Method Article
インタクトな機能的な膜タンパク質へのヌクレオチド結合をリアルタイムで測定
要約
このプロトコルは、細胞環境において受容体に結合するアデニンヌクレオチドをリアルタイムで測定する方法を提示する。結合は、トリニトロフェニルヌクレオチド誘導体と非標準的な蛍光アミノ酸で標識されたタンパク質との間のフェルスター共鳴エネルギー移動(FRET)として測定されます。
要約
私たちは、細胞または膜環境におけるアデニンヌクレオチドの無傷の機能的な膜貫通受容体への結合を測定する方法を開発しました。この方法は、蛍光性非標準アミノ酸ANAPでタグ付けされたタンパク質の発現、およびANAPと蛍光(トリニトロフェニル)ヌクレオチド誘導体との間のFRETを組み合わせたものです。ANAPタグ付きKATP イオンチャネルへのヌクレオチド結合の例を、屋根のない原形質膜および切除した裏返しの膜パッチで電圧クランプ下で測定した例を紹介します。後者は、リガンド結合とチャネル電流の同時測定を可能にし、タンパク質機能を直接読み取ることができます。データの処理と分析は、潜在的な落とし穴やアーティファクトとともに、広範囲に議論されています。この方法は、KATP チャネルのリガンド依存性ゲーティングに関する豊富な機構的洞察を提供し、他のヌクレオチド調節タンパク質または適切な蛍光リガンドを同定できる任意の受容体の研究に容易に適応させることができる。
概要
タンパク質のいくつかの重要なクラスは、リガンド結合によって直接制御されています。これらは、可溶性酵素から、受容体チロシンキナーゼ、Gタンパク質共役受容体(GPCR)、およびイオンチャネルを含む膜埋め込みタンパク質にまで及びます。GPCRとチャネルは、現在のすべての創薬標的のそれぞれ~34%と~15%を占めています1,2。したがって、リガンド-受容体相互作用に関する機構的な洞察を提供する方法の開発には、かなりの生化学的および医学的関心があります。光親和性標識や放射性リガンド結合研究など、リガンド結合を測定するための従来の方法は、大量の部分的に精製されたタンパク質を必要とし、通常、非生理学的条件および時間スケールで実行されます。理想的な方法は、少量のタンパク質のみを必要とし、細胞または膜環境で発現されたインタクトなタンパク質に対して実行でき、リアルタイムで監視でき、タンパク質機能の直接読み取りと互換性があります。
フェルスター共鳴エネルギー移動(FRET)は、蛍光でタグ付けされた2つの分子3間の近接を検出する方法である。FRETは、励起されたドナー蛍光色素が非放射方式でアクセプタ分子(通常は別の蛍光色素)にエネルギーを伝達するときに発生します。エネルギー移動は、ドナー蛍光発光の消光およびアクセプター発光の増感(アクセプターがフルオロフォアの場合)をもたらす。伝達効率は、ドナーとアクセプターの間の距離の6乗 に依存する。さらに、FRETが発生するためには、ドナーとアクセプターが近接(通常は10nm未満)になければなりません。そのため、FRETを利用して、蛍光標識されたタンパク質受容体と蛍光リガンドとの間の直接結合を測定することができます。
いくつかの異なるタンパク質は、細胞内または細胞外のアデニンヌクレオチド(ATP、ADP、AMP、cAMP)に結合することによって調節または活性化される。多くのトランスポータータンパク質は、ATP結合カセットトランスポーターやNa+/K+ポンプ4,5のようなP型ATPアーゼなど、反応サイクルにATP加水分解を必要とします。ATP感受性K+(KATP)チャネル、嚢胞性線維症膜貫通コンダクタンス制御因子(CFTR)、および環状ヌクレオチド調節チャネルはすべて、細胞内アデニンヌクレオチドの結合によってゲートされるイオンチャネルであり、細胞代謝およびシグナル伝達の変化に非常に敏感です6,7,8.プリン作動性P2XおよびP2Y受容体は、神経伝達物質として、または組織損傷の結果として放出され得る細胞外ATPの変化に応答する9。膜タンパク質に結合するアデニンヌクレオチドをリアルタイムで測定するためのFRETベースのアッセイを開発しました。我々は以前、この方法をKATPチャネル10,11へのヌクレオチド結合を研究するために適用した。
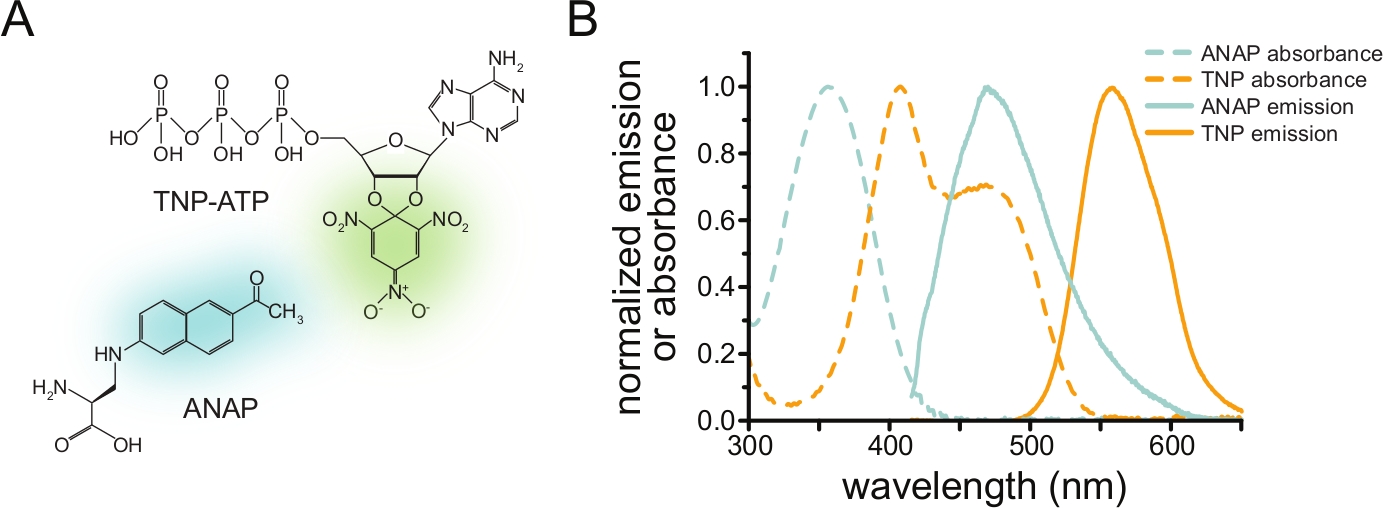
FRETを介したヌクレオチド結合を測定するには、まず目的のタンパク質を蛍光色素でタグ付けする必要があります。蛍光タグは、FRETが発生するリガンド結合部位に十分近くなるように、目的のタンパク質に部位特異的に挿入する必要があり、タグがタンパク質の全体的な構造と機能に影響を与えないように特別な注意が払われます。これを達成するために、我々はChatterjeeらによって開発された技術を採用し、アンバー終止コドン抑制を使用して蛍光非標準アミノ酸(l-3-(6-アセチルナフタレン-2-イルアミノ)-2-アミノプロピオン酸;ANAP)を目的のサイトで12。ANAP標識タンパク質と蛍光性トリニトロフェニル(TNP)ヌクレオチド誘導体との間のヌクレオチド結合をFRETとして測定します(図1A)。ANAPの発光スペクトルはTNPヌクレオチドの吸光度スペクトルと重なり、FRETが発生するために必要な条件です(図1B)。ここでは、2つの異なるタイプの結合実験の概要を説明します。第1に、ANAP標識KATPチャネルの細胞内側へのヌクレオチド結合は、ガラスカバースリップ10、11、13、14上に原形質膜の付着断片を残して超音波処理によって屋根を外した細胞において測定される。
第2の方法では、ANAP標識KATP チャネルへのヌクレオチド結合をメンブレンパッチ中で電圧クランプ下で測定し、イオン電流と蛍光の同時測定を可能にする。これら2つの実験的アプローチを組み合わせることにより、結合の変化をチャネル機能の変化と直接相関させることができる11。典型的な結果、潜在的な落とし穴、およびデータ分析について説明します。
プロトコル
1.カバースリップの準備
注意: これらの手順は、滅菌済みの組織培養フード内で行う必要があります。量は10皿の準備のために与えられます。
- オートクレーブ処理した30 mmホウケイ酸カバーガラス10枚を35 mm未処理の滅菌皿10個に個別に入れ、2 mLの滅菌蒸留水で1回すすぎます。
- 1 mL の 0.1% w/v ポリ-L-リジン溶液を滅菌蒸留水に希釈して、総容量を 10 mL にします (最終濃度 0.01% w/v)。よく混合し、各カバースリップに1 mLをピペットで入れ、室温で20分間インキュベートします。
- ポリ-L-リジンを吸引し、各カバースリップを少なくとも2 mLの滅菌蒸留水で2回洗浄します。完全に乾くまで、つまり少なくとも3時間放置します。
2. HEK-293T細胞の播種
注意: これらの手順は、組織培養フード内で行う必要があります。HEK-293T細胞は、その低電流バックグラウンドおよび培養中の増殖の容易さのために選択された。このプロトコルは、他の細胞型に適合させることができる。
- HEK-293T細胞の80〜90%コンフルエントT75フラスコを12 mLリン酸緩衝生理食塩水(PBS)で1回すすぎ、2 mLトリプシンと2〜5分間インキュベートするか、細胞が完全に剥離してほぼ完全に解離するまでインキュベートします。
- 10%ウシ胎児血清、100 U / mLペニシリン、および100 μg/mLストレプトマイシンを添加した10 mLのダルベッコ改変イーグル培地(DMEM)を加えて細胞を再懸濁します。フラスコの底を静かにピペットで固定して、残りの細胞の塊を分解します。
- コーティングされたカバースリップを含む所望の数の35 mmディッシュに2 mLの添加DMEMを加えます。各ディッシュに100 μLの再懸濁細胞を加えます。37°Cで一晩インキュベートします。
3. トランスフェクション
注意: これらの手順は、組織培養フード内で行う必要があります。10皿のトランスフェクションのための量が与えられます。部位特異的なANAPを組み込むためには、標識を意図した位置のDNAコドンをアンバー(TAG)終止コドンで置換しなければならない。このコンストラクトは、2つのプラスミド、pANAPおよびpeRF1-E55D12,15と同時トランスフェクトされています。pANAPは、ANAP特異的tRNA/tRNAシンテターゼペアのいくつかのコピーをコードする。ANAPの存在下で、このプラスミドのトランスフェクションは、アンバー終止コドンを認識するANAPで荷電したtRNAを生成します。peRF1-E55Dは、完全長のANAPタグ付きタンパク質の収量を増加させるドミナントネガティブリボソーム放出因子をコードしています。
- ANAPによる標識を目的としたコンストラクト用に、10 μg pANAP、10 μg peRF1-E55D、およびDNAを含む1.5 mLチューブを準備します。未添加のDMEMで最終容量を500 μLにします。
- 別のチューブで、DNA 1 μg ごとに脂質ベースのトランスフェクション試薬 ( 材料表参照) を 3 μL 調製し、未添加の DMEM で最終容量を 500 μL にします。
- DNA とトランスフェクション試薬の混合物を 1 本のチューブにまとめ、室温で 20 分間インキュベートします。
- 400 μLの1 mM ANAPストック(30 mM NaOH中のトリフルオロ酢酸塩)を20 mLの添加DMEMに加えて、最終濃度20 μM ANAPにします。プレーティングした細胞の古い培地を、ディッシュあたり2 mLのANAP含有培地と交換します。
- DNAトランスフェクションミックスの10%を各ディッシュにピペットで移します。実験前に33°Cで2〜4日間インキュベートします。33°Cでインキュベーションすると、細胞分裂が遅くなり、細胞あたりのタンパク質収量が増加します16。
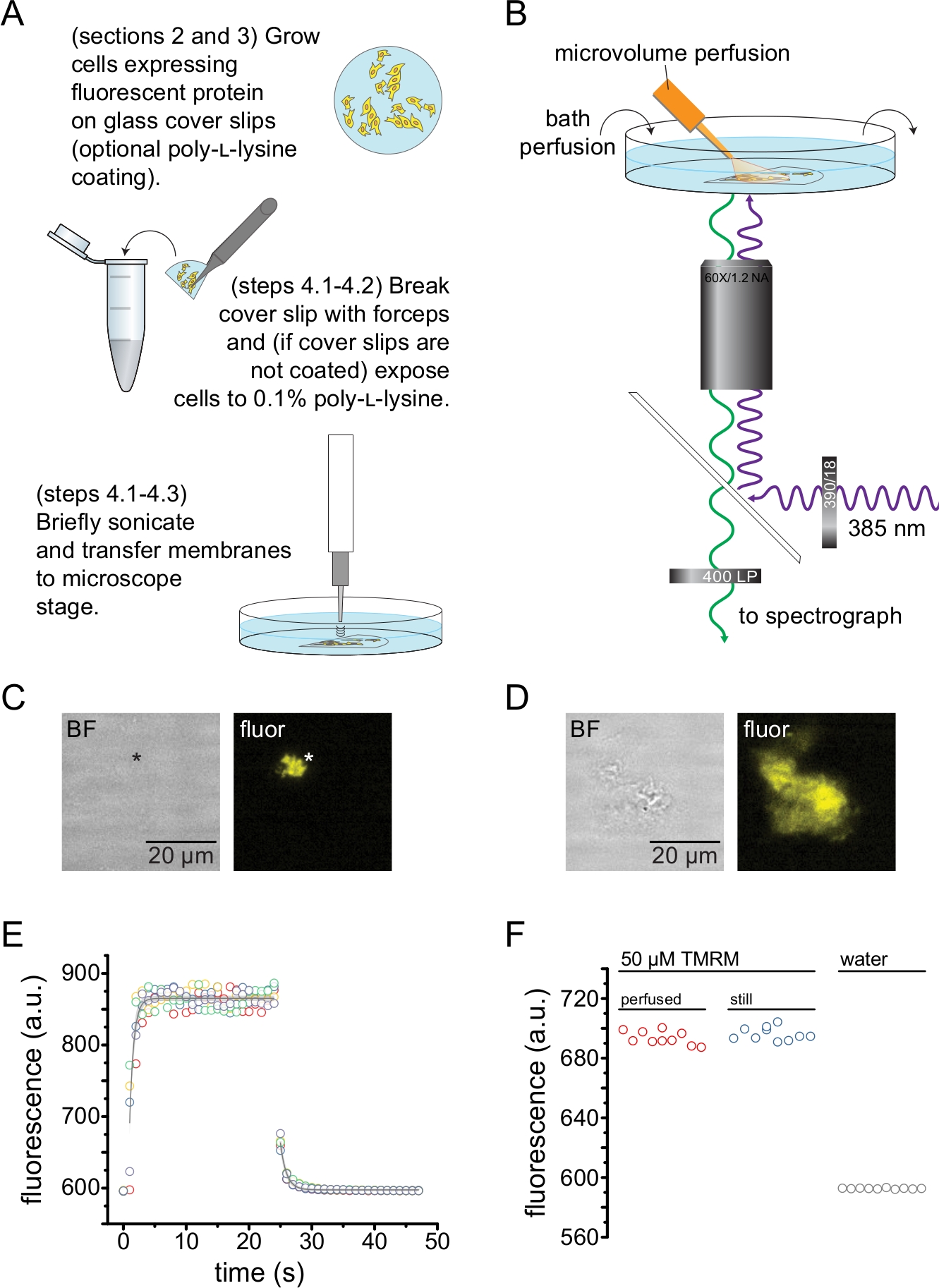
4. 屋根なし膜実験
- 一対の鉗子を使用して、トランスフェクトされた細胞を含むカバースリップをより小さな断片に分割します。
- 以下のいずれかの手順に従って、セルの屋根を外します。
- プレコートされたカバースリップを使用する場合は、フラグメントをPBSですすぎ、2 mLのPBSを含む35 mmディッシュの底に置きます。サンプルの3〜5 mm上に配置されたプローブ超音波処理器(50 W、20%-40%振幅、3 mmプローブ)を使用して簡単に超音波処理し、細胞の屋根を外し、付着した原形質膜断片を残します(図2A、C)。
注:ソニケーターのパワー、持続時間、およびサンプル上のプローブの高さはすべて、カバースリップを完全に露出させることなく、屋根のない膜の高収率を得るために変化させることができます。 - プレコートされたカバースリップを使用しない場合は、カバースリップフラグメントをPBSですすぎ、0.1%w / vポリ-L-リジンを含むチューブに~30秒間浸してから、簡単に超音波処理します(ステップ4.2.1のように)屋根を外し、屋根のない/部分的に屋根のない原形質膜断片を残します(図2A、C、D)。ポリ-L-リジンへの短時間の曝露は、カバーガラス13への付着を改善することが実証されている。
- プレコートされたカバースリップを使用する場合は、フラグメントをPBSですすぎ、2 mLのPBSを含む35 mmディッシュの底に置きます。サンプルの3〜5 mm上に配置されたプローブ超音波処理器(50 W、20%-40%振幅、3 mmプローブ)を使用して簡単に超音波処理し、細胞の屋根を外し、付着した原形質膜断片を残します(図2A、C)。
- 超音波処理された断片を2mLの浴溶液を含むカバーガラス底35mm皿に入れ、高NA、60倍水浸対物レンズを備えた倒立顕微鏡にマウントする。顕微鏡のカメラポートは、高感度CCDカメラと直列に分光器に接続されています。蠕動ポンプを使用して、バスチャンバー(0.5 – 1 mL / min)をバッファーで灌流します。緩衝液の組成は、研究中のタンパク質によって異なります。
注意: ユーザーが作動距離の長い対物レンズにアクセスできない場合、カバースリップの高さが余分なため、屋根のない膜の破片に焦点を合わせることができない場合があります。別の方法は、ポリ-L-リジンガラス底を有する皿に直接細胞を播種することである(例については 材料表 を参照)。これにより、2枚のガラスを通して焦点を合わせることに関連する画像の潜在的な収差も減少します。これらの収差は、取得されたスペクトルの形状に影響を与えません。 - チャネル蛍光を探すことにより、ANAP標識チャネルを発現する屋根のない膜断片を同定します(図2C、D)。
注:目的のタンパク質を含む屋根のない膜を識別するために、追加の蛍光標識(発光スペクトルがANAP発光スペクトルと区別できる場合)を使用することをお勧めします。図2のC、Dの実験は、C末端蛍光タンパク質タグを有するANAP標識チャネルで実施した。 - 分光器マスクを顕微鏡のカメラポートと分光器の間に部分的にかみ合わせます(~10%上げます)。マスクの影がカメラ画像に表示されます。顕微鏡ステージを調整して、屋根のないメンブレンを分光器マスクに合わせます。屋根のない膜の明視野と蛍光画像を取得します。これらは、分析する対象領域を選択するために使用されます。
- マイクロボリューム灌流システムの先端を屋根のない膜に近づけます。
注:バックグラウンド蛍光を低減するために、灌流システムの流出はホウケイ酸ガラス製のカスタムチップに置き換えられました。 - 蛍光スペクトルを画像化するには、390/18 nmバンドパス励起フィルターと416 nmエッジダイクロイックを介して385 nmのLEDでメンブレンを励起します。放射された光を400nmのロングパス発光フィルターに通します(図2B)。
- 分光器のマスクをかみ合わせ、放出された光が通過することを確認します。分光器のグレーティング(300溝/ mm)をかみ合わせます。格子を配置すると、分光器によって回折された光がCCDカメラのチップに投影され、スペクトル画像が生成されます(図3A)。これらの画像は、 y 次元の空間情報を保持します。 x 次元は波長に置き換えられます。
- 任意選択で、目的のタンパク質が蛍光タンパク質でタグ付けされている場合、適切なフィルターセットを使用して蛍光タンパク質のスペクトル画像を取得する。
- 実験の開始時に、ヌクレオチドを含まない緩衝液を灌流しながら、0.1〜10秒以上の曝露を1回以上行います。これらは、実験の残りの部分でデータを修正および正規化するために使用されます (以下のセクション 5 を参照)。
注:露光時間の選択は、達成される発現レベル、蛍光色素の輝度、および光学系によって異なります。露光時間は、信号を最大化し、観察された漂白速度を最小限に抑えるように選択する必要があります。4.10で与えられた暴露時間範囲は、平衡結合測定に適しているが、より遅い速度変化を測定するために有用であり得る10。短い露光時間を使用してより速い動態を追跡する能力は、ハードウェアではなく、タンパク質発現レベルと光退色によって制限されます。 - TNP-ATP(通常は浴溶液中で調製)の濃度の範囲を適用して、濃度-反応曲線を確立します。各溶液を少なくとも1分間灌流して定常状態に達することを確認し、各濃度を少なくとも1分間浴液で洗い流します。
注:灌流システムが急速に平衡に達し(図2E)、TNP-ATPの正しい局所濃度を達成できることを確認することが重要です(図2F)。 - 各濃度と各ウォッシュアウトの最後に(ステップ4.10で使用したのと同じ期間で)曝露します。
5. スペクトル解析
注: これらの手順は、GitHub にある分析コード "pcf.m" で使用するために記述されています。https://github.com/mpuljung/spectra-analysis10。追加のコードと代替コードは、https://github.com/smusher/KATP_paper_201911にあります。ここでは、ユーザーが独自のコードを作成したり、データを手動で分析したりできるように、ソフトウェアによって実行される操作について説明しました。
- コマンド ラインにプログラム名 ("pcf") を入力して、解析プログラムを起動します。
- ファイル/フォルダを開くダイアログボックスが開き、「ROI用のファイルを選択」というプロンプトが表示されたら、屋根のないメンブレンの明視野画像と蛍光画像に関連付けられたファイル名を選択します。出力ファイルの名前を入力するためのプロンプトがコマンドラインに表示されます。
- ファイル名を入力してEnterキーを押します。
- ソフトウェアが明視野画像と蛍光画像を表示したら、ソフトウェアプロンプトに従って、屋根のない膜片または切除されたパッチ(セクション6を参照)の位置に対応するスペクトル画像内の関心領域(ROI)を選択します。膜が取り付けられていないカバースリップまたはディッシュのセクションに対応する同じスペクトル画像(ROIと同じ波長範囲を表す)の背景領域を選択します(図3A)。ROIの上部をクリックしてEnterキーを押すように求められ、ROIの下部をクリックしてEnterキーを押してから、バックグラウンド領域に対してこのプロセスを繰り返します。
- ファイル/フォルダを開くダイアログボックスが開き、「FPスペクトルのファイルを選択」というプロンプトが表示されたら、蛍光タンパク質(FP)スペクトルに関連付けられたファイル名を選択します(オプションのステップ4.9)。FPスペクトラムが集録されていない場合は、別のスペクトラムファイルを選択してください。FPスペクトルは、タグ付きタンパク質とバックグラウンド蛍光を区別するための品質管理として機能します。
- 「解析用のファイルを選択」というプロンプトでファイル/フォルダを開くダイアログボックスが開いたら、漂白補正に必要なファイルを含め、ANAPスペクトルに対応するすべてのファイル(ステップ4.10から4.12まで)を選択します。
- 「漂白収集用のファイルを選択」というプロンプトでファイル/フォルダを開くダイアログボックスが開いたら、実験開始時にヌクレオチドフリー溶液で取得した初期スペクトル、または補正に使用するヌクレオチドフリー溶液で洗浄中に取得したスペクトルに対応するステップ5.6のファイルのサブセットを選択します(ステップ4.10から4.12まで)。
- スペクトルを生成するために各画像を線平均化する、すなわち、各波長におけるROIまたはバックグラウンド領域の y 次元内のすべてのピクセルの強度を平均化する。(図3B)。ROIから取得した平均スペクトルから得られた平均バックグラウンドスペクトルを差し引いて、非結合TNP-ATPからバックグラウンド蛍光および蛍光を除去します(図3C)。これらの手順は、ソフトウェアによって自動的に実行されます。
- 差し引かれたスペクトルのANAPピークを中心とした5 nmウィンドウの強度を平均することにより、各曝露のANAP強度を決定します(通常~470 nmですが、ANAP残基の局所微小環境に応じて異なる場合があります)。
注: 図3D は、ANAPタグ付きチャネルを発現する屋根のない膜断片の連続した10秒間の曝露から得られた6つのスペクトルを示しています。挿入図は、各スペクトルのピークの平均強度を示す。ソフトウェアは、取得した最初のスペクトルのピーク波長を自動的に検出し、この値を全体を通して使用します。強度はソフトウェアによって自動的に計算されます。 - 特定のばく露のANAP強度(F)を、ステップ4.10で取得した時系列の最初のばく露のANAP強度(Fmax)で割ることにより、各実験のANAP強度を正規化します。この場合も、ソフトウェアはこれらの計算を自動的に実行します。
- データを取得するには、以下の手順を実行してください。
- ANAP光退色を補正するには、まず、単一の指数関数的減衰(F / Fmax)= A*exp(-t / τ)+(1-A)、ここでtは累積露光時間、τは時定数、Aは振幅)を、TNP-ATPアプリケーション間の中間洗浄ステップまたはTNP-ATPでの洗浄前に行われた複数の初期露光のいずれかに適合させます(図3D、 挿入図)。
注意: ソフトウェアはこのフィットを表示し、それを受け入れるか拒否するかを確認するプロンプトを表示します。フィットが拒否された場合、漂白補正用のファイルを選択する別の機会が提供されます。 - 正規化された(ステップ5.10の)ANAPスペクトルを、各時点でのステップ5.11.1からの指数適合の予測値で割ります(図3E)。
注: 示されている例では、50 秒で観測された正規化されたピーク蛍光は 0.65 で、指数適合からの予測蛍光は 0.64 です。漂白を補正するには、観測値(0.65、図3E挿入図、空の円)を予測値(0.64、図3E挿入図、破線)で割って、補正値(~1、図3E挿入図、色付きの円)を生成します。漂白補正が適切であれば、ヌクレオチドの非存在下で得られたすべての曝露からのANAPの強度はほぼ等しくなければならない(図3E)。これらの計算は、ソフトウェアによって自動的に実行されます。 - 出力を、データをプロットした画像と、生のスペクトル、減算されたスペクトル、光退色のために補正されたスペクトル、および各ファイルのピークデータを含むタブ付きスプレッドシートとして取得して、さらに分析を行うことができます。
- ANAP光退色を補正するには、まず、単一の指数関数的減衰(F / Fmax)= A*exp(-t / τ)+(1-A)、ここでtは累積露光時間、τは時定数、Aは振幅)を、TNP-ATPアプリケーション間の中間洗浄ステップまたはTNP-ATPでの洗浄前に行われた複数の初期露光のいずれかに適合させます(図3D、 挿入図)。
6. パッチクランプ蛍光測定実験
- ピペット溶液で満たされたときに、厚肉のホウケイ酸ガラスキャピラリーからパッチピペットを1.5MΩから2.5MΩの抵抗まで引き出します。ピペット溶液の組成は、研究中のタンパク質によって異なります。
- トランスフェクトした細胞を含むカバースリップを、2 mLの浴液を含むカバーガラス底35 mmディッシュに移し、高NA、60倍水浸対物レンズを備えた倒立顕微鏡にマウントします。蠕動ポンプを使用して、浴室(0.5 – 1 mL / min)をバス溶液で灌流します。ピペット溶液に関しては、浴溶液は研究中のタンパク質によって異なります。
- 細胞膜での蛍光を探すことにより、ANAP標識チャネルを発現する細胞を同定します。
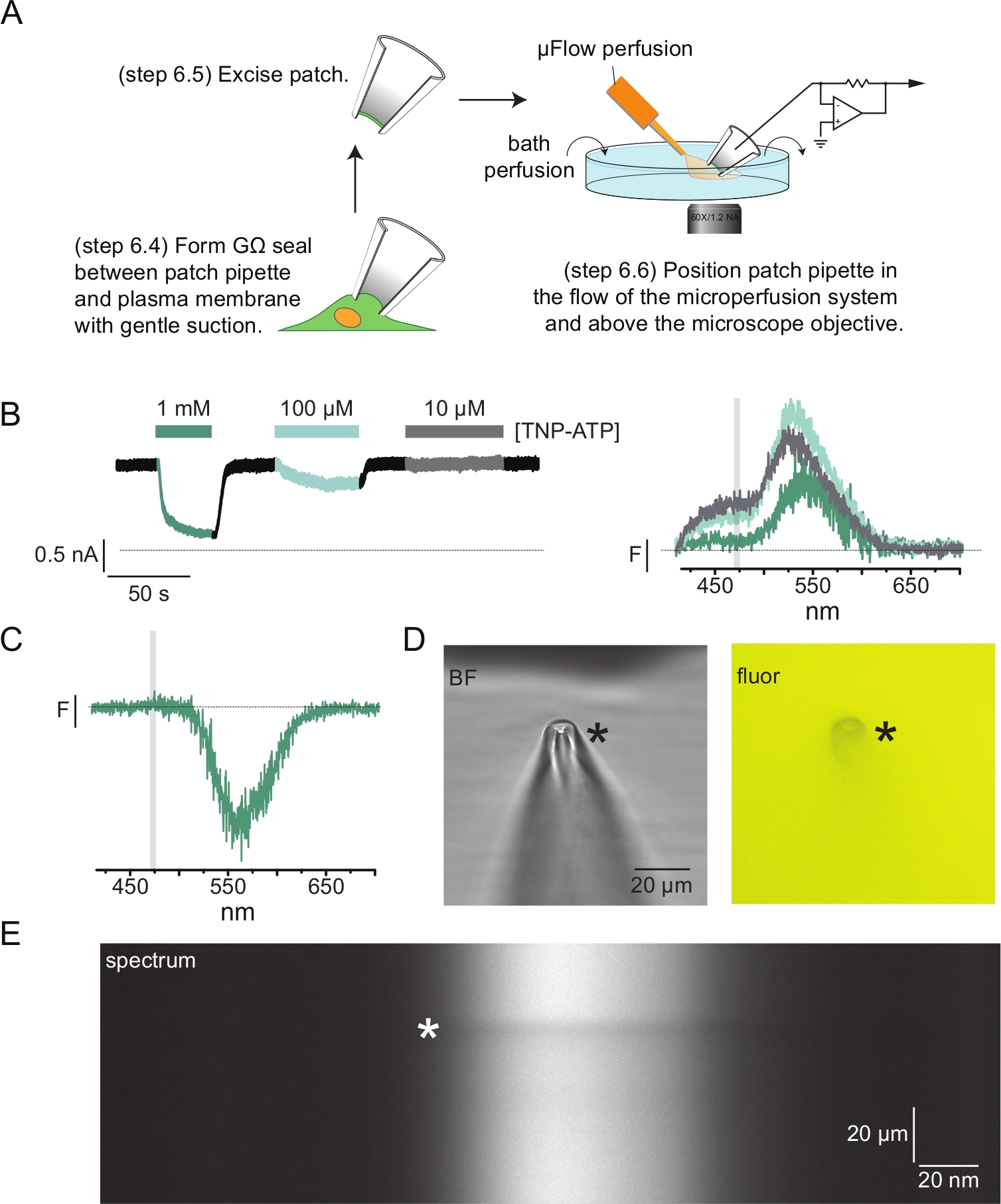
- パッチピペットにピペット溶液を入れます。ピペットに穏やかな陽圧を加え、バスチャンバーに入れます。ピペットを細胞の膜に押し付け、穏やかな吸引を加えてGΩシールを実現します(図4A)。
- ピペットホルダーを細胞から素早く離してパッチを切除します(図4A)。
注:この方法でパッチを切除すると、タンパク質の細胞質ドメインが灌流系にさらされた裏返しのパッチが形成されます。研究中のヌクレオチド結合部位の位置が細胞質でない場合は、PCF実験を行うためにアウトサイドアウトパッチまたは全細胞記録を使用する必要があります。 - パッチピペットの先端を灌流システムの先端に近づけ、パッチが分光器マスクのスリット内にあることを確認します(図4A)。
- ステップ4.10-4.12のようにTNP-ATPと画像スペクトルを適用し、同時にヌクレオチド適用に対するイオン電流応答を記録します。
注意: ピペットガラスは、取得した画像に空間収差や反射をもたらす可能性があります。ただし、これらの収差は取得したスペクトルの形状に影響を与えず、反射励起光は分光器またはロングパス発光フィルターのいずれかを使用して蛍光から簡単に分離できます。 - スペクトルを分析します。切り出したパッチから画像化されたスペクトルは、パッチピペットのガラスからTNP-ATPが排除されるため、未結合のTNP-ATP蛍光の過剰減算を示す可能性があります(図4C-E)。この過減算はANAP発光スペクトルに影響しないため、無視してかまいません。
注:切除パッチの蛍光シグナルは屋根のないメンブレンよりも低くなるため、ANAPを急速に漂白することなく十分に高い信号対雑音比が得られる露光時間を使用することが重要です。
結果
図2は、超音波処理によって得られた屋根のない膜断片中の蛍光タンパク質へのヌクレオチド結合を測定するための基本的な実験セットアップを示しています(図2A、B)。屋根のない膜を得るためには、ポリ-L-リジンでコーティングされたカバースリップ上で細胞を直接培養するか、未処理のガラス上で細胞を培養し、屋根を開ける前にポリ-L-リジン(水中0.1%)に短時間さらすかの2つの異なるアプローチを使用しました。図2Cは、オレンジ蛍光タンパク質(OFP)でタグ付けされたKATPチャネルを発現するHEK−293T細胞からの典型的な屋根のない膜断片を描写する。屋根のない膜は明視野画像では事実上見えず、タグ付き膜タンパク質の蛍光またはオクタデシルローダミンB13などの膜色素による対比染色によって同定されました。屋根のない膜に加えて、HEK-293T細胞の超音波処理も部分的に屋根のない細胞断片を生成しました(図2D)10,17。これらの破片は明視野で見えた。これは、カバーガラスへの接着が不十分な波立たせられた原形質膜の結果である可能性があります。あるいは、これらの断片は、細胞内オルガネラ由来の小胞および膜を含み得る。そのため、細胞内膜に関連する標識標的タンパク質は翻訳後プロセシングおよびアセンブリの中間段階を反映している可能性があるため、「真の」屋根のない膜からのみ画像を取得することが好ましい。ポリ-L-リジンコーティングガラス上で細胞を培養することは、超音波処理時に「真の」屋根のない膜のより高い収率をもたらしたため、推奨されます。
マイクロボリューム灌流システムを蛍光ヌクレオチドに適用して、典型的な実験で必要な量を最小限に抑えました(図2B)。提供されたポリイミドコーティングされたガラスチップは、灌流セットアップで手で引っ張られたホウケイ酸ガラスチップに置き換えられ、蛍光バックグラウンドが減少しました。イメージングされる屋根のない膜の周囲のヌクレオチド蓄積を最小限に抑えるために、浴室全体を緩衝液でゆっくりと灌流しました。そのため、マイクロボリューム灌流システムからの溶液変化率を測定し、関心領域で意図したリガンド濃度を達成できたこと、つまり、灌流システムからのリガンドが屋根のない膜に到達する前に入浴媒体に直接希釈されていないことを確認したいと考えました。これらの可能性を制御するために、水で灌流されたカバーガラス底皿の表面を対象としたマイクロボリューム灌流システムからのテトラメチルローダミン-5-マレイミド(TMRM)の50μM溶液のウォッシュインおよびウォッシュアウトを測定しました(図2E)。溶液交換速度論は再現性があり、ウォッシュインとウォッシュアウトの両方で時定数が1秒未満の単一の指数関数的減衰によってよく説明されました。このような溶液交換時間は、現在のセットアップでリガンドの結合と非結合の速度論を測定する能力を制限します。カバースリップの表面で目的のリガンド濃度を達成できたことを確認するために、マイクロボリューム灌流システムによってカバースリップに送達された50 μM TMRMの蛍光強度を、スチルバス中の50 μM TMRMと比較しました(図2F)。強度に差は認められず、浴を灌流した場合でも、マイクロボリューム灌流システムによるカバースリップ表面の適切なリガンド濃度を達成できることが確認されました。
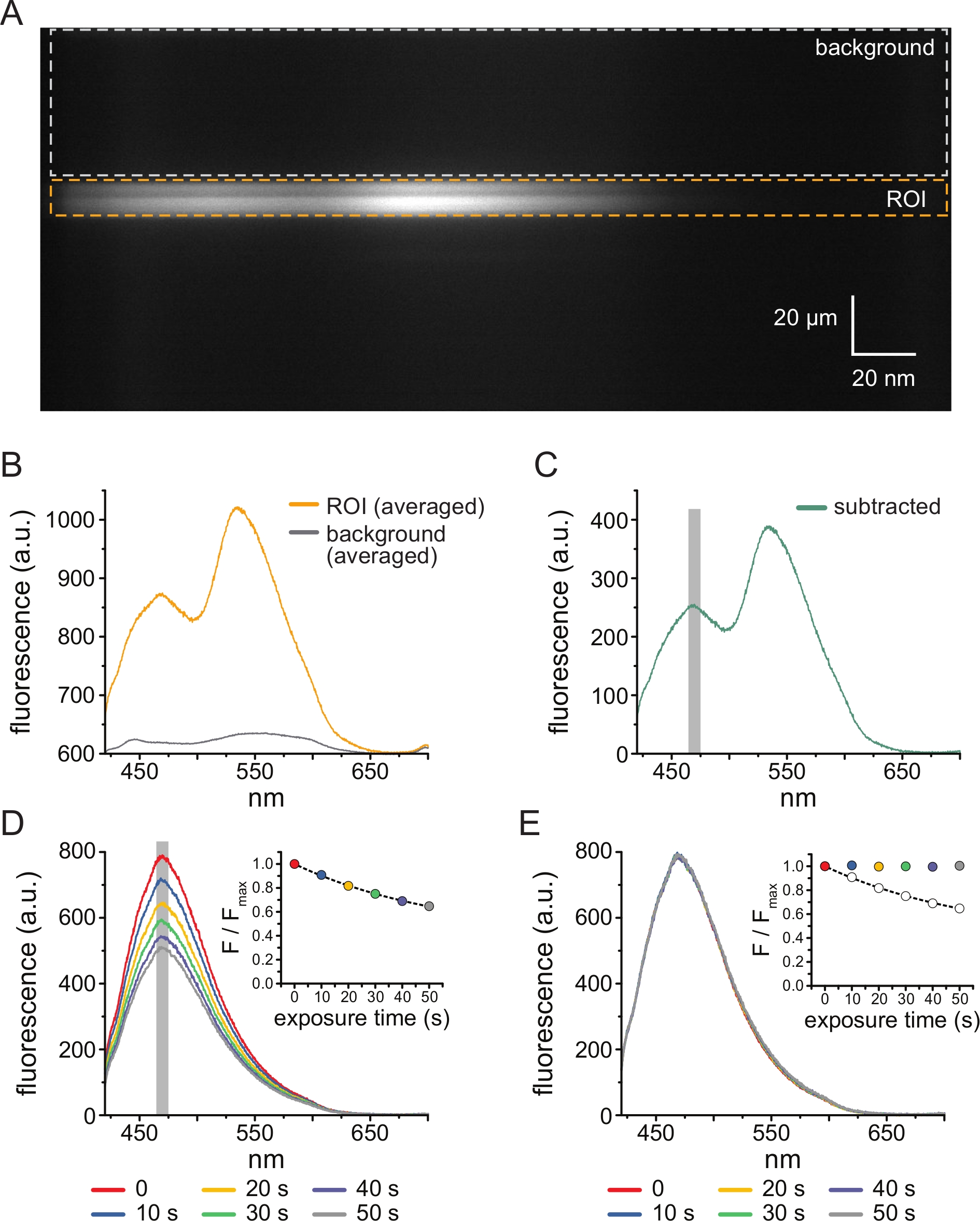
図3A は、5μM TNP-ATPに曝露したHEK-239T細胞由来の屋根のない膜中のANAPタグ付きKATP チャネルから得られたスペクトル画像を示す。このような画像を得るために、屋根のない膜から放出された光を、CCDカメラと直列に分光計を通して導いた。放出された蛍光は格子から回折され、カメラチップに投影され、スペクトルが生成されました。結果の画像は y 次元の空間情報を保持しますが、 x 次元は波長に置き換えられました。屋根のない膜に対応する関心領域(ROI)はオレンジ色で囲まれています。画像には、ANAPとTNP-ATPのピーク発光に対応する2つの高輝度領域が明らかです。これは、 図3Bに示す波長ごとの平均(ROI全体にわたる)スペクトルで最もよく理解されました。ピーク~470 nmは、KATPに取り込まれたANAPに対応する。ピーク~535 nmはTNP-ATPに対応する。溶液中のTNP−ATPのバックグラウンド蛍光および直接励起を補正するために、バックグラウンド領域(図3A、灰色)を各画像から選択した。平均化されたバックグラウンドスペクトルを 図3Bに示します。最終的なスペクトルは、平均化されたROIスペクトルから平均化されたバックグラウンドスペクトルを差し引くことによって得られました(図3C)。
ANAPはアーティファクトを光退色する傾向があります。 図3D は、多重露光後のピークANAP蛍光の減少を示す。TNP-ATP非存在下での数回の曝露(またはTNP-ATP濃度間の洗浄による)からのピーク蛍光を単一指数関数的減衰に適合させ、これを光退色アーチファクトの補正に使用しました(図3E)。低ヌクレオチド濃度から高濃度および高ヌクレオチド濃度から低濃度の両方から濃度反応実験を行うことをお勧めします。漂白補正によって追加のアーティファクトが導入されない場合、結果は同等になるはずです11。
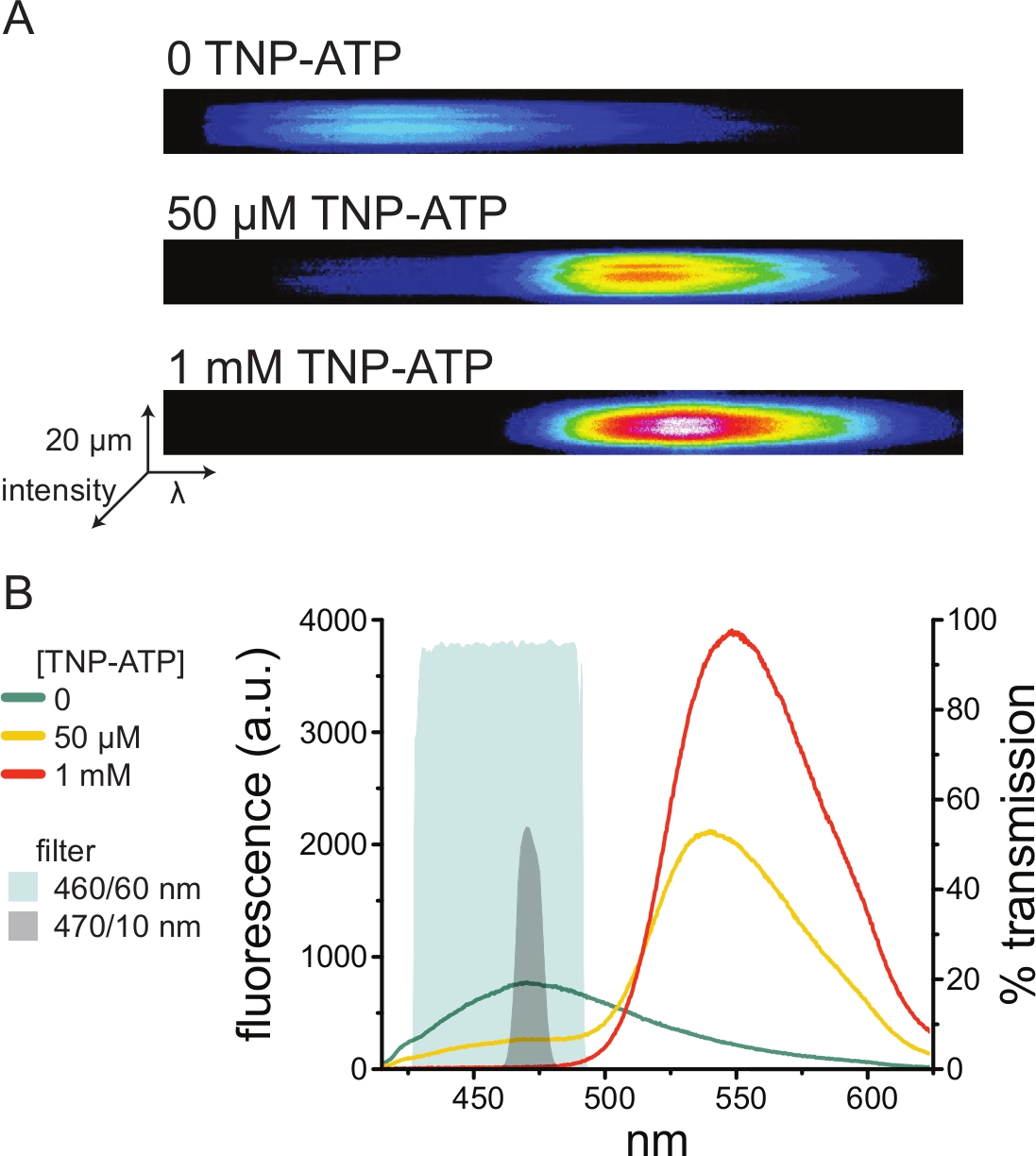
図5Aは、TNP−ATPの非存在下および存在下でANAPタグ付きKATPチャネルを発現する細胞から得られた屋根のない膜からの代表的なスペクトル画像を示す。補正されたスペクトルを図5Bに示します。発光スペクトルを観察すると、ドナーとアクセプターの蛍光発光の間に明確な分離がありました。トランスフェクションされていないHEK-293T細胞由来のナイーブ原形質膜へのTNP-ATPの非特異的結合が観察されたため、FRETをドナー(ANAP)蛍光の減少として定量することが推奨されます10,11。このピークは標識受容体に特異的であった。
それらの受容体の立体構造変化を誘導するリガンドについては、単独での結合研究は、リガンド結合プロセスに関する直接的で機構的に意味のある情報を提供しない18。リガンド結合の濃度応答関係は、固有の結合親和性だけでなく、リガンド結合によって誘発される立体構造の変化、およびリガンドがない場合に立体構造を変化させる受容体の固有の傾向にも依存します。リガンド-受容体相互作用を強調するプロセスをよりよく理解するために、結合測定をタンパク質機能の読み取りを提供する実験と組み合わせることができます。この目的のために、イオンチャネルは、電圧クランプを使用して1分子レベルまでのサブミリ秒の時間分解能で電流を測定できるため、理想的なモデルシステムです。歴史的に、対の電流および蛍光測定は、電位依存性および配位子依存性イオンチャネルの開閉(ゲーティング)に関する重要な洞察を提供してきました19、20、21。様々な環状ヌクレオチド調節チャネルへのイオン電流および蛍光環状ヌクレオチド結合を同時に測定する実験が行われている22、23、24。これらの研究では、結合時の量子収率を増加させるリガンドが採用されました。パッチ近傍の溶液の体積における非結合リガンドからの蛍光は、共焦点顕微鏡を用いてパッチを画像化することによって差し引くことができる22、23。我々の研究では、結合はANAP蛍光の減少を用いて測定された。このシグナルはチャネルに特異的であり、ANAPとTNP-ATP間のFRETは距離に強く依存するため(~43 Åで最大半分)、非特異的に結合したヌクレオチドと結合していないヌクレオチドによるシグナルの汚染は回避されました。
図4A は、典型的なパッチクランプ蛍光測定(PCF)実験を示しています。生理食塩水充填ホウケイ酸ガラスピペット(電圧クランプ増幅器に接続)とANAPタグ付きKATPを発現する細胞との間に高抵抗(GΩ)シールを形成した。シール形成後、ピペットを細胞から引き離し、細胞内ヌクレオチド結合部位へのアクセスを可能にした。次に、ピペットを分光器マスクのスリットを中心とした顕微鏡対物レンズ上に配置し、マイクロボリューム灌流システム(ホウケイ酸ガラスチップで修正)の流出をピペットに近づけました(図4D)。電圧を制御し、パッチ内のチャネルから電流を測定しました。ANAPタグ付きKATP チャネルからの代表的な電流とスペクトルを 図4Bに示し、スペクトルを電流に一致させるために色分けしています。発光スペクトルは、屋根のない膜と同様に、バックグラウンドと漂白について補正されました。

図1:ANAPとTNP-ATPは適切なFRETペアを作ります。 (A)ANAPおよびTNP-ATPの構造。蛍光部分が強調表示されています。(B)ANAPおよびTNP-ATPの吸光度および蛍光発光スペクトル。FRETには、ANAP発光とTNP-ATP吸光度のオーバーラップが必要です。Puljung et al. (クリエイティブ・コモンズ表示ライセンス, https://creativecommons.org/licenses/by/4.0/)10 より転載。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図2:屋根のない原形質膜におけるヌクレオチド結合の測定。 (a)蛍光膜タンパク質を発現する接着細胞からの屋根のない原形質膜の調製のための概略図。ポリ-L-リジンコーティングまたは未処理のカバースリップで増殖した細胞については、説明書が記載されています。(B)屋根のない膜のヌクレオチド結合を測定するための実験セットアップ。(C)オレンジ蛍光タンパク質(OFP)タグ付きKATP チャネルを発現する細胞に由来する完全に屋根のない原形質膜の明視野および蛍光画像。アスタリスクは膜の位置を示し、明視野画像ではほとんど見えません。OFPは、531/40nmのバンドパスフィルタと562nmのエッジダイクロイックを介して広い565nmのLEDで励起され、放出された光は593/40nmのバンドパスフィルタを通して収集されました。(D)オレンジ蛍光タンパク質(OFP)タグ付きKATP チャネルを発現する細胞に由来する部分的に屋根のない膜断片の明視野および蛍光画像。(E)Bで説明したセットアップを使用して取得した溶液交換時間コース。 5つの技術的な反復が示されています。マイクロボリューム灌流システムに50 μMテトラメチルローダミン-5-マレイミド(TMRM)をロードしました。浴を~0.5mL/minの速度で水で灌流した。ウォッシュオン(蛍光の増加)およびウォッシュアウト(蛍光の減少)タイムコースからのデータは、 F = A*exp(-x/τ)+ y0の形式の単一の指数関数的減衰に適合した。ウォッシュインの時定数(τ)は~0.6秒であった。TMRMは、540/25 nmバンドパスフィルタと565 nmエッジダイクロイックを介して広い565 nmのLEDで励起され、放出された光は605/55 nmバンドパスフィルタを介して収集されました。(F)Bと同様にマイクロボリューム灌流システムを用いて塗布したTMRMの50 μM溶液と50 μM TMRMを含むスチルバスの蛍光強度の比較。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図3:バックグラウンド減算と漂白補正。 (A)ANAP標識KATPチャネルを発現する細胞からの屋根のない原形質膜のスペクトル画像(y次元の空間情報、x次元の波長)。5μMのTNP-ATPを、図2Bに示すセットアップを使用して適用しました。オレンジ色のボックスは、屋根のない膜に対応する関心領域(ROI)を示します。灰色のボックスは、スペクトルの補正に使用される背景領域を示します。(B)AにおけるROI及びバックグラウンド領域の波長ごとの平均から得られる発光スペクトル。 (C)Bにおける平均化されたROIスペクトルから平均化されたバックグラウンドスペクトルを差し引いたスペクトル。平均強度を決定するために使用されたANAPピークの周りの5nmのウィンドウは、灰色の影付きの領域として示されています。(D)ANAP標識KATPチャネルを発現する細胞からの屋根のない原形質膜の6回の連続した10秒曝露から得られたスペクトル。光退色に起因する蛍光の減少に注意してください。挿入図は、F/Fmax = A*exp(-t/τ) + (1-A) という形式の単一の指数関数的減衰による正規化されたピーク蛍光適合を示しています。挿入図のシンボルは、スペクトルに一致するように色分けされています。(E)Dと同じスペクトルを光退色のために補正した。挿入図は、Dからの正規化されたピーク蛍光を白丸として示し、補正されたピーク蛍光は塗りつぶされた円を使用して示されます。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図4:パッチクランプ蛍光法(PCF)を用いたヌクレオチド結合とチャネル電流の同時測定。 (A)ヌクレオチド結合およびイオン電流を測定するための実験セットアップを示す概略図。(B)ANAP標識KATP チャネルを発現する細胞から切り出したメンブレンパッチから取得した電流の例(左)とスペクトル(右)。電流は-60mVの保持電位で記録され、20kHzでデジタル化され、5kHzでフィルタリングされました。灰色の影付きの領域は、ANAP強度が定量化された波長範囲に対応します。Usher et al. (クリエイティブ・コモンズ表示ライセンス, https://creativecommons.org/licenses/by/4.0/)11 より転載。(C)1mM TNP-ATPに曝露されたANAP標識KATP チャネルを発現する細胞から切り出したメンブレンパッチから取得したスペクトル。なお、TNP-ATP蛍光が観測される波長範囲に対応する負のピーク。灰色の影付きの領域は、B.Usherら(クリエイティブコモンズ表示ライセンス、https://creativecommons.org/licenses/by/4.0/)11の下で公開されたUsherらから適応されたように、ANAP蛍光を定量化するために使用される波長範囲を示します。(D)1 mM TNP-ATPに曝露したパッチピペットの明視野および蛍光画像。アスタリスクはピペットの先端を示します。(E)1 mM TNP-ATP中の同じパッチピペットのスペクトル画像。アスタリスクはピペットの位置を示します。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図5:ANAP標識K ATPチャネルへのTNP-ATP 結合。(A)TNP-ATPの非存在下または50 μMまたは1 mMのTNP-ATPの存在下で、ANAP標識KATP チャネルを発現する細胞からの屋根のない原形質膜のスペクトル画像。強度はヒートマップとして表示されます。(B)TNP-ATPによるANAP蛍光の消光を示すAの画像からの波長ごとの平均スペクトル。影付きの領域は、分光器が利用できない場合にANAPクエンチングを測定するために使用できる2つの異なるバンドパスフィルターを表しています。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

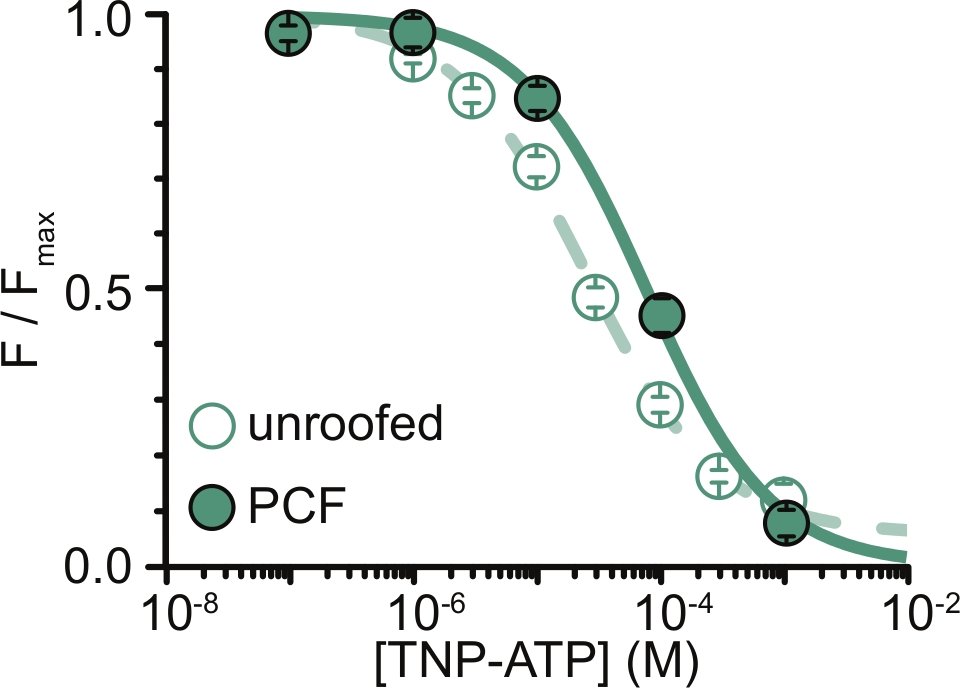
図6:屋根のない膜およびPCFにおけるTNP-ATPによるANAP標識KATPチャネルの消光。Usher et al. (クリエイティブ・コモンズ 表示 ライセンス, https://creativecommons.org/licenses/by/4.0/)11 のデータのオーバーレイ。データはヒル方程式に適合した:F / F max = E max +(1 – E max)/(1 + 10(EC50 –[TNP-ATP])* h)。Fは測定された蛍光であり、Fmaxはヌクレオチドの非存在下での最大蛍光であり、Emaxは飽和ヌクレオチド濃度での最大消光であり、そしてhはヒルスロープである。EC50(クエンチが最大の半分となるヌクレオチド濃度)および[TNP-ATP]は対数値である。屋根なし膜:EC50 = -4.59(25.7 μM)、h = 0.82、Emax = 0.93。PCF: EC50 = -4.11 (77.6 μM), h = 0.87, Emax = 1.00.この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
ディスカッション
我々は、インタクトな膜タンパク質へのアデニンヌクレオチド結合をリアルタイムで測定する方法を開発しました。私たちの方法は、アンバー終止コドン抑制12、セルアンルーフ14、電圧クランプ蛍光測定/ PCF 19,20,21,22,23,24,25を使用したANAPによるタンパク質の標識など、他のいくつかの確立された技術に基づいています.これらのアプローチの合成により、高い空間分解能および時間分解能でヌクレオチド結合の測定が可能になります。実際、以前の研究では、このアプローチを使用して、同じタンパク質複合体上の異なる結合部位を区別することができました10,11。重要なことに、この技術は、タンパク質機能を維持する条件下で、細胞環境中の少量のタンパク質に直接適用することができます。当社の結合法をイオンチャネル電流の直接的な電気生理学的読み出しと組み合わせて使用 することで、チャネルゲーティング11の分子基盤に関する豊富な洞察を得ることができます。
分光計は非標準の実験装置であるため、バンドパスフィルターを使用してANAP強度を相対的に絶縁して監視することもできます。 図5B は、このような2つのフィルタのスペクトル特性を示しています。470/10 nmバンドパスフィルターは、TNP-ATPからの蛍光シグナルを効果的に遮蔽し、ピークANAP蛍光とよく重なります。ただし、このフィルターのピーク透過率は約50%に過ぎないため、薄暗いメンブレン(または電圧クランプ下の切除メンブレンパッチ)から良好な信号を得ることが困難になる可能性があります。別のオプションは、460/60nmバンドパスフィルタです。470/10 nmフィルターと比較して、460/60 nmフィルターとTNP-ATP発光ピークの足の間にはわずかに多くのオーバーラップがあります。しかし、460/60nmのバンドパスは、ANAPピークの広い範囲で90〜95%の透過率を有し、蛍光発光シグナルを後押しすることが期待される。
ANAPは、環境に敏感な蛍光色素12、26、27です。ピーク発光および量子収率は、目的のタンパク質上の取り込み部位によって異なり、タンパク質の立体構造が変化するにつれて変化する可能性があります。このような変化は、発光スペクトルからすぐに明らかになりますが、フィルターを使用してANAP強度を測定する場合ほど明白ではありません。いずれにしても、ヌクレオチド結合後のANAP周辺の局所環境の変化のために蛍光シグナルが変化しないことを実証するには、適切なコントロールが必要です。非標識ヌクレオチドを用いた対照実験は、ANAP強度の変化がANAPとTNPヌクレオチドの間のFRETの結果であることを確認するのに役立ちます。TNPヌクレオチドは、トランスフェクトされていない細胞由来の膜(原形質膜または天然の膜タンパク質)に非特異的に結合することができます10。このシグナルは標識チャネルに特異的であるため、ドナー蛍光の減少として結合を定量化します。ただし、ドナー蛍光の変化が実際に標識受容体11への直接結合の結果であることを確認するために、各アゴニスト/受容体ペアに対して追加の対照実験(既知の場合はヌクレオチド結合部位を変異させるなど)を行うことをお勧めします。最後に、ANAPラベルに加えて蛍光タンパク質タグを含むコンストラクトを使用することをお勧めします。これは、標識された受容体蛍光をバックグラウンド/自家蛍光と区別するのに役立ちます。バックグラウンド蛍光は、発光スペクトル10のピークおよび形状によってANAPと区別することができるが、そのような決定は、フィルタセットのみが使用される場合、非常に困難であり得る。さらに、蛍光受容体を発現する細胞および屋根のない膜は、ANAPを励起することなく、過度の光退色のリスクなしに蛍光タンパク質タグを使用して識別できます。
PCF記録の多くでは、高いTNP-ATP濃度でスペクトルに強い負のピークが観察されました(図4C)。この負のピークは、バックグラウンド減算プロトコルのアーティファクトです。 図4D は、1 mM TNP-ATPに曝露したパッチピペットの明視野および蛍光画像を示す。ピペット壁の体積からTNP-ATPが排除されたためにピペットチップに影が見え、焦点面内で最も顕著になります。 図4E のスペクトル画像は、この影に対応する暗いバンドを示しています。この暗いバンドの上または下の領域をバックグラウンド減算に使用すると、負のピークが生成されます。重要なことに、このピークはTNP-ATP発光に対応する波長範囲で発生し、ANAPクエンチングの測定に影響を与えませんでした。
私たちの実験の主な制限は、蛍光を測定するためにANAPタグ付きコンストラクトの適切な原形質膜発現を得ることでした。パッチが一度に1つしか取得できないPCFとは異なり、サイズが大きく、屋根のないメンブレンをディッシュ全体をすばやくスキャンできるため、PCFよりも屋根のないメンブレンから高品質のスペクトルを取得するのは一般的に簡単でした。我々の実験では、屋根のない膜とPCF実験のデータは類似していましたが、同等ではありませんでした(図6)11。しかし、パッチピペット内のタンパク質は、屋根のない膜内のタンパク質とは異なる機能状態にある可能性があるため、これが普遍的な観察であるべき先験的な理由はありません。
ここでは、我々のANAPタグ付きコンストラクトの発現を最大化する試み、特に細胞培養温度を33°Cに下げる試みがなされている10、11、16。我々の経験では、ANAPが保存的置換となるタンパク質中の部位を同定しようとしても、一貫して良好に発現するコンストラクトが得られなかった。タンパク質領域全体を系統的にスキャンしてANAP取り込み部位を探し、表面発現候補をスクリーニングすることに成功しました10。ANAPラベリングシステムはアフリカツメガエル卵母細胞でも機能し、はるかに大きな膜パッチを切除できるため、シグナル対ノイズが増加します26,27,28。
発現レベルが高いほどシグナルが明るくなると予想されますが、蛍光の測定に必要な最小チャンネル数は、蛍光色素の輝度、光退色の程度、励起光の強度、焦点面など、いくつかの要因によって異なります。理論的には、推定は、前に示したように蛍光強度とチャネル電流を相関させることによって行うことができます28,29。ただし、このような推定値の信頼性を高めるには、単一チャネルのコンダクタンスとチャネルのオープン確率に関するある程度の知識が必要です。上記の要因に加えて、蛍光シグナルは、電圧クランプされていないピペットガラスに付着した小胞または原形質膜のセクションに関連するチャネルによっても影響を受けます。
この方法は、他のヌクレオチド感受性イオンチャネルの研究に容易に適合する。CFTRは、KATP30、31の副スルホニル尿素受容体サブユニットと構造的に類似している。KATPと同様に、CFTRゲーティングはヌクレオチド結合によって制御されており、我々の方法7の明らかな将来の標的となっています。プリン作動性P2X受容体は、細胞外ATP9によってゲートされるイオンチャネルである。TNP-ATPは、P2X受容体32,33のアンタゴニストとして作用する。したがって、P2Xアゴニストとの競合アッセイで使用される可能性がありますが、P2X活性化の研究には役立ちません。あるいは、ANAP発光と十分なスペクトルオーバーラップを有する他の蛍光ATP誘導体を使用して、活性化を研究してもよい。Alexa−647−ATPは蛍光P2Xアゴニスト34である。Alexa-647とANAPの間で計算されたR0は~85 Åであり、これはP2Xへの直接結合がチャネルに組み込まれたANAPの実質的な消光をもたらすことを意味する。しかしながら、そのような長いR0はまた、隣接するサブユニットに結合したAlexa−647−ATPからの消光をもたらし、非特異的ヌクレオチド結合がFRETをもたらす可能性を高める。P2X受容体のリガンド結合部位は細胞外であるため、結合測定は無傷の細胞、全細胞電圧クランプ、または外側の膜パッチで行われます。私たちの方法は、反応サイクルをATPに依存する電気発生および非電気発生トランスポーターおよびポンプ、およびGタンパク質共役型P2Y受容体の結合および活性化の研究にも拡張できます。最後に、アデニンヌクレオチド結合(TNP-ATP、TNP-ADP、TNP-AMP)を測定するためにこの方法を開発しましたが、同じアプローチを使用して、適切な蛍光リガンドが同定されたほぼすべての受容体への結合を研究することができます。
開示事項
著者は利益相反を宣言しません。
謝辞
優れた技術支援を提供してくれたラウル・テロン・エクスポジトに感謝します。この研究は、バイオテクノロジーおよび生物科学研究評議会(BB / R002517 / 1;MCPおよびFMA)およびウェルカムトラスト(203731 / Z / 16 / A;SGU)
資料
| Name | Company | Catalog Number | Comments |
| T75 tissue-culture treated flask | StarLab | CC7682-4875 | |
| 0.1% w/v poly-L-lysine | Sigma-Aldrich | P8920 | |
| 30 mm borosilicate cover glass slips | VWR | 631-0174 | |
| 35 mm non-treated sterile dishes | CytoOne | CC7672-3340 | |
| 35 mm cover glass bottom dish | WPI | FD35-PDL-100 | |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 31966021 | |
| Foetal bovine serum (FBS) | Gibco | 10500-064 | |
| Penicillin/Streptomycin | Gibco | 15140-122 | |
| TrypLE select (tryosin) | Gibco | 12563-011 | Trypsin/EDTA reagent |
| Phosphate buffered saline (PBS) | Gibco | 14040-091 | |
| UltraPure distilled water | Invitrogen | 10977-035 | |
| HEK293T cells | ATTC | CRL-3216 | Used between passages 5-30 |
| ANAP-TFA | AsisChem | ASIS-0014 | Reconstituted in 30 mM NaOH to a final concentration of 1 mM |
| pANAP expression plasmid | Addgene | Plasmid #48696 | Encodes tRNA/tRNA synthetase pair for expression of ANAP-tagged protein |
| peRF1-E55D | Chin Lab (MRC Laboratory of Molecular Biology, Cambridge, UK) | Jason Chin: DOI: 10.1021/ja5069728 | Encodes dominant-negative eukaryotic ribosomal release factor |
| TransIT-LT1 | Mirus Bio | MIR 2300 | Lipopolyplex transfection reagent |
| Thick-walled borosilicate glass capillaries | Harvard Apparatus | GC150F-15 | |
| Tetramethylrhodamine-5-maleimide | Sigma-Aldrich | 94506 | |
| TNP-ATP | Jena Bioscience | NU-221L | Delivered at 10 mM in water |
| Nikon Eclipse TE2000-U inverted microscope microscope | Nikon | ||
| 60x water immersion objective (1.4 NA) | Nikon | MRD07602 | |
| 4-Wavelength High-Power LED Head | ThorLabs | LED4D245 | 385/490/565/625 nm LEDs |
| Four-Channel LED Driver | ThorLabs | DC4100 | |
| 390/18 nm band-pass excitation filter | ThorLabs | MF390-18 | For ANAP excitation |
| 400 nm long-pass emission filter | ThorLabs | FEL0400 | For imaging ANAP spectra |
| 416 nm edge dichroic | ThorLabs | MD416 | For imaging ANAP spectra |
| 460/60 nm band-pass emission filter | ThorLabs | MF460-60 | Suggested wide band-pass filter for imaging ANAP fluorescence (Figure 4B) |
| 470/10 nm band-pass emission filter | ThorLabs | FB470-10 | Suggested narrow band-pass filter for imaging ANAP fluorescence (Figure 4B) |
| 531/40 band-pass excitation filter | Brightline | FF01-531/40-25 | For orange fluorescent protein (OFP) excitation |
| 540/25 nm band-pass excitation filter | Chroma | D540/25X | For tetramethylrhodamine-5-maleimide (TMRM) excitation |
| 562 nm edge dichroic | Semrock | FF562-Di03 | For imaging OFP fluorescence |
| 565 nm edge dichroic | Chroma | 565DC | For imaging TMRM fluorescence |
| 593/40 nm band-pass excitation filter | Brightline | FF01-387/11-25 | For imaging OFP fluorescence |
| 605/55 nm band-pass emission filter | Chroma | D605/55M | For imaging TMRM fluorescence |
| IsoPlane-160 Imaging Spectrometer | Princeton Instruments | IsoPlane-160 | |
| PIXIS 400BR_eXcelon Camera | Princeton Instruments | PIXIS: 400BR_eXcelon | |
| Axopatch 200B amplifier | Molecular Devices | Axopatch 200B-2 | |
| Digidata 1440A digitizer | Molecular Devices | Digidata 1440A | |
| Probe sonicator | Sonics & Materials | VC-50 | For unroofing |
| REGLO digital peristaltic pump | Ismatec | ISM 832 | For bath perfusion |
| Microvolume perfusion system | ALA Scientific Instruments | ALA μFlow-8 | For TNP-ATP perfusion |
| pClamp 10.6.2 | Molecular Devices | Recording and analysing currents | |
| Lightfield 5.20.1507 | Princeton Instruments | Acquisition software for images and spectra | |
| Matlab | Mathworks | For data analysis | |
| Python 3.8.1 | Python Software Foundation | For data analysis |
参考文献
- Garcia, M. L., Kaczorowski, G. J. Ion channels find a pathway for therapeutic success. Proceedings of the National Academy of Sciences of the United States of America. 113 (20), 5472-5474 (2016).
- Hauser, A. S., Attwood, M. M., Rask-Andersen, M., Schioth, H. B., Gloriam, D. E. Trends in GPCR drug discovery: new agents, targets and indications. Nature Reviews Drug Discovery. 16 (12), 829-842 (2017).
- Lakowicz, J. R. . Principles of fluorescence spectroscopy. 3rd edn. , (2006).
- Higgins, C. F., Linton, K. J. The ATP switch model for ABC transporters. Nature Structural & Molecular Biology. 11 (10), 918-926 (2004).
- Toyoshima, C., Cornelius, F. New crystal structures of PII-type ATPases: excitement continues. Current Opinion in Structural Biology. 23 (4), 507-514 (2013).
- Craven, K. B., Zagotta, W. N. CNG and HCN channels: two peas, one pod. Annual Review of Physiology. 68, 375-401 (2006).
- Csanady, L., Vergani, P., Gadsby, D. C. Strict coupling between CFTR's catalytic cycle and gating of its Cl- ion pore revealed by distributions of open channel burst durations. Proceedings of the National Academy of Sciences of the United States of America. 107 (3), 1241-1246 (2010).
- Vedovato, N., Ashcroft, F. M., Puljung, M. C. The Nucleotide-Binding Sites of SUR1: A Mechanistic Model. Biophysical Journal. 109 (12), 2452-2460 (2015).
- Burnstock, G. Introduction to the Special Issue on Purinergic Receptors. Advances in Experimental Medicine and Biology. 1051, 1-6 (2017).
- Puljung, M., Vedovato, N., Usher, S., Ashcroft, F. Activation mechanism of ATP-sensitive K(+) channels explored with real-time nucleotide binding. Elife. 8, 41103 (2019).
- Usher, S. G., Ashcroft, F. M., Puljung, M. C. Nucleotide inhibition of the pancreatic ATP-sensitive K+ channel explored with patch-clamp fluorometry. Elife. 9, 52775 (2020).
- Chatterjee, A., Guo, J., Lee, H. S., Schultz, P. G. A genetically encoded fluorescent probe in mammalian cells. Journal of the American Chemical Society. 135 (34), 12540-12543 (2013).
- Gordon, S. E., Senning, E. N., Aman, T. K., Zagotta, W. N. Transition metal ion FRET to measure short-range distances at the intracellular surface of the plasma membrane. Journal of General Physiology. 147 (2), 189-200 (2016).
- Heuser, J. The production of 'cell cortices' for light and electron microscopy. Traffic. 1 (7), 545-552 (2000).
- Schmied, W. H., Elsasser, S. J., Uttamapinant, C., Chin, J. W. Efficient multisite unnatural amino acid incorporation in mammalian cells via optimized pyrrolysyl tRNA synthetase/tRNA expression and engineered eRF1. Journal of the American Chemical Society. 136 (44), 15577-15583 (2014).
- Lin, C. Y., et al. Enhancing Protein Expression in HEK-293 Cells by Lowering Culture Temperature. PloS One. 10 (4), 0123562 (2015).
- Usukura, J., et al. Use of the unroofing technique for atomic force microscopic imaging of the intra-cellular cytoskeleton under aqueous conditions. Journal of Electron Microscopy. 61 (5), 321-326 (2012).
- Colquhoun, D. Binding, gating, affinity and efficacy: the interpretation of structure-activity relationships for agonists and of the effects of mutating receptors. British Journal of Pharmacology. 125 (5), 924-947 (1998).
- Mannuzzu, L. M., Moronne, M. M., Isacoff, E. Y. Direct physical measure of conformational rearrangement underlying potassium channel gating. Science. 271 (5246), 213-216 (1996).
- Zheng, J., Zagotta, W. N. Gating rearrangements in cyclic nucleotide-gated channels revealed by patch-clamp fluorometry. Neuron. 28 (2), 369-374 (2000).
- Zheng, J., Zagotta, W. N. Patch-clamp fluorometry recording of conformational rearrangements of ion channels. Science's STKE. 2003 (176), 7 (2003).
- Biskup, C., et al. Relating ligand binding to activation gating in CNGA2 channels. Nature. 446 (7134), 440-443 (2007).
- Kusch, J., et al. Interdependence of receptor activation and ligand binding in HCN2 pacemaker channels. Neuron. 67 (1), 75-85 (2010).
- Wu, S., et al. State-dependent cAMP binding to functioning HCN channels studied by patch-clamp fluorometry. Biophysical Journal. 100 (5), 1226-1232 (2011).
- Cha, A., Bezanilla, F. Characterizing voltage-dependent conformational changes in the Shaker K+ channel with fluorescence. Neuron. 19 (5), 1127-1140 (1997).
- Kalstrup, T., Blunck, R. Dynamics of internal pore opening in K(V) channels probed by a fluorescent unnatural amino acid. Proceedings of the National Academy of Sciences of the United States of America. 110 (20), 8272-8277 (2013).
- Kalstrup, T., Blunck, R. S4-S5 linker movement during activation and inactivation in voltage-gated K(+) channels. Proceedings of the National Academy of Sciences of the United States of America. 115 (29), 6751-6759 (2018).
- Dai, G., Aman, T. K., DiMaio, F., Zagotta, W. N. The HCN channel voltage sensor undergoes a large downward motion during hyperpolarization. Nature Structural & Molecular Biology. 26 (8), 686-694 (2019).
- Liu, C., et al. Patch-clamp fluorometry-based channel counting to determine HCN channel conductance. Journal of General Physiology. 148 (1), 65-76 (2016).
- Hwang, T. C., et al. Structural mechanisms of CFTR function and dysfunction. Journal of General Physiology. 150 (4), 539-570 (2018).
- Puljung, M. C. Cryo-electron microscopy structures and progress toward a dynamic understanding of KATP channels. Journal of General Physiology. 150 (5), 653-669 (2018).
- Kasuya, G., et al. Structural insights into the competitive inhibition of the ATP-gated P2X receptor channel. Nature Communications. 8 (1), 876 (2017).
- Virginio, C., Robertson, G., Surprenant, A., North, R. A. Trinitrophenyl-substituted nucleotides are potent antagonists selective for P2X1, P2X3, and heteromeric P2X2/3 receptors. Molecular Pharmacology. 53 (6), 969-973 (1998).
- Bhargava, Y., Nicke, A., Rettinger, J. Validation of Alexa-647-ATP as a powerful tool to study P2X receptor ligand binding and desensitization. Biochemical and Biophysical Research Communications. 438 (2), 295-300 (2013).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved