
Method Article
Endokardiyal ve Koroner Endotel Hücrelerinin Sıçan Kalbinin Ventriküler Serbest Duvarından İzolasyon
Bu Makalede
Özet
Biz bir sindirim tampon ardışık doku sindirim yoluyla sıçan kalplerinden endokardiyal ve koroner endotel hücrelerinin izolasyon için bir protokol sunmak, tekrarlayan santrifüj döngüleri hücre toplama, ve anti-sıçan CD31 mikroboncuklar kullanarak hücre arıtma.
Özet
Endokardiyal endotel hücrelerinin (EECs) ve koroner endotel hücrelerinin (MK) kökeni, gelişimi, belirteçleri ve fonksiyonları arasında farklılık gösterdiği gösterilmiştir. Sonuç olarak, bu iki hücre popülasyonları kalp hastalıklarında benzersiz rol oynamaktadır. İzole endotel hücrelerini içeren güncel çalışmalar, hem EEC'lerden hem de MED'lerden oluşan hücre popülasyonlarını araştırmaktedir. Bu protokol, hücreye özgü karakterizasyonu için bu iki hücre popülasyonunu bağımsız olarak yalıtmak için bir yöntem özetler. Sol ve sağ ventrikül süzme duvarın toplanmasının ardından, dış yüzeyve iç yüzeyden gelen endotel hücreleri sindirim tampon çözeltisi kullanılarak ayrı ayrı serbest bırakılır. Dış yüzeyin ardışık sindirimi ve iç endokardiyal tabaka iki endotel hücre popülasyonunun ayrılmasını korumuştur. AÇK'ların ve MDC'lerin ayrı ayrı yalıtımı, her popülasyona özgü işaretçilerin tanımlanması yla daha da doğrulanır. Fare kalbinde daha önce yayınlanan tek hücreli RNA profilleme dayanarak, Npr3, Hapln1, ve Cdh11 gen ekspresyonu EECs benzersizdir; Fabp4iken , Mgll, ve Cd36 gen ekspresyonu CECs benzersizdir. qPCR verileri, bu karakteristik belirteçlerin kendi örneklerinde zenginleştirilmiş ifadesini ortaya çıkardı, başarılı EEC ve CEC izolasyonunun yanı sıra hücre fenotipinin bakımını göstererek hücreye özgü fonksiyonel analizin daha da artmasını sağladı.
Giriş
Bu makalede, sıçan kalplerinden endokardiyal endotelyal hücrelerin (EECs) ve koroner endotel hücrelerinin (MK) diseksiyonu ve sonraki izolasyonu için ayrıntılı bir protokol (Gladka ve ark.1'denmodifiye) verilmektedir. Bu hücre popülasyonlarını bağımsız olarak araştırma yeteneği, potansiyel tedavi hedefleri olarak hizmet verebilecek çeşitli kalp hastalıklarının altında yatan hücre tipine özgü mekanizmaların araştırılmasını sağlayacaktır. Ancak, bu hücre popülasyonlarının bağımsız olarak toplanması için başarılı bir yöntem henüz yayınlanmamıştır.
Cecs kökeni açısından AÇCs farklıdır, belirteçleri, ve kalp gelişimi ve hastalık sırasında fonksiyonları1,2,3,4,5,6,7., EEC'ler kardiyak mezoderm3ventral yüzeyinden kaynaklanmaktadır. Onlar VEGF ve HIF sinyalyanıt olarak Flk1+ progenitor hücrelerinden ortaya çıkar ve gelişmekte olan kalbin üç ayrık bölgelerin en iç tabakasını oluşturur: atriyum, ventrikül, ve sinüs venöz3,6. Genetik soy takibi sinüs venöz pluripotent endokardiyal hücrelerin subepikardiyal tabaka oluşturmak için göç venöz hücreleri türetmek düşündürmektedir3. Daha sonra, subepikardiyal tabaka koroner arterler ve damarlar içine farklılaşır, CECs de dahil olmak üzere, hangi periferik ventriküler serbest duvar boyunca kalır3,4. Endotel yolu bu endokardiyal VEGFC, ELA / APJ ve SOX17 sinyal3,4,,6,8,9tarafından düzenlenir. Ventriküler endokardiyum bilinmeyen birmekanizma3 tarafından interventriküler septum daha az MDCs türetilmiştir . Daha sonra, EECs ve CECs arasında lokalize farklılaşma mglldahil olmak üzere bu iki hücre popülasyonları, özel belirteçleri tarafından önerilmektedir, Fabp4, ve CD36 ifade MDCs, veya Npr3, Cdh11, ve AçCs Hapln1 ifade3,5,10.
EEC'ler ve MCD'ler kardiyak fonksiyonda farklı roller oynarlar. Mezenkimal geçiş için endokardiyal, kapak oluşumu, oda olgunlaşması, çıkış yolu regülasyonu ve atriyoventriküler kanal gelişimi EECs6'yabağlıdır. Alternatif olarak, CECs vazomotor sesi ve koroner arterlerin iltihabı katkıda11. Fonksiyondaki bu varyanslar hastalık gelişiminde bireyselleştirilmiş rollere nedenolur 4,12. Örneğin, kanıtlar, arızalı EEC'lerin konjenital kapak hastalığına yol açabileceğini düşündürmektedir6,nonkompansyon olmayan miyokardiyum6, atriyoventriküler septal etki6, endokardiyal fibroelastoz13, hipoplastik sol kalp sendromu13, ventriküler hipoplazi13, ve kardiyak hipertrofi12. Benzer şekilde, çalışmalar anormal MK koroner arter hastalığı katkıda bulduk14 ve tromboz11.
Hem araştırma hem de klinik ortamlarda kullanılabilen bu iki hücre popülasyonu hakkında kapsamlı bilgi sahibi olmak için EDC ve MED'lerin başarılı bir şekilde izole edilmesi gerekmektedir. Bu hücre popülasyonlarının büyüme ve farklılaşma faktörlerinin belirlenmesi, endotel alt tiplerinin indüklenen pluripotent kök hücrelerden (iPSC) farklılaşmasıiçin bir referans sağlayacaktır. Ayrıca, AÇK'ların ve CeC'lerin gelişimi, düzenlenmesi ve işlevindeki varyansların tam olarak tanımlanması, hücre tipine özgü bir şekilde çok sayıda kalp hastalığından sorumlu genomik ve epigenomik faktörlerin anlaşılması için hayati önem taşımaktadır. Bu makalede, ECO'ların ve MSK'ların bağımsız olarak başarılı bir şekilde toplanması için adımlar özetlenmiştir ve hücre tipine özgü belirteçlerin gen ifade düzeylerini değerlendirerek ayrışma kanıtı sağlar.
Protokol
Tüm hayvan prosedürleri Stanford Üniversitesi Laboratuvar Hayvan Bakımı İdari Paneli (APLAC31608) tarafından onaylanmıştır.
1. Tamponların hazırlanması
- Tablo 1'delistelenen reaktifleri kullanarak sindirim tamponu hazırlayın.
- DNase I stok çözeltisini hazırlayın: 2.000 birim/mL (1 mg/mL) stok çözeltisi için RNase içermeyen suda DNase I'i çözün. Aliquot ve stok çözeltisini -20 °C'de saklayın.
- Liberaz çözeltisini hazırlayın: 5 mg/mL'lik bir stok çözeltisi için rnase içermeyen su ile liberaz çözün. 30 dk. Aliquot için 4 °C'de bir rulo üzerinde döndürün ve stok çözeltisini -20 °C'de saklayın.
- 0,5 M ethylenediaminetetraastic asit (EDTA) ve 1x fosfat tamponlu salin (PBS) ile %10 büyükbaş serum albumin (BSA) stok çözeltisini 2 mM EDTA ve %0,5 BSA'lık son konsantrasyoniçin seyrelterek sıralama tamponunu hazırlayın.
2. Kalp koleksiyonu
NOT: Bu protokolde 50-100 g ağırlığında altı Adet Sprague Dawley sıçanı kullanıldı. Cinsiyet farkı gözlenmedi.
- Co2 ile sıçan ötenazi ve bir diseksiyon platformu üzerinde bir supine pozisyonda yerleştirin. Ekstremiteleri aşağı pin ve% 70 etanol ile hem karın ve göğüs sterilize.
- Karnı makasla açın ve bağırsakların arkasında bulunan arka vena kava kısmına 21 G iğne yerleştirin. Şırınga pistonunu geri çekin, karaciğer daha açık görünene kadar vendeki kanı geri çekin ve yeterli kan alınmasını gösteren daha fazla kan çekilememiştir.
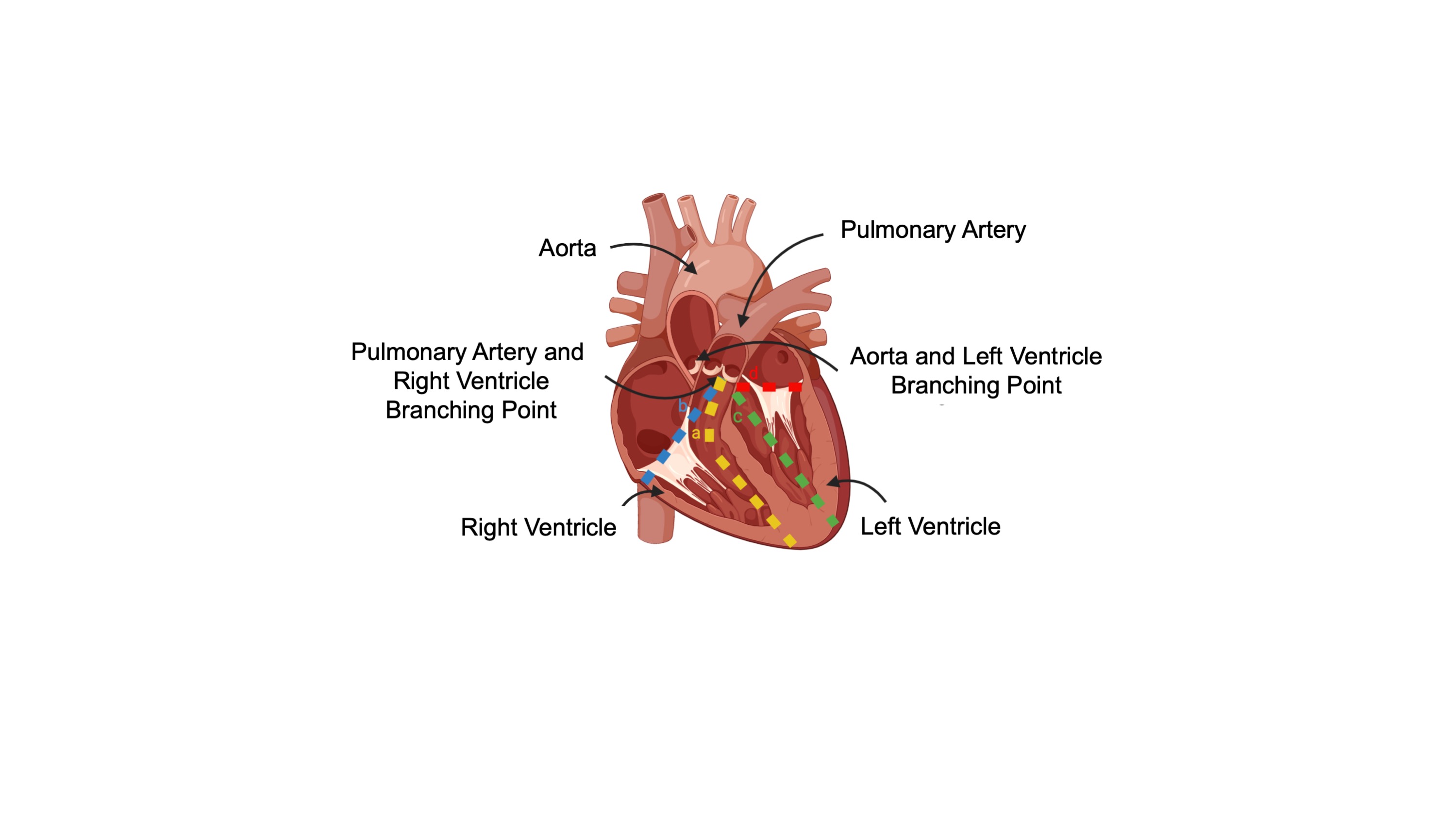
- Makas kullanarak, akciğer ve kalbe zarar vermemek için göğsü dikkatlice açın. Forseps ile aort kemer / atriyum kaldırın ve aort, pulmoner arter, pulmoner venler ve vena kava kurtarmak ve kalp kaldırmak için kesti.
- Aşırı kanı gidermek için kalbi 50 mL soğuk 1x Hanks'in dengeli tuz çözeltisi (HBSS) tampon3x ile yıkayın.
- Mikrodises mikroskobu altında, sağ ventrikülsüz duvarı, Dulbecco'nun modifiye Kartal ortamı (DMEM) içeren 5 mL'lik bir tüpe çıkarın (Şekil 1A,B).
- Onun düz, arka yüz sol ve sağ tarafı tanımlamak için kalp lay.
- Pulmoner arter bulun, venlerin ve arterlerin en ön kalbin üst dallanma ve sağ ventrikül odasına aşağı pulmoner arter ile kesti. Kalbin ön yüzünde, tepe ulaşana kadar septum boyunca kesilmiş. Pulmoner arter ve sağ ventrikül oda kavşak noktasına kadar septum boyunca kalbin arka tarafında apeks kesmeye devamedin (Şekil 1A).
- Pulmoner arter ve sağ ventrikül kavşak noktasından başlayarak, sağ ventrikül serbest duvarı kalbin geri kalanından kurtarılıncaya kadar, kalbin hem ön hem de arka tarafını bir önceki diseksiyona dik olarak ve septumdan uzaklaştırın (Şekil 1B).
- Mikrodisesmikroskop altında, sol ventrikül serbest duvarını DMEM içeren 5 mL'lik bir tüpe çıkarın (Şekil 1C,D).
- Pulmoner arter arkasında kalbin üst dallanma, aort bulun ve sol ventrikül serbest duvar kesmek için adım 2.5 açıklanan yöntemi tekrarlayın.
3. AET sindirim
- Hem sağ hem de sol ventrikülsüz duvar dokularını iç yüzeyi düz, aşağı bakacak şekilde 60 cm'lik bir kültür kabına yerleştirin(Şekil 2).
NOT: İç yüzey, Şekil 2A,C' de gösterildiği gibi hafif içbükey şekilli ventriküliç yüzüdür. - Tabağa 0,5-1 mL sindirim tamponu ekleyin ve pipetin ucunu doğrudan dokunun altına yerleştirin. Sadece iç yüzey batırılana kadar devam edin.
- Yemeği %5 CO2, 37 °C inkübatörde 5 dakika kuluçkaya yatırın.
- Sindirimi durdurmak için kültür yemeğine EC ortamı ekleyin.
NOT: SINDIRIM tamponu miktarını 1:5 oranı olarak EC ortamına saklayın. - Ventriküllerin iç yüzeyini EC ortamıile yıkamak için 1 mL'lik pipet kullanın ve akıntıyı 40 mm'lik bir süzgeçten 50 mL'lik bir toplama tüpüne aktarın(Şekil 2E). Aşağı saflaştırma için buz üzerinde koleksiyonları tutun.
4. CEC sindirim
- İç tabakadan(Şekil 2B,D)kirlenmeden sol ventrikülün dış yüzeyini kesin ve her birini sindirim tamponunun 1 mL'lik içeren ayrı bir 5 mL tüpüne yerleştirin.
NOT: Sol ventrikül kalın ventrikül olarak tanımlanabilir ve dış yüzey biraz içbükey ventrikül dış yüzü olarak belirlenebilir. - Diseksiyon makası kullanarak ventriküler duvarı küçük, 1 mm3 adet ebathalinde doğuz.
- Tüpü 37 °C'lik bir su banyosunda yaklaşık 15-20 dk. Girdap her 2-3 dakikada bir kuluçkaya yatırın.
- Pipet 4 mL EC orta 5 mL tüp içine sindirim sona erdirmek için.
- Çözeltiyi 5 mL serolojik pipetle 40 mm'lik bir süzgeçten 50 mL'lik bir tüpe aktarın(Şekil 2E). Aşağı saflaştırma için buz üzerinde koleksiyonları tutun.
5. Hücre toplama
- Tüpleri oda sıcaklığında 10 dakika (RT) için 300 x g'de santrifüj edin ve süpernatantı aspire edin.
- Pelet kırmızı görünüyorsa, 1x kırmızı kan hücresinin (RBC) 1-2 mL'lik lysing tamponu(Şekil 2F)peletini yeniden askıya alın.
NOT: RBC yoksa, adım 6'ya geçin. - Tüpleri 37 °C'lik bir su banyosunda 5 dakika kuluçkaya yatırın.
- Pipet 10 mL PBS 50 mL tüpler içine lysis durdurmak için.
- Tüpleri RT'de 10 dk için 300 x g'da santrifüj edin ve süpernatantı aspire edin.
6. AT sıralama
- 50 mL tüplere 90 μL sıralama tamponu ve 10 μL anti-CD31 PE antikor ekleyin.
- Tüpleri girdap ve sonra 4 ° C buzdolabında 10 dakika kuluçka.
- Tüpleriçine tampon sıralama 10 mL ekleyin, iyice karıştırın ve sonra RT 10 dakika için 300 x g santrifüj.
- Supernatant aspire ve sıralama tampon 80 μL ve anti-PE mikroboncuklar 20 μL pelet resuspend.
- Girdap tüpler ve 15 dakika boyunca 4 °C'de kuluçka.
- Tüplere 10 mL'lik tampon ekleyerek hücreleri yıkayın, iyice karıştırın ve rt'de 10 dakika boyunca 300 x g'de santrifüj edin.
- Supernatant aspire ve sıralama tampon 500 μL pelet resuspend.
- Süperparamanyetik küreleri içeren bir sütunu manyetik ayırıcıya yerleştirin ve 3 mL sıralama arabelleğiyle sütunu durulayın.
- Sütun "akış durdurma" sonra, pipet 500 μL hücre süspansiyon sütunlar içine ve sonra sıralama tampon ile sütunları yıkayın. Yıkama 3x, her 3 mL sıralama tampon(Şekil 2G)ile tekrarlayın.
- Kolonları ayırıcıdan çıkarın ve yeni bir 15 mL toplama tüpünün üzerine yerleştirin.
- Pipetle, sütunlara 5 mL EC orta ekleyin ve bir sütun pistonkullanarak çözümü itin.
- Toplama tüplerini RT'de 10 dk için 300 x g'da santrifüj edin ve süpernatantı aspire edin.
- Üreticinin talimatlarına uygun olarak, Bir kit kullanarak EEC ve CEC örneklerinin hücre peletinden RNA'yı ayıklayın. En az 500 ng RNA elde edilebilir.
NOT: RNA ekstraksiyonundan sonra numune -80 °C'de saklanabilir. - Üreticinin talimatlarına uyarak rastgele bir astar ve kit kullanarak cDNA almak için 500 ng RNA'yı ters yazır.
- Endokardiyal ve endotel popülasyonlarının doğrulanması için nicel PCR gerçekleştirin (Şekil 2H). Astar dizileri Tablo 2'delistelenmiştir.
- 384 qPCR plakanın kuyularına 5 μL EEC veya CEC cDNA (1 ng/μL) ekleyin. Astar sayısı başına üç kuyu olması için Yeterli kuyulara EEC veya CEC cDNA'sını ekleyin.
- 6 μL 2x qPCR SYBR yeşil arabelleği, her 10 μM astarın 0,5 μL'si ile karıştırın ve bunları her aday genine karşılık gelen tek bir kuyuya ekleyin.
- Plakayı rt'de 300 x g'da 1 dk plastik film ve santrifüj ile kapatın.
- Plakayı bir qPCR makinesine yerleştirin ve ardından programı 95 °C'de 3 dk, ardından 15 s için 95 °C'de ve ardından 55 °C'de 60 s. Sonraki iki adımı 45 devir için tekrarlayın.
Sonuçlar
AET ve CEC yalıtımı süreci Şekil 2'deaçıklanmıştır. AçS ve MED'lerin başarılı izolasyonu pan-endotel hücre belirteçlerinin yanı sıra iki alt tip popülasyona farklı olanların varlığı değerlendirilerek belirlendi. Tahmin edildiği gibi, qPCR β-actingöreli , EMC endokardiyal belirteçleri Npr3daha yüksek düzeyde ifade , Hapln1, ve Cdh11 CECs göre(Şekil 3A). Aynı şekilde, CEO'lar eecs(Şekil 3B)ile karşılaştırıldığında koroner belirteçleri Fabp4, Mgllve Cd36 yüksek düzeyde ifade . Ayrıca, hem EMC'ler hem de CECs pan-EC marker gen Cdh5ifade , CEC biraz daha yüksek düzeyde(Şekil 3C).

Şekil 1: Kalp diseksiyonu diyagramı. (A) Sağ ventrikülsüz duvarı septumdan ayırmak için yapılan ilk kesim. (B) Sağ ventrikülü tamamen serbest leştirmek için yapılan ikinci kesik. (C) İlk kesim septum sol ventrikül serbest duvar ayırmak için yapılır. (D) Sol ventrikülü tamamen serbest leştirmek için yapılan ikinci kesik. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: CeC'lerin ve EEC'lerin sindirim düzeninin diyagramı ve hücre sıralamaiçin aşağıdaki düzenleme. (A) En innermost serbest ventriküler duvar ve (B) en dışta ventriküler serbest duvar (C) sindirim tamponu batırılmış veya (D) sırasıyla sindirim tamponu sindirilir. (E) Hücre çözeltilerinin toplanması ve filtrasyonu ile(F) kırmızı kan hücresi (RBC) lysis ve (G) manyetik aktive hücre sıralama (MACS) CD31 antikor kullanarak. (h) Saflaştırılmış eK'ler qPCR kullanılarak gen ekspresyonu doğrulaması için işlenmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: MDC ve EEC'lerin izolasyonunun doğrulanması na qPCR verileri. (A) AET belirteçlerinin Gen ekspresyonu düzeyleri Npr3, Cdh11, ve Hapln1 AÇC'lerde ve MK'lerde gerçek zamanlı PCR ile ölçüldü. (B) EEC'lerde ve CEC'lerde MGLL, Fabp4ve Cd36'nın gen ekspresyonu düzeyleri gerçek zamanlı PCR ile ölçüldü. (C) Pan-EC işaretçisi Cdh5, AÇM'larda ve CEC'lerde gerçek zamanlı PCR ile ölçüldü. (her grupta n = 3). Çubuklar ortalama ± SEM. *p < 0,05, **p < 0,01 vs EEC, eşleşmemiş t-testi temsil eder. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
| Tablo 1. Sindirim Tampon (1.5ml) | |||
| Hisse Senedi Con. | Son Con. | Birim | |
| Liberase TM | 5 mg/ml | 0.5 mg/ml | 150 ul |
| Dnase I | 1 mg/ml | 20 ug/ml | 30 ul |
| HEPES | 1 M | 10 mM | 15 ul |
| DMEM | - | - | 1305 ul |
Tablo 1: Sindirim tamponu tarifi.
| Tablo 2. Astar Dizileri | ||
| Gen Adı | Ileri | Ters |
| Npr3 | TCCTTGCAAATCATGTGGCCTA | GGAATCTTCCCGCAGCTC |
| Cdh11 | GTGAATGGGACTGGGACTGG | GTAATTTCTGGGGCCCTGC |
| Hapln1 | CCAGCTAAGTGGGACTCGAAG | GGGCCATTTTCAGCTTGGATG |
| Mgll | CCCGGGGCCCAAAGAC | GAAGATGAGGGCCTTGGGTG |
| Fabp4 | AGAAGTGGGAGTTGGCTTCG | ACTCTCTGACCGGATGACGA |
| Cd36 | GCAAAACGACTGCAGGTCAA | CCCGGTCACTTGGTTTTCTGA |
| Cdh5 | CCATTGAGACAGACCCCGAC | TGTGGAACGTACTGCTGG |
| B-aktin | TCTGTGTGTGGATTGGTGGCTC | CGGACTCATCGTACTCCTGC |
Tablo 2: Astar Dizileri
Tartışmalar
MED'ler ve EEC'ler kökenleri, belirteçleri ve işlevleri farklıdır ve bu nedenle gelişim ve hastalıkta benzersiz roller oynayabilir. Endotel hücre izolasyonu için mevcut protokoller makrovasküler dokularla sınırlıdır, EEC'lerin toplanması ihmal, böylece CeC çalışma kısıtlayan- ve EEC özgü fonksiyonların. Bu bilgi, gelecekteki çalışmalar için iPSC'lerin MSK ve AEC'lere farklılaşması için bir referans sağlayacak ve çeşitli kalp hastalıkları için potansiyel terapötik hedefler için bu hücre popülasyonlarının incelenmesini kolaylaştıracağı için, bu iki popülasyonu bağımsız olarak izole etmek ve incelemek esastır. Bu yeni protokol, yetişkin sıçanların ventriküler serbest duvarının dış yüzeyinden EMC'lerin iç yüzeyden ve MED'lerin izolasyonu için bir yöntem özetlendir.
Bu protokoldeki her adımın zamanlamasını tam olarak denetlemek çok önemlidir. EEC sayısı sıçan kalp dokusunda çok sınırlı olduğundan, biz hücresel hasarı önlemek için kalbin iç tabakasının sindirim süresini en aza indirdi ve daha da önemlisi, CEC kontaminasyon. Enzimatik reaksiyonu sonlandırmak için EC orta çözeltisinin hemen eklenmesi de yüksek hücre canlılığını korumak için çok önemlidir. Hücre koleksiyonunu takip eden kırmızı pelet, RBC kontaminasyonmiktarına bağlı olarak çok sayıda RBC'nin varlığını göstermektedir, RBC'lerin bozulması için 1-2 mL RBC lysis tamponu eklenebilir. Mevcut protokolü kullanarak,105 AÇ'ı altı fare kalplerinden izole edebildik. Bu hücreler daha fazla genişleme ve karakterizasyon için bir hücre kültür çanak üzerine tohumlu olabilir.
Serbest duvar toplama için doku hazırlanırken, kalp hbss ile kalan RBC'lerin çoğunluğu kaldırmak için yıkandı. Sindirim tamponunun bileşimi endotel hücrelerinin yeterli şekilde serbest kalmasını sağlar, liberase sindirim enzimi maruz hücrelerin doku şeritler nerede, DNase I DNA'nın yapışkan kalitesi ile inhibe edilebilir hücre detachment teşvik etmek için ölü hücrelerden DNA ortadan kaldırır, HEPES tampon pH dengeler, ve DMEM daha iyi hücre bakımı için amino asitler ve vitaminler daha yüksek konsantrasyon ile değiştirilmiş bir bazal orta ve liberase etkinleştirmek için gerekli kalsiyum içerir.
İnsan15 ve fare kalp10elde edilen tek hücreli RNA dizilimi (scRNA-seq) göre, belirli AT popülasyonlarına atfedilen çeşitli belirteçler bildirilmiştir. İzole edilmiş EEC'lerin (yani, Npr3, Cdh11ve Hapln1)ve AÇC'lerin (yani Mgll, Fabp4ve Cd36)saflığını incelemek için en zengin genlerden bazılarını seçtik. Β β-Actin, Npr3, Cdh11ve Hapln1 belirteçlerine göre EMC'lerde MDC'lere göre ekspresyon artışı gösterilmiştir. Benzer şekilde, Mgll, Fabp4ve Cd36 belirteçlerinin ekspresyonu, EEC'lere göre CEC'lerde daha fazlaydı. Her örnek için benzersiz olarak ifade edilmiş işaretler, sırasıyla AÇ'lar ve CeC'ler için karakteristik işaretlerle uyumludur ve başarılı yalıtımı gösterir.
Ancak mevcut protokol, izolasyon sırasında iki AK popülasyonu arasındaki çapraz kontaminasyonu, dikkatle kontrol edilen sıralı sindirimlerle bile ekarte edemez. Bu nedenle, bazı hücre yüzey işaretleri daha fazla arıtma için uygulanabilir. Örneğin, NPR3 genellikle endokardiyum10'uetiketlerken, APJ CEC'lerin16'sınınçoğunluğunu izleyebilir. Bu iki belirteç hücre yüzeyinde ifade edildiğinden, floresan aktif hücre sıralama (FACS) ve farklı EC popülasyonlarını daha da arındırmak için kullanılan antikorlar için kullanılabilirler. Buna ek olarak, insan15 ve fare10 kalp scRNA-seq AÇC'larda Npr3 zenginleştirme teyit edebilir, ve Cd36 potansiyel OLARAK CEC arıtma için kullanılabilir.
Sonuç olarak, sunulan protokol, AÇC'lerin ve MSK'ların fare kalbinden bağımsız olarak izole edilmesinin ana hatlarını ortaya koyuyor. Hücre yalıtımı ile etkinleştirilen hücresel özelliklerin kapsamlı tanımlanması, önemli downstream uygulamaları için kullanılabilir.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Yazarlar, Dr. Lingli Wang'ı hayvan protokolüne yardımlarından ve Stanford'daki Pediatri Bölümü'ne altyapı desteği için çok teşekkür ediyor. Bu çalışma NIH/NHLBI K99 HL135258 (M.G.'ye) tarafından desteklenmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) | Gibco | 15630080 | 1M |
| 15ml Tubes | Eppendorf | 0030122151 | |
| 50ml Tubes | Nunc | 339652 | |
| 5ml Tubes | Eppendorf | 30119401 | |
| ACK Lysing Buffer | Gibco | A1049201 | |
| Anti-CD31 PE | Miltenyi Biotec | 130-115-505 | |
| Anti-PE microbeads | Miltenyi Biotec | 130-048-801 | |
| Bovine serum albumin (BSA) | Miltenyi Biotec | 130-091-376 | 10% |
| CFX384 Touch Real-Time PCR Detection System | Bio-rad | 1855485 | For qPCR |
| DNAse I | Worthington | LK 003172 | 1 mg/mL in water |
| Dulbecco's Modified Eagle Medium (DMEM) | Gibco | 10313021 | |
| Endothelial Cell Growth Medium-2 (EGM-2) | Lonza | CC-3162 | EC Medium |
| Ethylendiaminetetraacetic Acid (EDTA) | Invitrogen | 15575020 | 0.5 M |
| Hank's Balanced Salt Solution (HBSS) X1 | Gibco | 14025092 | |
| Hard-Shell 384-Well PCR Plates, thin wall, skirted, clear/white | Bio-rad | HSP3805 | For qPCR |
| High-Capacity cDNA Reverse Transcription Kit | Applied Biosystems | 4368813 | |
| Liberase TM | Sigma Aldrich | 5401119001 | 5 mg/mL |
| MACS LS Column | Miltenyi Biotec | 130-042-401 | For EC isolation |
| MACS MultiStand | Miltenyi Biotec | 130-042-302 | For EC isolation |
| Microseal 'B' PCR Plate Sealing Film, adhesive, optical | Bio-rad | MSB1001 | For qPCR |
| Narrow Pattern Forceps | F.S.T | 11002-12 | For heart harvest |
| Nylon Sterile Strainer | Falcon | 352340 | 40 mm |
| Phosphate Buffered Saline (PBS) X1 | Gibco | 1001023 | |
| PowerUp SYBR Green Master Mix | Applied Biosystems | A25780 | For qPCR |
| Quick-RNA Microprep Kit | Zymo Research | R1050 | For RNA extraction |
| S&T Forceps - SuperGrip Tips | F.S.T | 00632-11 | For heart harvest |
| Strabismus Scissors - Tungsten Carbide | F.S.T | 14574-11 | For heart harvest |
| Vannas Spring Scissors - Microserrated | F.S.T | 15007-08 | For heart harvest |
Referanslar
- Gladka, M. M., et al. Single-Cell Sequencing of the Healthy and Diseased Heart Reveals Cytoskeleton-Associated Protein 4 as a New Modulator of Fibroblasts Activation. Circulation. 138 (2), 166-180 (2018).
- Koren, C. W., Sveinbjornsson, B., Smedsrod, B. Isolation and culture of endocardial endothelial cells from Atlantic salmon (Salmo salar) and Atlantic cod (Gadus morhua). Cell and Tissue Research. 290 (1), 89-99 (1997).
- Tian, X., et al. Subepicardial endothelial cells invade the embryonic ventricle wall to form coronary arteries. Cell Research. 23 (9), 1075-1090 (2013).
- Wu, B., et al. Endocardial cells form the coronary arteries by angiogenesis through myocardial-endocardial VEGF signaling. Cell. 151 (5), 1083-1096 (2012).
- Li, Y., Lui, K. O., Zhou, B. Reassessing endothelial-to-mesenchymal transition in cardiovascular diseases. Nature Reviews in Cardiology. 15 (8), 445-456 (2018).
- Zhang, H., Lui, K. O., Zhou, B. Endocardial Cell Plasticity in Cardiac Development, Diseases and Regeneration. Circulation Research. 122 (5), 774-789 (2018).
- Yu, S. Y., et al. Isolation and characterization of human coronary artery-derived endothelial cells in vivo from patients undergoing percutaneous coronary interventions. Journal of Vascular Research. 46 (5), 487-494 (2009).
- Red-Horse, K., Ueno, H., Weissman, I. L., Krasnow, M. A. Coronary arteries form by developmental reprogramming of venous cells. Nature. 464 (7288), 549-553 (2010).
- Kamimura, T., Yamagishi, T., Nakajima, Y. Avian coronary endothelium is a mosaic of sinus venosus- and ventricle-derived endothelial cells in a region-specific manner. Development, Growth and Differentiation. 60 (2), 97-111 (2018).
- Zhang, H., et al. Endocardium Minimally Contributes to Coronary Endothelium in the Embryonic Ventricular Free Walls. Circulation Research. 118 (12), 1880-1893 (2016).
- Kinlay, S., Libby, P., Ganz, P. Endothelial function and coronary artery disease. Current Opinion in Lipidology. 12 (4), 383-389 (2001).
- Jacques, D., Bkaily, G. Endocardial endothelial cell hypertrophy takes place during the development of hereditary cardiomyopathy. Molecular and Cell Biochemistry. 453 (1-2), 157-161 (2019).
- Grossfeld, P., Nie, S., Lin, L., Wang, L., Anderson, R. H. Hypoplastic Left Heart Syndrome: A New Paradigm for an Old Disease. Journal of Cardiovascular Development and Disease. 6 (1), (2019).
- Li, Y., et al. Down-regulated RGS5 by genetic variants impairs endothelial cell function and contributes to coronary artery disease. Cardiovascular Research. , (2019).
- Miao, Y., et al. Single-Cell RNA-Seq Reveals Endocardial Defect in Hypoplastic Left Heart Syndrome. bioRxiv. , (2019).
- Chen, H. I., et al. The sinus venosus contributes to coronary vasculature through VEGFC-stimulated angiogenesis. Development. 141 (23), 4500-4512 (2014).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır