
Method Article
İnsan Blastoidlerinin Modellenmesi Blastosist Gelişimi ve İmplantasyonu için Protokol
* Bu yazarlar eşit katkıda bulunmuştur
Bu Makalede
Özet
Verimli, zamanında ve sırayla blastosist benzeri hücreler üreten insan blastoidlerinin oluşumunu özetleyen bir protokol.
Özet
Kök hücrelerden (blastoid) oluşan insan blastosistinin bir modeli, bilimsel ve tıbbi ilerlemeleri destekleyecektir. Bununla birlikte, öngörücü gücü, blastosist gelişim dizilerini (morfogenez, spesifikasyon, modelleme) verimli, zamanında ve sadık bir şekilde özetleme ve blastosist aşamasını yansıtan hücreler oluşturma yeteneğine bağlı olacaktır. Burada, PXGL koşullarında kültürlenen ve daha sonra Hippo için üç kez inhibe edilen, büyüme faktörü β dönüştüren naif insan pluripotent kök hücrelerinin ve hücre dışı sinyal düzenlemeli kinaz yolaklarının blastoidleri oluşturmak için etkili bir şekilde morfogeneze maruz kaldığını gösteriyoruz (% >70). Gelişimsel zamanlama (~ 4 gün) ile eşleşen blastoidler, trofoblast ve epiblastın analoglarını üreterek blastosist spesifikasyon dizisini açar, ardından ilkel endoderm ve polar trofoblastların analoglarının oluşumu izler. Bu, blastosiste (>% 96) benzer hücrelerin transkripsiyonel olarak oluşmasına ve implantasyon sonrası analogların azınlığına neden olur. Blastoidler, uteroda olduğu gibi hormonal olarak uyarılmış endometriyal hücrelere yönlü olarak bağlanma potansiyelini kazanan polar bölgenin (NR2F2 +) olgunlaşması ile işaretlenmiş embriyonik-abembriyonik ekseni oluşturarak etkili bir şekilde modellenir. Böyle bir insan blastoidi, in vitro insan gelişimini ve implantasyonunu incelemek için ölçeklenebilir, çok yönlü ve etik bir modeldir.
Giriş
Deneysel modellerin eksikliği, erken insan embriyogenezinin anlaşılmasını sınırlamıştır. Embriyonik gelişimin insana özgü yönleri hakkındaki mevcut bilgiler, araştırma için bağışlanan fazla in vitro fertilizasyon (IVF) embriyolarından elde edilmektedir. Bununla birlikte, sınırlı kullanılabilirlik, deneysel manipülasyonların zorlukları ve embriyoların değişken kalitesi bilimsel araştırmaları engellemektedir. Aksine, insan embriyolarının sadık bir in vitro modeli, karmaşık deneysel manipülasyonlara izin verecek ve böylece insan embriyoları üzerindeki araştırmayı tamamlamak için etik bir fırsat sağlayacaktır 1,2,3,4. Daha önce geliştirilmiş bir fare blastosist modeli, fare embriyonik kök hücrelerini ve trofoblast kök hücrelerini birleştirdi5. Bu ayrıntılı protokolde, elemental blastosist kriterlerine uygun naif pluripotent kök hücrelerden insan blastosisti modeli oluşturmak için bir yöntem tanımlanmıştır6.
İnsan blastoidleri için dört kriter. Burada, insan blastoidlerinin standartlaştırılmış bir tanımını oluşturmak amacıyla, dört minimum kriter öneriyoruz. Kapsamlı olmasa da, bu kriterler insan blastoidlerinin oluşumuna izin veren parametreleri değerlendirmek için bir temel oluşturabilir (Şekil 1A). (1) Blastoidler, morfoloji ve epiblast (Epi), trofektoderm (TE) ve ilkel endoderm (PrE) olmak üzere üç soyun analoglarının üretilmesi açısından verimli bir şekilde oluşmalıdır. Verimsizliğin yetersiz bir başlangıç hücre durumuna veya / ve kültür durumuna (örneğin, blastoid ortam) işaret etmesi muhtemeldir. (2) Blastoidler, gelişimsel sekansa (Epi / TE ilk, PrE / polarTE son) 7,8 ve zamanlamaya (indüksiyon ~ 3 gün; embriyonik günler 5-7) 7,9'a göre üç soyun analoglarını üretmelidir. (3) Blastoidler, blastosist aşamasının analoglarını oluşturmalı, ancak implantasyon sonrası aşamaların (örneğin, implantasyon sonrası epiblast, trofoblast veya amniyon hücreleri) analoglarını oluşturmamalıdır. (4) Son olarak, blastoidler blastosist implantasyonu ve gelişiminin fonksiyonel özelliklerini özetleyebilmelidir. Bu protokolü kullanarak, insan blastoidleri çoklu hücre hatları (% >70) kullanarak verimli bir şekilde oluşur, blastosist hücresel analoglarını sırayla ve 4 gün içinde üretebilir ve analoglar transkripsiyonel olarak blastosist aşamasına benzer (birkaç analize dayanarak% >96)6,10,11. Son olarak, blastoidler embriyonik-abembriyonik ekseni sağlam bir şekilde üretir, bu da hormonal olarak uyarılmış endometriyal hücrelerle polar bölge boyunca etkileşime girmelerini sağlar ve genişletilmiş kültür üzerindeki soyları sağlam bir şekilde genişletir (zaman eşdeğeri: embriyonik gün 13).
İlk hücre durumunun önemi. İnsan pluripotent kök hücreleri (hPSC'ler), hassas gelişimsel aşamaları yakalamaya çalışan farklı durumlarda stabilize edilebilir. Bu durumlar, hala optimal olmamasına rağmen, implantasyon öncesi (~ embriyonik günler 5-7) veya implantasyon sonrası benzeri (~ embriyonik günler 8-14) epiblast evresi12'deki hücreleri kısıtlayan kültür koşulları tarafından sürdürülür. Transkriptomik analiz, PD0325901, XAV939, Gö6983 ve lösemi inhibitör faktöründe (LIF; PXGL naif hPSC'ler olarak adlandırılır) kültürlenen hPSC'lerin, fibroblast büyüme faktörü (FGF) 2 ve aktivin15'te (astarlanmış hPSC'ler 12) ve insan genişletilmiş pluripotent kök hücrelerinde (hEPSC'ler) kültürlenen hPSC'lere kıyasla blastosist epiblastına daha benzer olduğunu göstermiştir. 18,19). Buna göre, astarlanmış hPSC'lerin transkriptomu, implantasyon sonrası / gastrulasyon öncesi cynomolgus maymun epiblastı20 ile en iyi şekilde eşleşir. Transpozon ekspresyonu, DNA metilasyonu ve X kromozom durumu gibi ek moleküler kriterler, naif durumun varyasyonlarının, astarlanmış duruma kıyasla blastosist epiblastına daha yakından benzediğini doğruladı17,21. Son olarak, naif hPSC hatları, PXGL kültür koşulları22 kullanılarak doğrudan blastosistlerden başarıyla türetilmiştir.
İnsan erken blastosist hücreleri henüz işlenmemiştir. Murin soy spesifikasyonu, blastosist evre23'ten önceki morula evresinden oluşur. Aksine, ayrışma ve yeniden agregasyon deneyleri, erken blastosistlerin insan trofektoderm hücrelerinin henüz işlenmediğini göstermiştir24. Buna göre, insan blastosistlerinin hücrelerinin tek hücreli RNA dizilimi (scRNAseq) ile analizi, ilk soy spesifikasyonunun (trofoblast / epiblast) blastosist boşluğunun7 oluşumundan sonra gerçekleştiğini göstermiştir. Bu ertelenmiş insan spesifikasyonu, fare PSC'leri epiblast soyuna büyük ölçüde bağlı kaldıklarında, hPSC'lerin trofoblast25,26,27 oluşturmak için güçlü olduğu gözlemleriyle ilişkilidir. Bu kombine gözlemler, naif hPSC'lerin bir blastosist aşamasını yansıtması ve üç blastosist soyunu oluşturma potansiyelini koruması olasılığına yol açtı. Son zamanlarda, hPSC'lerin ekstraembriyonik analogları belirleme gücünün, naiften astarlanmış duruma ilerleme sırasında trofektodermden amniyona kayması önerilmiştir27. Bu nedenle, naif hPSC'ler implantasyon öncesi aşama 17,18,21'e daha benzerdir ve implantasyon sonrası analogları oluşturmaya eğilimli olan astarlanmış hPSC'ler 27, hEPSC'ler 16 veya ara yeniden programlanmış durumlar 28'e kıyasla trofoblastlar oluşturmak için gelişmiş bir kapasiteye sahiptir10 (Şekil 1B ). Bu nedenle başlangıç hücre durumu, uygun ekstraembriyonik analogları oluşturmak için çok önemlidir. Dönüştürülmüş trofektoderm analoglarının kapsamlı bir yan yana analizi yapılmaya devam etse de, erken blastosisti yansıtan bir PXGL naif durumu, yüksek doğruluklu blastoidler oluşturmak için önemli görünmektedir.
Sinyal yolaklarının inhibisyonu ile spesifikasyon ve morfogenezin uyarılması. Hippo sinyal yolunun inhibisyonu, farelerde, ve insanlarda trofoblast spesifikasyonunu yönlendiren korunmuş bir mekanizmadır 9,29,30. Ayrıca, 2013 yılından bu yana, NODAL (A83-01) ve hücre dışı sinyal regüle kinaz (ERK; PD0325901 veya eşdeğeri) ve kemik morfogenetik protein (BMP) sinyal yolaklarının aktivasyonu, trofoblast soyu 25,31,32,33,34 ile ilişkili transkripsiyonel ağı aktive etmek için astarlanmış hPSC'leri tetikler. Ayrıca, son zamanlarda birkaç rapor hem NODAL hem de ERK yolunun inhibisyonu ve BMP'nin aktivasyonunun, naif hPSC'lerdentrofoblast farklılaşmasını kolaylaştırdığını doğrulamıştır 25,31,32,33,34. Son olarak, trofoblast spesifikasyonu naif bir durumdan tetiklenirse, hücreler trofektoderm26'nın gelişimsel ilerlemesinin yönlerini özetler. Bununla birlikte, blastosist trofektodermini yansıtan kendi kendini yenileyen çizgiler in vitro olarak stabilize edilmemiştir. Trofoblast spesifikasyonunu takiben, HDAC inhibisyonu ile birlikte epidermal büyüme faktörünün (EGF) ve Wnt sinyal yollarının indüklenmesi, trofoblast gelişimsel progresyonunu34,35 kolaylaştırabilir ve hücreleri, implantasyon sonrası sitotrofobikler 18,35'i yansıtan insan trofoblast kök hücrelerinin (hTSC'ler) hatlarına stabilize edebilir. Bu çizgiler hem blastosistlerden hem de plasenta dokularından türetilebilir35.
PrE olarak adlandırılan ikinci ekstraembriyonik soy, trofoblastlardan sonra belirtilir ve epiblast 7,9'dan kaynaklanır. Murine PrE36'nın aksine, insan muadilinin FGF sinyalinin 37,38'den bağımsız olduğu düşünülmektedir. Ekstraembriyonik endodermi (nEnd olarak adlandırılır) yansıtan çizgiler, naif hPSC'lerden aktivin A, Wnt ve LIF39 kullanılarak sinyal yolaklarının indüklenmesiyle oluşturulmuştur. Embriyo inhibisyon deneyleri ile tutarsız olarak, ERK inhibisyonunun in vitro39 gibi nEND hücrelerinin oluşumunu önlediği gösterilmiştir. Şimdiye kadar, bu tür çizgiler doğrudan blastosistlerden türetilmemiştir.
Son zamanlarda, erken embriyonun modelleri, daha önce hTSC'ler 35 ve nEND hücreleri39 için geliştirilen ortamın varyasyonlarının birleştirilmesiyle oluşturulmuştur, böylece dönüştürücü büyüme faktörü - β (TGF-β), EGF ve Wnt sinyal yollarının aktivatörleri kullanılarak28,40. Bu embriyo modelleri düşük verimlilikte (% 10 -% 20) oluşur ve implantasyon sonrası epiblast, trofoblast, amniyon, gastrula, mezodermal dokular (~ embriyonik gün14) ve sitotrofoblastların analogları dahil olmak üzere implantasyon öncesi aşama10'a benzeyen hücreler oluşturur. Aksine, Hippo, ERK ve TGF-β yolaklarının üçlü inhibisyonu blastosist benzeri hücreleri içeren blastoidlerin oluşumunu etkili bir şekilde yönlendirir41. İlk hücre durumu ile birlikte, üçlü yolak inhibisyonunun (Hippo, ERK, TGF-β) yüksek doğrulukta blastoidler oluşturmak için ikinci temel parametre olduğunu öneriyoruz (Şekil 1B).
ScRNAseq kullanarak hücre durumunu ve yansıyan aşamayı değerlendirme. Blastoidleri oluşturan hücrelerin durumları scRNAseq analizi ile değerlendirilebilir. Spesifik embriyonik aşamalara transkripsiyonel benzerlikleri, tek başına blastoid hücreler kullanılarak ve implantasyon sonrası aşamaları20,35 yansıtan astarlanmış hPSC'ler veya hTSC'lerle karşılaştırılarak ölçülebilir. Farklı tanım düzeyleri kullanarak küme analizleri yapmak, tanım azaldığında alt popülasyonların aşamalı olarak nasıl birleştiğini ortaya çıkarır ve böylece kümelerin benzerliklerini ortaya çıkarır. Küme sayısındaki optimalite42 olarak ölçülebilse de, yüksek çözünürlüklü kümeleme, örneğin implantasyon sonrası aşamaları yansıtan küçük anormal alt popülasyonların nihai varlığı hakkında da bilgi verir10. Kümeler arasında farklı şekilde eksprese edilen genler, aşamaya özgü soyları tanımlayan referans gen kümelerinin ekspresyon seviyelerini değerlendirerek gelişim sürecinde analogları hakkında bilgi sağlayabilir. Bu, blastoid alt popülasyonlarının zenginleştirilmesinin ya denetimsiz mesafe haritaları (örneğin, üstten zenginleştirilmiş genler kullanılarak) ya da gen seti zenginleştirme analizi (GSEA) ile ölçülmesini sağlar43. Bu blastoid protokolünü kullanarak, üç blastosist soyunu transkripsiyonel olarak yansıtan sadece üç ana küme oluşur. Bir küme hem başlangıçtaki naif hPSC'leri hem de blastoidlerin epiblast analoğunu içerir. Farklı zaman noktalarındaki hücrelerin analiz edilmesi, soy spesifikasyonunun sıralı doğasını göstermiştir (trofoblastlar 24 saat içinde ve ilkel endoderm hücreleri 60 saat içinde belirtmeye başlar). Yüksek çözünürlüklü bir kümeleme, gastrulasyon aşaması embriyolarına (muhtemelen mezoderm veya amniyon) özgü genleri eksprese eden bir hücre alt popülasyonunu (% 3.2) yakaladı. Not olarak, ilk naif hPSC'ler, dahaönce tarif edildiği gibi, implantasyon sonrası benzeri hücrelerin% 5'ini de oluşturuyordu. İkinci bir analizde, blastoid hücreler, evre eşdeğerliğini çıkarmak için farklı aşamalarda45,46,47 farklı aşamalarda concepti'den izole edilen referans hücrelerle siliko olarak birleştirilebilir. Burada preimplantasyon concepti 45,46, in vitro kültürlü blastosistler 45 ve gastrulasyon evresi embriyoları 47'den izole edilen hücreler referans noktası olarak kullanılmıştır. Bu protokol kullanılarak, yüksek çözünürlüklü kümelenme ile ortaya çıkarılan uyumsuz blastoid hücrelerin gerçekten implantasyon sonrası mezoderm ve amniyon ile kümelendiği ölçülmüştür. Gelecekteki adımlarda, transkriptom kıyaslaması, transpozon ekspresyonu, DNA metilasyonu ve X kromozomu durumunun analizi ile tamamlanmalıdır ve bu da gelişimsel aşamaların21.
İnsan blastoidlerinin eksen oluşumunu ve diğer işlevlerini değerlendirmek. Olgun bir blastosist, implantasyon için embriyonik-abembriyonik eksen desenli trofoblastların oluşumu ile karakterizedir. Bu blastoid protokolünü kullanarak, proksimal trofoblastların (örneğin, NR2F2 + / CDX2-) olgunlaşmasıyla örneklenen ve endometriyal organoid hücrelere yalnızca hormonal olarak uyarıldığında bağlanma kapasitesini kazanan sağlam bir eksen oluşur48,49. Epiblastı oluşturmayan trofosferlerle karşılaştırma, bu iç hücrelerin endometriyuma ilk bağlanmaya aracılık etmek için abutting trofoblastların olgunlaşmasını indüklediğini göstermektedir. Sinomolgus maymun blastosistleri50 için tasarlanmış genişletilmiş bir kültür ortamında kültürlendiğinde, blastoidden gelen üç soyun da organizasyonları bu gelişim aşamasını yansıtmasa da, altı ek gün boyunca (13. günün zaman eşdeğeri) sürekli olarak genişler.
Yüksek verimli ve yüksek kaliteli insan blastoidlerinin anlamı. Model organizmalarda keşfedilen gelişimsel ilkelerin korunması, sınırlı erişim ve genetik ve fiziksel olarak manipüle edilmesindeki teknik zorluklar nedeniyle insan conceptusunda test edilmesi doğal olarak zordur. Yüksek verimli ve yüksek doğruluklu bir blastoid modeli, bilimsel ve biyomedikal keşiflerin temelinde yer alan yüksek verimli genetik ve ilaç ekranlarına izin verecektir. Ek olarak, biyolojik süreçleri değiştirmek ve kaydetmek için karmaşık genetik modifikasyonların dahil edilmesi bu tür çalışmaları tamamlayacaktır. Genel olarak, naif PXGL hPSC'lerin üçlü inhibisyonunun (Hippo, TGF-β, ERK), dört minimum kritere uygun yüksek kaliteli insan blastoidlerinin verimli oluşumu için iletken olduğunu öneriyoruz. Bu protokolün ölçeklenebilir ve çok yönlü doğası, daha sonra insan blastosistleri kullanılarak doğrulanabilecek hedefli hipotezler üretmeyi uygun kılar. Bu nedenle, insan blastoidleri, in vitro araştırmalar için insan conceptus'un kullanımının yerini almayacak, ancak bilimsel ve biyomedikal keşif sürecinin kalbinde daha önce erişilemeyen deneysel yaklaşımlarla araştırmayı yönlendirmenin güçlü bir yolu olarak hareket edebilir. Protokol, insan blastoidlerinin nasıl oluşturulacağını ve ayrıca blastoid içinde bulunan hücrelerin nasıl analiz edileceğini gösterir.
Protokol
Uluslararası Kök Hücre Araştırmaları Derneği (ISSCR) Kök Hücre Araştırmaları ve Klinik Çeviri Kılavuzları, insan blastoidleri üzerine yapılan araştırmalara ancak özel bir bilimsel ve etik inceleme süreci aracılığıyla inceleme ve onaydan sonra izin verilmesini önermektedir 3,4. Tüm deneysel prosedürler, Avusturya Bilim Akademisi Moleküler Biyoteknoloji Enstitüsü (IMBA) insan araştırma etik komitesinin Rivron_Stellungnahme_2020-04-22 onayı altındaki yönergeleri izlenerek gerçekleştirilmiştir. Bu yönergelere uygunluk, araştırma sonuçlarının bilimsel dergilerde yayınlanması için gereklidir.
1. PXGL durumunda insan naif embriyonik kök hücre kültürü
NOT: Naif hPSC'ler ilgili laboratuvarlardan temin edilebilir. Burada kullanılan hatlar Yasuhiro Takashima (şu anda CiRA, Kyoto, Japonya'da) ve Austin Smith'in (şu anda Living Systems Institute, Exeter, İngiltere'de) laboratuvarlarından elde edilmiştir. Alternatif olarak, naif hPSC'ler, daha önce13,14'te açıklandığı gibi astarlanmış hPSC'lerin hatlarından şirket içinde sıfırlanabilir. Naif hPSC'ler birden fazla pasaj için kararlı görünür (> 15) ancak kültürün kalitesi zamanla değişebilir. Naif hPSC'nin kalitesi düşerse, yeni bir şişe hücreyi eritin veya astarlanmış PSC'lerden de novo naif hPSC'ler üretin. Tüm medya kompozisyonları için Ek Tablo 1'e bakınız.
- Işınlanmış fare embriyonik besleyici (MEF) tabakasının hazırlanması
- Naif hPSC'lerin geçişinden bir gün önce, aşağıda açıklanan adımları izleyerek ışınlanmış MEF katmanlarına sahip 6 delikli bir hücre kültürü plakası hazırlayın.
- 6 kuyucuklu bir hücre kültürü plakasını, kuyu başına PBS'de 1 mL% 0.1 jelatin ile kaplayın. Plakayı 30 dakika boyunca 37 ° C'de inkübe edin. Jelatin çözeltisini çıkarın.
- MEF ortamını 37 °C'de hazırlayın.
- MEF'leri 37 ° C'de bir su banyosunda sadece küçük bir buz kümesi kalana kadar çözün. Bir P1000 pipet kullanarak şişenin hacmini 1 mL hazırlanmış MEF ortamı ile çözün.
- Hücre süspansiyonunu 15 mL'lik bir tüpe aktarın. Süspansiyonu 4 dakika boyunca 200 x g'de döndürün. Süpernatantı aspire edin ve taze MEF ortamı ekleyerek MEF peletini yeniden askıya alın (kuyu başına 1,5 mL için yeterlidir).
- Hücre sayma slaytlarını kullanarak hücreleri sayın ve kuyu başına 300.000 hücre ekleyin ve plakayı 37 ° C'de bir normoksi inkübatöre aktarın.
NOT: MEF'ler zaman içinde ayrılırsa, rutin ortam değişimi sırasında PXGL ortamına yeni MEF'ler eklenebilir.
- İnsan naif pluripotent kök hücrelerinin geçişi
- hPSC'leri geçmeden önce, mikroskop altında morfolojilerini kontrol edin. Koloniler tipik olarak parlak, tanımlanmış sınırları olan kubbe şeklinde bir morfolojiye sahiptir. Bireysel koloniler daha düz bir morfoloji gösteriyorsa veya farklılaşmış koloniler ortaya çıkmaya başlarsa, adım 1.2.8'deki talimatları izleyin.
- Ortamı aspire edin ve hücreleri PBS ile bir kez yıkayın. 6 delikli bir plakanın kuyucuğu başına 500 μL hücre ayırma çözeltisi ekleyin.
- Hücreleri 37 ° C'de 5 dakika boyunca inkübe edin. Bir P1000 pipet kullanın ve kolonileri tek hücrelere ayırmak için hücreleri birkaç kez pipetleyin.
- Hücreleri toplayın ve bunları yıkama tamponu içeren 15 mL'lik bir tüpe aktarın (6 delikli bir plakanın kuyusu başına 1 mL). Hücreleri 4 dakika boyunca 200 x g'de döndürün.
- Süpernatantı aspire edin ve peleti taze PXGL ortamında yeniden askıya alın (kuyu başına 1,5 mL için yeterlidir). Rutin olarak pasaj için 1:3-1:6 bölme oranını göz önünde bulundurun.
NOT: Her 3-4 pasajdan sonra veya hücre morfolojisine bağlı olarak hücre kültürünün kalitesi düşerse (örneğin, popülasyonda düz kolonilerin ortaya çıkması), geçişten sonraki ilk 24 saat boyunca ortama 10 μM Y-27632 ve büyüme faktörü bazal membran ekstraktı (5 μL / kuyu) ekleyin. - hPSC'leri yeniden kaplamadan önce, MEF ortamını aspire ederek ve hücreleri PBS ile bir kez yıkayarak plakaları taze MEF'lerle hazırlayın. Ardından, MEF'leri içeren 6 delikli bir plakanın kuyusu başına 1,5 mL hPSC hücre süspansiyonunu aktarmak için bir P1000 pipet kullanın.
NOT: Pipetlemenin, kuyu alanı boyunca hücrelerin homojen bir şekilde tohumlanmasına yol açtığından emin olun. Bu, homojen boyutlara ve hücrelerin göreceli senkronizasyonuna sahip kolonilerin büyümesini sağlayacaktır. - Kültür hPSC'leri hipoksik koşullar altında nemlendirilmiş bir ortamda 37 °C'de bulunur. 24 saat sonra hPSC'ler takılmalıdır. Çok sayıda yapışkan olmayan (veya yüzen) hücre, pasajdan sonra yaşayabilirlik veya bağlanma problemini yansıtır.
- Ortamı kuyucuk başına günlük 1,5 mL PXGL ortamı ile değiştirin. Her 3-4 günde bir hPSC'leri geçirin veya blastoid oluşum deneyleri için kullanın.
NOT: hPSC'leri çözdükten sonra, bir blastoid deneyine başlamadan önce bunları en az üç pasaj boyunca geçirin.
2. Blastoidlerin Oluşumu
- Naif PSC agregalarının oluşumu
- Deneye başlamadan önce PXGL ortamını, N2B27 bazal ortamı, yıkama arabelleğini, PBS'yi ve toplama ortamını hazırlayın ve önceden ısıtın. Aşağıda açıklanan adımları izleyerek MEF'leri blastoid oluşturmak için hPSC'lerin süspansiyonundan hariç tutun.
- MEF hariç tutma için, 6 delikli bir plakanın kuyucuğuna 1 mL% 0.1 jelatin ekleyerek ve 30-90 dakika boyunca 37 ° C'de inkübe ederek jelatin kaplı bir plaka hazırlayın.
- Hücreleri hasat etmek için, ortamı aspire edin ve hücreleri 1 mL PBS ile yıkayın.
- 500 μL hücre ayırma çözeltisi ekleyin (6 delikli bir plakanın kuyucuğu başına) ve 5 dakika boyunca 37 ° C'de inkübe edin.
- Kolonilerin tek hücrelere ayrışmasını takip etmek için mikroskop altındaki hücreleri kontrol edin (birkaç çok hücreli küme daha sonra nazik pipetleme ile ayrıştırılabilir).
- Hücre ayrılma çözeltisini 1 mL yıkama tamponu ile seyreltin. 5 ila 10 kez hafifçe pipetleyerek hücreleri plakadan toplayın. Hücre süspansiyonunu 15 mL'lik bir tüpe aktarın. Hücreleri 4 dakika boyunca 200 x g'de döndürün.
- Süpernatanı aspire edin, hücreleri 1.5 mL PXGL ortamında (6 delikli bir plakanın kuyucuğu başına) yeniden askıya alın ve hücreleri MEF dışlaması için jelatin kaplı plakalara tohumlayın ve 60-90 dakika boyunca 37 ° C'de inkübe edin.
- Naif hücreler MEF dışlaması için tohumlandıktan sonra, PBS'yi mikro kuyulardan çıkarın ve kuyuları 200 μL bazal N2B27 ortamı (1 mikrowell çipi başına) ile dengeleyin ve 37 ° C'de 60 dakika boyunca inkübe edin.
- Bağlanmamış naif hücreleri içeren süpernatantı toplayın, 15 mL'lik bir tüpe aktarın ve hücreleri 4 dakika boyunca 200 x g'de döndürün.
- Medyayı aspire edin ve hücreleri 1 mL N2B27 bazal ortamda yeniden askıya alın. Hücre sayma slaytlarını kullanarak hücreleri sayın. Hücreleri 4 dakika boyunca 200 x g'de döndürün.
- Ortamı aspire edin ve 50 μL başına 30.000 hücrelik bir hücre yoğunluğu elde etmek için N2B27 ortamında bulunan uygun miktarda 10 μM Y-27632 ekleyin.
NOT: Optimal ilk tohumlama hücre sayısı, farklı hücre hatları arasında değişebilir. Örneğin, 50-60 hücre/mikrokuyu tohumlamak için, 30.000 hücre (bazı hücrelerin mikro kuyunun dışına düştüğü göz önüne alındığında fazlalık dahil), 430 mikrokuyu içeren 96 kuyu plakasının 1 kuyusuna ekilir. Uygun olmayan bir başlangıç hücresi sayısı, boşluksuz küçük agrega oluşumuna veya 250 μm'den daha fazla kavitasyonlu yapı oluşumuna neden olabilir. - Ortamı dengeli mikrowell dizilerinden aspire edin ve 10 μM Y-27632 ile 25 μL N2B27 ortam ekleyin. 50 μL hücre süspansiyonu ekleyin ve 15 dakika boyunca 37 ° C'de inkübe edin (hücreler mikro kuyunun dibine düşene kadar). Ardından, 10 μM Y-27632 ile desteklenmiş 125 μL N2B27 ortamı ekleyin.
- Blastoid gelişimi
- 24 saat içinde, mikro kuyu çipi üzerinde naif hPSC'lerin agregaları gözlemlenebilir (0. gün). Blastoid oluşumunu başlatmak için PALLY ortamını hazırlayın ve aşağıda özetlenen adımları izleyin.
- 30 dakika boyunca 37 °C'de ön sıcak PALLY ortamı.
NOT: 1-Oleoil Lizofosfatidik asit (LPA) ve 10 μM Y-27632 kullanımdan hemen önce eklenmelidir. Optimal LPA konsantrasyonu 0.5-5 μM arasında değişir. Bu, blastoidler için kullanılan bireysel hPSC hatları için titre edilmelidir. - Toplama ortamını aspire edin. Mikro kuyucuklara 200 μL önceden ısıtılmış PALLY ortamı ekleyin. Hücre kültürü plakasını 37 ° C'de hipoksik bir inkübatöre geri yerleştirin. Medya değişikliğini 1. günde tekrarlayın.
- 2. günde, PALLY ortamını çıkarın ve LPA ve 10 μM Y-27632 ile desteklenmiş 200 μL N2B27 ortamı ekleyin.
NOT: 2. günde agregaların çoğunluğu büyümeye devam eder. Bununla birlikte, bazı agregalar küçük boşluklar oluşturur. 4. güne kadar PALLY içinde blastoidleri sürekli olarak kültüre alın veya 2. günden itibaren in vitro kültür ortamı (IVC1). Bununla birlikte, bu ortam değişikliğini takiben, olgun blastoidlerde PrE oluşumunu arttırır. - 3. günde medya değişikliğini tekrarlayın. Tam blastoid oluşumu 4. günde gerçekleşir.
NOT: Blastoidlerin, 7. gün insan blastosistlerinin morfometrisine (örneğin, 150 -250 μm arasında bir çap aralığı; trofektoderm benzeri hücrelerin bir epiteli ile çevrili bir iç küme) dayanan tam morfogenez geçirdikleri ve polar trofektoderm benzeri hücreler (NR2F2 + / CDX2-) ve PrE benzeri hücreler (GATA +) oluşturdukları zaman tamamen gelişmiş oldukları düşünülmektedir. Bu, immünofloresan boyama veya floresan ile aktive edilmiş hücre sıralama (FACS) kullanılarak değerlendirilebilir.
3. 96 kuyucuklu ultra düşük ataşmanlı mikroplakalarda blastoid oluşumu
- Yukarıda 2.1.1 - 2.1.11 arasında açıklanan adımları izleyerek blastoid oluşumu için naif hPSC'lerin hücre süspansiyonunu hazırlayın.
- Ortamı aspire edin ve ortamın 100 μL'si başına 70 hücrenin hücre yoğunluğunu elde etmek için 10 μM Y-27632 içeren uygun miktarda N2B27 ortamı ekleyin.
NOT: Optimal ilk tohumlama hücre sayısı, farklı hücre hatları arasında değişebilir. Örneğin, 70 hücre / kuyu, çoğu hücre satırı için en uygun hücre numarası olabilir. - Kuyucukların dibindeki hücreleri kümelemek için plakayı oda sıcaklığında 2 dakika boyunca 200 x g'de santrifüj yapın.
- Plakayı hipoksik kültür koşulları altında 37 ° C'de bir inkübatörde inkübe edin. 24 saat içinde kuyucuklarda naif hPSC agregaları gözlemlenebilir (0. gün).
- 2x PALLY ortamı hazırlayın. Kuyucuklara 100 μL önceden ısıtılmış 2x PALLY ortamı ekleyin.
- Hücre kültürü plakasını 37 ° C'de hipoksik bir inkübatöre geri yerleştirin. 24 saat sonra, ortamın yarısını (100 μL) aspire edin ve 100 μL önceden ısıtılmış PALLY ortamı ile değiştirin. 4. güne kadar adımı tekrarlayın. Agregaları aspire etmediğinizden emin olun.
NOT: 2. günde, agregaların çoğunluğu büyümeye devam eder. Bununla birlikte, bazı agregalar küçük sıvı dolu boşluklara sahiptir. 4. günde, kavitasyonlu yapıların çoğu, blastosist benzeri yapılar oluşturmak için tam morfogeneze uğrar.
4. Trofosferlerin oluşumu
- Trofosfer oluşumu için, adım 2.1.1'den (MEF dışlaması) adım 2.1.12'ye (tohumlama protokolünün son adımı) kadar blastoid oluşum protokolünü izleyin.
- Naif hPSC'lerin agregaları 24 saat sonra oluştuktan sonra, erken trofektodermi temsil eden trofosferlerin oluşumu için 3 μM SC-144 ile desteklenmiş PALY (LIF'siz) ve olgun trofektodermi temsil eden trofosferlerin oluşumu için 2 μM XMU-MP-1 ile desteklenmiş PALLY ile agregasyon ortamını değiştirin.
- Ortamı günlük olarak yenileyin. Tam trofosfer oluşumu 4. günde gerçekleşir.
5. scRNAseq kullanarak blastoid hücrelerin durumunun ve yansıyan aşamasının analizi
- Blastoidleri almak ve ayrışma gerçekleştirmek için, sallanan inkübatörü 37 ° C'ye ısıtın ve 100 rpm'ye ayarlayın.
- İlk 96 delikli plakadan blastoidleri toplayın ve cam kılcal damarla donatılmış bir ağız pipeti kullanarak bunları U tabanlı 96 delikli bir plakanın birden fazla kuyucuğuna aktarın.
NOT: Blastoid olmayan yapılarla (>% 30) kontaminasyonu önlemek için blastoidler (< %70) morfometrik kriterlere (benzersiz bir iç küme ile boyut = 150-250 μm) göre seçilmelidir. - Bir kez stereomikroskop altında görüntüleyerek bir P200 kullanarak 200 μL PBS ile yıkayın. 50 μL kollajenaz içeren bir kuyuya aktarın ve çalkalayan inkübatörde 30 dakika boyunca inkübe edin.
- Blastoidleri 100 μL 10x tripsin-EDTA içeren bir kuyuya aktarın ve iyice karıştırın. Sallanan inkübatörde 20 dakika boyunca kuluçkaya yatırın.
- P200 pipet kullanarak blastoidleri tek hücreli olarak ayırın. Hücreleri FACS tamponlu 15 mL'lik bir tüpe aktarın (PBS'de% 1 FBS).
- Üç soyun analoglarının spesifik oranlarını yakalamak için, TE ve PrE analoglarını sırasıyla TROP2 ve PDGFRa antikorları ile lekeleyin.
NOT: İnsan blastosistlerindeki PrE hücrelerinin sayısı, fare blastosistlerine kıyasla daha azdır, bu da in vitro fertilizasyon (IVF) veya bir tür farkı yoluyla oluşan blastosistlerin gelişimsel kusurlarını yansıtabilir. Blastoidlerde, PrE analogları TE ve EPI analoglarından daha az miktarda bulunur ve immünofloresan görüntüleme ile GATA4 + hücrelerinin sayımı üzerine hücrelerin% 7.4'ünü temsil eder. Ayrıca, ayrışma süreci, blastoid ayrışması, PDGFRa etiketlemesi ve FACS analizi üzerine hücrelerin %1-2'sini temsil etmek için PrE analogları olarak farklı hücre tiplerinin oranlarında önyargılara neden olabilir. - FACS, her üç soy analogundan hücreleri, akıllı-seq2 analizi için bir lizis tamponu içeren 384 plakaya ayırır. DAPI boyama ile işaretlenmiş ölü hücreleri hariç tutun (üreticinin talimatlarına göre gerçekleştirilir).
- Hücre durumlarını (hücre tipi ve gelişim evresi) değerlendirmek için, blastoidden alınan transkriptomik verileri uygun kontrollerle karşılaştırın.
6. Blastoid gelişimsel ilerlemesini değerlendirmek için genişletilmiş kültür
- Kültür insan blastoidleri bazal membran matris kaplı plakalar (cam taban) üzerinde.
- Plakayı bazal membran matrisi ile kaplayın.
- Morfolojiyi değerlendirmek ve kaydetmek için blastoidleri görsel olarak inceleyin.
NOT: Sadece kompakt ICM ile klasik içi boş bilyalı blastosist morfolojisini gösteren blastoidler daha da büyüme ve gelişme potansiyeline sahiptir. - 96 delikli plakanın bir kuyucuğuna 100 μL CMRL orta-1 ekleyin ve blastoidler denge için transfer edilmeden en az 2 saat önce plakayı inkübatöre yerleştirin.
- Bir stereomikroskop kullanarak, blastoidleri iyi morfolojiye sahip görsel olarak tanımlayın, blastoid ortamın izlerini gidermek için seçilen blastoidleri 100 μl CMRL orta-1 içeren 96 kuyucuklu bir plaka kuyucuğuna aktarın.
- Blastoidleri, önceden dengelenmiş CMRL ortamı-1'i içeren kuyuya aktarın. Plakayı inkübatöre yerleştirin ve gece boyunca 37 ° C'de inkübe edin.
NOT: 96 kuyucuk plakasından oluşan bir kuyucukta 5 adede kadar blastoid kültürlenebilir. Tek bir kuyucukta çok fazla blastoid bulunması, çoklu blastoidlerin agregalarının oluşumuna yol açabilir. - Ertesi gün, blastoidleri mikroskop altında görsel olarak inceleyin. Blastoidler takılıysa,% 5 bazal membran matrisi ile desteklenmiş 100 μL önceden dengelenmiş CMRL ortam-1 ekleyin. Plakayı inkübatöre yerleştirin ve gece boyunca 37 ° C'de inkübe edin.
- Ertesi gün, blastoidleri mikroskop altında izleyin. Ortamın yarısını (100 μL) çıkarın ve% 5 bazal membran matrisi ile desteklenmiş 100 μL önceden dengelenmiş CMRL ortam-2 ile değiştirin.
- Sonraki günlerde, ortamın yarısını (100 μL), %5 bazal membran matrisi ile desteklenmiş, önceden dengelenmiş CMRL ortam-3 ile değiştirin. Plakayı inkübatöre yerleştirin ve gece boyunca 37 ° C'de inkübe edin. Her gün 4-6 güne kadar in vitro kültür boyunca tekrarlayın.
NOT: Blastoidleri, in vitro kültürlenmiş insan embriyolarının 13. gününün zaman eşdeğerine karşılık gelen genişletilmiş kültür koşullarında 6 güne kadar kültürlü blastoidlerimiz var.
7. İmmün boyayan blastoidler
- Ortamı aspire edin. Numuneleri PBS ile 5 dakika boyunca 3 kat yıkayın.
- PBS'ye 200 μL soğuk% 4 paraformaldehit (PFA) ekleyin ve numuneleri oda sıcaklığında 30 dakika sabitleyin. PFA çözeltisini çıkarın ve numuneleri PBS ile 10 dakika boyunca 3 kat yıkayın.
NOT: Blastoidler mikrowell çipleri üzerinde kültürlenmişse, aşağıdaki adımlar için blastoidleri çipten 96 delikli U tabanlı plakalara aktarın. - Blastoidleri kuyucuk başına 100 μL blokaj çözeltisinde (% 0.3 Triton-X 100 ve% 10 normal eşek serumu içeren PBS) 60 dakika boyunca geçirgen hale getirin ve bloke edin.
NOT: Antikorların konakçı türüne bağlı olarak, serumu buna göre uyarlayın. - Engelleme çözümünü kaldırın. Taze bloke edici çözelti içinde seyreltilmiş 100 μL birincil antikorlar ekleyin ve numuneleri gece boyunca 4 ° C'de inkübe edin.
NOT: Birincil antikorların konsantrasyonları, üreticinin talimatlarına göre belirlenmelidir. - Numuneleri PBS'de (yıkama çözeltisi) %0,1 Triton-X 100 ile 3 kat 10 dakika boyunca yıkayın. 20 μg / mL Hoechst nükleer boyası ile birlikte bloke edici çözeltiye 100 μL ikincil antikorlar ekleyin ve numuneleri oda sıcaklığında 1 saat inkübe edin. Numuneleri ışıktan koruyun.
NOT: İkincil antikorların konsantrasyonları, üreticinin talimatlarına göre belirlenmelidir. - Numuneleri 10 dakika boyunca yıkama çözeltisi ile 3 kat yıkayın. Görüntüleme için, numuneleri PBS'de cam tabana μ kaydırağa aktarın.
NOT: Montaj ortamı, görüntüleme için kullanılan hedefe göre seçilmelidir. Örneğin, PBS'deki% 80 gliserol, yağ hedeflerini kullanırken numuneleri monte etmek için kullanmak mümkündür.
Sonuçlar
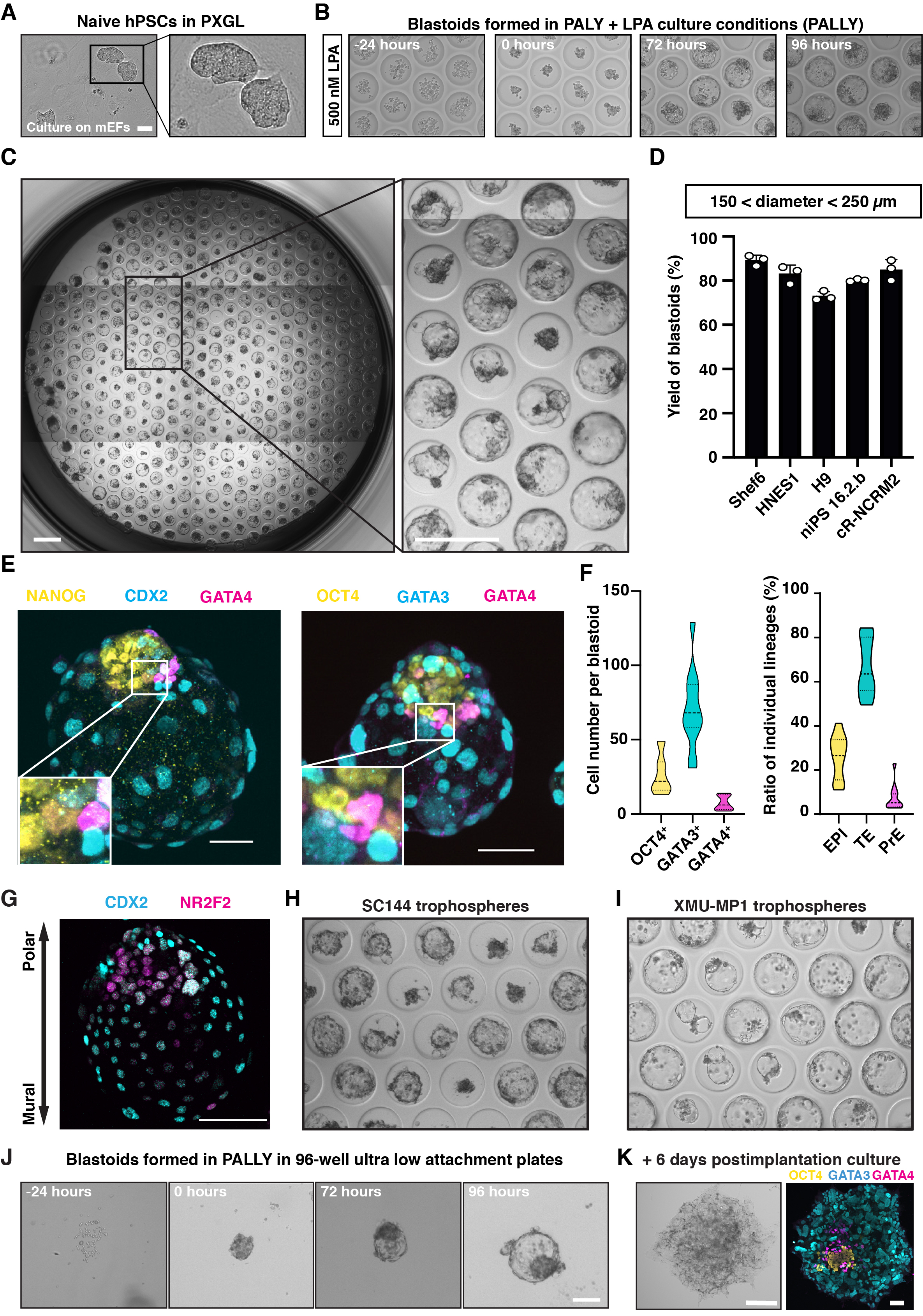
Tipik olarak, PXGL'de kültürlenen naif hPSC'ler (Şekil 2A), PALLY indüksiyonundan 48 ila 72 saat sonra ortaya çıkan ve 96 saat içinde 150-250 μm çapa ulaşan agrega ve kavitasyonlu yapılardır (Şekil 2B). Optimal (1) tohumlama hücre sayıları, (2) N2B27 ile kültür öncesi agregasyon süresi (0 ila 24 saat), (3) bireysel kimyasal bileşenlerin (özellikle LPA) konsantrasyonu ve (4) PALLY işleminin süresi kullanılarak, indüksiyon verimliliği, morfometrik parametrelere (toplam 150-250 μm boyutu, tek düzenli boşluk, tek iç hücre kümesi; Şekil 2C,D) ve üç soyun varlığı. Optimal olmayan bir başlangıç hücre durumu ve / veya indüksiyon koşulları, daha az verimli veya hiç blastoid oluşumuna neden olmaz. Maksimum verimlilik sağlamak ve sadece implantasyon öncesi analoglar oluşturmak için, naif PXGL hPSC'lerin yüksek kaliteli bir kültürünü kullanmak çok önemlidir. Bu, FACS tarafından yüzey belirteçleri SUSD2 (naif durum) ve CD24 (astarlanmış durum) için pozitif hücrelerin yüzdesi ölçülerek değerlendirilebilir. Hedef dışı ekstraembriyonik soylara (örneğin, amniyon, ekstraembriyonik mezoderm) özgü ek yüzey belirteçleri de yararlı olacaktır, ancak bildiğimiz kadarıyla şu anda mevcut değildir. Elde edilen oluşum verimliliği bildirilen sonuçlardan düşükse, blastoid ortamın tüm bileşenlerini, özellikle PBS'de yeniden yapılandırılmış LPA'yı dikkatlice kontrol etmek önemlidir ve bir GPCR ligandı olarak, DMSO'da yeniden oluşturulan sentetik moleküllere kıyasla daha kararsız olabilir. Çoğu durumda, verim maksimum olmasa bile, kavitasyonlu yapılar hala üç blastosist soyundan oluşur. Üç blastosist soyunun ortaya çıkışı ve embriyonik-abembriyonik eksenin oluşumu, belirteçlerin immünofloresan boyanması ile doğrulanabilir (EPI: NANOG, OCT4, TE: GATA3, Polar-TE NR2F2, Mural-TE: CDX2, PrE: GATA4; Şekil 2E,G). Sadece TE'den oluşan trofosferler, hücreler arası iletişimin rolünü daha da incelemeye yardımcı olur. Trofosferler indüksiyondan sonraki 96 saat içinde %50-%60 verimlilikte oluşabilir (Şekil 2H,I). Blastoid oluşumu sadece ev yapımı mikrowell dizilerinde değil, aynı zamanda indüksiyon koşullarının optimizasyonu ile ticari olarak temin edilebilen ultra düşük ataşman 96 kuyu plakalarında da gerçekleştirilebilir (bkz. Protokol ve Şekil 2J). Blastoidler ayrıca, in vitro farklılaşma protokolü ile 13. gün embriyosunun zaman eşdeğeri olan 6 gün daha fazla gelişme kapasitesine sahiptir (Şekil 2K).
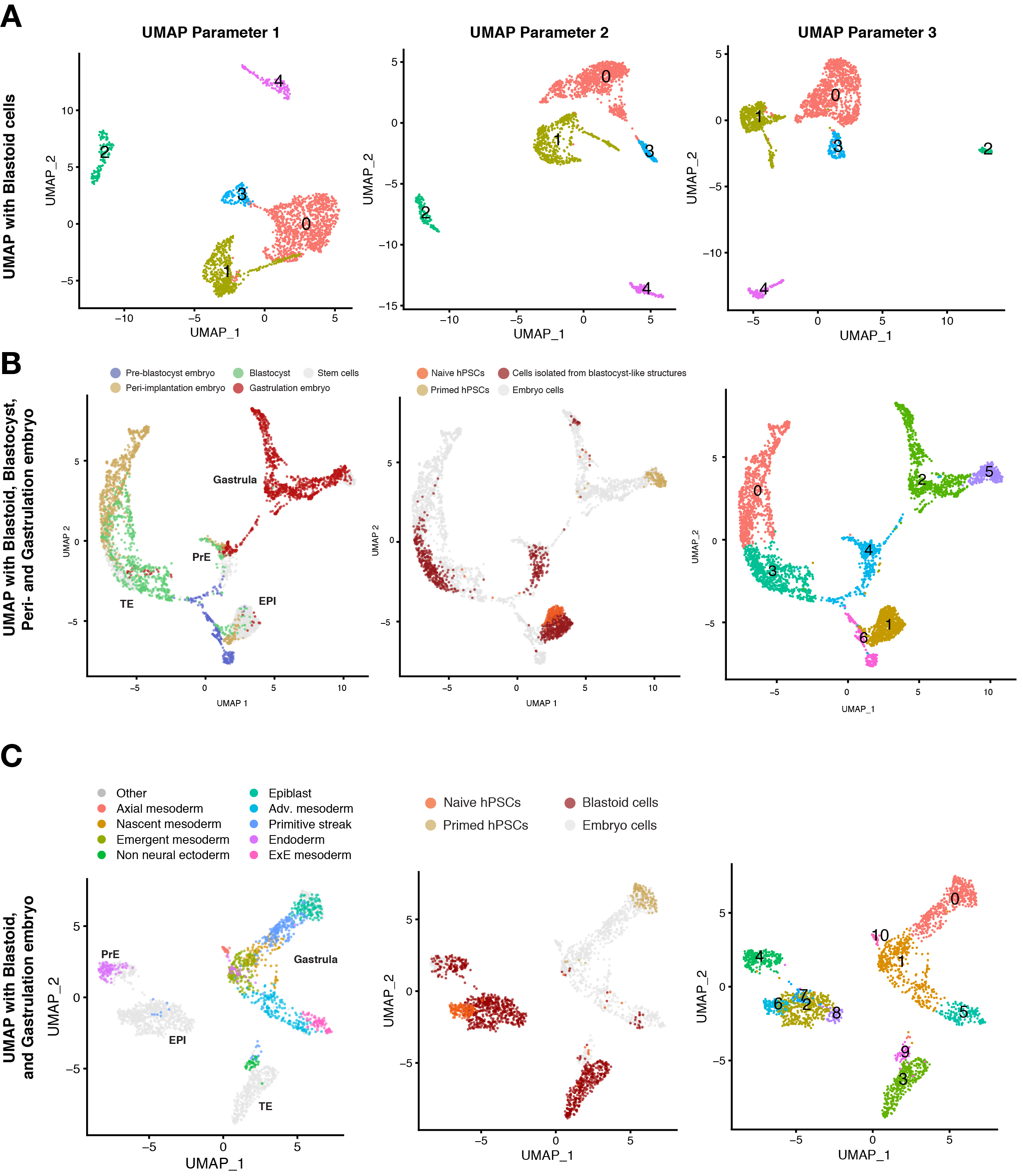
Blastoid hücrelerin hücre durumunu daha da karakterize etmek için, tek hücreli RNA dizileme teknolojisi kullanılmalıdır. UMAP, hücre durumlarının dağılımını görselleştirmek için yaygın olarak uygulanır ve tek tek hücre durumlarının yakınlığını değerlendirmek için denetimsiz kümeleme analizi yapılır. Tek hücreli veri analizindeki farklı parametreler, hücrelerin UMAP'lerde nasıl görüntülendiğini etkileyebilir, böylece farklı uzamsal ve göreli konumlara ve şekillere sahip kümelere yol açabilir (Şekil 3A). Bununla birlikte, bu analizde, hücreler, kümelemeyi gerçekleştirmek ve verilerin görselleştirilmesini gerçekleştirmek için kullanılan parametrelerden bağımsız olarak, üç blastosist soyunun yüksek güvenle ayırt edilmesini sağlayan belirgin şekilde tekrarlanabilir şekilde farklı kümeleme profilleri görüntüler. Farklı gelişim aşamalarında toplanan embriyolardan elde edilen hücreleri referans olarak kullandık. Bu veri kümelerinin birleştirilmesi, blastoidden gelen trofektoderm analoğunun çoğunluğunun implantasyon öncesi trofektoderm ile kümelendiğini, ancak implantasyon sonrası trofoblastlarla kümelenmediğini göstermektedir (Şekil 3B). Bu sonuçlar bağımsız bir konsorsiyum10 tarafından da doğrulandı.
Carnegie evre 7 (CS7) gastrulatan embriyolar referans haritasına dahil edildiğinde, küçük bir blastoid hücre popülasyonu (%3) bu embriyoların mezoderm ve amniyon soyları ile kümelenmiştir (Şekil 3C). Amniyon benzeri hücreler referans haritasına dahil edildiğinde, küçük bir blastoid hücre popülasyonu (<% 2), bu tür amniyon benzeri hücrelerle kümelenir.
Genel olarak, sadece tek bir düzenli boşluk, tek bir iç hücre kümesi, 150-250 μm arasında değişen toplam büyüklükte, üç blastosist soyunun transkriptomik analoglarını içeren ve büyük ölçüde diğer soylardan (örneğin, amniyon, mezoderm, ekstraembriyonik mezoderm) yoksun olan yapılar insan blastoidleri olarak kabul edilir.

Şekil 1: Yüksek doğrulukta blastoidler oluşturmak için dört özellik ve iki yaklaşım. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: Naif hPSC agregalarından türetilen blastoid ve trofosferler. (A) Işınlanmış MEF ile birlikte kültürlenmiş PXGL ortamında kültürlenmiş naif hPSC'leri gösteren faz kontrastlı görüntüler. Ölçek çubuğu: 50 μm. (B) 500 nM LPA (PALLI ortamı) ile yapışkan olmayan bir hidrojel mikrowell dizisi üzerinde kültürlenen naif hPSC agregalarının morfolojik değişimini gösteren faz kontrastlı görüntüler. Ölçek çubuğu: 200 μm. (C) İnsan blastoidleri 96 saat sonra bir mikrowell dizisi üzerinde oluşmuştur. Ölçek çubukları: 400 μm. (D) Farklı naif hPSC hatlarından (n = 3 mikrowell dizileri) optimize edilmiş LPA konsantrasyonu ile PALLY kültür koşulu tarafından indüklenen bir insan blastoidi içeren mikro kuyucukların yüzdesinin miktarı. (E) İnsan blastoidlerinin epiblast (EPI) belirteçleri (sarı) NANOG ve OCT4 ile immünofloresan boyaması; TE belirteçleri (camgöbeği) CDX2 ve GATA3; ve ilkel endoderm belirteci (macenta) SOX17 ve GATA4. Ölçek çubuğu: 100 μm. (H-I) OCT4, GATA3 ve GATA4'ün immünofloresan boyamasına dayanan blastoiddeki (96 h) her bir soya ait hücre sayısının (solda) ve hücre yüzdesinin (sağda) miktarı. (G) CDX2 (camgöbeği) ve NR2F2 (macenta) için insan blastoidlerinin immünofloresan boyaması. (F) Mikrowell dizisi üzerindeki erken ve geç evre trofosferlerin faz kontrastlı görüntüleri, sırasıyla 3 μM SC144 (H) veya 2 μM XMU-MP-1 (I) ilavesiyle indüklenir. (J) 500 nM LPA (PALLI ortam) ile ultra düşük ataşman 96 kuyu plakasında kültürlenmiş naif hPSC'lerin morfolojik değişimini gösteren faz kontrastlı görüntüler. (K) Postimplantasyon kültür durumunda 6 gün boyunca yetiştirilen blastoidde OCT4 (sarı), GATA3 (camgöbeği) ve GATA4 (macenta) için faz kontrastlı görüntü (solda) ve immünofloresan boyama (sağda). Ölçek çubuğu: 100 μm. Bu rakam 6,10'dan uyarlanmıştır. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Blastoid bileşiminin tek hücre dizilimi ile karakterizasyonu. (A) Blastoidin (24 saat, 60 saat, 96 saat), naif hPSC'lerin, astarlanmış hPSC'lerin ve hTSC'nin (implantasyon sonrası sitotrofoblastı temsil eder) farklı zaman noktalarından türetilen tek hücrelerin transkriptomunun UMAP'inde farklı parametrelerle denetimsiz kümeleme analizi. (B) Blastoid (96 saat), naif hPSC'ler ve astarlanmış hPSC'lerden türetilen hücrelerin transkriptomunun, preimplantasyon, peri-implantasyon (in vitro kültürlü blastosistler) ve gastrulasyon (Carnegie evre 7, yani E16-19 arasında) aşamalarının insan embriyolarından yayınlanmış veri setleriyle entegre edilmiş transkriptomun UMAP'si. Bireysel hücreler, insan embriyolarındaki (solda), blastoid türevli hücrelerdeki veya kök hücrelerdeki (orta) kökenlerine ve denetimsiz kümeleme analizinin sonucuna (sağda) göre renklendirilir. (C) blastoidlerden, naif hPSC'lerden, astarlanmış hPSC'lerden türetilen ve gastrulasyondan (Carnegie evre 7, yani E16-19 arasında) evre embriyosundan yayınlanmış veri seti ile entegre edilmiş hücrelerin transkriptomunun UMAP'leri. Bireysel hücreler, insan embriyolarındaki (solda), blastoid türevli hücrelerdeki veya kök hücrelerdeki (orta) kökenlerine ve denetimsiz kümeleme analizinin sonucuna (sağda) göre renklendirilir. Bu rakam6'dan uyarlanmıştır. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
Ek Tablo 1: Bu çalışmada kullanılan tüm medya kompozisyonu. Bu tabloyu indirmek için lütfen tıklayınız.
Tartışmalar
Bu çalışmada, basit ve sağlam bir protokol kullanarak yüksek verimliliğe sahip insan blastoidlerinin nasıl kurulacağını adım adım gösteriyoruz. Naif PXGL hPSC'lerin toplanması ve üçlü inhibisyonu üzerine, blastoidler verimli bir şekilde (>% 70) oluşur ve 4 gün içinde sırayla 3 blastosist analogu üretir. Blastoidlerin verimliliğindeki ve kalitesindeki sınırlamalar (örneğin, hedef dışı hücrelerin varlığı), başlangıç durumu optimalin altındaysa ortaya çıkabilir. Not olarak, PXGL hPSC'lerin implantasyon sonrası aşamaları yansıtan hücrelerin yaklaşık% 5'ini içerdiğini ölçtük. Bu hücreler yüksek kaliteli blastoidlerin oluşumunu sınırlayabilir. Blastosist epiblastını yansıtan ilk naif PXGL durumunun ötesinde, bir başka önemli faktör blastoid oluşumu için kullanılan ortamdır. Blastosist benzeri hücreleri hızla oluşturmak ve hedef dışı, implantasyon sonrası benzeri hücrelerin oluşumunu önlemek için, üçlü yolak inhibisyonunun (Hippo, ERK, TGF-β) gerekli olduğunu öneriyoruz. Farklı hücre hatları, ERK / TGF-β inhibisyonu üzerine farklı blastoid verimleri verirken (genellikle% 10 -% 20 civarında), LPA'ya maruz kalmak, katı morfometrik ve soy spesifikasyon kriterleri kullanırken, tüm hücre hatlarında eşit derecede yüksek blastoid verimi oluşumuna neden olur. LPA muhtemelen fare ve insan 8,51'deki epiblast ve trofektoderm soyları arasındaki ilk soy ayrımında kritik bir rol oynayan Hippo yolunun inhibisyonu üzerinde hareket eder. LPA ile blastoid verimliliğinin önemli ölçüde iyileştirilmesi, blastosistte rol oynayan Hippo yolu aracılı iç-dış hücre spesifikasyon mekanizmalarının blastoid oluşumu sırasında birlikte seçildiğini göstermektedir. Mevcut bir sınırlama, insan blastosisti veya blastoidleri kültürlemek için kullanılan protokollerin 7-13. günde (blastosist / blastoid oluşumundan sonra) yetersiz bir optimalliği nedeniyle, implantasyon sonrası gelişimi ne ölçüde uygun şekilde modelleyebileceğimizi değerlendiremediğimiz gerçeğinde yatmaktadır.
Blastoid hücrelerin transkriptomik durumunu analiz etmek, scRNAseq, yeterli referans haritaları ve biyoinformatik yöntemler kullanılarak kolayca elde edilebilir. Daha önce, transkriptomik analiz, PXGL'de kültürlenen hPSC'lerin, astarlanmış duruma kıyasla blastosist epiblastına daha benzer olduğunu göstermiştir. Verilerin analizindeki sınırlamalar, referans haritası yalnızca blastosist evre hücrelerini içeriyorsa ortaya çıkabilir. Referans haritası, potansiyel hedef dışı hücrelerin varlığını değerlendirmek için implantasyon sonrası embriyolardan kaynaklanan hücreleri içermelidir. Gelecekte, blastoid hücreleri karşılaştırmak için, implantasyon öncesi ve sonrası insan conceptusunun tüm dokularını içeren bir referans haritası son derece değerli olacaktır. Ek olarak, örneğin transkriptom, kromatin erişilebilirliği ve DNA metilasyonu dahil olmak üzere multi-omik tek hücreli referans haritaları daha da yardımcı olacaktır. Son olarak, embriyo modellerinden ve referans concepti'den hücreler arasındaki benzerlikleri nicel olarak değerlendirmek ve hedef dışı hücreleri pozitif olarak tanımlamak için standartlaştırılmış biyoinformatik yöntemler, sonuçları tarafsız bir şekilde analiz etmeye ve karşılaştırmaya yardımcı olacaktır.
Hippo, TGF-β ve ERK yolaklarının üçlü inhibisyonu ile oluşan blastoidler toplamda, 1) yüksek verimli morfogenez, 2) soy spesifikasyonunun doğru dizisi, 3) transkriptom seviyesinde blastosist benzeri hücrelerin yüksek saflığı, 4) peri-implantasyon gelişimini modelleme kapasitesinin dört özelliğine sahiptir. Blastoidlerin bu özellikleri, blastosist gelişimi ve implantasyonu üzerine hipotezler oluşturmayı kolaylaştıracaktır, ancak embriyonik gelişimin erken aşamalarını özetlemezler. İnsan blastosistinin sınırlı erişilebilirliği ve çok yönlülüğünün aksine, blastoidler, blastosist gelişimi ve implantasyonunun fonksiyonel araştırmaları için genetik ve ilaç taramalarına uygundur. Gelecekte, bu tür temel bilgiler IVF medya formülasyonunun iyileştirilmesine, döllenme sonrası kontraseptiflerin geliştirilmesine ve erken gebeliğin daha iyi yönetilmesine katkıda bulunabilir.
Açıklamalar
Avusturya Bilimler Akademisi, Moleküler Biyoteknoloji Enstitüsü, insan blastoid oluşumu ve blastoid-endometriyum etkileşim testi için protokolleri açıklayan EP21151455.9 patent başvurusunda bulundu. HK, AJ, HHK ve NR bu patentin mucitleridir. Diğer tüm yazarlar rakip çıkarlar olmadığını beyan eder.
Teşekkürler
Bu proje, Avrupa Birliği'nin Horizon 2020 araştırma ve inovasyon programı (ERC-Co hibe anlaşması No.101002317 'BLASTOID: erken insan embriyogenezi için bir keşif platformu') kapsamında Avrupa Araştırma Konseyi'nden (ERC) finansman almıştır. H.H.K., Avusturya Bilim Fonu (FWF), Lise Meitner Programı M3131-B tarafından desteklenmektedir. H9 ve H9-GFP hücre hatlarını paylaştığı için Yasuhiro Takashima'ya ve HNES1, Shef6, niPSC 16.2b ve cR-NCRM2 hücre hatlarını paylaştığı için Austin Smith, Peter Andrews ve Ge Guo'ya teşekkür ederiz. Hossein Baharvand'a endometriyal organoidleri paylaştığı için teşekkür ederiz. Joshua M. Brickman'a, PrE farklılaştırılmış hücrelerden ve nEND hücrelerinden izole edilen RNA'yı paylaştığı için teşekkür ederiz. Shankar Srinivas'a, peri-gastrulasyon embriyosunun tek hücreli RNA dizileme verilerini paylaştığı için teşekkür ederiz. Aleksand Bykov ve Luisa Cochella'ya SMARTSeq2 kütüphanesinin hazırlanmasında teknik yardım için teşekkür ederiz. IMBA'daki NGS, Biyooptik ve Kök Hücre tesisine kritik yardımları için teşekkür ederiz.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Neurobasal media | in house | ||
| DMEM/F12 | in house | ||
| 100X N2 supplemen | Gibco | 17502048 | |
| 50X B27 supplement | Gibco | 17504044 | |
| 100X Glutamax | Gibco | 35050038 | |
| 100 mM Sodium Pyruvate | Gibco | 11360039 | |
| MEM-Non-essential amino acids | Gibco | 11140050 | |
| 1 M Hepes | in house | ||
| 50 mM 2-Mercaptoethanol | Thermofisher | 31350010 | |
| 100X Penicillin-Streptomycin | Sigma-Aldrich | P0781 | |
| Bovine Serum Albumin solution | Sigma-Aldrich | A7979 | |
| PD0325901 | Medchem express | HY-10254 | |
| XAV-939 | Medchem express | HY-15147 | |
| Gö 6983 | Medchem express | HY-13689 | |
| Human recombinant Leukemia Inhibitory Factor | in house | ||
| A83-01 | Medchem express | HY-10432 | |
| 1-Oleoyl Lysophosphatidic acid (LPA) | Peprotech | 2256236 | |
| Y-27632 | Medchem express | HY-10583 | |
| CMRL medium | Gibco | 21530027 | |
| Fetal Bovine Serum (FBS) | Sigma-Aldrich | F7524 | |
| KnockOut Serum Replacement (KSR) | Thermofisher | 10-828-028 | |
| Accutase | Biozym | B423201 | cell detachment solution |
| Geltrex | Thermofisher | A1413302 | growth factor basement membrane extract |
| TROP2 antibody | R&D systems | MAB650 | |
| PDGFRα antibody | R&D systems | AF307 | |
| SC-144 | Axon | 2324 | |
| XMU-MP-1 | Med Chem Express | HY-100526 | |
| Matrigel | basement membrane matrix | ||
| Countess cell counting chamber slides | Thermo fisher | cell counting slides | |
| DAPI Staining Solution | Miltenyi Biotec | 130-111-570 |
Referanslar
- Rivron, N., et al. Debate ethics of embryo models from stem cells. Nature. 564 (7735), 183-185 (2018).
- Hyun, I., Munsie, M., Pera, M. F., Rivron, N. C., Rossant, J. Toward Guidelines for Research on Human Embryo Models Formed from Stem Cells. Stem Cell Reports. 14 (2), 169-174 (2020).
- Clark, A. T., et al. Human embryo research, stem cell-derived embryo models and in vitro gametogenesis: Considerations leading to the revised ISSCR guidelines. Stem Cell Reports. 16 (6), 1416-1424 (2021).
- Lovell-Badge, R., et al. ISSCR Guidelines for Stem Cell Research and Clinical Translation: The 2021 update. Stem Cell Reports. 16 (6), 1398-1408 (2021).
- Rivron, N. C., et al. Blastocyst-like structures generated solely from stem cells. Nature. 557 (7703), 106-111 (2018).
- Kagawa, H., et al. Human blastoids model blastocyst development and implantation. Nature. , 04267-04268 (2021).
- Meistermann, D., et al. Integrated pseudotime analysis of human pre-implantation embryo single-cell transcriptomes reveals the dynamics of lineage specification. Cell Stem Cell. 28 (9), 1625-1640 (2021).
- Gerri, C., et al. Initiation of a conserved trophectoderm program in human, cow and mouse embryos. Nature. 587 (7834), 443-447 (2020).
- Gerri, C., Menchero, S., Mahadevaiah, S. K., Turner, J. M. A., Niakan, K. K. Human Embryogenesis: A Comparative Perspective. Annual Review of Cell and Developmental Biology. 36, 411-440 (2020).
- Zhao, C., et al. Reprogrammed iBlastoids contain amnion-like cells but not trophectoderm. bioRxiv. , 2021.05.07.442980 (2021).
- Zijlmans, D. W. Integrated multi-omics reveal polycomb repressive complex 2 restricts human trophoblast induction. Nat. Cell Biol. 24, 858-871 (2022).
- Nichols, J., Smith, A. Naive and primed pluripotent states. Cell Stem Cell. 4 (6), 487-492 (2009).
- Guo, G., et al. Epigenetic resetting of human pluripotency. Development. 144 (15), 2748-2763 (2017).
- Takashima, Y., et al. Resetting transcription factor control circuitry toward ground-state pluripotency in human. Cell. 158 (6), 1254-1269 (2014).
- Thomson, J. A. Embryonic stem cell lines derived from human blastocysts. Science. 282 (5391), 1145-1147 (1998).
- Yang, Y., et al. Derivation of Pluripotent Stem Cells with In Vivo Embryonic and Extraembryonic Potency. Cell. 169 (2), 243-257 (2017).
- Stirparo, G. G., et al. Integrated analysis of single-cell embryo data yields a unified transcriptome signature for the human pre-implantation epiblast. Development. 145 (3), 158501 (2018).
- Castel, G., et al. Induction of Human Trophoblast Stem Cells from Somatic Cells and Pluripotent Stem Cells. Cell Reports. 33 (8), 108419 (2020).
- Posfai, E., et al. Evaluating totipotency using criteria of increasing stringency. Nature Cell Biology. 23 (1), 49-60 (2021).
- Nakamura, T., et al. A developmental coordinate of pluripotency among mice, monkeys and humans. Nature. 537 (7618), 57-62 (2016).
- Theunissen, T. W., et al. Molecular Criteria for Defining the Naive Human Pluripotent State. Cell Stem Cell. 19 (4), 502-515 (2016).
- Guo, G., et al. Naive Pluripotent Stem Cells Derived Directly from Isolated Cells of the Human Inner Cell Mass. Stem Cell Reports. 6 (4), 437-446 (2016).
- Rossant, J. Genetic Control of Early Cell Lineages in the Mammalian Embryo. Annual Review of Genetics. 52, 185-201 (2018).
- De Paepe, C., et al. Human trophectoderm cells are not yet committed. Human reproduction. 28 (3), 740-749 (2013).
- Amita, M., et al. Complete and unidirectional conversion of human embryonic stem cells to trophoblast by BMP4. Proceedings of the National Academy of Sciences of the United States of America. 110 (13), 1212-1221 (2013).
- Io, S., et al. Capturing human trophoblast development with naive pluripotent stem cells in vitro. Cell Stem Cell. 28 (6), 1023-1039 (2021).
- Guo, G., et al. Human naive epiblast cells possess unrestricted lineage potential. Cell Stem Cell. 28 (6), 1040-1056 (2021).
- Liu, X., et al. Modelling human blastocysts by reprogramming fibroblasts into iBlastoids. Nature. 591 (7851), 627-632 (2021).
- Hirate, Y., et al. Polarity-dependent distribution of angiomotin localizes Hippo signaling in preimplantation embryos. Current biology: CB. 23 (13), 1181-1194 (2013).
- Cockburn, K., Biechele, S., Garner, J., Rossant, J. The Hippo pathway member Nf2 is required for inner cell mass specification. Current Biology: CB. 23 (13), 1195-1201 (2013).
- Xu, R. H., et al. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nature Biotechnology. 20 (12), 1261-1264 (2002).
- Krendl, C., et al. GATA2/3-TFAP2A/C transcription factor network couples human pluripotent stem cell differentiation to trophectoderm with repression of pluripotency. Proceedings of the National Academy of Sciences of the United States of America. 114 (45), 9579-9588 (2017).
- Xu, R. H., et al. Basic FGF and suppression of BMP signaling sustain undifferentiated proliferation of human ES cells. Nature Methods. 2 (3), 185-190 (2005).
- Horii, M., Bui, T., Touma, O., Cho, H. Y., Parast, M. M. An Improved Two-Step Protocol for Trophoblast Differentiation of Human Pluripotent Stem Cells. Current Protocols in Stem Cell Biology. 50 (1), 96 (2019).
- Okae, H., et al. Derivation of Human Trophoblast Stem Cells. Cell Stem Cell. 22 (1), 50-63 (2018).
- Yamanaka, Y., Lanner, F., Rossant, J. FGF signal-dependent segregation of primitive endoderm and epiblast in the mouse blastocyst. Development. 137 (5), 715-724 (2010).
- Kuijk, E. W., et al. The roles of FGF and MAP kinase signaling in the segregation of the epiblast and hypoblast cell lineages in bovine and human embryos. Development. 139 (5), 871-882 (2012).
- Roode, M., et al. Human hypoblast formation is not dependent on FGF signalling. Developmental Biology. 361 (2), 358-363 (2012).
- Linneberg-Agerholm, M., et al. Naïve human pluripotent stem cells respond to Wnt, Nodal and LIF signalling to produce expandable naïve extra-embryonic endoderm. Development. 146 (24), (2019).
- Yu, L., et al. Blastocyst-like structures generated from human pluripotent stem cells. Nature. 591 (7851), 620-626 (2021).
- Yanagida, A., et al. Naive stem cell blastocyst model captures human embryo lineage segregation. Cell Stem Cell. 28 (6), 1016-1022 (2021).
- Zappia, L., Oshlack, A. Clustering trees: a visualization for evaluating clusterings at multiple resolutions. GigaScience. 7 (7), (2018).
- Subramanian, A., et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proceedings of the National Academy of Sciences of the United States of America. 102 (43), 15545-15550 (2005).
- Messmer, T., et al. Transcriptional Heterogeneity in Naive and Primed Human Pluripotent Stem Cells at Single-Cell Resolution. Cell Reports. 26 (4), 815-824 (2019).
- Zhou, F., et al. Reconstituting the transcriptome and DNA methylome landscapes of human implantation. Nature. 572 (7771), 660-664 (2019).
- Petropoulos, S., et al. Single-Cell RNA-Seq Reveals Lineage and X Chromosome Dynamics in Human Preimplantation Embryos. Cell. 167 (1), 285 (2016).
- Tyser, R. C. V., et al. A spatially resolved single cell atlas of human gastrulation. bioRxiv. , (2020).
- Turco, M. Y., et al. hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nature Cell Biology. 19 (5), 568-577 (2017).
- Boretto, M., et al. Development of organoids from mouse and human endometrium showing endometrial epithelium physiology and long-term expandability. Development. 144 (10), 1775-1786 (2017).
- Ma, H., et al. In vitro culture of cynomolgus monkey embryos beyond early gastrulation. Science. 366 (6467), (2019).
- Nishioka, N., et al. The Hippo signaling pathway components Lats and Yap pattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass. Developmental Cell. 16 (3), 398-410 (2009).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır