
Method Article
الهندسة الوراثية ديسكويديوم ديكتيوستيليوم "الخلايا استناداً إلى" التحديد والنمو في البكتيريا
In This Article
Summary
ديسكويديوم ديكتيوستيليوم كائن نموذج شعبية لدراسة العمليات الخلوية المعقدة مثل الهجرة الخلية والالتقام، والتنمية. الأداة المساعدة للكائن الحي يتوقف على إمكانية التلاعب بالجينات. نقدم هنا، أساليب ترانسفيكت الخلايا ديسكويديوم ديكتيوستيليوم أن التغلب على القيود القائمة لتثقيف الخلايا في سائل الإعلام.
Abstract
ديسكويديوم ديكتيوستيليوم كائن حي نموذجية مثيرة للاهتمام لدراسة عمليات تمييز الخلية أثناء التنمية والخلية الإشارات والأسئلة الهامة البيولوجيا الخلوية الأخرى. التكنولوجيات المتاحة للتلاعب وراثيا بخلايا ديكتيوستيليوم متطورة جداً. ترانسفيكشنز يمكن أن يؤديها باستخدام علامات التحديد مختلفة وعلامة إعادة ركوب الدراجات، بما في ذلك جزئ مثلى والطفرات insertional. وهذا معتمد من قبل جينوم مشروحة جيدا. بيد أن هذه النهج هي الأمثل لخطوط الخلايا أكسينيك المتنامية في الثقافات السائل ويصعب من تطبيق غير أكسينيك البرية من نوع الخلايا، التي تتغذى على البكتيريا فقط. الطفرات التي تكون موجودة في سلالات أكسينيك تخل برأس الإشارات، مما تسبب في ماكروبينوسيتوسيس المفرطة المطلوبة لتغذية، وتضعف الهجرة الخلية، مما يفند تفسير توصيل الإشارة وتجارب إنزيمية في هذه السلالات. محاولات سابقة للتلاعب وراثيا بخلايا غير أكسينيك تفتقر إلى الكفاءة والإجراءات التجريبية المعقدة المطلوبة. قمنا بتطوير بروتوكول تعداء بسيطة، للمرة الأولى، ويتغلب على هذه القيود. هذه السلسلة من التحسينات الكبيرة في علم الوراثة الجزيئي ديكتيوستيليوم السماح بالتلاعب بها بسهولة كما سلالات مختبرية قياسية البرية من نوع الخلايا. بالإضافة إلى المزايا لدراسة عمليات الإشارات وحركية غير المعطوب، يمكن طفرات التي تعطل النمو القائم على ماكروبينوسيتوسيس الآن سهولة معزولة. وعلاوة على ذلك، سير العمل تعداء كامل هو تسارع إلى حد كبير، مع المؤتلف الخلايا التي يمكن أن تتولد في أيام بدلاً من أسابيع. هناك ميزة أخرى أن علم الوراثة الجزيئي كذلك يمكن أن يؤديها مع معزولة طازجة البرية من نوع ديكتيوستيليوم عينات من البيئة. وهذا يمكن أن تساعد على توسيع نطاق النهج المستخدمة في هذه المجالات البحثية.
Introduction
جنس ديكتيوستيليوم هي أميبا الحية التربة الاجتماعية التي تتغذى أساسا على البكتيريا. يتم وضعها في الأسرة في اللغات متموريات، عدد كبير من الأنواع وقد تم عزل التي يمكن تجميعها في أربعة أطوار مختلفة1. وقد أصبح الأنواع ديسكويديوم ديكتيوستيليوم (ديسكويديوم د) كائن نموذج شعبية لدراسة العمليات الخلوية المعقدة مثل الهجرة الخلية والبلعمه. للتحكم وتوحيد الشروط التجريبية، خلية أكسينيك ووضعت الخطوط التي تستطيع أن تنمو في المتوسطة السائلة المعقدة أو تم تعريفها في حالة عدم وجود البكتيريا2. Ax2، Ax3، تكتسب أهمية خاصة، وعزل سلالات Ax4، التي كانت كلها ولدت في السبعينات وفي نهاية المطاف مستمدة من البرية واحد NC43. ووضعت أدوات للهندسة الوراثية في هذه السلالات أكسينيك، أسفر عن التقديم المنشورة الأولى في عام 19874،5. كذلك وضعت بروتوكولات والأمثل للاستخدام تحت ظروف أكسينيك6،7.
التكيف مع هذه البروتوكولات إلى البرية من نوع ديسكويديوم دال- حاولت السلالات التي لا تستطيع أن تنمو في المرق السائل من عدة مختبرات. ومع ذلك، هذا لا أصبحت ناجحة تماما منذ بروتوكولات تعداء معقدة وتفتقر إلى الكفاءة، وجزئياً بسبب قدرة البكتيريا على العمل كبالوعة لل8،الكواشف انتقائية9. نتيجة لذلك يأتي الأساس الجزيئي جميع البيانات المتعلقة ديسكويديوم د من أحفاد عزل البرية من نوع واحد. لقد أردنا للتغلب على هذا التحديد، ووضع طريقة لتعديل الخلايا ديسكويديوم دال- مستقلة عن قدرتها على النمو في المتوسط السائل وراثيا. ويمكن تفسير الحاجة إلى مثل هذا أسلوب بالملاحظة أنه كان يفترض في الماضي أن الطفرات مما يسمح بزيادة أكسينيك كانت محايدة أساسا ولا تنتقص من فيزيولوجيا الخلية. هذا الافتراض صحيح جزئيا فقط. وبصفة عامة، هناك اثنين من الاختلافات الملحوظة؛ أولاً، بين مختلف عزل سلالات أكسينيك، والثانية، وعند مقارنة هذه السلالات أكسينيك مع يعزل البرية غير أكسينيك8،9.
ولعل أكثر العوامل حسما هو الرئيسية أكسينيك الجينات، فاسب، التي اعتبرت مؤخرا NF1 راسجاب. المهمة الرئيسية ل NF1 راسجاب لكبح جماح رأس النشاط3. حذف الإنزيم في جميع سلالات أكسينيك يؤدي إلى النشاط المفرط رأس يتجلى كتشكيل بقع Ras نشطة كبيرة. هذه البقع Ras الموسع يؤدي إلى تراكم PIP3 في غشاء البلازما. هذه البقع الظهور مصادفة PIP3 ورأس النشطة هي قالب لتشكيل دائرية [روفل] أن يغلق في نهاية المطاف، ويؤدي إلى تشكيل ماكروبينوسوميس10. والنتيجة زيادة مفرطة في النشاط ماكروبينوسيتيك. ماكروبينوسيتوسيس عملية يحركها أكتين. مسابقة لمكونات سيتوسكيليتال للتشكيل أما ماكروبينوسوميس أو بسيودوبودس هو النتيجة. تأثيره على سلوك الخلية ينعكس في منع إنزيمية خلايا النباتية لحمض الفوليك11كاملة تقريبا. هذه البقع PIP3 الموسع على نطاق واسع جداً ثابتة. حتى في الخلايا المتعطشة، تظل البقع PIP3 ويمكن أن يساء تفسيره بسيودوبودس، الذي يمكن أن يسبب مشاكل تفسير الدراسات المتعلقة به إلى المخيم.
وفي بعض الحالات، أن الطفرة NF1 مفيد تجريبيا. وهذا يقودنا إلى حافز ثاني لتطوير أسلوب تعداء للبكتريا نمت ديسكويديوم دال- الخلايا، نظراً للزيادة في معدل ماكروبينوسيتوسيس يجعل الخلايا أكسينيك قيمة للتحقيق في الجوانب الأساسية لهذه العملية12 . ومع ذلك، ألغت الطفرات في الجينات المطلوبة ماكروبينوسيتوسيس، مثل PI3-مؤنزم ورأس10، تقريبا النمو أكسينيك، مما يجعل من الضروري التعامل مع هذه الخلايا من خلال النمو في البكتيريا. وهناك سبب آخر يجعل القائم على البكتيريا ترانسفيكشنز قيمة هو تزايد استخدام ديكتيوستيليدس لاستكشاف المسائل في التطور متعدد سيلولاريتي13،14، الاعتراف بالقربى15،16، ويعزل الإيثار سلوك الخلوية، التي تعتمد أساسا على استخدام البرية الطازجة معزولة من نوع17. جميع المذكورة يمكن تيسير مجالات البحث بأساليب تتسم بالكفاءة للتحوير الوراثي يعزل البرية، وهي غير أكسينيك ولا تنمو في المرق السائل.
لدينا بروتوكولات تسمح للتغلب على أوجه القصور الموصوفة. أخذت معا، إمكانية أداء التلاعبات الجينية مع البكتيريا نمت الخلايا ديسكويديوم دال- يحمل فوائد لجميع الباحثين ديكتيوستيليوم ، حتى لو أنها مجرد زيادة سرعة عملية الاختيار المقرر أن أسرع نمو الاميبا (ح 4 مضاعفة الوقت) على البكتيريا مقارنة بالنمو في وسائل الإعلام أكسينيك (10 ح مضاعفة الوقت).
Protocol
1-إعداد الخلايا والمواد
-
إعداد المخزن المؤقت سورمك
- تحضير 100 مل 100 × سورمك (سورنسن المخزن المؤقت بما في ذلك مجكل2 وكاكل2) المخزن المؤقت بتذويب 20.36 ز خ2ص4 (15 ملم) وز 5.47 Na2هبو4·7 ح2س (2 مم) في 100 مل الماء س2ddH. وآثاره الحل في درجة حرارة الغرفة (RT) وإحضار وحدة التخزين إلى 100 مل مع dH2o.
ملاحظة: المخزن المؤقت الناتج بالرقم الهيدروجيني 6 ولا تحتاج إلى مزيد من ضبط. - إنتاج 1000 مل 1 × يعمل الحل في ddH2سين إضافة 50 ميليلتر كل مجكل2 و كاكل2 لتحقيق تركيزات النهائي لكل 50 ميكرومتر. فلتر تعقيم الحل باستخدام عامل تصفية 0.22 ميكرومتر.
ملاحظة: دائماً إضافة مجكل2 وكاكل2 إلى المخزن المؤقت 1 x لتجنب ترسب الأملاح في 100 × الحل الأسهم. - وبدلاً من ذلك، إعداد2 المخزن المؤقت (2.2 ز خ2ص4 و 0.7 ز ك2هبو4 ل 1 لتر من المخزن المؤقت) وتستكمل مع 50 ميكرومتر مجكل2 وميكرومتر 50 كاكل2 (يشار إليها هنا2MC). استخدم هذا المخزن المؤقت في جميع أنحاء بدلاً من سورمك.
- تحضير 100 مل 100 × سورمك (سورنسن المخزن المؤقت بما في ذلك مجكل2 وكاكل2) المخزن المؤقت بتذويب 20.36 ز خ2ص4 (15 ملم) وز 5.47 Na2هبو4·7 ح2س (2 مم) في 100 مل الماء س2ddH. وآثاره الحل في درجة حرارة الغرفة (RT) وإحضار وحدة التخزين إلى 100 مل مع dH2o.
-
إعداد البكتيريا كمصدر للغذاء دال-ديسكويديوم
- استخدام مستعمرة واحدة من ك. الغازية وتطعيم 1 لتر من رطل-متوسط (مرق ليسوجيني). استخدام قارورة 2 ل. واسمحوا البكتيريا التي تنمو بين عشية وضحاها في 37 درجة مئوية مع الهز 220 لفة في الدقيقة.
ملاحظة: إذا كانت مطلوبة كميات كبيرة من البكتيريا، استخدام الوسائط الغنية مثل 2xTY (خميرة استخراج المتوسطة تريبتوني) أو سوب (سوبر الأمثل مرق) بدلاً من رطل. في حالة استخدام البكتيريا الغازية ك. غير مسموح بها بسبب قيود الأمان، يمكن استخدام BL21 كولاي بدلاً من ذلك. - حصاد الخلايا في اليوم التالي بالغزل منهم إلى أسفل في اثنان أنابيب الطرد المركزي 500 مل في x ~ 6,600 ز للبكتيريا 20 دقيقة أغسل مرة واحدة مع 500 مل من سورمك المخزن المؤقت.
- ريسوسبيند بيليه في 20 مل سورمك. تحقق OD600 (الكثافة البصرية 600 nm) استخدام شحني. تضعف مع نفس المخزن المؤقت ل التطوير التنظيمي600 من حوالي 100.
ملاحظة: البكتيريا الغازية ك يصعب بيليه، ذلك بسرعة عالية نسبيا للغزل أسفل البكتيريا أمر ضروري لتجنب فقدان البكتيريا الغذاء. الحل الأسهم البكتيرية الناتجة يمكن تخزينها لمدة تصل إلى 4 أشهر في الثلاجة عند درجة 4 مئوية والحفاظ على فائدتها كمصدر للغذاء دال-ديسكويديوم. 1 لتر من بين عشية وضحاها تعليق الغازية ك. نمت في المتوسط رطل عادة غلة 20 مل التطوير التنظيمي لحوالي 100600 .
تنبيه: للتأكد من وجود بكتيريا المعدة أحادية الغازية ك.، القيام بكافة الخطوات تحت غطاء محرك السيارة.
- استخدام مستعمرة واحدة من ك. الغازية وتطعيم 1 لتر من رطل-متوسط (مرق ليسوجيني). استخدام قارورة 2 ل. واسمحوا البكتيريا التي تنمو بين عشية وضحاها في 37 درجة مئوية مع الهز 220 لفة في الدقيقة.
-
إعداد H40 انهانسر المخزن المؤقت
- تحضير 100 مل من محلول المخزن المؤقت وحل ز 0.952 من هيبيس في ddH2س المياه وإضافة 100 ميليلتر مجكل2 من حل أسهم م 1. ضبط على درجة الحموضة 7 استخدام كوه للمعايرة. تعقيم المخزن المؤقت باستخدام عامل تصفية 0.22 ميكرومتر أو اﻷوتوكﻻف. استخدام هيبيس خال من الأحماض ولا ملح الصوديوم.
- القيام بإعداد بلازميد يتبع البروتوكول الخاص بالشركة المصنعة، واستخدام مجموعات الملخصة في الجدول للمواد. استخدام البلازميدات إيجاز في الجدول 1.
ملاحظة: نوعية الحمض النووي يستخدم تعداء أمر حاسم. اختيار ترانسفيكتانتس ديسكويديوم دال المتزايد على البكتيريا بالمتطلبات المحددة للمروجين القيادة في الكاسيت الاختيار والتعبير (انظر المناقشة).
| اسم بلازميد | المقاومة/التحديد في البكتيريا | التحديد/المقاومة في ديكتيوستيليوم | العلامة |
| والبلازميدات التعبير اكستراتشروموسومال | |||
| pDM1203 | أمبيسيلين | G418 | لا |
| pDM1207 | أمبيسيلين | G418 | N--المحطة الطرفية بروتينات فلورية خضراء |
| pDM1208 | أمبيسيلين | G418 | N--المحطة الطرفية مشري |
| pPI159 | أمبيسيلين | G418 | منيون الطرفي ن |
| pPI437 | أمبيسيلين | G418 | مسكارليت ن--المحطة الطرفية |
| pPI54 | أمبيسيلين | G418 | MTurquoise2 ن--المحطة الطرفية |
| pDM1209 | أمبيسيلين | G418 | ج-محطة بروتينات فلورية خضراء |
| pDM1210 | أمبيسيلين | G418 | ج-محطة مشري |
| pPI143 | أمبيسيلين | G418 | ج-محطة منيون |
| pPI459 | أمبيسيلين | G418 | ج-محطة مسكارليت |
| pPI142 | أمبيسيلين | G418 | ج-محطة mTurquoise2 |
| والبلازميدات المكوك | |||
| pDM344 | أمبيسيلين | لا | لا |
| pDM1019 | أمبيسيلين | لا | N--المحطة الطرفية بروتينات فلورية خضراء |
| pDM1018 | أمبيسيلين | لا | متشيري ن--المحطة الطرفية |
| pPI152 | أمبيسيلين | لا | منيون الطرفي ن |
| pPI418 | أمبيسيلين | لا | مسكارليت ن--المحطة الطرفية |
| pPI150 | أمبيسيلين | لا | MTurquoise2 ن--المحطة الطرفية |
| pDM1021 | أمبيسيلين | لا | ج-محطة بروتينات فلورية خضراء |
| pDM1020 | أمبيسيلين | لا | ج-محطة مشري |
| pPI153 | أمبيسيلين | لا | ج-محطة منيون |
| pPI457 | أمبيسيلين | لا | ج-محطة مسكارليت |
| pPI151 | أمبيسيلين | لا | ج-محطة mTurquoise2 |
| والبلازميدات التعبير اكستراتشروموسومال إيندوسيبلي | |||
| pDM1038 | أمبيسيلين | هيجروميسين | لا |
| pDM1047 | أمبيسيلين | هيجروميسين | N--المحطة الطرفية بروتينات فلورية خضراء |
| pDM1046 | أمبيسيلين | هيجروميسين | N--المحطة الطرفية مشري |
| pPI450 | أمبيسيلين | هيجروميسين | منيون الطرفي ن |
| pPI452 | أمبيسيلين | هيجروميسين | مسكارليت ن--المحطة الطرفية |
| pPI449 | أمبيسيلين | هيجروميسين | MTurquoise2 ن--المحطة الطرفية |
| pDM1049 | أمبيسيلين | هيجروميسين | ج-محطة بروتينات فلورية خضراء |
| pDM1048 | أمبيسيلين | هيجروميسين | ج-محطة مشري |
| pPI470 | أمبيسيلين | هيجروميسين | ج-محطة منيون |
| pPI460 | أمبيسيلين | هيجروميسين | ج-محطة مسكارليت |
| pPI469 | أمبيسيلين | هيجروميسين | ج-محطة mTurquoise2 |
| استهداف والبلازميدات ملاذ act5 | |||
| pDM1501 | أمبيسيلين | هيجروميسين | لا |
| pDM1513 | أمبيسيلين | هيجروميسين | N--المحطة الطرفية بروتينات فلورية خضراء |
| pDM1514 | أمبيسيلين | هيجروميسين | متشيري ن--المحطة الطرفية |
| pPI231 | أمبيسيلين | هيجروميسين | منيون الطرفي ن |
| pPI419 | أمبيسيلين | هيجروميسين | مسكارليت ن--المحطة الطرفية |
| pPI228 | أمبيسيلين | هيجروميسين | MTurquoise2 ن--المحطة الطرفية |
| pDM1515 | أمبيسيلين | هيجروميسين | ج-محطة بروتينات فلورية خضراء |
| pDM1516 | أمبيسيلين | هيجروميسين | ج-محطة مشري |
| pPI230 | أمبيسيلين | هيجروميسين | ج-محطة منيون |
| pPI458 | أمبيسيلين | هيجروميسين | ج-محطة مسكارليت |
| pPI229 | أمبيسيلين | هيجروميسين | ج-محطة mTurquoise2 |
| والبلازميدات التعبير ريمي | |||
| pDM1220 | أمبيسيلين | هيجروميسين | لا |
| pDM1351 | أمبيسيلين | هيجروميسين | N--المحطة الطرفية بروتينات فلورية خضراء |
| pDM1259 | أمبيسيلين | هيجروميسين | N--المحطة الطرفية مشري |
| pPI465 | أمبيسيلين | هيجروميسين | منيون الطرفي ن |
| pPI468 | أمبيسيلين | هيجروميسين | مسكارليت ن--المحطة الطرفية |
| pPI466 | أمبيسيلين | هيجروميسين | MTurquoise2 ن--المحطة الطرفية |
| pDM1352 | أمبيسيلين | هيجروميسين | ج-محطة بروتينات فلورية خضراء |
| pDM1305 | أمبيسيلين | هيجروميسين | ج-محطة مشري |
| pPI471 | أمبيسيلين | هيجروميسين | ج-محطة منيون |
| pPI467 | أمبيسيلين | هيجروميسين | ج-محطة مسكارليت |
| pPI472 | أمبيسيلين | هيجروميسين | ج-محطة mTurquoise2 |
| المستهدفة في إطار البلازميدات | |||
| pDM1355 | أمبيسيلين | هيجروميسين | ج-محطة بروتينات فلورية خضراء |
| pPI461 | أمبيسيلين | هيجروميسين | ج-محطة مشري |
| pPI462 | أمبيسيلين | هيجروميسين | ج-محطة منيون |
| pPI464 | أمبيسيلين | هيجروميسين | ج-محطة مسكارليت |
| pPI463 | أمبيسيلين | هيجروميسين | ج-محطة mTurquoise2 |
| والبلازميدات المغلوب | |||
| pDM1079 | أمبيسيلين | بلاستيسيدين | لا |
| pDM1080 | أمبيسيلين | نورسيوثريسين | لا |
| pDM1081 | أمبيسيلين | هيجروميسين | لا |
| pDM1082 | أمبيسيلين | G418 | لا |
| والبلازميدات التعبير لجنة المساواة العرقية | |||
| pDM1483 | أمبيسيلين | نورسيوثريسين | لا |
| pDM1489 | أمبيسيلين | هيجروميسين | لا |
| pDM1488 | أمبيسيلين | G418 | لا |
الجدول 1: قائمة بلازميد ترانسفيكشنز غير أكسينيك.
-
إعداد الخلايا ديكتيوستيليوم تعداء
- تنمو ك الغازية لالتقاء في SM المتوسطة (المتوسطة الغنية بالمغذيات) بين عشية وضحاها في "الثقافات الرايت" يمكن تخزينها لمدة تصل إلى أسبوعين في 4 درجات مئوية.
- إضافة حوالي 400 ميليلتر من هذا التعليق البكتيرية على طبق أجار SM (ببتون 10 غرام/لتر؛ والخميرة استخراج 1 غرام/لتر؛ الجلوكوز 10 غرام/لتر؛ خ2بو4 1.9 غرام/لتر؛ ك2هبو4 × 3 ح2س، 1.3 غرام/لتر؛ مجسو4 ز 0.49 اللامائى/لتر؛ أجار 1.7 في المائة)، وتنتشر بشكل متساو. حلقة عقيمة وتلقيح مع الخلايا ديكتيوستيليوم . انتشار الخلايا عند حافة واحدة اللوحة.
- احتضان لوحة عند 22 درجة مئوية لمدة يومين التأكد من مناطق نمو كبير بما فيه الكفاية تعداء.
ملاحظة: لسلالات ديكتيوستيليوم لا تجعل مناطق نمو كبير (مثلاً، Ax3، DH1، أو JH10)، أو للخبرة المجربون، استخدم مسح اللوحات بدلاً من ذلك. لهذا، اتبع الإرشادات في خطوات 1.5.4 إلى 1.5.6. - حلقة عقيمة وتلقيح مع خلايا ديكتيوستيليوم (حوالي 2-4 × 105 الخلايا). نقل الخلايا إلى 800 ميليلتر من الكثيفة الغازية ك تعليق في الخلايا الدمتوسط مزيج من بيبيتينج صعودا وهبوطاً.
- نقل 400 ميليلتر ميليلتر 200، 100 ميليلتر و 50 ميليلتر في لوحات أجار SM الطازجة. إضافة إلى كل لوحة 400 ميليلتر إضافية SM الغازية ك. تعليق، وتنتشر بالتساوي، والجاف.
- احتضان لوحات عند 22 درجة مئوية لمدة 2 أيام حتى تصبح اللوحات شفافة.
ملاحظة: نظراً لمعدل نموها أسرع، تنتج البكتيريا في البداية حديقة المتلاقية والزحاريه في وقت لاحق "واضحة" اللوحة البكتيريا. الوقت اللازم لهذه العملية يمكن أن تختلف تبعاً للخلفية إجهاد وقدرة خلايا متحولة تنمو على البكتيريا.
2-تعداء الخلايا ديكتيوستيليوم استناداً إلى التحديد البكتيرية

الشكل 1 : سير العمل تعداء الخلايا نمت البكتيريا ديكتيوستيليوم . يتم سرد الخطوات تعداء على النحو التالي. تنمو خلايا ديسكويديوم دال على صفيحة SM المصنف مع البكتيريا الغازية ك. (أحمر). حصاد الخلايا فقط من الجبهة التغذية (الأخضر)، تجنب الخلايا التي تقوم فعلا بوضع (أخضر داكن). تغسل الخلايا في H40. ريسوسبيند الخلايا لكثافة نهائي من 2-4 × 107 خلايا/مل. مزيج من تعليق خلية مع 1-2 ميكروغرام للحمض النووي. نقل الخليط إلى انهانسر ومبومو ونبض خلايا. نقل الخلايا مباشرة بعد انهانسر لطبق مع سورمك والبكتيريا. السماح للخلايا لاسترداد ح 5 قبل إضافة علامة اختيار. البلازميدات اكستراتشروموسومال، إضافة التحديد إلى الطبق مباشرة. ترانسفيكتانتس مرئية عادة بعد أيام ~ 2. لخطية بنيات التي تهدف لدمج واحدة في الجينوم، إعداد تخفيف ثلاثة كما هو مبين وإضافة التحديد. البكتيريا هي مأخوذة من OD600 = 100 حل الأسهم. مزج الأنابيب جيدا ونقل الخلايا إلى لوحات زراعة الأنسجة 96-جيدا مسطحة القاع. استخدام لوحات اثنين للتخفيف. "الماصة؛" 150 ميليلتر من تعليق خلية في كل بئر. يستغرق حوالي 5 أيام حتى مستعمرات ضيق مرئية. بئر الأحمر إظهار مثال عن المبلغ المعتاد للخلايا المحولة بنجاح الحصول على (تعديل من المنشور السابق22اللوحة العلوية). الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}
- إعداد لوحات مع تعليق ك الغازية ، أضف 10 مل من المخزن المؤقت سورمك التي تحتوي على البكتيريا الغازية ك لكثافة OD600 = 2 (إضافة 200 ميليلتر OD استعداد600 = 100 ك. الغازية حل الأسهم للمطلوب تركيز البكتيريا) في 10 سم تعامل زراعة الأنسجة طبق بيتري.
ملاحظة: طبق هذا مطلوب في وقت لاحق لزراعتها transfected الخلايا ديسكويديوم د . وبدلاً من ذلك، يمكن استخدام لوحة 6-جيدا-زراعة أنسجة. في حالة تعداء البلازميدات اكستراتشروموسومال، لوحة 6-جيدا-زراعة الأنسجة أكثر كفاءة في استخدام الموارد. استخدام 2 مل من الغازية ك. سورمك (OD600 = 2) تعليق كل بئر. -
إعداد الخلايا ديكتيوستيليوم
- استخدام حلقة تطعيم المتاح 10 ميليلتر، تتخلص الخلايا من مناطق النمو (3 سم تقريبا) من لوحة الثقافة (حافة المنطقة التي تم مسحها) أو إزالة لوحة. نقل الخلايا إلى 1.5 مل أنبوب يحتوي على 1 مل من المثلج H40 المخزن المؤقت.
ملاحظة: توقيت حصاد الخلايا من إزالة لوحات أمر حاسم. حصاد غلة مبكرا قليلاً جداً كمية من الخلايا، أثناء الحصاد للزيادات في أواخر خطر العائد جزئيا تطوير خلايا. - تغسل الخلايا من الغزل إلى أسفل لمدة 2 دقيقة ب 000 1 × ز أو فلاش-الغزل 2 s في 10,000 س ز. تجاهل المادة طافية وريسوسبيند الخلايا الموجودة في المخزن المؤقت H40 بكثافة نهائي من 2-4 × 107 خلايا/مل. تبقى الخلايا الباردة أثناء الإجراء تعداء كله. استخدام ملاط الماء المثلج تأمين الاتصال المباشر من الأنابيب والجليد.
- استخدام حلقة تطعيم المتاح 10 ميليلتر، تتخلص الخلايا من مناطق النمو (3 سم تقريبا) من لوحة الثقافة (حافة المنطقة التي تم مسحها) أو إزالة لوحة. نقل الخلايا إلى 1.5 مل أنبوب يحتوي على 1 مل من المثلج H40 المخزن المؤقت.
-
انهانسر
- إضافة 100 ميليلتر من الخلايا إلى أنبوب مع 1-2 ميكروغرام للحمض النووي. مزيج دقيق من بيبيتينج صعودا وهبوطاً.
- نقل الخليط خلية/الحمض النووي في ومبومو انهانسر مبردة مسبقاً (الفجوة 2 مم).
- نبض الخلايا باستخدام إعدادات مربع الموجه التالية: 350 8 مللي ثانية والبقول 2، الخامس والفاصل الزمني لنبض ق 1.
ملاحظة: لا تقم بإضافة أكثر من 2 ميكروغرام للحمض النووي. مبالغ أعلى سامة للخلايا، وتقليل كفاءة تعداء. وأضاف المجموع حجم الحمض النووي لا ينبغي أن تتجاوز 5 ميليلتر. - نقل الخلايا فورا في وقت سابق استعداد 10 سم على طبق بيتري مع سورمك الغازية ك. وتسمح للخلايا لاسترداد ح 5.
ملاحظة: فحص الخلايا في طبق بتري تحت مجهر مقلوب. الخلايا تظهر جولة مباشرة بعد انهانسر ولكن سيعود إلى شكلها أميبية بعد حوالي 30 دقيقة، عندما كان لها ما يكفي من الوقت لإرفاقها بشكل صحيح على السطح.
-
اختيار ترانسفيكتانتس: اعتماداً على ما إذا كان يستهدف تعداء لإنشاء المغلوب، وطريقة الكبس، أو تدق في act5 ، أو للتعبير عن بروتين مراسل فلورسنت من بلازميد اكستراتشروموسومال، القيام بعملية الاختيار عن طريق أحد فيما يلي وصف الأساليب.
ملاحظة: وخلافا للخلايا المزروعة أكسينيكالي، الخلايا نمت البكتريا شديدة المقاومة بلاستيسيدين. التحديد، ولذلك يتم دائماً الاضطلاع باستخدام G418 أو هيجروميسين (انظر المناقشة).- طرق الرافضة وتدق الإضافية act5 تدق الإضافية
- فصل الخلايا بعناية من طبق بيتري بها مرارا وتكرارا إجبار السائل ماصة على السطح.
- قم بإعداد ثلاثة تخفيف في تعليق سورمك الغازية ك. (OD600 = 2) وإضافة عامل انتقائية وفقا للمقاومة المستخدمة (انظر الشكل 1).
- انخفاض تمييع: مزيج مل 9 من تعليق خلية مع 20.4 مل سورمك و 600 ميليلتر الغازية ك حل الأسهم.
- إضعاف المتوسط: مزيج ميليلتر 900 من تعليق خلية مع مل 28.5 من سورمك و 600 ميليلتر الغازية ك حل الأسهم.
- تمييع عالية: مزيج ميليلتر 90 من تعليق خلية مع 29.3 مل سورمك و 600 ميليلتر الغازية ك حل الأسهم.
- إضافة 30 ميليلتر علامة الاختيار (100 × الحل الأسهم).
- توزيع تخفيف مستعدين في لوحات زراعة الأنسجة 96-جيدا مسطحة القاع ميليلتر 150 بيبيتينج من تعليق خلية في كل بئر.
ملاحظة: ويهدف هذا الإجراء إلى استنساخ واحدة الشاشة بدلاً من السكان. وتحيط التحديد حوالي 5-7 أيام، اعتماداً على بنية استخدامها.
- البلازميدات اكستراتشروموسومال
- إضافة علامة اختيار مباشرة للطبق 10 مل (انظر الشكل 1).
ملاحظة: ليست هناك حاجة لإعداد تخفيف، نظراً لوجود أي رغبة السكان الاستنساخ. أسرع عملية اختيار والبلازميدات اكستراتشروموسومال يرجع إلى أرقام عالية نسخة موجودة في الخلايا ديسكويديوم دال . ويمكن توقع ترانسفيكتانتس بعد 32 ساعة إلى 4 أيام.
تنبيه: أن المضادات الحيوية المستخدمة كعلامة التحديد مواد سامة. ارتداء القفازات.
- إضافة علامة اختيار مباشرة للطبق 10 مل (انظر الشكل 1).
- طرق الرافضة وتدق الإضافية act5 تدق الإضافية
-
شاشة النسخ التي تم الحصول عليها ترانفيكتانتس إيجابية. لمحاولات المغلوب أو طريقة الكبس، اتبع الإرشادات التي تظهر في الخطوة 2.5.1. تحقق من نجاح تعداء البلازميدات اكستراتشروموسومال باستخدام الإرشادات التي تظهر في الخطوة 2.5.2.
- لطرق الرافضة، تدق الإضافية، وتدق act5 الإضافية، أداء الشاشة الأولى عن طريق PCR للتأكد من إدماج بناء محور الجينوم الصحيح.
ملاحظة: إلى أقصى حد احتمال الاستنساخ السكان، استخدام تمييع أعلى يمكن أن ينتج ترانسفيكتانتس بعد التحديد. هدف للوحات التي هي ثلث الحد الأقصى من الآبار التي تحتلها.- لتوسيع نطاق السكان الاستنساخ، نقل الحيوانات المستنسخة التي نمت بعد التحديد من لوحة 96-جيدا-زراعة الأنسجة في لوحة 12-جيدا-زراعة أنسجة تنمو خلايا كافية لعزل الحمض النووي. إمداد كل بئر مع 1 مل من سورمك الغازية ك. (OD600 = 2) وعلامة التحديد الطازجة.
ملاحظة: يوم 1 عادة ما تكون كافية للحصول على روافد مناسبة أيضا لعزل الحمض النووي. - القيام بعزل الحمض النووي الجينومي ميني، حصاد الخلايا من بئر المتلاقية وعزل الحمض النووي باستخدام مجموعة أدوات استخراج الحمض النووي مصغرة اتباع إرشادات الشركة المصنعة.
- استخدام الحمض النووي الجينومي معزولة جنبا إلى جنب مع الإشعال المناسبة و تق-بوليميريز (انظر الجدول للمواد) على الشاشة PCR إينتيجراندس إيجابية.
ملاحظة: بعد تأكيد الاندماج الإيجابي في موضع الجينوم الصحيحة، ينبغي إجراء تحليل لطخة جنوب. وهذا ما يضمن قدرا أكبر من الثقة في عدم وقوع أحداث الإدراج إضافية في مناطق الجينوم غير محدد.
- لتوسيع نطاق السكان الاستنساخ، نقل الحيوانات المستنسخة التي نمت بعد التحديد من لوحة 96-جيدا-زراعة الأنسجة في لوحة 12-جيدا-زراعة أنسجة تنمو خلايا كافية لعزل الحمض النووي. إمداد كل بئر مع 1 مل من سورمك الغازية ك. (OD600 = 2) وعلامة التحديد الطازجة.
- للبروتينات الفلورية مراسل، وتحديد الخلايا إيجابية فلورسنت بصريا وتحقق مع مجهر الأسفار. وبدلاً من ذلك، إجراء وصمة عار غربية مع الأجسام المضادة المناسبة لتحديد الهوية الكيميائية الحيوية.
- لطرق الرافضة، تدق الإضافية، وتدق act5 الإضافية، أداء الشاشة الأولى عن طريق PCR للتأكد من إدماج بناء محور الجينوم الصحيح.
| البرنامج بكر | ||
| الخطوة | درجة الحرارة | الوقت |
| تمسخ الأولى | 94 درجة مئوية | 30 s |
| دورات 30 | 94 درجة مئوية | 15-30 ق |
| 42 درجة مئوية | 15-60 ثانية | |
| 68 درجة مئوية | 1 دقيقة/كيلو بايت | |
| التمديد النهائي | 68 درجة مئوية | 5 دقيقة |
| عقد | 4-10 درجة مئوية | |
| تكوين رد فعل | ||
| المكون | رد 25 ميكروليتر | التركيز النهائي |
| التمهيدي إلى الأمام 10 ميكرومتر | 0.5 ميليلتر | 0.2 ميكرون |
| 10 ميكرون عكس التمهيدي | 0.5 ميليلتر | 0.2 ميكرون |
| قالب الحمض النووي | متغير (ca. 5 ميليلتر) | < 1,000 نانوغرام |
| 2 x ميكس ماجستير مع "معيار المخزن المؤقت" بما في ذلك بوليميراز (انظر الجدول للمواد) | ميليلتر 12.5 | س 1 |
| المياه خالية من نوكلاس | إلى 25 ميليلتر | < 1,000 نانوغرام |
الجدول 2: تكوين البرنامج وعينه PCR للتضخيم من دال-ديسكويديوم الحمض النووي-
النتائج
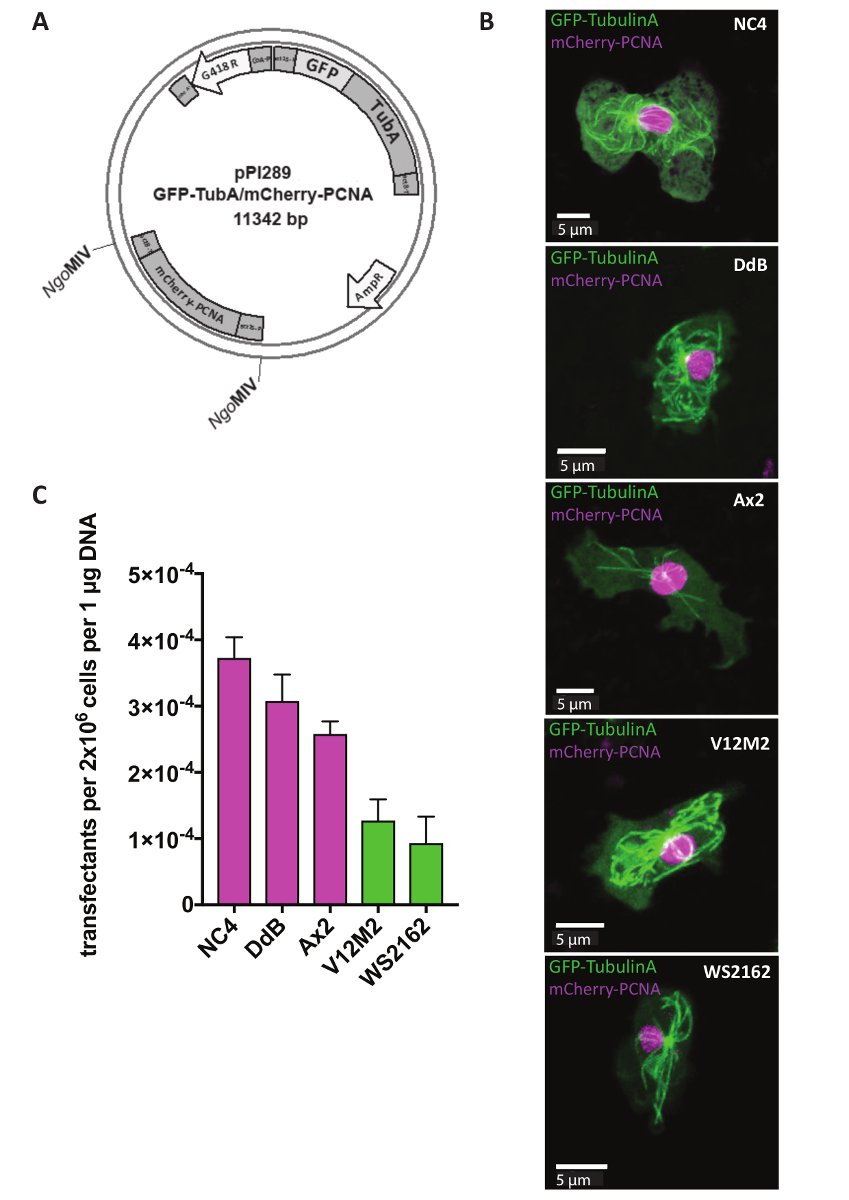
وتستخدم البلازميدات اكستراتشروموسومال مراسل الدراسات، التي تهدف إلى التعرف على توطين بعض البروتينات داخل خلية أو التغييرات في البنية الخلوية لخلايا متحولة. للعديد من النهج، مثل رصد دورة الخلية، من الأهمية بمكان أن نعرب عن اثنين من الصحفيين في نفس الوقت. الآن هذا من الممكن استخدام نظامنا بلازميد اكستراتشروموسومال مراسل المزدوج (الجدول 1). في اليوم الأول، تم transfected الخلايا قبل إضافة علامة اختيار G418 بعد ح 5 (الشكل 1). في المثال، NC4، DdB، Ax2، ويعزل البرية مشتقة بشكل مستقل V12M2 و WS2162 (تكميلية الجدول 1) كانت transfected مع pPI289 بلازميد، الذي يشفر للتجارة والنقل-توبولينا، علامة ل microtubules ومشري-منها، بروتين الذي يستخدم لرصد دورة الخلية (الشكل 2أ). بعد ح 32، لوحظ الخلايا تحت المجهر. أعربت غالبية الخلايا عن كل البروتينات الفلورية المسمى الانصهار، متسقة مع السابقة تقارير ذلك التعبير عن اثنين من الصحفيين من نفس بلازميد يظهر التعبير مستويات مماثلة، ويكاد يكون من المستحيل عند استخدام البلازميدات مختلفة اثنين 18،19. خلية ممثل لكل خط الخلية (NC4، DdB، Ax2، V12M2، و WS2162) معربا عن المراسل المزدوج المطلوب يظهر في الشكل 2ب. ويرد في الشكل2 (ج)الكفاءة تعداء. إظهار خطوط الخلايا المستمدة من NC4 كفاءات تعداء أفضل. ومع ذلك، حصل على لخطوط الخلايا V12M2 و WS2162، إلى حد كبير ارتفاع عدد ترانسفيكتانتس.

الشكل 2 : التعبير عن بلازميد اكستراتشروموسومال. هي transfected البلازميدات اكستراتشروموسومال (A) مباشرة في شكل دائري. على سبيل مثال، يتم إظهار pPI289 مراسل المزدوج. المواقع MIV المنظماتالتي تشير إلى إدراج المراسل الثاني في بلازميد التعبير اكستراتشروموسومال. (ب) Z-الإسقاط خلية الممثل الإعراب عن التجارة والنقل-توبولينا (هيولى) ومنها مشري (النووية إلى حد كبير) لخمسة خطوط مختلفة البرية من نوع الخلية المستخدمة (NC4، DdB، Ax2، V12M2، WS2162). وتم حساب الكفاءة (ج) تعداء للخلية خمسة بنود المصورة في (ب). سيظهر هو المتوسط التجربتين. أشرطة الخطأ تشير إلى ± التنمية المستدامة. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}
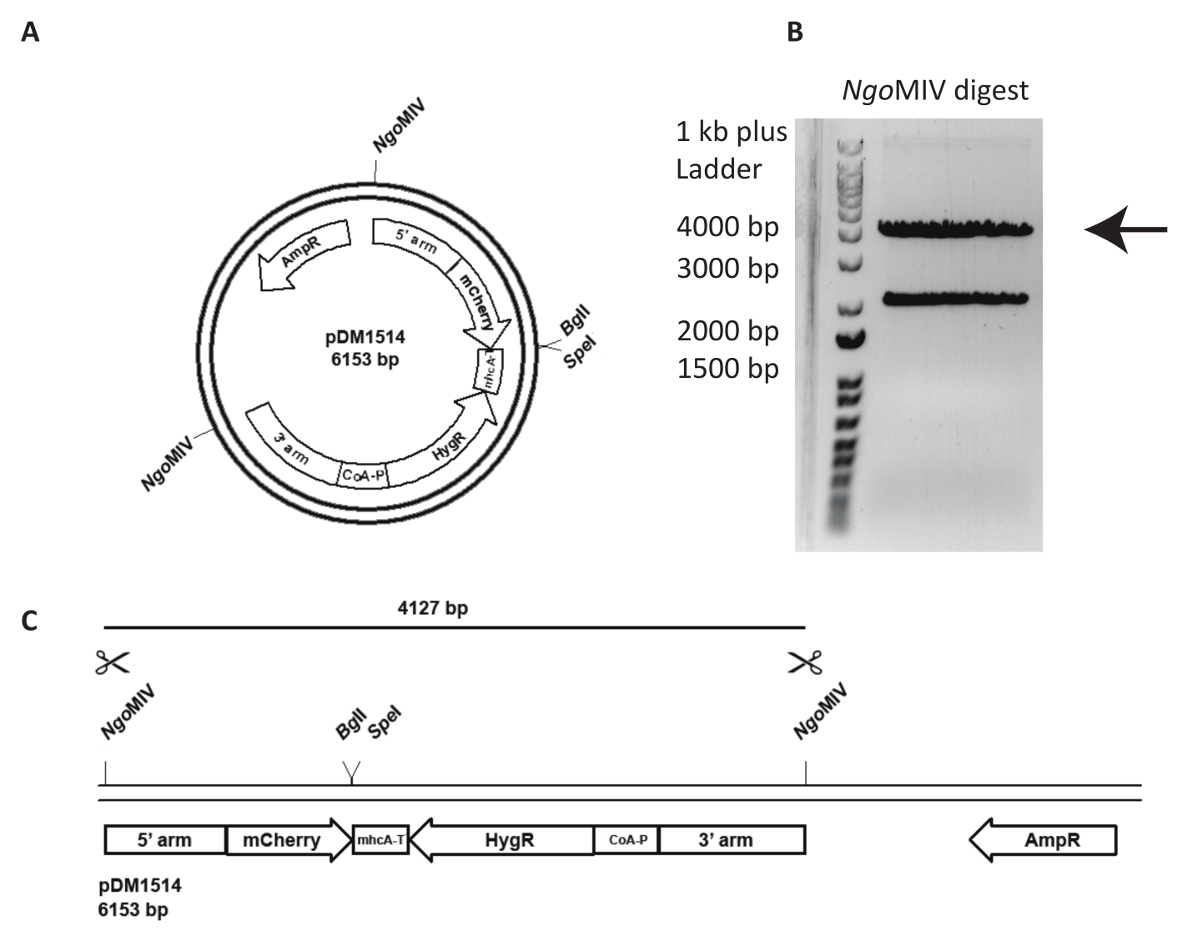
ناقل تستهدف الاندماج مكان محدد الجينوم هو أكثر تحديا ويتطلب تحليلاً أكثر حذراً من خط الخلية التي تم إنشاؤها. في الشكل 3، تتم محاولة act5-متشيري كي في NC4. أولاً، يجب أن خطيا بلازميد زيادة تواتر الأحداث جزئ عقب تعداء. لهذا، يتم قطع pDM1514 بلازميد مع المنظماتMIV. يتم الحصول على شريطين بعد تشغيل الخلاصة على [اغروس] هلام. الفرقة 4127 bp يتضمن بناء المطلوب (الشكل 3). تعداء، هضم الحمض النووي يجب أن تستخلص من الجل وتنقيته باستخدام مجموعة أدوات استخراج جل اتباع إرشادات الشركة المصنعة.

الشكل 3 : إعداد تدق في act5 والحمض النووي تعداء- ويرد مثال على استخدام قانون5 طريقة الكبس pDM1514 بلازميد. يتم سرد الخطوات لإعداد كما يلي. (أ) قبل انهانسر، لينيريزي بلازميد استخدام المواقع MIV المنظماتالمشار إليها. (ب، ج) تشغيل بلازميد قص على [اغروس] هلام حتى يتم فصل شريطين المتوقعة بشكل صحيح. قطع الفرقة bp 4127 تتضمن الأسلحة جزئ، مشري، والمقاومة-الكاسيت، وجل استخراج الحمض النووي. الحمض النووي جاهز الآن تعداء. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}
تم استخدام الحمض النووي المنقي تعداء الخلايا NC4. وبعد 5-6 أيام تحديد، تم الحصول على الحيوانات المستنسخة. تعداء تمثيلية الكفاءات ومقدار إيجابية استنساخ المحددة ل act5 عدة طرق في محاولات يرد في الجدول 3. استنساخ اثنين من تعداء NC4 تم اختيارها عشوائياً وتحليلها من قبل بكر (الشكل 4أ)، وسواء أظهرت أنماط توقع الفرقة المتوقع لضرب في وكذلك التحقق من صحتها مع تحليل لطخة الجنوبية لضمان إدماج صورة واحدة الحدث بناء في الجينوم20. يظهر وصمة عار لتكامل واحدة واضحة في المكان المطلوب act5 (الشكل 4ب). يمكن الآن استخدام خط الخلية مشري NC4::act5 الذي تم إنشاؤه في التجارب.

الشكل 4 : التحقق من act5-مشري كيس في NC4- الخطة (A) والتحكم في تقارير إتمام المشروعات للتحقق التكامل الإيجابي في محور act5 . واستخدمت الإشعال المشار إليه لتحليل اثنين من الحيوانات المستنسخة مستقل والأصل. إظهار كلا استنساخ العصابات المتوقعة للمقاومة-كاسيت والمصب (P1) أو المنبع التمهيدي (P2)، التي لم تكن موجودة في السلالة الأبوية. ويؤكد تركيبة التمهيدي (P1/P2) إدماج كاسيت مشري والمقاومة الصحيحة في محور act5 . يظهر NC4 البرية من نوع الفرقة bp 2800 المتوقعة، بينما كل استنساخ كي تفتقر إلى هذه الفرقة، وبدلاً من ذلك عرض منتج PCR عن 1400 قاعدة أزواج أكبر. (ب) نظام لتقييد استخدام تشفير والجنوب وصمة عار. كلا استنساخ تدق في إظهار الفرقة bp 3400 أصغر حجماً، وهو نتيجة لاندماج بناء محور العمل5، على وجه التحديد إضافية Bclأنا الموقع في كاسيت المقاومة هيجروميسين. أنا المواقع البرية من نوع عنصر التحكم يظهر kb 5.8 المتوقعة الناتجة عن يقع المصب اثنين Bcl. تم اقتصاص وصمة عار وتقسم إلى الوضوح. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}
| بناء المستهدفة في محور act5 | اسم بلازميد | عدد من الآبار المحتلة | عدد النسخ المحددة | استنساخ الإيجابية | صحة الحيوانات المستنسخة (%) | سلالة ديكتيوستيليوم المستخدمة | الكفاءة تعداء (ترانسفيكتانتس/2 × 10 ^ 6/1 ميكروغرام الحمض النووي) |
| ليفيكت--مشري | pPI226 | 7 | 7 | 1 | 14.2 | AX2 | 3.5 × 10 ^-6 |
| ليفياكت--بروتينات فلورية خضراء | pPI227 | 12 | 12 | 5 | 41.6 | AX2 | 6 × 10 ^-6 |
| بروتينات فلورية خضراء | pDM1513 | 3 | 3 | 2 | 66.6 | AX2 | 1.5 × 10 ^-6 |
| متشيري | pDM1514 | 3 | 3 | 1 | 33.3 | AX2 | 1.5 × 10 ^-6 |
| بروتينات فلورية خضراء | pDM1513 | 66 | 9 | 5 | 55.5 | AX2 | 3.3 × 10 ^-5 |
| مشري | pDM1514 | 221 | 12 | 10 | 83.3 | AX2 | 1.1 × 10 ^-4 |
| H2B مشري | pPI420 | 3 | 3 | 1 | 33.3 | AX2 | 1.5 × 10 ^-6 |
| H2B-متشيري | pPI420 | 7 | 7 | 6 | 85.7 | AX2 | 3.5 × 10 ^-6 |
| مشري | pDM1514 | 10 | 10 | 7 | 70 | DdB | 5 × 10 ^-6 |
| مشري | pDM1514 | 240 | 12 | 11 | 91.6 | DdB | 1.2 × 10 ^-4 |
| مشري | pDM1514 | 320 | 12 | 12 | 100 | NC4 | 1.6 × 10 ^-4 |
الجدول 3: تعداء الكفاءة ومقدار الحصول على ترانسفيكتانتس إيجابية لتوليد act5 كيس في الخلفيات سلالة مختلفة 22 .
محور العمل5 يقدم التعبير متجانسة نسبيا ل مراسل المتكاملة21. تسمح الخلايا NC4::act5-مشري ولدت ميكس التجارب التي يتعين القيام بها مع act5 تدق الوظائف الأخرى باستخدام بروتين فلورسنت مختلفة مثل التجارة والنقل. لإبراز الميزة الكبرى لهذا النظام، يتم عرض تجارب الاختلاط مع Ax2::act5-التجارة والنقل . بسبب عدم القدرة على ترانسفيكت غير أكسينيك البرية من نوع الخلايا، لا يمكن إجراء هذا النوع من النهج قبل. تجارب ميكس أداة هامة لتحليل سلوك الخلية، لأنها تسمح مقارنة مباشرة بين خطوط الخلايا المختلفة التي تعاني من ظروف تجريبية مماثلة. NC4 الخلايا تنمو بشكل أسرع في المروج البكتيرية من Ax2 الخلايا (الشكل 5ألف). قد يكون هذا بسبب قدرة أعلى على البلعمه البكتيريا أو تحسين القدرة على التحرك، وتظهر به نحو مصدر لغذاء. استخدام مقايسة إنزيمية فولات أجار الأطفال دون سن الخامسة، تم إجراء مقارنة مباشرة للسكان مشري NC4::act5 و Ax2::act5-التجارة والنقل ، تبين أن NC4::act5-مشري ، أكثر كفاءة بكثير في مجال الاستشعار عن حمض الفوليك. بعد ح 4، تمكنت خلايا NC4::act5-مشري أكثر من الزحف تحت [اغروس] من الخلايا Ax2::act5-التجارة والنقل (الشكل 5ب). تحليل المقاييس الموحدة إنزيمية للخلايا المهاجرة تحت [اغروس] كشفت عن أن الخلايا NC4::act5-مشري كانت أسرع وأظهرت استجابة تشيموتاكتيك أقوى من الخلايا Ax2::act5-التجارة والنقل (الشكل 5ه ج ).

الشكل 5 : استخدام قانون5 كيس لتجارب مزيج المستندة إلى الصور إنزيمية. (A) NC4::act5-مشري والإعراب عن Ax2::act5-التجارة والنقل تم تحليل البروتين الفلورسنت المشار إليها من موضع العمل5 للقدرة أن ينمو في حديقة بكتيرية. بعد 4 أيام، جرى قياس القطر البلاك نشأت من الخلايا ديكتيوستيليوم مطلي الانفرادي. عدم أكسينيك act5:: الخلايا NC4 جعل لويحات أكبر كثيرا من الخلايا Ax2::act5-التجارة والنقل أكسينيك (يعني ± التنمية المستدامة، * * * ف < 0.0001، n = 3، تغيير حجم شريط 5 ملم). (ب) لاستخدام إنزيمية الفولات تحت [اغروس] الاعتداء22 مباشرة مقارنة قدرات chemotactic مشري NC4::act5 و Ax2::act5-التجارة والنقل المستخدمة في (أ)، نمت الزحارية كلا سلالات البكتريا-كانت مختلطة بنسبة 50: 50. سمح للخلايا بالزحف تحت [اغروس]. بعد ح 4، تم تصويرها الخلايا التي تم ترحيل إعداد التدرج الفولات باستخدام مجهر [كنفوكل]. ثم تم تحديد عدد الخلايا NC4::act5-مشري و Ax2::act5-التجارة والنقل . NC4::act5-متشيري خلايا كانت أكثر كفاءة في مجال الاستشعار عن حمض الفوليك. حول أكثر إذ NC4::act5-مشري تم العثور على خلايا مقارنة بالخلايا Ax2::act5-التجارة والنقل (يعني ± التنمية المستدامة، * * * ف < 0.0001، n = 6، مقياس بار 100 ميكرومتر). (ج، د) تم تصويره في الخلايا لمدة 60 دقيقة، وحسبت سرعة ومؤشر تشيموتاكتيك. بعد الاختيار المسبق للخلايا تستجيب معظم تشيموتاكتيكالي (فقط تلك التي هاجرت تحت [اغروس])، Ax2::act5-بروتينات فلورية خضراء الخلايا أظهرت القيم الأدنى للسرعة الخلية وانزيمية. وجرى تحليل الخلايا خمسين كل خط الخلية. (± متوسط SD، * p < 0.01، n = 3). (ه) يتم رسم المسارات من NC4::act5-متشيري وتدق Ax2::act5-جفباكت5-وظائف أكثر من 60 دقيقة، تبين حركة موجهة أكثر نحو مصدر تشيمواتراكتينت الخلايا NC4::act5-متشيري (± متوسط SD، * * * ف < 0.0001، n = 6). الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}
يمكن أيضا استخدام خطوط الخلية تدق في act5 للتدفق الخلوي. كما هو الحال مع النمو في البكتيريا، وهناك اختلافات كبيرة في التنمية بين سلالات أكسينيك والأنواع البرية غير أكسينيك. بعد التنمية والهيئات الاثمار NC4 الخلايا تقريبا مرتين كبيرة كتلك المستمدة من الخلايا Ax2. إذا كان يتم خلط الخلايا، يتم الحصول على هيئة الاثمار متوسطة حجم. واستخدمت لتحليل مساهمة خطوط الخلية على حد سواء أكثر كمياً، التدفق الخلوي. هذه التحاليل تبين بوضوح أن الهيئات الاثمار المتوسطة الحجم نتيجة لمستويات مختلفة من مساهمة من كلا خطوط الخلايا. في حين تتكون خلايا NC4::act5-مشري حوالي 75% الجراثيم المقاسة، أسهم Ax2::act5-التجارة والنقل 25 في المائة فقط، تكشف عن ميزة اللياقة بدنية محتملة لسلالات غير أكسينيك (الشكل 6). منذ رصد التحليل لا السكان خلية ساق، وهناك احتمالان لتفسير عدم التوازن في التنمية بين Ax2 و NC4. أحد الاحتمالات أن الخلايا Ax2 تسهم أساسا السكان خلية ساق، بدلاً من دخول السكان خلية بوغ. وبدلاً من ذلك، قد أدخل المزيد من الخلايا NC4 دورة التنموي، مع Ax2 تأخر نسبيا، ونتيجة لذلك غير قادر على المساهمة في الجمعية العامة لهيئة الاثمار. مواصلة تطوير النهج الأخرى إمكانية ترانسفيكت غير أكسينيك البرية من نوع الخلايا ويبسط إجراءات تجريبية إلى حد كبير.

الشكل 6: قانونكيس 5 يسمح بتحليل لتجارب المزيج باستخدام التدفق الخلوي- (أ) NC4::act5-مشري ، و التجارة والنقل Ax2::act5 وضعت كل على حدة أو في خليط 50/50 على ألواح غير المغذيات أجار. وتشكل الخلايا NC4::act5-متشيري أكبر الهيئات الاثمار من Ax2::act5-التجارة والنقل. بدلاً من ذلك، يظهر هذا المزيج حجم متوسط (مقياس بار 5 مم). الأسفار [كنفوكل] الخفيفة الميكروسكوب يوحي بكمية أعلى من جراثيم NC4::act5-متشيري في رؤساء بوغ المستمدة من يمزج. (ب) التحديد الكمي لهذه الملاحظة، كميات الجراثيم في بوغ المقطوع رؤساء من تجربة خلط في (أ) تم تحليلها بواسطة التدفق الخلوي. حوالي 75% الجراثيم نشأت من الخلايا NC4::act5-مشري ، مع 25 في المائة فقط من الخلايا Ax2::act5-التجارة والنقل . (ج) توزيع الممثل من جراثيم من كلا خطوط الخلايا تظهر في تدفق الخلوي مبعثر. حوالي 0.05% تظهر إيجابية إشارات متشيري والتجارة والنقل، مما يوحي بأنه قد حدثت عمليات الانصهار باراسيكسوال أو أن الجراثيم هي التمسك ببعضها البعض. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
{kind=link}
| خط الخلية | وراثيا من الخلفية | رقم المرجع | ونشرت قبل | نوع |
| AX2 (Ka) | DBS0235521 | بلومفيلد et al., 2008 | نوع البرية | |
| NC4 (S) | بلومفيلد et al., 2008 | نوع البرية | ||
| act5::mCherry 5 استنساخ | ICS | HM1912 | باشكي et al., 2018 | تدق في act5 |
| act5::GFP 2 استنساخ | AX2 | HM1930 | باشكي et al., 2018 | تدق في act5 |
| V12M2 | بلومفيلد et al., 2008 | نوع البرية | ||
| WS2162 | بلومفيلد et al., 2008 | نوع البرية |
التكميلية الجدول 1: سلالات مختلفة من ديكتيوستيليوم درس.
Discussion
وقد تم استخدام الخلايا ديكتيوستيليوم غير أكسينيك، والبرية من نوع محدودة جداً حتى الآن في الأبحاث الجزيئية. الأساليب المتاحة للهندسة الوراثية لهذه السلالات تفتقر إلى الموثوقية والكفاءة23، منع اعتمادها العام. يمكن استخدام المواد التي تم إنشاؤها والبروتوكولات المقدمة هنا لأي سلالة ديسكويديوم دال- مستقلة عن قدرته أن ينمو في المتوسط السائل. تجدر الإشارة إلى أن هذا البروتوكول هو الأمثل لخطوط الخلايا المستمدة من NC4. الكفاءة تعداء للسلالات المعزولة طازجة من البرية تختلف عن NC4، كما أننا لاحظنا قبل وترد هنا ل V12M2 و WS2162،من89. شروط انهانسر خاصة ويبدو أن لها تأثير كبير على الكفاءة تعداء وقد تتطلب مزيدا من التحسين لبعض السلالات. وبصفة عامة، لوحظت على كمية كافية من ترانسفيكتانتس في جميع سلالات اختبارها حتى الآن، تبين أن هذه الأساليب قابلة للتطبيق. عدد ترانسفيكتانتس الإيجابية التي تم الحصول عليها باستخدام سلالات المستمدة من NC4 أعلى بالمقارنة بغيرها من السلالات البرية من نوع غير أكسينيك، ولكن في جميع الحالات، يتم الحصول على العدد الكافي من ترانسفيكتانتس للسماح لمزيد من التجارب. هذا، بالإضافة إلى بساطة لدينا البروتوكول، وهو تحسن كبير مقارنة بمحاولات سابقة8،9.
تغطي جميع الإجراءات الجينية القياسية هذه البروتوكولات غير أكسينيك جديدة وإضافة المزيد من المزايا، حيث يمكن إجراء ترانسفيكشنز سرعة وكفاءة في نفس الوقت. يمكن الحصول على النسخ الإيجابية في أيام بدلاً من أسابيع، منذ نمو في النصفين البكتيريا الوقت شعبة. والبلازميدات المنشأة حديثا أيضا العمل في ظل ظروف أكسينيك، ويمكن بشكل روتيني استخدامها تحت ظروف النمو على حد سواء، وهو آخر النهوض بهذه الأساطير22. كما أدخلت في البروتوكول، قد بلازميد المستخدمة لاختيار على البكتيريا المتطلبات الخاصة التي تعتبر حاسمة بالنسبة لنجاح تعداء. على وجه الخصوص، المروجين القيادة شرائط التعبير والمقاومة هامة لنجاح تعداء. مروج غالباً ما تستخدم act6 (أكتين 6)24 للقيادة أشرطة الكاسيت المقاومة أو التعبير يفتقر إلى الكفاءة عندما تزرع الخلايا على البكتيريا. في نظامنا بلازميد، داعية نشطة للغاية act15 (أكتين 15) محركات جميع الأشرطة التعبير، بينما الأشرطة المقاومة تحت سيطرة أحد المروجين coA (كواكتوسين A)، سواء التي تنشط في ظل الظروف أكسينيك، وفي الخلايا المزروعة في البكتيريا. متطلبات للتعبير عن مراسل بنيات فضلا عن طريقة الكبس وثوابت المغلوب تجعل من الضروري استخدام مرجع بلازميد لدينا، ولكن للأسف يحد استخدام الموجهات التي تستخدم المروج الكفاءة act6 بالفعل بإنشاء.
الكفاءة المروج تكتسي أهمية حاسمة خاصة ترانسفيكشنز التي تعتمد على حدث واحد تكامل في موضع الجينوم الصحيح. سبب لنا تحسين المروجين القيادة تعبير الجينات المقاومة، ينتج ما يكفي من البروتين المقاومة من التكاملات محور واحد. وكان شاغلا رئيسيا تحابي تكاملات متعددة في مجال المجين عند استخدام هيجروميسين أو G418 كما هو موضح. لا تحابي تكاملات متعددة وقد لوحظ حتى الآن، كما ذكرت سابقا لتحديدات G418 باستخدام نظم المروج الأكبر سنا25. وهذا يعني أن كل من هيجروميسين و G418 علامات التحديد مناسبة لتوليد تدق الرافضة نظيفة وتدق ins. للأسف، بلاستيسيدين علامة التحديد لا يعمل تحت ظروف غير أكسينيك. هذا عيب رئيسي لأسلوبنا، منذ كاسيت المقاومة بلاستيسيدين تستخدم بشكل روتيني لإنشاء بنيات المغلوب في ديسكويديوم د. الثوابت التي تم إنشاؤها بالفعل مع أشرطة المقاومة بلاستيسيدين ستحتاج إلى أن يكون ريديريفيد باستخدام واحدة من علامات التحديد قابلة للتطبيق. وثمة إمكانية أخرى للتغلب على هذا القيد الجمع بين أكسينيك أنشئت مؤخرا ديسكويديوم دال كريسبر التكنولوجيا مع هذا البروتوكول تعداء26. جيل مناسبة، واحدة دليل الكشف (سجرناس) أبسط وأسرع من إعادة بناء كاملة بنيات المغلوب. لاتجاهات المستقبل، إمكانية توليد عدة طرق الرافضة استخدام كريسبر/Cas9 جنبا إلى جنب مع هذا البروتوكول تعداء جذابة وقد تمهد الطريق للعديد من الباحثين في المجتمع ديكتيوستيليوم . ومع ذلك، ينبغي أن تدرس قابلية نظام التعبير عابر المنشأة المستخدمة كريسبر/Cas9 في أكسينيك الخلايا المزروعة بعناية.
يوفر act5 تدق في نظام عرض نظام تكامل آمنة وموثوق بها لتوليد خطوط الخلايا مستقرة في ديسكويديوم دال، مع استفادة مواقع مماثلة في سائر الكائنات الحية22. كما يعتمد على حدث واحد تكامل في الجينوم ويوفر العديد من الإمكانيات للاتجاهات البحثية المختلفة. مروج act5 تنشط بقوة وضمانات التعبير متجانسة تقريبا27 مستقلة عن شروط زراعة. البروتينات الفلورية مراسل يمكن دمجها بسهولة في هذا المكان ملاذا والبلازميدات استهداف هندسيا باستخدام. يمكن أن يكون هذا مفيداً، على سبيل المثال، لأغراض تتبع الخلية، كما هو موضح هنا في تجربة مزيج. إظهار الخلايا تقلب أدنى خلية إلى التعبير، التي يمكن أن تساعد في الخلية الآلي تتبع. الأهم من ذلك، يظهر إدخال تسلسل المطلوب في موضع العمل5 محايدة فينوتيبيكالي28،29. حسبما التعبير لا تعتمد على علامة اختيار للحفاظ على التعبير البروتين، كما رأينا في إينتيجرانتس عشوائي أو خطوط الخلايا الحاملة لناقل اكستراتشروموسومال، نظام act5 يمكن أن تكون مفيدة لتجارب الإنقاذ، كذلك. منذ خطوط الخلية متجانسة، يمكن تحليل كافة الخلايا. وهذا لا يمكن استخدام بلازميد اكستراتشروموسومال للتعبير، التي يوجد تباين كبير في التعبير.
وبالإضافة إلى هذه التحسينات العامة لجميع سلالات، يتيح القدرة على استخدام غير أكسينيك البرية من نوع الخلايا للأبحاث الجزيئية تقييما لآثار الطفرات المتراكمة في سلالات المختبر الحالي. تم العثور على عدة ترتيبات جديدة الوراثية منذ اعتماد سلالة Ax2 في عام 1970، كما يتضح من تحليلات ميكرواري المنشورة سابقا26. ولوحظت تعمل الاصطناعية بسبب وجود هذه الطفرات. على سبيل المثال، يظهر متحولة المبلغ هو2 التصاق انخفض في مختبر واحد سلالة الخلفية والالتصاق المتزايد في آخر3. مثل هذه البيانات المتضاربة يمكن حلها الآن بتكرار هذه التجربة في سلالة الجد المشترك (في هذه الحالة، DdB). وقد تبين قوة هذا النهج مؤخرا لل30،جتباسي رأس الصغيرة31.
القدرة على إجراء التجارب الجينية في أي سلالة ديسكويديوم دال- يوسع إمكانيات البحث الاتجاهات الجديدة التي تعتمد على استخدام يعزل البرية، لا سيما تلك التي تنطوي على التطور الاجتماعي والتعرف على الأقرباء و دورة الجنسي22.
Disclosures
الكتاب قد لا يوجد تضارب في الكشف عن.
Acknowledgements
يشكر المؤلفون مختبر كاي للإدخال لهذا الأسلوب و LMB المجهري والتدفق الخلوي مرافق للدعم العلمي والتقني الممتاز. تم تمويل هذا العمل من "مجلس البحوث الطبية في" منحة MC_U105115237 ر. ر. كاي، بسرك (مجلس بحوث العلوم البيولوجية والتكنولوجيا الأحيائية) منحة BB/K009699/1 إلى ر. ر. كاي، منح "أبحاث السرطان" في بريطانيا A15672 R. H. Insall، "ويلكوم ترست كبار زمالة" 202867/Z/16/Z لجوناثان R تشب، ولجنة نهر الميكونج التمويل (MC_U12266B) إلى "وحدة جامعة لمكب لجنة نهر الميكونج" في دوري إبطال أوروبا.
Materials
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Eppendorf Microcentrifuges 5424/5424R | ThermoScientific | 05-400-002 | |
| Eppendorf 5702R Centrifuges with A-4-38 Model Rotor | ThermoScientific | 12823252 | |
| Eppendorf Mastercycler Nexus Thermal Cyclers | Sigma Aldrich | EP6331000025 | |
| Gene Pulser X Cell, including CE & PC modules | BioRad | 1652660 | |
| Safety Cabinet Foruna - SCANLAF | Labogene | ||
| BioPhotometer Plus | Eppendorf | ||
| CLSM 710 | Zeiss | ||
| Media & Agaroses | |||
| LoFlo Medium | Formedium | LF0501 | |
| Seakem GTG Agarose | Lonza | 50071 | |
| Low EEO Agarose | BioGene | 300-200 | |
| Purified Agar | Oxoid | LP0028 | |
| SM broth | Formedium | SMB0102 | |
| 2x LB broth | Formedium | LBD0102 | |
| Chemicals | |||
| SYBR Safe DNA Gel Stain | ThermoScientific | S33102 | |
| 1 Kb Plus DNA Ladder | ThermoScientific | 10787018 | |
| 1 Kb DNA Ladder | NEB | N3232L | |
| HEPES free acid | Sigma Aldrich/Merck | 391340 EMD | |
| KH2PO4 | VWR | P/4800/53 | |
| Na2HPO4 * 2 H2O | VWR | 10028-24-7 | |
| MgCl2 * 6 H2O | Sigma Aldrich/Merck | 5982 EMD | |
| CaCl2 * 2 H2O | Sigma Aldrich | 442909 | |
| Folic Acid | Sigma Aldrich | F7876 | |
| KOH | VWR | 26668.263 | |
| Cultur Dishes | |||
| 96 Well Cell Culture Cluster, Flat Bottom with Low Evaporation Lid, Tissue Culture Treated | Corning Incorporated | 3595 | |
| 6 Well Cell Culture Cluster, Flat Bottom with Lid, Tissue Culture Treated | Corning Incorporated | 3516 | |
| 100 x 20 mm style Tissue Culture Dish, Tissue Culture Treated | Corning Incorporated | 353003 | |
| 50 mm Glass Bottom Microwell Dishes No1.5 | MatTek Corporation | P50G-1.5-30-F | |
| Antibiotics | |||
| Geneticin G418-sulphate | Gibco by Life Technologies | 11811-023 | |
| Hygromycin B Gold | InvivoGen | ant-hg-1 | |
| Additional Consumables | |||
| 2 mm gap electroporation cuvettes, long electrode | Geneflow Limited | E6-0062 | |
| 10 µL Inoculating Loop, Blue 10 Micro L | ThermoScientific | 129399 | |
| Spreader, L-shaped, sterile | greiner bio-one | 730190 | |
| Combitips advanced 5 mL | Eppendorf BIOPUR | 30089669 | |
| Kits | |||
| ZR Plasmid Miniprep Classic | Zymoresearch | D4016 | |
| Quick DNA Miniprep Kit | Zymoresearch | D3025 | |
| Zymoclean DNA Recovery Kit | Zymoresearch | D40002 | |
| Enzymes | |||
| Restriction enzymes | NEB | ||
| OneTaq 2X Master Mix with Standard Buffer | NEB | M0482L | |
| T4 DNA Ligase | NEB | M0202S | |
| PrimeSTAR Max DNA Polymerase | TakaraBio | R045A |
References
- Schaap, P. Evolutionary crossroads in developmental biology: Dictyostelium discoideum. Development. 138 (3), 387-396 (2011).
- Sussman, R., Sussman, M. Cultivation of Dictyostelium discoideum in axenic culture. Biochemical and Biophysical Research Communications. 29, 53-55 (1967).
- Bloomfield, G., Tanaka, Y., Skelton, J., Ivens, A., Kay, R. R. Widespread duplications in the genomes of laboratory stocks of Dictyostelium discoideum. Genome Biology. 9 (4), 75 (2008).
- Witke, W., Nellen, W., Noegel, A. Homologous recombination in the Dictyostelium alpha-actinin gene leads to an altered mRNA and lack of the protein. The EMBO Journal. 6, 4143-4148 (1987).
- De Lozanne, A., Spudich, J. A. Disruption of the Dictyostelium myosin heavy chain gene by homologous recombination. Science. 236, 1086-1091 (1987).
- Howard, P. K., Ahern, K. G., Firtel, R. A. Establishment of a transient expression system for Dictyostelium discoideum. Nucleic Acids Research. 16, 2613-2623 (1988).
- Knecht, D., Pang, K. M. Electroporation of Dictyostelium discoideum. Methods in Molecular Biology. 47, 321-330 (1995).
- Wetterauer, B., et al. Wild-type strains of Dictyostelium discoideum can be transformed using a novel selection cassette driven by the promoter of the ribosomal V18 gene. Plasmid. 36, 169-181 (1996).
- Lloyd, M. M., Ceccarelli, A., Williams, J. G. Establishment of conditions for the transformation of nonaxenic Dictyostelium strains. Developmental Genetics. 11, 391-395 (1990).
- Bloomfield, G., et al. Neurofibromin controls macropinocytosis and phagocytosis in Dictyostelium. eLife. 4, (2015).
- Veltman, D. M., et al. A plasma membrane template for macropinocytic cups. eLife. 5, (2016).
- Veltman, D. M., Lemieux, M. G., Knecht, D. A., Insall, R. H. PIP(3)-dependent macropinocytosis is incompatible with chemotaxis. The Journal of Cell Biology. 204 (4), 497-505 (2014).
- Hoeller, O., Kay, R. R. Chemotaxis in the absence of PIP3 gradients. Current Biology. 17, 813-817 (2007).
- Chubb, J. R., Wilkins, A., Thomas, G. M., Insall, R. H. The Dictyostelium RasS protein is required for macropinocytosis, phagocytosis and the control of cell movement. Journal of Cell Science. 113, 709-719 (2000).
- Schaap, P., et al. Molecular phylogeny and evolution of morphology in the social amoebas. Science. 314, 661-663 (2006).
- Du, Q., Kawabe, Y., Schilde, C., Chen, Z. H., Schaap, P. The Evolution of Aggregative Multicellularity and Cell-Cell Communication in the Dictyostelia. Journal of Molecular Biology. 427 (23), 3722-3733 (2015).
- Hirose, S., Benabentos, R., Ho, H. I., Kuspa, A., Shaulsky, G. Self-recognition in social amoebae is mediated by allelic pairs of tiger genes. Science. 333 (6041), 467-470 (2011).
- Strassmann, J. E., Queller, D. C. Evolution of cooperation and control of cheating in a social microbe. Proceedings of the National Academy of Sciences of the United States of America. 108 (2), 10855-10862 (2011).
- Wolf, J. B., et al. Fitness Trade-offs Result in the Illusion of Social Success. Current Biology. 25 (8), 1086-1090 (2015).
- Veltman, D. M., Akar, G., Bosgraaf, L., Van Haastert, P. J. A new set of small, extrachromosomal expression vectors for Dictyostelium discoideum. Plasmid. 61 (2), 110-118 (2009).
- Southern, E. M. Detection of specific sequences among DNA fragments separated by gel electrophoresis. Journal of Molecular Biology. 98 (3), 503-517 (1975).
- Paschke, P., et al. Rapid and efficient genetic engineering of both wild type and axenic strains of Dictyostelium discoideum. PLoS One. 13 (5), 0196809 (2018).
- Woznica, D., Knecht, D. A. Under-agarose chemotaxis of Dictyostelium discoideum. Methods in Molecular Biology. 346, 311-325 (2006).
- Dubin, M., Nellen, W. A versatile set of tagged expression vectors to monitor protein localisation and function in Dictyostelium. Gene. 465 (1-2), 1-8 (2010).
- de Hostos, E. L., et al. Dictyostelium mutants lacking the cytoskeletal protein coronin are defective in cytokinesis and cell motility. Journal of Cell Biology. 120, 163-173 (1993).
- Sekine, R., Kawata, T., Muramoto, T. CRISPR/Cas9 mediated targeting of multiple genes in Dictyostelium. Scientific Reports. 8 (1), 8471 (2018).
- Sadelain, M., Papapetrou, E. P., Bushman, F. D. Safe harbours for the integration of new DNA in the human genome. Nature Reviews Cancer. 12 (1), 51-58 (2011).
- Rosengarten, R. D., et al. Leaps and lulls in the developmental transcriptome of Dictyostelium discoideum. BMC Genomics. 16, 294 (2015).
- Tunnacliffe, E., Corrigan, A. M., Chubb, J. R. Promoter-mediated diversification of transcriptional bursting dynamics following gene duplication. Proc Natl Acad Sci USA. 115 (33), 8364-8369 (2018).
- Gebbie, L., et al. Phg2, a kinase involved in adhesion and focal site modeling in Dictyostelium. Molecular Biology of the Cell. 15, 3915-3925 (2004).
- Jeon, T. J., Lee, D. J., Merlot, S., Weeks, G., Firtel, R. A. Rap1 controls cell adhesion and cell motility through the regulation of myosin II. Journal of Cell Biology. 176, 1021-1033 (2007).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved