway of a [2Fe2S] Cluster Iron-Sulfur Protein. J. Am. Chem. Soc. 136 (32), 11236-11239 (2014).
- Kriegel, S., Uchida, T., Osawa, M., Friedrich, T., Hellwig, P. Biomimetic Environment to Study E. coli Complex I through Surface-Enhanced IR Absorption Spectroscopy. Biochemistry. 53 (40), 6340-6347 (2014).
- El Khoury, Y., Van Wilderen, L. J. G. W., Bredenbeck, J. Ultrafast 2D-IR spectroelectrochemistry of flavin mononucleotide. J. Chem. Phys. 142 (21), 212416(2015).
- Ash, P. A., et al. Synchrotron-Based Infrared Microanalysis of Biological Redox Processes under Electrochemical Control. Anal. Chem. 88 (13), 6666-6671 (2016).
- Nakamoto, K. Infrared and Raman Spectra of Inorganic and Coordination Compounds. Part B. , John Wiley & Sons, inc. (2009).
- Bard, A., Faulkner, L. R. Electrochemical Methods: Fundamentals and Applications. , John Wiley & Sons, Inc. (2001).
- Murphy, B. J., et al. Discovery of Dark pH-Dependent H+ Migration in a [NiFe]-Hydrogenase and Its Mechanistic Relevance: Mobilizing the Hydrido Ligand of the Ni-C Intermediate. J. Am. Chem. Soc. 137 (26), 8484-8489 (2015).
- Ash, P. A., et al. Electrochemical and Infrared Spectroscopic Studies Provide Insight into Reactions of the NiFe Regulatory Hydrogenase from Ralstonia eutropha with O2 and CO. J. Phys. Chem. B. 119 (43), 13807-13815 (2015).
- Lee, C. -Y., et al. Theoretical and experimental investigation of surface-confined two-center metalloproteins by large-amplitude Fourier transformed ac voltammetry. J. Electroanal. Chem. 656 (1-2), 293-303 (2011).
- Adamson, H., et al. Electrochemical evidence that pyranopterin redox chemistry controls the catalysis of YedY, a mononuclear Mo enzyme. Proc. Natl. Acad. Sci. U.S.A. 112 (47), 14506-14511 (2015).
- De Lacey, A. L., Fernández, V. M., Rousset, M., Cammack, R. Activation and Inactivation of Hydrogenase Function and the Catalytic Cycle: Spectroelectrochemical Studies. Chem. Rev. 107 (10), 4304-4330 (2007).
- Lubitz, W., Ogata, H., Rüdiger, O., Reijerse, E. Hydrogenases. Chem Rev. 114 (8), 4081-4148 (2014).
- Greene, B. L., Wu, C. -H., McTernan, P. M., Adams, M. W. W., Dyer, R. B. Proton-Coupled Electron Transfer Dynamics in the Catalytic Mechanism of a [NiFe]-Hydrogenase. J. Am. Chem. Soc. 137 (13), 4558-4566 (2015).
- Greene, B. L., Wu, C. -H., Vansuch, G. E., Adams, M. W. W., Dyer, R. B. Proton Inventory and Dynamics in the Nia-S to Nia-C Transition of a [NiFe] Hydrogenase. Biochemistry. 55 (12), 1813-1825 (2016).
- Greene, B. L., Vansuch, G. E., Wu, C. -H., Adams, M. W. W., Dyer, R. B. Glutamate Gated Proton-Coupled Electron Transfer Activity of a [NiFe]-Hydrogenase. J. Am. Chem. Soc. 138 (39), 13013-13021 (2016).
- Sezer, M., et al. Role of the HoxZ Subunit in the Electron Transfer Pathway of the Membrane-Bound [NiFe]-Hydrogenase from Ralstonia eutropha Immobilized on Electrodes. J. Phys. Chem. B. 115 (34), 10368-10374 (2011).
- Heering, H. A., Wiertz, F. G. M., Dekker, C., de Vries, S. Direct Immobilization of Native Yeast Iso-1 Cytochrome c on Bare Gold: Fast Electron Relay to Redox Enzymes and Zeptomole Protein-Film Voltammetry. J. Am. Chem. Soc. 126 (35), 11103-11112 (2004).
- dos Santos, L., Climent, V., Blanford, C. F., Armstrong, F. A. Mechanistic studies of the "blue" Cu enzyme, bilirubin oxidase, as a highly efficient electrocatalyst for the oxygen reduction reaction. Phys. Chem. Chem. Phys. 12 (42), 13962-13974 (2010).
- Lukey, M. J., et al. How Escherichia coli Is Equipped to Oxidize Hydrogen under Different Redox Conditions. J. Biol. Chem. 285 (6), 3928-3938 (2010).
- Jones, A. K., et al. Enzyme Electrokinetics: Electrochemical Studies of the Anaerobic Interconversions between Active and Inactive States of Allochromatium vinosum [NiFe]-hydrogenase. J. Am. Chem. Soc. 125 (28), 8505-8514 (2003).
- Quinson, J., et al. Comparison of carbon materials as electrodes for enzyme electrocatalysis: hydrogenase as a case study. Faraday Trans. 172 (0), 473-496 (2014).
- Shafaat, H. S., Rüdiger, O., Ogata, H., Lubitz, W. [NiFe] hydrogenases: A common active site for hydrogen metabolism under diverse conditions. Biochim. Biophys. Acta, Bioenerg. 1827 (8-9), 986-1002 (2013).
- Ash, P. A., Hidalgo, R., Vincent, K. A. Proton transfer in the catalytic cycle of NiFe hydrogenases: insight from vibrational spectroscopy. ACS Catalysis. , (2017).
- Krishnan, S., Armstrong, F. A. Order-of-magnitude enhancement of an enzymatic hydrogen-air fuel cell based on pyrenyl carbon nanostructures. Chem. Sci. 3 (4), 1015-1023 (2012).
- Baffert, C., et al. Covalent Attachment of FeFe Hydrogenases to Carbon Electrodes for Direct Electron Transfer. Anal. Chem. 84 (18), 7999-8005 (2012).
- Rüdiger, O., et al. Enzymatic Anodes for Hydrogen Fuel Cells based on Covalent Attachment of Ni-Fe Hydrogenases and Direct Electron Transfer to SAM-Modified Gold Electrodes. Electroanal. 22 (7-8), 776-783 (2010).
- Barth, A., Zscherp, C. What vibrations tell about proteins. Q. Rev. Biophys. 35 (4), 369-430 (2002).
- Jiang, X., et al. Resolving voltage-dependent structural changes of a membrane photoreceptor by surface-enhanced IR difference spectroscopy. Proc. Natl. Acad. Sci. U.S.A. 105 (34), 12113-12117 (2008).
- Kottke, T., Lòrenz-Fonfrìa, V. A., Heberle, J. The Grateful Infrared: Sequential Protein Structural Changes Resolved by Infrared Difference Spectroscopy. J. Phys. Chem. B. 121 (2), 335-350 (2017).
- Callender, R., Dyer, R. B. The Dynamical Nature of Enzymatic Catalysis. Acc. Chem. Res. 48 (2), 407-413 (2015).
- Ciaccafava, A., et al. When the inhibitor tells more than the substrate: the cyanide-bound state of a carbon monoxide dehydrogenase. Chem. Sci. 7 (5), 3162-3171 (2016).
- George, S. J., Ashby, G. A., Wharton, C. W., Thorneley, R. N. F. Time-Resolved Binding of Carbon Monoxide to Nitrogenase Monitored by Stopped-Flow Infrared Spectroscopy. J. Am. Chem. Soc. 119 (27), 6450-6451 (1997).
- McPherson, I. J., Ash, P. A., Jacobs, R. M. J., Vincent, K. A. Formate adsorption on Pt nanoparticles during formic acid electro-oxidation: insights from in situ infrared spectroscopy. Chem Commun. 52 (85), 12665-12668 (2016).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados