
Method Article
Подготовка образцов методом 3D-коррелятивного фрезерования сфокусированного ионного пучка для криоэлектронной томографии высокого разрешения
В этой статье
Резюме
В данной работе мы представляем конвейер для 3D-коррелятивного фрезерования сфокусированного ионного пучка для подготовки клеточных образцов для криоэлектронной томографии. 3D-положение интересующих флуоресцентно меченых белков сначала определяется с помощью криофлуоресцентной микроскопии, а затем подвергается измельчению. Протокол подходит для клеток млекопитающих, дрожжей и бактерий.
Аннотация
Криоэлектронная томография (криоэлектронная томография) стала методом выбора для исследования клеточной ультраструктуры и молекулярных комплексов в их нативном, замороженно-гидратированном состоянии. Однако крио-ЭТ требует, чтобы образцы были достаточно тонкими, чтобы не рассеивать и не блокировать падающий электронный пучок. Для толстых ячеистых образцов это может быть достигнуто с помощью криофокусированного ионного пучка (FIB). Этот протокол описывает, как нацеливаться на определенные клеточные участки во время измельчения FIB с использованием 3D-коррелятивного подхода, который объединяет данные трехмерной флуоресцентной микроскопии с информацией, полученной с помощью электронного микроскопа FIB. Используя этот метод, редкие клеточные события и структуры могут быть обнаружены с высокой точностью и визуализированы с молекулярным разрешением с помощью криотрансмиссионной электронной микроскопии (крио-ПЭМ).
Введение
Измельчение сфокусированным ионным пучком позволяет получать тонкие биологические образцы из криофиксированных образцов без проблем, обычно связанных с механическими сечениями, таких как следы ножа и артефакты сжатия1. В сочетании с криоэлектронной томографией измельчение FIB позволяет проводить биологические исследования морфологии клеток с высоким разрешением и определять структуру высокомолекулярных комплексов непосредственно внутри клеток с субнанометровым разрешением 2,3,4. В то время как широко распространенные виды, такие как рибосомы, легко обнаруживаются в случайно вырезанных ламелях FIB, многие клеточные процессы полагаются на колокализацию нескольких комплексов или локализуются в определенных участках внутри клетки. Следовательно, требуется эффективное наведение, чтобы не потерять интересующую биологическую особенность в процессе измельчения и ограничиться случайными попаданиями. Поэтому необходим коррелятивный подход, объединяющий данные сканирующего электронного микроскопа (SEM)-FIB и криофлуоресцентного светового микроскопа (FLM). Несмотря на то, что начальную корреляцию можно опустить и объединить данные FLM и cryo-ET только после регистрации ПЭМ5,6, фрезерование сфокусированного ионного пучка под контролем флуоресценции позволяет заранее точно выбрать область измельчения, что приводит к более эффективному сбору данных. С момента его зарождения7 применение 3D-коррелированного измельчения FIB в биологических исследованиях было ограничено до тех пор, пока мы недавно не сообщили об обнаружении нового компартмента с разделением фаз жидкость и жидкостью (LLPS) у дрожжей с помощью этого метода8.
Здесь описан обобщенный 3D-протокол криокоррелированной световой и электронной микроскопии (CLEM), который может быть использован для изучения широкого спектра образцов, начиная от бактерий и заканчивая дрожжами и клетками млекопитающих. Несмотря на то, что эксперименты проводились с использованием определенного набора приборов, отдельные этапы не привязаны к конкретному оборудованию и могут быть легко перенесены в другие системы в качестве расширения существующих протоколов 3,5. Перечень тестируемого оборудования и предлагаемые настройки приведены в Таблице материалов и Таблице 1. Четыре ключевых этапа трубопровода: (1) подготовка образца, (2) локализация интересующих объектов с помощью криофлуоресцентной микроскопии, (3) 3D-коррелированное измельчение сфокусированного ионного пучка и (4) локализация целевых структур для сбора данных крио-ЭТ на ламелях в криопросвечивающем электронном микроскопе (рис. 1).
протокол

Рисунок 1: Краткое описание рабочего процесса с выборкой критических шагов. Весь протокол разделен на четыре этапа в зависимости от используемого оборудования: пробоподготовка, включая погружную заморозку, криофлуоресцентную микроскопию, криофокусированное измельчение ионным пучком и криоэлектронную микроскопию. Для каждого шага выделяется несколько ключевых моментов. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
1. Культивирование клеток и погружная заморозка сеток
- Культивирование выбранных клеток и оптимизация стратегий маркировки и обработки при комнатной температуре перед переходом к криоэкспериментам. Интересующие нас мишени либо маркируются с помощью флуоресцентных белков, либо с помощью живого окрашивания (например, Halo-Tag, LysoTracker, BODIPY, окрашивание живыми антителами и т. д.). Если обработка химическими или биологическими агентами (малыми молекулами, специальными средами, миРНК и т.д.) необходима для исследования интересующего биологического процесса, оптимизируйте условия (например, время, концентрацию) с помощью визуализации живых клеток FLM.
- Убедитесь, что интересующие участки могут быть успешно локализованы над фоном в достаточном количестве клеток, используя настройки визуализации, которые максимально соответствуют более поздним криоусловиям (т.е. NA, время экспозиции и т. д.).
- Выбор и подготовка сеток
- Выберите сетки с размером отверстия и расстоянием между ними, соответствующими ячейкам и используемым реперным меткам (см. шаг 1.3.1). Не используйте сплошную пленку без отверстий, так как это может привести к слишком большому остаточному буферу после промокания и, таким образом, снизить эффективность витрификации и затруднить обнаружение реперных шариков. Для длительного контакта клеток с сетками убедитесь, что опора решетки и материал пленки являются биосовместимыми.
- Очистите крио-ЭМ решетки плазмой, чтобы сделать их более гидрофильными. Для использования в адгезивных клеточных культурах стерилизовать решетки после плазменной очистки УФ-излучением в течение 20 мин в ламинарном колпаке. Опционально сетки могут быть предварительно обработаны соединениями, которые способствуют адгезии клеток, такими как поли-L-лизин или конканавалин А, как описано ниже.
ПРИМЕЧАНИЕ: Как правило, в коррелятивном рабочем процессе крио-FIB успешно используются следующие комбинации сетки/образца: Дрожжи: Cu или Au, 200 меш, углерод R1/4 или пленка SiO2 , опционально покрытая конканавалином А; Кишечная палочка: Cu или Au, 200 меш, пленка R1/4 углерода илиSiO2 ; Chlamydomonas reinhardtii: Cu или Au, 200 меш, R1/2 или R1/4 углерод или пленка SiO2 ; HeLa: Au, 200 меш, пленка R1/4 SiO2 , покрытая поли-L-лизином; HEK293: Au, 200 меш, пленка R1/4 SiO2 , покрытая поли-L-лизином. - Покрытие Concanavalin A для улучшения прикрепления дрожжевых клеток:
- Приготовьте покровный раствор 1 мг/мл конканавалина А в 10 мМ буфере HEPES с 100 мкМ CaCl2, pH 8,5. Поместите одну каплю (50 мкл) раствора покрытия и две капли дистиллированной воды отдельно на кусок парафиновой пленки.
- Возьмите очищенную плазмой сетку пинцетом с обратной силой и осторожно вставьте ее в каплю раствора покрытия, избегая движений перпендикулярно сетке, чтобы не повредить пленку.
- После ~5 с инкубации промойте решетку два раза, вставив ее в капли воды аналогичным образом. Наконец, промокните лишнюю жидкость, приложив фильтровальную бумагу к обратной стороне решетки, и дайте решетке полностью высохнуть, прежде чем выпускать ее из пинцета. Используйте высушенные сетки для погружной заморозки.
- Поли-L-лизиновое покрытие для суспензионных культур и адгезивных клеток:
- Готовят покровный раствор 1 мг/мл поли-L-лизина в 0,1 М буфере бората натрия, рН 8,5.
- Поместите очищенные плазмой решетки в подходящую чашку для клеточной культуры и стерилизуйте в течение 20 минут УФ-излучением.
- Аккуратно добавьте достаточное количество раствора для покрытия, чтобы покрыть все решетки, и инкубируйте при 37 °C не менее 2 часов. Аспирируйте жидкость и осторожно промойте решетки два раза PBS перед посевом клеток до желаемой концентрации.
- Подготовка ячеек и реперных шариков
ПРИМЕЧАНИЕ: Реперные шарики необходимы для 3D-регистрации данных флуоресценции с изображениями, полученными в микроскопе FIB/SEM, чтобы обеспечить 3D-коррелятивное фрезерование FIB.- Выбирайте шарики, распознаваемые во всех методах визуализации, т.е. FLM, SEM и IB (рекомендуемый диаметр 0,5-1 мкм), но убедитесь, что они не затмевают клеточную целевую структуру во время флуоресцентной визуализации, чтобы было легче дифференцировать шарики и интересующие биологические особенности. Удаляйте цитотоксические консерванты в реперных шариках (например, NaN3) в соответствии с инструкциями производителя.
ПРИМЕЧАНИЕ: Для более легкого различения интересующих биологических особенностей и реперных точек полезно, чтобы спектры флуоресцентного излучения перекрывались лишь частично, чтобы сигналы можно было различать на основе различий интенсивностей в каналах FLM. - Если используется суспензионная культура, вырастите клетки до подходящей плотности (например, дрожжи OD 600 = 0,8, E. coli OD 600 = 0,8-1,0, C. reinhardtii 1500 клеток/мкл) и выполните обработку, необходимую для эксперимента, такую как смена среды, добавление химикатов, голодание и т. д. Прикрепите очищенные плазмой сетки к пинцету в соответствии с требованиями метода погружения (ручной/автоматический) и нанесите 4 мкл клеточной суспензии, смешанной с ~1 x 105 шариков/мкл реперной суспензии шариков, на пленочную сторону сеток.
ПРИМЕЧАНИЕ: Определите оптимальное разведение клеток и реперных точек в экспериментах по титрованию (например, проверив крио-FLM или FIB/SEM, см. ниже). Тем не менее, для большинства клеток, выращенных в суспензии, окончательная концентрация ~1 x 105 шариков/мкл реперных шариков размером 1 мкм (разведение 1:20 от запаса; см. Таблицу материалов для получения подробной информации) оказалась хорошей отправной точкой. - Если используется адгезивная культура, решетки очищают плазмой и стерилизуют с помощью УФ-излучения для асептической культуры. При необходимости предварительно покройте решетки составами, способствующими адгезии клеток (например, поли-l-лизином, фибронектином, ламинином; см. шаг 1.2.2). Высевайте и выращивайте клетки на сетках в обычных чашках для культивирования или чашках с подразделениями для сеток.
- Обращайтесь с образцами так, как это необходимо для эксперимента, и поддерживайте клетки в оптимальных условиях до полного замораживания (например, 37 °C/5% CO2 для HEK/HeLa). Аккуратно снимите сетки с чашки для культур и прикрепите их к пинцету. Нанести 4 мкл питательной среды, смешанной с реперными речами (1 x 105 шариков/мкл для 1 мкм реперных точек) на клеточную сторону.
- Выбирайте шарики, распознаваемые во всех методах визуализации, т.е. FLM, SEM и IB (рекомендуемый диаметр 0,5-1 мкм), но убедитесь, что они не затмевают клеточную целевую структуру во время флуоресцентной визуализации, чтобы было легче дифференцировать шарики и интересующие биологические особенности. Удаляйте цитотоксические консерванты в реперных шариках (например, NaN3) в соответствии с инструкциями производителя.
- Погружная заморозка клеток с помощью ручной или автоматизированной процедуры замораживания. По возможности промокайте сетку только со стороны, противоположной ячейкам, чтобы предотвратить механическое повреждение клеток (Рисунок 2А). В автоматизированных погружных системах с двумя руками этого можно добиться, поместив лист политетрафторэтилена (например, тефлона) вместо промокательной бумаги на прокладку, обращенную к ячейкам. Переложите решетки в ящики для хранения и храните их в жидком азоте (lN2) до использования.
ВНИМАНИЕ: lN2 и другие криогены могут вызвать серьезное повреждение глаз и кожи. Используйте средства индивидуальной защиты (СИЗ) и работайте только в хорошо проветриваемом помещении, чтобы избежать накопления опасных концентрацийN2 .
ПРИМЕЧАНИЕ: Все последующие этапы лучше всего выполнять в жидкой фазе lN2 , чтобы избежать загрязнения поверхности ячейки и ламелей, так как это может усложнить последующую обработку. Уменьшите контакт с плавающими кристаллами льда, всегда используя чистый жидкий азот (например, фильтр для удаления плавающего льда), исключив ненужные этапы переноса и, по возможности, работая в среде с контролируемой влажностью. - Установите и закрепите погружно-замороженные сетки в AutoGrids с вырезом и ячейками вверх (рис. 2A) для последующей криофлуоресцентной визуализации и фрезерования FIB. Чтобы обеспечить правильное выравнивание образцов в ПЭМ, направление измельчения должно быть ортогонально оси наклона крио-ET. Соответственно, разместите метки ориентации (например, лазерную гравировку или съемные маркерные точки) на автосетках перед обрезкой, чтобы облегчить это выравнивание (рис. 2A).
- Проверьте качество сетки (рис. 2B) на крио-FLM и FIB/SEM. Оптимизируйте плотность клеток, время и усилие промокания, чтобы получить равномерное распределение клеток и шариков. Используйте изображение отраженного света на криофлуоресцентном микроскопе или используйте крио-FIB-SEM, чтобы убедиться, что и клетки, и реперные шарики хорошо видны (рис. 2B, белые стрелки).
- При необходимости повторите погружение в более благоприятные условия, например, изменяя концентрацию клеток и/или время промокания. После того, как подходящие параметры погружения найдены, не повторяйте скрининг сетки для каждого нового раунда экспериментов.

Иллюстрация 2: Просеивание подходящих сеток с помощью SEM и IB . (A) Метки ориентации должны быть размещены на автосетках перпендикулярно направлению фрезерования, чтобы упростить правильную загрузку в ПЭМ. Ячейки монтируются лицевой стороной вверх в собранную AutoGrid. (B) После погружного замораживания решетки проверяются в SEM для оценки и оптимизации условий погружения: a) В сетке не должно быть слишком много ячеек. Например, для клеток HeLa не используйте более 1-4 клеток на квадрат. Для более мелких клеток, таких как Saccharomyces cerevisiae (см. здесь), были признаны полезными скопления из 4-6 клеток. б) Реперные шарики (белые стрелки) должны быть хорошо видны, и вокруг ячеек не должно быть слишком много буфера. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
2. Криофлуоресцентная световая микроскопия
- Для каждой сетки получите обзор в (широкопольном) флуоресцентном и дифференциально-интерференционном контрасте (DIC) или отраженном режиме и выберите подходящие квадраты сетки с флуоресцентным сигналом. Выбирайте поля зрения, которые содержат как интересующие ячейки, так и достаточное количество реперных маркеров (6-12).
- Следите за тем, чтобы ячейки и бусины были распределены равномерно, не слишком плотно и ближе к центру каждого квадрата. Выбирайте только те квадраты, которые доступны как для прибора FIB-SEM, так и для TEM, т.е. те, которые находятся на расстоянии не менее трех квадратов от края сетки на сетке 200 сеток (рис. 3A, внутри красного круга).
- На каждом из выбранных квадратов сетки получите флуоресцентный стек с шагом фокусировки, подходящим для последующей деконволюции, т. е. <1/2 предела осевого разрешения. Если возможно, используйте объективы с высокой числовой апертурой (NA), чтобы увеличить количество фотонов и точность локализации.
- В конфокальном микроскопе с объективом NA 0,9 регистрируйте стеки с шагом 300 нм, превышая значение Найквиста. При необходимости запишите несколько цветовых стеков (рис. 2B). Храните сетки под lN2 до дальнейшего использования.
ПРИМЕЧАНИЕ: Чтобы определить оптимальный размер шага, выберите значения, рассчитанные программным обеспечением для управления микроскопом, или используйте онлайн-инструменты9. Проверьте наличие просачивания сигнала между каналами, так как чрезмерный протекание вреден для экспериментов по колокализации. Тем не менее, некоторые из них могут быть полезны для коррекции хроматических аберраций в многоцветных стеках.
- В конфокальном микроскопе с объективом NA 0,9 регистрируйте стеки с шагом 300 нм, превышая значение Найквиста. При необходимости запишите несколько цветовых стеков (рис. 2B). Храните сетки под lN2 до дальнейшего использования.
- Деконволютируйте стеки с помощью соответствующего программного обеспечения10,11 и повторно нарежьте7 из них, если требуется изотропный размер пикселя. Деконволюция, как и при комнатной температуре, очищает сигнал FLM и может повысить точность локализации (рис. 3C).

Рисунок 3: Выбор квадратов для сбора стека FLM и улучшение данных с помощью деконволюции. (A) Обзор сетки, погруженной в дрожжевые клетки, экспрессирующие eGFP-Ede1 (зеленый) и mCherry-Atg8 (пурпурный). Выбирайте позиции с хорошим распределением бусин и ячеек, но избегайте краев сетки (заштрихованы красным). Прямоугольниками обозначены позиции с хорошим распределением клеток, где были взяты флуоресцентные стеки. (B) Проекция максимальной интенсивности (MIP) многоцветного стека, взятая на квадрат с желтым прямоугольником (из А) после деконволюции. Деконволюция стеков FLM значительно очищает нежелательные фоновые сигналы и помогает более точно локализовать бусины в z, как видно из гауссовских аппроксимаций до (C) и после деконволюции (D) (аппроксимация была выполнена в 3DCT и показана для валика, отмеченного цифрой 1). На изображениях показаны увеличенные MIP-изображения красного канала (возбуждение: 552 нм, излучение: 585-650 нм). Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
3. Фрезерование сфокусированным ионным пучком
- Загрузите сетки в прибор cryo-FIB-SEM и используйте метки выреза и/или ориентации, чтобы обеспечить правильную ориентацию для последующего размещения в ПЭМ (Рисунок 2A). Убедитесь, что направление фрезерования перпендикулярно оси наклона ПЭМ.
- Используйте систему впрыска газа (ГИС; CpMePtMe3) в положениях ступени, заданных установкой FIB-SEM для покрытия решеток защитным металлоорганическим слоем. Не наносите слишком много, так как это может повлиять на локализацию реперного валика в ПЭМ в дальнейшем. Используйте установку для нанесения плазменного покрытия на металлическую платину, чтобы уменьшить зарядку образца.
ПРИМЕЧАНИЕ: Если настройки для покрытия GIS недоступны, их можно легко найти, выполнив последовательные циклы короткого покрытия (~2 с) с последующим фрезерованием FIB. Убедитесь, что образец по-прежнему может быть успешно разрезан при средних токах (~100 пА) без видимой окантовки защитного металлоорганического слоя по краям ламелей. Как время, так и расстояние стрелки ГИС (по отношению к образцу) являются важными параметрами, которые следует учитывать. Не используйте иглу GIS при комнатной температуре (например, 45 °C), но как можно холоднее, чтобы обеспечить равномерное покрытие (25-27 °C). - Запишите обзор сетки SEM и выполните 2D-корреляцию с обзорами FLM, чтобы найти квадраты сетки, для которых были записаны флуоресцентные стеки. Вручную проверьте оба вида или используйте различные программные пакеты 7,10,12 для поиска квадратов сетки. В данном случае основное внимание уделяется набору инструментов 3D-корреляции (3DCT)7, который использует 3D-трансформацию твердых тел с изотропным масштабированием между видами. Превосходное руководство по функциям 3DCT доступно в Интернете13.
- Выберите и отметьте не менее четырех соответствующих позиций, например, ориентиры, такие как стержни сетки или отверстия в опорной пленке, в обзоре сетки FLM и SEM (щелчок правой кнопкой мыши) и рассчитайте трансформацию между отмеченными точками (коррелят).
- Затем поместите маркеры в центр соответствующих квадратов сетки, для которых были получены стеки FLM, и спрогнозируйте их положение в представлении SEM (коррелировать; Рисунок 4А).
- Для каждого коррелированного квадрата сетки сделайте изображение слаботочного (≤10 пА) ионного пучка (IB) под выбранным углом фрезерования FIB (10°-25° для челнока с предварительным наклоном 45°). Выберите поле зрения (т. е. положение и увеличение), которое соответствует данным флуоресценции. Для сеток с 200 сетками собирайте данные флуоресценции и FIB/SEM, содержащие одиночные квадраты сетки, включая полосы сетки (см. рис. 3A и рис. 4A).
ПРИМЕЧАНИЕ: Фрезерование должно выполняться под как можно меньшим углом, чтобы избежать потери значительного углового диапазона во время крио-ET и позволить идентифицировать достаточное количество реперных шариков. Например: при наклоне ступени 17°, предварительном наклоне челнока 45° и наклоне балки FIB 52° относительно отвеса, предварительный наклон ламели составляет 10°, что почти соответствует предпочтительному угловому диапазону ±60° в ПЭМ при наклоне от -50° до +70°, максимальному для многих криодержателей ТЕМ. - Сделайте СЭМ-изображение того же квадрата, чтобы помочь идентифицировать соответствующие шарики во флуоресцентном и ионном лучах.
- Выполните регистрацию деконволюрированного стека 3D FLM и вида 2D ионного пучка для каждой позиции с помощью 3DCT, как описано в следующих шагах (рис. 4B).
- Загрузите соответствующий перерезанный стек 3D FLM и вид ионного пучка (IB) в 3DCT.
ПРИМЕЧАНИЕ: Многоцветные флуоресцентные данные могут быть загружены в виде трех отдельных одноканальных стековых файлов. - Выберите 4 реперных шарика в данных флуоресценции и щелкните правой кнопкой мыши по списку позиций, чтобы определить их 3D-положение с помощью гауссовской аппроксимации сигнала по осям, осям и z. Выделите соответствующие бусины на изображении IB и выполните начальную 3D-корреляцию (корреляцию).
- Итеративно добавляйте больше бусин в флуоресцентное изображение, уточняйте их 3D-положение и прогнозируйте их положение в представлении IB, чтобы быстро добавить больше бусин в регистрацию и проверить точность корреляции. В 3DCT для оценки согласованности корреляции предоставляются значения среднеквадратичной ошибки (RMSE)7.
- Убедитесь, что значения RMSE малы и находятся на уровне точности локализации (~300 нм). Чтобы определить точность корреляции, намеренно опустите некоторые реперные шарики, четко идентифицируемые как во флуоресценции, так и в ионном пучке на этапе регистрации. Для этого сравните их прогнозируемое и фактическое расположение на изображении ионного пучка. Если прогнозируемое положение значительно отличается от реального, повторите первоначальную корреляцию с новым набором реперных точек.
ПРИМЕЧАНИЕ: Корреляция 6-8 бусин оказалась достаточной для точной регистрации стеков FLM и представлений IB. Тем не менее, добавление большего количества реперных точек (до 12-15) в широком диапазоне значений z (например, путем выбора бусин на полосе сетки или в соседних квадратах) может повысить точность корреляции.
- Убедитесь, что значения RMSE малы и находятся на уровне точности локализации (~300 нм). Чтобы определить точность корреляции, намеренно опустите некоторые реперные шарики, четко идентифицируемые как во флуоресценции, так и в ионном пучке на этапе регистрации. Для этого сравните их прогнозируемое и фактическое расположение на изображении ионного пучка. Если прогнозируемое положение значительно отличается от реального, повторите первоначальную корреляцию с новым набором реперных точек.
- Выберите целевые сигналы сотовой связи, подогнайте их 3D-положение в стеке FLM и примените преобразование для прогнозирования целевых положений в представлении IB (рис. 4B).
ПРИМЕЧАНИЕ: Любая запись в списке позиций FLM, которая не имеет аналога в списке IB, будет рассматриваться как сигнал, подлежащий прогнозированию.
- Загрузите соответствующий перерезанный стек 3D FLM и вид ионного пучка (IB) в 3DCT.
- Для каждого коррелированного квадрата перенесите прогнозируемые положения интересующих объектов в прибор FIB-SEM и разместите шаблоны фрезерования ламелей (рис. 4C). Переносите положения вручную (например, измеряя расстояние до видимых ориентиров, таких как ячейки или реперные шарики) или используйте автоматизацию и сценарии, как это реализовано, например, в SerialFIB14. Если на ячейку приходится несколько сигналов, разместите шаблоны так, чтобы включить как можно больше точек интереса (POI) в одну и ту же ламель, чтобы увеличить пропускную способность.
- Сначала грубо, а затем тонко фрезеруют ламели до конечной толщины 150-250 нм. Избегайте шагов (например, разрезов для снятия напряжений15), которые вызывают провисание ламели и, таким образом, приводят к смещению фактического интересующего элемента по отношению к ранее полученным стопелям FLM. Используйте либо ручные3, либо автоматизированные процедуры фрезерования14,16,17,18 FIB. При любом методе убедитесь, что интересующий элемент остается в центре ламели, симметрично утончая ее сверху и снизу.
- Чтобы оценить точность фрезерования для каждой ламели, выполните ту же регистрацию, что и на шаге 3.4. Однако на этот раз используйте окончательное изображение IB после фрезерования FIB и проверьте, содержатся ли прогнозируемые положения интересующих объектов в конечной ламели. В качестве альтернативы можно наложить повернутые проекции стеков FLM, полученные из выходных данных 3DCT и пользовательских сценариев19, с окончательным изображением IB (рис. 4C, маленькая вставка).

Рисунок 4: 3D-коррелятивная процедура фрезерования FIB . (A) 2D-2D корреляция FLM (слева) и SEM (справа) обзоров сетки используется для определения местоположения квадратов сетки, на которых ранее были сделаны флуоресцентные стеки. (B) Для каждого выбранного квадрата, после 3D-2D регистрации соответствующих реперных точек в 3DCT (цветные прямоугольники), в данных FLM выбираются позиции интересующих биологических особенностей. На основе предсказания соответствующих положений на изображении ионного пучка (красные кружки) выбираются места для подготовки ламелей. (C) Изображения сканирующего электронного микроскопа (SEM) и ионного пучка (IB) используются для удержания мишени в центре во время фрезерования. Конечная толщина 150-250 нм была признана достаточной для дальнейшей переработки. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
4. Коррелятивный ПЭМ
- Загрузите сетки в ПЭМ, убедившись, что ориентация ламелей (как видно из вырезов или меток ориентации) перпендикулярна оси наклона.
ПРИМЕЧАНИЕ: Микроскопами разных производителей можно управлять с помощью различного программного обеспечения, например, Tomo5, TOM или SerialEM20. Здесь основное внимание уделяется последнему. - Получите монтаж сетки и обзоры для каждого квадрата сетки, содержащего ламели. Убедитесь, что увеличение и время экспозиции подходят для визуализации реперных шариков на изображениях ПЭМ без значительного увеличения общей дозы электронов. Получение ПЭМ-карт (монтажей) каждой ламели с высоким разрешением.
- Приводка и 3D-2D сопоставляют стек FLM с квадратом сетки TEM и обзорами ламелей в 3DCT. Используйте ту же процедуру, что описана в шаге 3.6, выбрав соответствующие положения шариков во флуоресценции (x, y, z - гауссова подгонка) и изображениях просвечивающего электронного микроскопа. Затем выберите интересующие позиции в каналах FLM и перенесите их в обзоры TEM. При необходимости используйте двухступенчатую процедуру, включающую в себя первую корреляцию между FLM и ПЭМ с малым увеличением, а вторую — между ПЭМ с малым и большим увеличением (рис. 5).
- Переносите координаты вручную (путем измерения расстояний до ориентиров), с помощью инструментов регистрации и картографирования, доступных в SerialEM20, или внешнего программного обеспечения, такого как CorRelator21.
- Настройка и запуск серии наклонов в коррелированных положениях. Используйте соответствующее увеличение, расфокусировку и общую дозу (подробности см. в таблице материалов и таблице 1 ). Съемку начинают с предварительного наклона, определяемого ламелем (см. также примечание на шаге 3.4) и используют дозосимметричную схему наклона22. Используйте ручную или пакетную сборку.

Рисунок 5: Локализация коррелированных позиций в ПЭМ. После успешного 3D-коррелятивного фрезерования FIB и переноса на просвечивающий электронный микроскоп выполняется 3D-2D-регистрация для каждого фрезерованного квадрата между реперными шариками (цветными прямоугольниками) в стеках FLM и обзорах ПЭМ для локализации потенциальных участков крио-ЭТ (красные круги). Затем можно получить более высокие обзоры ламелей (увеличение) для более точной настройки томограмм. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
Результаты
Протокол предусматривает обход конвейера, используемого для обнаружения EH-домен-содержащего и эндоцитоз-зависимого белка 1 (Ede1)-зависимого эндоцитарного белкового отложения (END), а также его деградации и захвата в аутофагических телах8. END представляет собой фазово-разделенный компартмент S. cerevisiae, который буферизует различные белки, участвующие в клатрин-опосредованном эндоцитозе (CME) после неудачных эндоцитарных событий. Одним из его основных компонентов является Ede1, который одновременно является компонентом CME и селективным рецептором аутофагии для деградации этого нового компартмента LLPS. Соответственно, для визуализации ЭСДН использовали EGFP-слияние Ede1 (EGFP-Ede1) под контролем промотора алкогольдегидрогеназы (АДГ), поскольку гиперэкспрессия Ede1 мешает ранним стадиям эндоцитоза и, следовательно, конститутивно индуцирует LLPS.
На погружно-замороженной сетке с гиперэкспрессией дрожжевых клеток EGFP-Ede1 и реперными маркерами 1 мкм были выбраны пять позиций для регистрации стека FLM в канале GFP (рис. 6A; TFS Corrsight; конфокальный режим, шаг фокусировки 300 нм, диапазон 10 мкм). Сетка была перенесена в прибор FIB (Quanta 3D FEG), и квадраты сетки, для которых были получены стеки FLM, были идентифицированы путем выполнения 2D-2D корреляции обзоров флуоресценции и сетки SEM (сравните шаг 3.2).
Для каждого из выбранных квадратов были получены изображения ионным пучком при слабом токе (10 пА, 1200-кратное увеличение), и соответствующие реперные точки были зарегистрированы в 3DCT. После отбора позиций с интересующей биологической особенностью и подгонки их 3D-положения в стеке FLM, найденное преобразование было применено к предполагаемым позициям END, и были выбраны места для подготовки ламелей (рис. 6B). В приведенных здесь примерах использовалась балка FIB, наклоненная на 11° относительно поверхности сетки (предварительный наклон челнока FIB 45°; наклон ступени 18°). Интересующие позиции были перенесены, и паттерны FIB были нарисованы вручную (рис. 6D) путем измерения расстояния прогнозируемых позиций относительно заметных ориентиров на изображении FIB (например, лунки, ледяные загрязнения, реперные шарики). Точность регистрации оценивалась путем преднамеренного исключения бусин, которые могли быть четко идентифицированы на изображениях FLM и IB, а затем сравнения их фактического и прогнозируемого положения в виде ионного пучка (например, алмаз на рисунке 6B, C). Корреляция для квадрата, показанного на рисунке 6C , оказалась точной (т.е. предсказанное положение бусин FLM идеально совпадало с их соответствующим положением IB, а 3DCT сообщал субпиксельные среднеквадратичные значения для регистрации). Таким образом, ламели были разрезаны в прогнозируемых положениях (участок Б) и тонко измельчены до толщины ~200 нм (окончательное смещение шаблона).

Рисунок 6: Репрезентативные результаты для 3D-коррелятивного таргетирования эндоцитарных белковых отложений (END) у дрожжей. (A) Обзор решетки перед фрезерованием с помощью СЭМ. Цветными прямоугольниками обозначены квадраты сетки, для которых заранее были взяты флуоресцентные стеки. (B-C) 3D корреляция в квадрате сетки. После регистрации нескольких соответствующих реперных шариков (цветных прямоугольников) в данных FLM (B, показанная здесь как проекция максимальной интенсивности) и изображении ионного пучка (C), точность 3D-регистрации была проверена путем прогнозирования положения шарика, обозначенного алмазом. Затем положения целевого сигнала (красные кружки) были предсказаны в виде ионного пучка для двух потенциальных участков фрезерования. (D) Увеличение Участка Б, показывающее прогнозируемые положения трех целевых точек (красные круги) и начальные паттерны фрезерования (желтые прямоугольники). Было предсказано, что четвертый флуоресцентный пунктум будет намного ниже, чем другие пункты, и, следовательно, не будет нацелен во время фрезерования (серый круг). Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
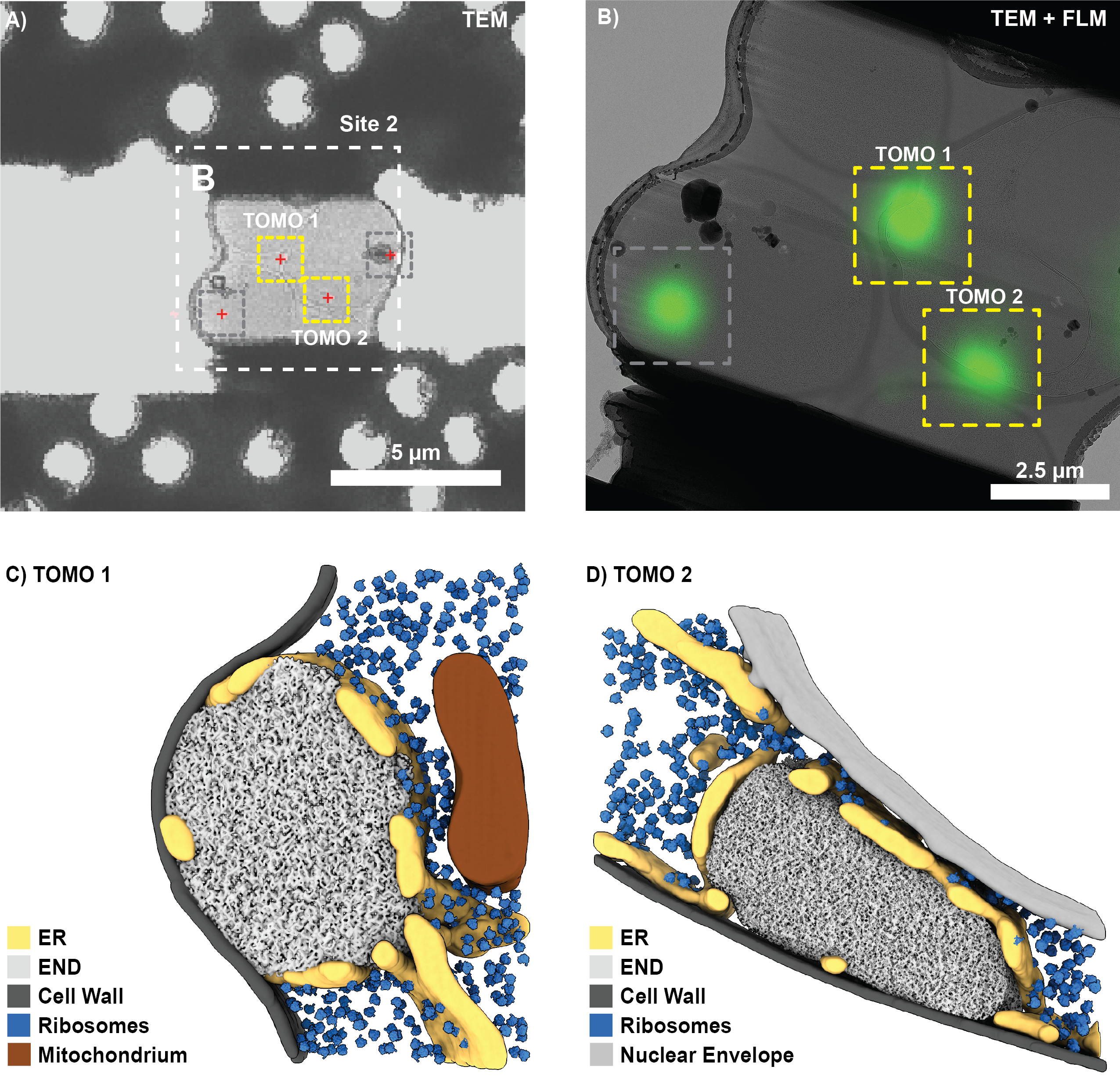
После успешного фрезерования FIB и переноса сетки на криопросвечивающий электронный микроскоп (Titan Krios работал на 300 кВ и был оснащен прямым электронным детектором Gatan K2 и биоквантовым энергетическим фильтром) в SerialEM был записан обзор сетки, который использовался для определения местоположения квадратов с ламелями. Для каждой ламели были получены обзорные изображения, а данные FLM были зарегистрированы в 3DCT (3D-2D) с использованием соответствующих реперных шариков. Затем положение интересующих биологических признаков (рис. 7А) было предсказано с помощью преобразования, рассчитанного по реперным шарикам. Обзоры ламелей, записанные при большем увеличении, были сшиты, а интересующие места соотнесены с помощью хорошо видимых ориентиров (например, реперных бусин). В качестве альтернативы классические обзоры CLEM могут быть созданы в различных программах10,12.
На основе корреляции были найдены четыре потенциальных участка для получения томограммы для ламели, показанной на рисунке 7А. Тем не менее, это также включает в себя положение, которое не было нацелено во время 3D-коррелированного фрезерования FIB (сравните Рисунок 6D; серый круг) и положение, заблокированное ледяным загрязнением (Рисунок 7; серые прямоугольники). Соответственно, томограммы могут быть записаны только для двух позиций (рис. 7Б). В целом был достигнут корреляционный успех ~75%, т.е. на предсказанных участках были найдены ламели, пережившие переход в структуры ПЭМ и ЭНД (12 коррелированных участков). После реконструкции томограммы, сегментации и сопоставления шаблонов отдельные структуры END могут быть визуализированы в их нативном контексте (рис. 7C, D). К ним относятся фенестрированный эндоплазматический ретикулум (ER), окружающий END, липидные капли, время от времени вступающие в контакт, и рибосомы, которые исключаются из компартмента LLPS. В совокупности это показывает, как 3D-коррелятивное измельчение FIB может предоставить информацию на молекулярном уровне о редких биологических процессах в интактных клетках.

Рисунок 7: Репрезентативные результаты визуализации END с помощью крио-ET. (A) Обзор участка измельчения с малым увеличением, показанный на рисунке 6, может быть легко соотнесен с проекцией максимальной интенсивности FLM (рисунок 6B) для локализации интересующих биологических особенностей (красные кресты). (B) На втором этапе можно сопоставить изображение с большим увеличением (сшитое) и установить положения для получения томограммы (желтые прямоугольники). Местоположения, полученные в результате внеплоскостного сигнала (серый прямоугольник, сравните рис. 6D), игнорировались. (К-Д) Используя этот 3D-коррелятивный подход FIB, эндоцитарный белковый депозит (END) может быть визуализирован в его нативной среде. Такие структуры, как эндоплазматический ретикулум (ER), рибосомы, мембраны и липидные капли, могут быть идентифицированы и визуализированы. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
| Настройки плазменного очистителя | |||
| Плазменный очиститель Harrick PDG-3XG : | Радиочастотная передача: "HI", 30 с; Плазма N2 | ||
| Настройки плунжера | |||
| TFS Vitrobot Mk IV: | 100% влажность; блотфорс = 8; блоттайм = 10 с; время ожидания 0 с; (это должно работать для большинства суспензионных и адгезивных клеток) | ||
| Координаты и тайминги FIB GIS | |||
| Quanta 3D FEG: | Наклон = 0, Поворот = -180, Положение Z = 13,5, Заданное значение температуры = 26,15° , Время = 8 с | ||
| TFS Scios: | Наклон = 0, Вращение = -180, Положение Z = 9,8, Заданное значение температуры = 28° C, Время = 7 с | ||
| TFS Aquilos 1: | Предустановленное программным обеспечением положение, заданное значение температуры = 28° , время = 7 с | ||
| TFS Aquilos 2: | Предустановленное программным обеспечением положение, заданное значение температуры = 28°, время = 7 с | ||
| Настройки установки для нанесения покрытий FIB | |||
| Система кворума: | В камере подготовки кворума: 10 мА, 40 с | ||
| TFS Scios: | 10 Вт, 500 В, 250 мА, 0,2 мбар, 15 с | ||
| TFS Aquilos 1: | 1кВ, 10 мА, 10 Па, 15 с | ||
| TFS Aquilos 2: | 1кВ, 10 мА, 10 Па, 15 с | ||
| Получение томограммы | |||
| Титан Криос Gi2 | Камера К2, энергетический фильтр Gatan Bioquantum | ||
| щель 20 эВ; схема симметричного наклона дозы (Хагена) с шагом 2°; Начните с +10° (предварительный наклон ламели!) до +70° и -50° | |||
| Титан Криос Gi4 | Сокол 4; Энергетический фильтр Selectris X | ||
| щель 10 эВ; схема симметричного наклона дозы (Хагена) с шагом 2°; Начните с +10° (предварительный наклон ламели!) до +70° и -50° | |||
| Приобретение FLM | |||
| Corrsight (конфокальный режим) | Цель: Zeiss EC Plan-Neofluar 40×/0.9 NA Pol; Параметры сбора стека: размер пикселя x-y = 161,25 нм, размер шага z = 300 нм. | ||
| Криоконфокальный монитор Leica SP8 | Объектив: Leica HCX PL APO 50x / 0,90 CLEM; Параметры сбора стека: размер пикселя x-y = 84 нм, размер шага z = 300 нм. | ||
Таблица 1: Перечень тестируемого оборудования и предлагаемые настройки.
Обсуждение
1. Критические шаги в протоколе
Оптимизация параметров клеточной культуры и погружения сетки имеет основополагающее значение для этого рабочего процесса. В начале проекта стоит потратить время на оптимизацию стратегий маркировки, распределение ячеек и реперных шариков, а также на тестирование различных параметров подготовки сетки и промокания. Работа с оптимально замороженным образцом значительно упростит последующую обработку.
Как и для любого эксперимента ПЭМ, необходимы образцы стекловидного тела. Для крупных клеток млекопитающих, таких как HeLa, предпочтительнее 1-2 клетки на квадрат сетки, но клетки все еще могут быть стекловидными при более высокой плотности. Опционально витрификация может быть улучшена в клетках млекопитающих (например, HEK293, HeLa) путем инкубации их с 2,5-10% (v/v) глицерином, добавленным в питательную среду за 10 мин до погружения23. Если это возможно, можно использовать сетчатый узор для обеспечения идеального размещения и распределения клеток, тем самым улучшая витрификацию и последующую корреляцию24.
В то время как определенные клетки могут быть выбраны во время рабочего процесса, слишком малое количество клеток, демонстрирующих интересующую биологическую особенность, значительно снизит общую пропускную способность. Для улучшения корреляции в POI-положительных клетках следует использовать достаточно яркие флуорофоры. Это особенно важно на уровнях эндогенной экспрессии. Мы обнаружили, что в криоусловиях mVenus часто работает лучше, чем EGFP, благодаря своей повышенной яркости25 и гипсохромному сдвигу, что делает ее пригодной для стандартных установок GFP-фильтров в криоусловиях26. Для неточечных целевых структур также следует учитывать компромисс между длиной волны и точностью локализации (дифракционный предел Аббе).
Эффективная 3D-корреляция также требует, чтобы сетки были механически стабильными и обрабатывались с большой осторожностью. В то время как стандартные золотые или медные сетки с углеродной подложкой могут быть использованы, вероятность успеха может быть значительно увеличена за счет использования более жестких пленок SiO2 в зависимости от проекта. Тем не менее, до сих пор окончательно не определено, является ли (а) механическая стабильность или (б) согласование коэффициентов теплового расширения (подложка и пленка) для уменьшения криосморщивания27 наиболее важным фактором для успешной 3D-корреляции. Кроме того, для сбора хрупких сеток Au можно использовать посуду, покрытую полидиметилсилоксаном5.
Помимо обеспечения стабильности образца, для получения высококачественных флуоресцентных стеков, пригодных для оптимального нацеливания при фрезеровании FIB, необходим тщательный выбор параметров визуализации FLM. В связи с этим также рекомендуется тестировать различные методы шумоподавления28 или деконволюции на данных FLM, так как это может значительно улучшить локализацию реперных и сотовых сигналов. При корреляции флуоресцентного сигнала с изображениями FIB-SEM важна хорошая выборка реперных шариков. Они должны быть хорошо распределены по ячейкам и, возможно, на разной высоте z. Также хорошей практикой является проверка согласованности корреляции путем проверки прогнозируемого и фактического положения бусин, которые были намеренно исключены из реперной модели, но могут быть четко соотнесены на глаз. Среднеквадратичные значения 3DCT также всегда следует учитывать для проверки согласованности регистрации.
Поскольку осаждение измельченного материала и остаточной воды из камеры FIB-SEM (т.е. повторное загрязнение) увеличивает эффективную толщину ламели за счет добавления аморфного материала с обеих сторон от нее, хранение тонко измельченных ламелей в микроскопе в течение длительного времени, как правило, снижает качество данных ПЭМ из-за дополнительных событий рассеяния электронов. Соответственно, фрезерование чаще всего выполняется в два этапа: сначала все позиции фрезеруются грубо (т.е. примерно до 800 нм), а затем тонко (до ~150-250 нм), и сетка сразу же выгружается после того, как последняя ламель будет готова. Тем не менее, лучшего успеха корреляции можно достичь, обрабатывая интересующие позиции по участку, поэтому выполняя грубое и тонкое фрезерование на одной и той же ламели непосредственно друг за другом, поскольку это не оставляет времени на изгиб или деформацию. Это, однако, уменьшает максимальное количество ламелей, которые могут быть произведены на решетку, в зависимости от скорости повторного загрязнения системы. При скорости 20 нм/ч в течение 1-1,5 ч производится 4-6 ламелей.
Смещение всей сетки или грубо фрезерованных ламелей >300 нм приведет к плохой или неудачной корреляции (см. также ограничения, обсуждаемые ниже). Поэтому его следует регулярно проверять, например, сравнивая изображения IB до, во время и после фрезерования FIB. Участки, которые показывают значительное движение (>300 нм), должны быть отброшены. Оптимизируйте пробоподготовку (т. е. выбор типа сетки, плотности ячеек и параметров погружения; см. раздел протокола 1) и стратегию измельчения, чтобы избежать этих перемещений. Изгиб ламелей может быть значительно уменьшен за счет фрезерования на месте, как описано в шаге 3.6, и уменьшения ширины ламели. Как упоминалось ранее, несмотря на то, что разрезы для снятия напряжений15 были разработаны для уменьшения изгиба ламелей, они часто приводят к согласованному движению разъединенных ламелей, тем самым эффективно предотвращая корреляцию. Для решения этой проблемы могут быть использованы интегрированные системы FLM.
2. Модификации и устранение неисправностей метода
Настоятельно рекомендуется провести тщательную характеристику образца при визуализации живых клеток перед переходом в криоусловия. Оптимизация клеточных образцов, схем лечения и знание того, какого сигнала следует ожидать до начала криорабочего процесса, может существенно повысить его успешность.
В представленном здесь рабочем процессе для получения изображений образцов используется автономный флуоресцентный микроскоп с криостоликом с последующим переносом сеток в сфокусированный ионно-лучевой микроскоп. Тем не менее, он был протестирован на системах, в которых флуоресцентный микроскоп интегрирован в камеру FIB-SEM, и поэтому для получения флуоресцентных изображений не требуется перенос образца 29,30,31. С помощью таких интегрированных систем можно получить изображения интересующих позиций во время и после фрезерования FIB, чтобы проверить наличие целевого флуоресцентного сигнала без увеличения риска загрязнения конечных ламелей. Тем не менее, важно помнить об оптических параметрах используемых микроскопов, поскольку, например, объектив с низким значением NA ограничивает точность, с которой могут быть локализованы реперные шарики и целевые сигналы. Тем не менее, интегрированные настройки FLM также помогут лучше справляться с небольшими деформациями сеток и ламелей, поскольку стеки FLM могут постоянно обновляться и сравниваться с актуальными представлениями SEM и IB.
В качестве альтернативы флуоресцентной визуализации ламели между фрезерованием FIB и сбором данных ПЭМ для проверки правильности размещения и фрезерования ламелейможно использовать корреляцию после ПЭМ 5,6.
На всех этапах коррелятивного рабочего процесса, но особенно во время ПЭМ, рекомендуется создавать наложение данных проецируемой флуоресценции на изображения FIB-SEM/TEM. Такие классические CLEM-представления помогают более интуитивно понять, какая часть клеток содержится внутри ламелей. Это также служит полезной проверкой работоспособности для проверки точности корреляции.
3. Ограничения метода
Для 3D-коррелятивного подхода FIB требуются образцы, которые могут быть снабжены реперными шариками. Соответственно, этот метод в настоящее время ограничен погружно-замороженными сетками. Для замороженных (тканевых) образцов под высоким давлением (HPF) в настоящее время можно выполнять только корреляции 2D-2D. Потенциально решением этой проблемы могут быть внутренние реперные маркеры (например, органеллы, окрашенные липидные капли)32,33. Окончательная успешность корреляции зависит от многих факторов, включая качество образца, установку флуоресцентной микроскопии, толщину ламели и размер целевой структуры. Точность корреляции с использованием описанного подхода 3D-регистрации оценивается в диапазоне 200-300 нм на итоговом изображении IB, что примерно соответствует типичной толщине фрезерованных ФИБ ламелей7. Соответственно, в настоящее время будет трудно нацелиться на клеточные структуры гораздо меньшего размера. Кроме того, чрезмерное движение на участке измельчения (>300 нм) также снижает точность корреляции, и эта проблема потенциально может быть решена с помощью установок FLV, интегрированных в приборы FIB/SEM. Ламели, которые демонстрируют сильную деформацию или изгиб во время фрезерования, в любом случае должны быть исключены из последующего рабочего процесса.
В целом, криофлуоресцентная визуализация в настоящее время ограничена дифракционным критерием Аббе. При более рутинном применении (и коммерциализации) методов крио-ФЛМ со сверхвысоким разрешением может стать возможным более точное нацеливание на клеточные структуры, особенно при интеграции в FIB/SEM для работы «на лету».
4. Значимость метода
Особенно по сравнению с нецелевыми и посткорреляционными методами, 3D-коррелированный подход к фрезерованию FIB позволяет выбрать подходящие позиции до этапа TEM, требующего много времени и ресурсов. Таким образом, это позволяет более эффективно собирать данные и планировать проекты. Кроме того, коррелированные данные флуоресценции добавляют слой информации, которая может иметь решающее значение для интерпретации томограмм и для интеграции результатов крио-ЭТ в многомасштабные проекты, особенно при работе с неструктурированными белковыми сборками или теми, которые слишком малы для сопоставления шаблонов и усреднения субтомограмм.
5. Важность и потенциальные области применения в будущем
В сочетании с передовыми рабочими процессами, такими как криолифт из образцов HPF 34,35, cryo-FIB-SEM volume 36 и флуоресцентная визуализациясверхвысокого разрешения 26,37,38,39, 3D-таргетная подготовка ламелей открывает перспективу не только препарирования биологических процессов в изолированных клетках, но и обеспечения доступа к образцам тканей и пациентов для измельчения FIB и криоэлектронной томографии. Таким образом, он позволит препарировать патологические процессы в высоком разрешении и, таким образом, станет неотъемлемым строительным блоком для биопсии на наноуровне.
Раскрытие информации
Авторы заявляют об отсутствии конфликта интересов.
Благодарности
Мы благодарим Ингу Вольф (Inga Wolf) за поддержку ИТ-инфраструктуры, Флориана Бека (Florian Beck) за вычислительную поддержку и Оду Х. Шиетца (Oda H. Schiøtz) за критическое прочтение рукописи. Финансирование было частично предоставлено через стипендию Александра фон Гумбольдта для репатриантов Филиппа С. Эрдманна и долгосрочную стипендию EMBO ALTF 764-2014 для Флориана Уилфлинга. Анна Бибер получила стипендию Ph.D. фонда «Берингер Ингельхайм».
Материалы
| Name | Company | Catalog Number | Comments |
| Autogrids | Thermo Fisher Scientific / Homemade | 1036173 (no cutout), 1205101 (with cutout) | |
| C-rings | Thermo Fisher Scientific | 1036171 | |
| Corrsight with cryo module | Thermo Fisher Scientific | FLM Alternative 1 | |
| Dynabeads MyOne COOH | Thermo Fisher Scientific | 65011 | recommended 1 µm fiducial beads |
| EM Grids R1/4 SiO2 | Quantifoil | N1-S13nAu20-01 | |
| Falcon 4 camera w. post-column Selectris X energy filter | Thermo Fisher Scientific | Camera/Filter Alternative 1 | |
| FIB Aquilos 1 | Thermo Fisher Scientific | FIB Alternative 1 | |
| FIB Aquilos 2 | Thermo Fisher Scientific | FIB Alternative 2 | |
| FIB Quanta 3D FEG | Thermo Fisher Scientific | FIB Alternative 3 | |
| FIB Scios | Thermo Fisher Scientific | FIB Alternative 4 | |
| K2 summit camera w. post-column energy filter 968 Quantum K2 | Gatan | Camera/Filter Alternative 2 | |
| Leica TCS SP8 with cryo module | Thermo Fisher Scientific | FLM Alternative 2 | |
| Plasma Cleaner PDC-3XG | Harrick | ||
| Teflon Sheet (0.25 mm) | plastx24.de | 11645 | Cut to same dimensions as filter paper |
| TEM Titan Krios XFEG 300 kV Gi2 | Thermo Fisher Scientific | TEM Alternative 1 | |
| TEM Titan Krios XFEG 300 kV Gi4 | Thermo Fisher Scientific | TEM Alternative 2 | |
| THUNDER Imager EM Cryo CLEM | Thermo Fisher Scientific | FLM Alternative 3 | |
| Vitrobot Mark IV | Thermo Fisher Scientific | alternativevly, use manaual plunger | |
| Whatman filter paper | Sigma Aldrich | 10311807 | 55 mm diamater; needs to be cut to fit the Vitrobot |
Ссылки
- Beck, M., Baumeister, W. Cryo-Electron tomography: Can it reveal the molecular sociology of cells in atomic detail. Trends in Cell Biology. 26 (11), 825-837 (2016).
- Plitzko, M., Villa, E., Schaffer, M., Baumeister, W. Opening windows into the cell focused-ion-beam milling for cryo-electron tomography. Current Opinion in Structural Biology. 23, 771-777 (2013).
- Schaffer, M., et al. Optimized cryo-focused ion beam sample preparation aimed at in situ structural studies of membrane proteins. Journal of Structural Biology. 197 (2), 73-82 (2017).
- Tegunov, D., Xue, L., Dienemann, C., Cramer, P., Mahamid, J. Multi-particle cryo-EM refinement with M visualizes ribosome-antibiotic complex at 3.5 Å in cells. Nature Methods. 18 (2), 186-193 (2021).
- Klein, S., Wachsmuth-Melm, M., Winter, S. L., Kolovou, A., Chlanda, P. Cryo-correlative light and electron microscopy workflow for cryo-focused ion beam milled adherent cells. Methods in Cell Biology. 162, Elsevier Inc. 273-302 (2021).
- Klein, S., et al. Post-correlation on-lamella cryo-CLEM reveals the membrane architecture of lamellar bodies. Communications Biology. 4 (1), 1-12 (2021).
- Arnold, J., et al. Site-Specific Cryo-focused ion beam sample preparation guided by 3D correlative microscopy. Biophysical Journal. 110 (4), 860-869 (2016).
- Wilfling, F., et al. A selective autophagy pathway for phase-separated endocytic protein deposits. Molecular Cell. 80 (5), 764-778 (2020).
- Scientific volume imaging, Nyquist calculator. , Available from: https://svi.nl/NyquistCalculator (2021).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Huygens Professional version 19.04. , Scientific Volume Imaging. The Netherlands. Available from: http://svi.nl (2021).
- de Chaumont, F., et al. Icy: an open bioimage informatics platform for extended reproducible research. Nature Methods. 9 (7), 690-696 (2012).
- Arnold, J. 3DCT. , Available from: https://3dct.semper.space/ (2021).
- Klumpe, S., et al. A modular platform for streamlining automated cryo-FIB workflows. bioRxiv. , 444745(2021).
- Wolff, G., et al. Mind the gap: Micro-expansion joints drastically decrease the bending of FIB-milled cryo-lamellae. Journal of Structural Biology. 208 (3), 0-3 (2019).
- Buckley, G., et al. Automated cryo-lamella preparation for high-throughput in-situ structural biology. Journal of Structural Biology. 210 (2), 107488(2020).
- Tacke, S., et al. A streamlined workflow for automated cryo focused ion beam milling. bioRxiv. , 963033(2020).
- Zachs, T., et al. Fully automated, sequential focused ion beam milling for cryo-electron tomography. eLife. 9, e52286(2020).
- Fung, H. K. H. tools3dct. , Available from: https://github.com/hermankhfung/tools3dct (2021).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology. 152 (1), 36-51 (2005).
- Yang, J. E., Larson, M. R., Sibert, B. S., Shrum, S., Wright, E. R. CorRelator: Interactive software for real-time high precision cryo-correlative light and electron microscopy. Journal of Structural Biology. 213 (2), 107709(2021).
- Hagen, W. J. H., Wan, W., Briggs, J. A. G. Implementation of a cryo-electron tomography tilt-scheme optimized for high resolution subtomogram averaging. Journal of Structural Biology. 197 (2), 191-198 (2017).
- Bäuerlein, F. J. B., et al. In situ architecture and cellular interactions of polyQ inclusions. Cell. 171 (1), 179-187 (2017).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Shaner, N. C., Steinbach, P. A., Tsien, R. Y. A guide to choosing fluorescent proteins. Nature Methods. 2 (12), 905-909 (2005).
- Kaufmann, R., et al. Super-resolution microscopy using standard fluorescent proteins in intact cells under cryo-conditions. Nano Letters. 14 (7), 4171-4175 (2014).
- Booy, F. P., Pawley, J. B. Cryo-crinkling: what happens to carbon films on copper grids at low temperature. Ultramicroscopy. 48 (3), 273-280 (1993).
- Krull, A., Buchholz, T. O., Jug, F. Noise2void-Learning denoising from single noisy images. Proceedings of the IEEE Computer Society Conference on Computer Vision and Pattern Recognition. 2019, 2124-2132 (2019).
- Gorelick, S., et al. PIE-scope, integrated cryo-correlative light and FIB/SEM microscopy. eLife. 8, 1-15 (2019).
- Delmic METEOR. , Available from: https://www.delmic.com/en/products/cryo-solutions/meteor (2021).
- T. F. Scientific. iFLM. , Available from: https://assets.thermofisher.com/TFS-Assets/MSD/Datasheets/iflm-aquilos-datasheet-ds0366.pdf (2021).
- Mahamid, J., et al. Liquid-crystalline phase transitions in lipid droplets are related to cellular states and specific organelle association. Proceedings of the National Academy of Sciences of the United States of America. 116 (34), 16866-16871 (2019).
- Scher, N., Rechav, K., Paul-Gilloteaux, P., Avinoam, O. In situ fiducial markers for 3D correlative cryo-fluorescence and FIB-SEM imaging. iScience. 24 (7), 102714(2021).
- Mahamid, J., et al. A focused ion beam milling and lift-out approach for site-specific preparation of frozen-hydrated lamellas from multicellular organisms. Journal of Structural Biology. 192, 262-269 (2015).
- Schaffer, M., et al. A cryo-FIB lift-out technique enables molecular-resolution cryo-ET within native Caenorhabditis elegans tissue. Nature Methods. 16 (8), 757-762 (2019).
- Wu, G. -H., et al. Multi-scale 3D cryo-correlative microscopy for vitrified cells. Structure. 28 (11), 1231-1237 (2020).
- Liu, B., et al. Three-dimensional super-resolution protein localization correlated with vitrified cellular context. Scientific Reports. 5, 13017(2015).
- Weisenburger, S., Jing, B., Renn, A., Sandoghdar, V. Cryogenic localization of single molecules with angstrom precision. SPIE NanoScience + Engineering. 8815, (2013).
- Tuijtel, M. W., Koster, A. J., Jakobs, S., Faas, F. G. A., Sharp, T. H. Correlative cryo super-resolution light and electron microscopy on mammalian cells using fluorescent proteins. Scientific Reports. 9 (1), 1-11 (2019).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены